Effects of High-Fat Diet Induced Obesity and Fructooligosaccharide Supplementation on Cardiac Protein Expression



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sample Collection
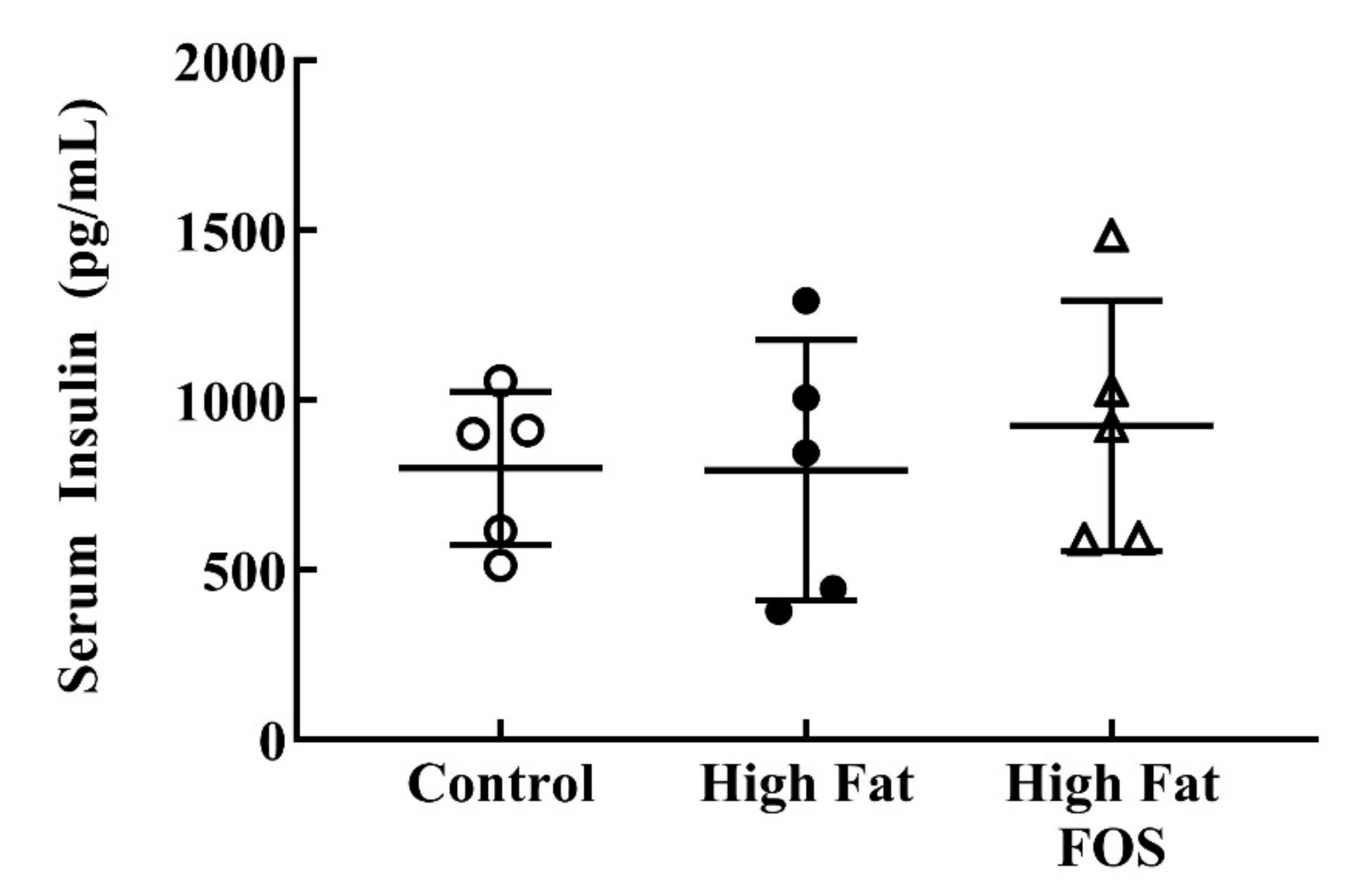
2.2. Serum Insulin and Glucose Measurements
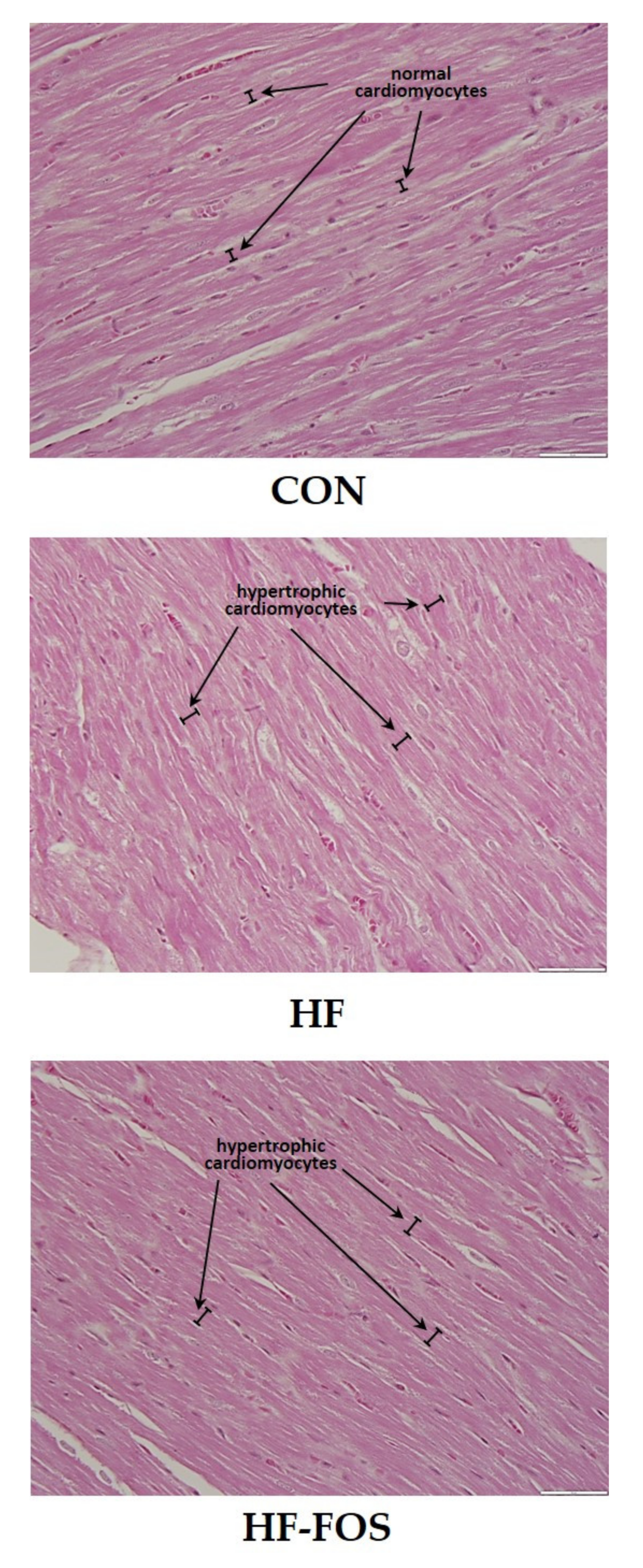
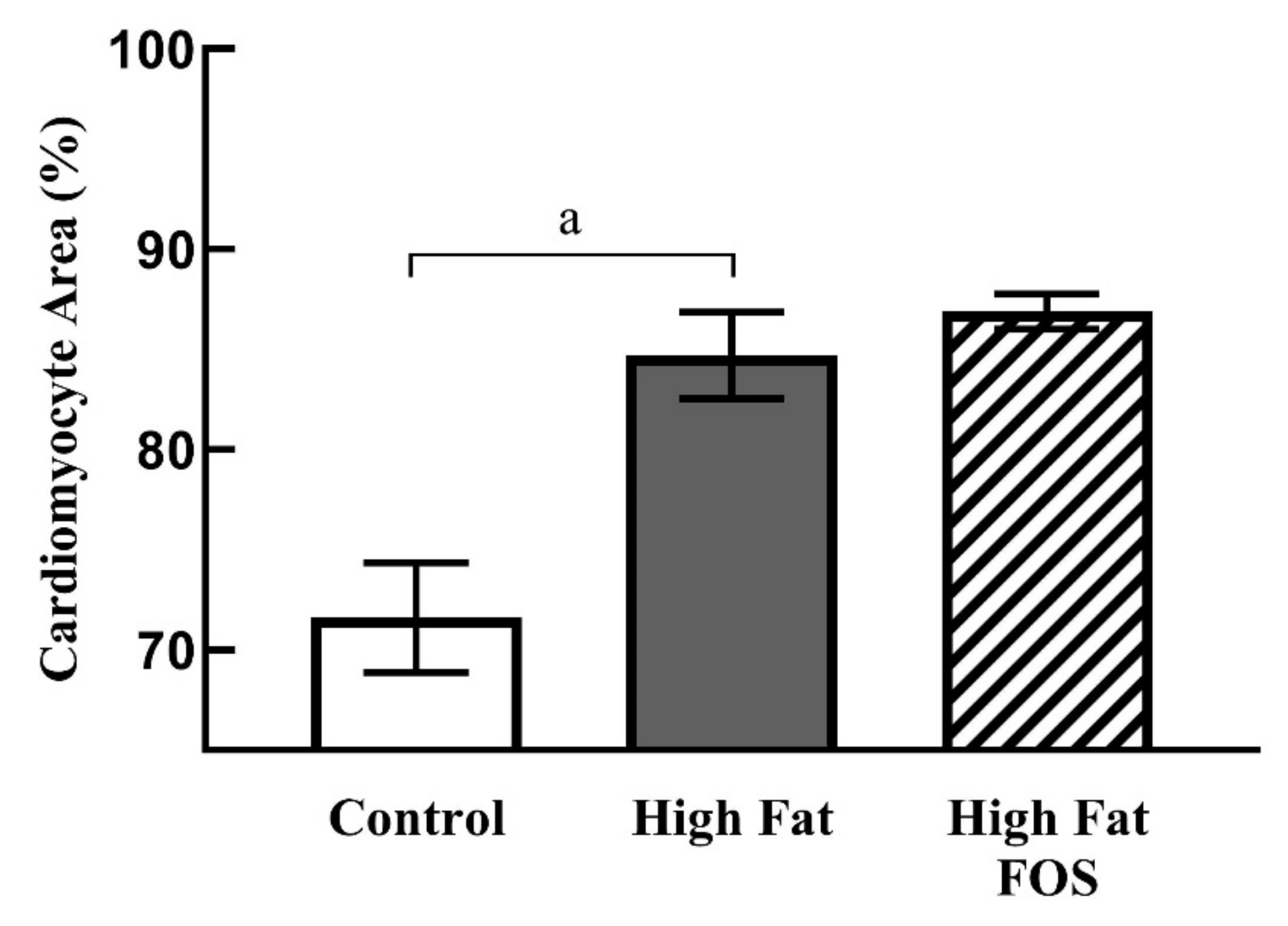
2.3. Histology
2.4. Mass Spectrometry
2.5. Protein Identification and Quantitation
2.6. Statistical Analyses
2.7. In-Silico Functional Analyses
3. Results and Discussions
3.1. Obesity
3.2. Glucose and Insulin
3.3. Histology
3.4. Cardiac Tissue Protein Expression
3.4.1. Lipid Metabolism (HF vs. CON Diet): Supplemental Table S2 and Table 2
3.4.2. Protein Metabolism (HF vs. CON Diet): Supplemental Table S2 and Table 2
3.4.3. Oxidative Stress (HF vs. CON Diet): Supplemental Table S2 and Table 2
3.4.4. Cellular Growth and Proliferation (HF vs. CON Diet): Supplemental Table S2 and Table 2
3.4.5. Lipid Metabolism (HF-FOS vs. HF Diet): Table 3 and Table 4
3.4.6. Protein Folding (HF-FOS vs. HF Diet): Table 3 and Table 4
3.4.7. Adhesion and Cytoskeletal Regulation (HF-FOS vs. HF Diet): Table 3 and Table 4
3.4.8. Cardiac Muscle Regulation (HF-FOS vs. HF Diet): Table 3 and Table 4
3.4.9. Carbohydrate Metabolism (HF-FOS vs. HF Diet): Table 3 and Table 4
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Benjamin, E.J.; Muntner, P.; Alonso, A.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Das, S.R.; et al. Heart Disease and Stroke Statistics-2019 Update: A Report From the American Heart Association. Circulation 2019, 139, e56–e528. [Google Scholar] [CrossRef] [PubMed]
- Peden, J.F.; Farrall, M. Thirty-five common variants for coronary artery disease: The fruits of much collaborative labour. Hum. Mol. Genet. 2011, 20, R198–R205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piche, M.E.; Poirier, P.; Lemieux, I.; Despres, J.P. Overview of Epidemiology and Contribution of Obesity and Body Fat Distribution to Cardiovascular Disease: An Update. Prog. Cardiovasc. Dis. 2018, 61, 103–113. [Google Scholar] [CrossRef]
- Travers, J.G.; Kamal, F.A.; Robbins, J.; Yutzey, K.E.; Blaxall, B.C. Cardiac Fibrosis: The Fibroblast Awakens. Circ. Res. 2016, 118, 1021–1040. [Google Scholar] [CrossRef] [Green Version]
- Zhai, A.B.; Haddad, H. The impact of obesity on heart failure. Curr. Opin. Cardiol. 2017, 32, 196–202. [Google Scholar] [CrossRef]
- Dixon, J.B. The effect of obesity on health outcomes. Mol. Cell Endocrinol. 2010, 316, 104–108. [Google Scholar] [CrossRef]
- Akiyama, T.; Tachibana, I.; Shirohara, H.; Watanabe, N.; Otsuki, M. High-fat hypercaloric diet induces obesity, glucose intolerance and hyperlipidemia in normal adult male Wistar rat. Diabetes Res. Clin. Pract. 1996, 31, 27–35. [Google Scholar] [CrossRef]
- Martins, F.; Campos, D.H.; Pagan, L.U.; Martinez, P.F.; Okoshi, K.; Okoshi, M.P.; Padovani, C.R.; Souza, A.S.; Cicogna, A.C.; Oliveira, S.A., Jr. High-fat Diet Promotes Cardiac Remodeling in an Experimental Model of Obesity. Arq. Bras. Cardiol. 2015, 105, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Abel, E.D.; O’Shea, K.M.; Ramasamy, R. Insulin resistance: Metabolic mechanisms and consequences in the heart. Arter. Thromb. Vasc. Biol. 2012, 32, 2068–2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva Figueiredo, P.; Carla Inada, A.; Marcelino, G.; Maiara Lopes Cardozo, C.; De Cássia Freitas, K.; De Cássia Avellaneda Guimarães, R.; Pereira de Castro, A.; Aragão do Nascimento, V.; Aiko Hiane, P. Fatty Acids Consumption: The Role Metabolic Aspects Involved in Obesity and Its Associated Disorders. Nutrients 2017, 9, 1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.V.; Li, D.L.; Hill, J.A. Heart failure and loss of metabolic control. J. Cardiovasc. Pharmacol. 2014, 63, 302–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz-Topete, D.; List, E.O.; Okada, S.; Kelder, B.; Kopchick, J.J. Proteomic changes in the heart of diet-induced pre-diabetic mice. J. Proteom. 2011, 74, 716–727. [Google Scholar] [CrossRef] [Green Version]
- Miele, L.; Giorgio, V.; Alberelli, M.A.; De Candia, E.; Gasbarrini, A.; Grieco, A. Impact of Gut Microbiota on Obesity, Diabetes, and Cardiovascular Disease Risk. Curr. Cardiol. Rep. 2015, 17, 120. [Google Scholar] [CrossRef]
- David, L.A.; Materna, A.C.; Friedman, J.; Campos-Baptista, M.I.; Blackburn, M.C.; Perrotta, A.; Erdman, S.E.; Alm, E.J. Host lifestyle affects human microbiota on daily timescales. Genome Biol. 2014, 15, R89. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.A.; Keogh, J.B.; Clifton, P.M. Probiotics, prebiotics, synbiotics and insulin sensitivity. Nutr. Res. Rev. 2018, 31, 35–51. [Google Scholar] [CrossRef]
- Le Bourgot, C.; Apper, E.; Blat, S.; Respondek, F. Fructo-oligosaccharides and glucose homeostasis: A systematic review and meta-analysis in animal models. Nutr. Metab. 2018, 15, 9. [Google Scholar] [CrossRef] [Green Version]
- John, G.K.; Wang, L.; Nanavati, J.; Twose, C.; Singh, R.; Mullin, G. Dietary Alteration of the Gut Microbiome and Its Impact on Weight and Fat Mass: A Systematic Review and Meta-Analysis. Genes 2018, 9, 167. [Google Scholar] [CrossRef] [Green Version]
- Choque Delgado, G.T.; Tamashiro, W. Role of prebiotics in regulation of microbiota and prevention of obesity. Food Res. Int. 2018, 113, 183–188. [Google Scholar] [CrossRef]
- Clarke, S.T.; Green-Johnson, J.M.; Brooks, S.P.; Ramdath, D.D.; Bercik, P.; Avila, C.; Inglis, G.D.; Green, J.; Yanke, L.J.; Selinger, L.B.; et al. beta2-1 Fructan supplementation alters host immune responses in a manner consistent with increased exposure to microbial components: Results from a double-blinded, randomised, cross-over study in healthy adults. Br. J. Nutr. 2016, 115, 1748–1759. [Google Scholar] [CrossRef] [Green Version]
- Ayoub, H.M.; McDonald, M.R.; Sullivan, J.A.; Tsao, R.; Meckling, K.A. Proteomic Profiles of Adipose and Liver Tissues from an Animal Model of Metabolic Syndrome Fed Purple Vegetables. Nutrients 2018, 10, 456. [Google Scholar] [CrossRef] [Green Version]
- Nesvizhskii, A.I.; Keller, A.; Kolker, E.; Aebersold, R. A statistical model for identifying proteins by tandem mass spectrometry. Anal. Chem. 2003, 75, 4646–4658. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.; Nesvizhskii, A.I.; Kolker, E.; Aebersold, R. Empirical statistical model to estimate the accuracy of peptide identifications made by MS/MS and database search. Anal. Chem. 2002, 74, 5383–5392. [Google Scholar] [CrossRef]
- Oberg, A.L.; Mahoney, D.W.; Eckel-Passow, J.E.; Malone, C.J.; Wolfinger, R.D.; Hill, E.G.; Cooper, L.T.; Onuma, O.K.; Spiro, C.; Therneau, T.M.; et al. Statistical analysis of relative labeled mass spectrometry data from complex samples using ANOVA. J. Proteome Res. 2008, 7, 225–233. [Google Scholar] [CrossRef]
- Da Huang, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, P.E.; Salapatek, A.M.; Wheeler, M.B. Glucagon-like peptide-1 receptor activation antagonizes voltage-dependent repolarizing K(+) currents in beta-cells: A possible glucose-dependent insulinotropic mechanism. Diabetes 2002, 51 (Suppl. 3), S443–S447. [Google Scholar] [CrossRef] [Green Version]
- Brady, T.M. The Role of Obesity in the Development of Left Ventricular Hypertrophy Among Children and Adolescents. Curr. Hypertens. Rep. 2016, 18, 3. [Google Scholar] [CrossRef] [Green Version]
- Calligaris, S.D.; Lecanda, M.; Solis, F.; Ezquer, M.; Gutierrez, J.; Brandan, E.; Leiva, A.; Sobrevia, L.; Conget, P. Mice long-term high-fat diet feeding recapitulates human cardiovascular alterations: An animal model to study the early phases of diabetic cardiomyopathy. PLoS ONE 2013, 8, e60931. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, S.A., Jr.; Martinez, P.F.; Guizoni, D.M.; Campos, D.H.; Fernandes, T.; Oliveira, E.M.; Okoshi, M.P.; Okoshi, K.; Padovani, C.R.; Cicogna, A.C. AT1 receptor blockade attenuates insulin resistance and myocardial remodeling in rats with diet-induced obesity. PLoS ONE 2014, 9, e86447. [Google Scholar] [CrossRef]
- Kronenberg, F.; Stuhlinger, M.; Trenkwalder, E.; Geethanjali, F.S.; Pachinger, O.; von Eckardstein, A.; Dieplinger, H. Low apolipoprotein A-IV plasma concentrations in men with coronary artery disease. J. Am. Coll. Cardiol. 2000, 36, 751–757. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Kohan, A.B.; Lo, C.M.; Liu, M.; Howles, P.; Tso, P. Apolipoprotein A-IV: A protein intimately involved in metabolism. J. Lipid Res. 2015, 56, 1403–1418. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Lu, Q.; Zhang, Y.; Tian, G. ApoB/apoA1 is an effective predictor of coronary heart disease risk in overweight and obesity. J. Biomed. Res. 2011, 25, 266–273. [Google Scholar] [CrossRef] [Green Version]
- Black, P.N.; DiRusso, C.C. Transmembrane movement of exogenous long-chain fatty acids: Proteins, enzymes, and vectorial esterification. Microbiol. Mol. Biol. Rev. 2003, 67, 454–472. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.D.; Wang, H.; Zhang, J.F.; Li, J.N.; Can, Y.; Qing, L.; Kung, H.F.; Zhang, Y.O. Enhanced peroxisomal beta-oxidation metabolism in visceral adipose tissues of high-fat diet-fed obesity-resistant C57BL/6 mice. Exp. Ther. Med. 2011, 2, 309–315. [Google Scholar] [CrossRef]
- Zhang, L.; Ussher, J.R.; Oka, T.; Cadete, V.J.; Wagg, C.; Lopaschuk, G.D. Cardiac diacylglycerol accumulation in high fat-fed mice is associated with impaired insulin-stimulated glucose oxidation. Cardiovasc. Res. 2011, 89, 148–156. [Google Scholar] [CrossRef] [Green Version]
- Pellieux, C.; Montessuit, C.; Papageorgiou, I.; Pedrazzini, T.; Lerch, R. Differential effects of high-fat diet on myocardial lipid metabolism in failing and nonfailing hearts with angiotensin II-mediated cardiac remodeling in mice. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H1795–H1805. [Google Scholar] [CrossRef] [Green Version]
- Drosatos, K.; Schulze, P.C. Cardiac lipotoxicity: Molecular pathways and therapeutic implications. Curr. Heart Fail. Rep. 2013, 10, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Lekli, I.; Das, S.; Das, S.; Mukherjee, S.; Bak, I.; Juhasz, B.; Bagchi, D.; Trimurtulu, G.; Krishnaraju, A.V.; Sengupta, K.; et al. Coenzyme Q9 provides cardioprotection after converting into coenzyme Q10. J. Agric. Food Chem. 2008, 56, 5331–5337. [Google Scholar] [CrossRef]
- Cao, F.; Zervou, S.; Lygate, C.A. The creatine kinase system as a therapeutic target for myocardial ischaemia-reperfusion injury. Biochem. Soc. Trans. 2018, 46, 1119–1127. [Google Scholar] [CrossRef] [Green Version]
- Zervou, S.; Yin, X.; Nabeebaccus, A.A.; O’Brien, B.A.; Cross, R.L.; McAndrew, D.J.; Atkinson, R.A.; Eykyn, T.R.; Mayr, M.; Neubauer, S.; et al. Proteomic and metabolomic changes driven by elevating myocardial creatine suggest novel metabolic feedback mechanisms. Amino Acids 2016, 48, 1969–1981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Canon, J.M.; Baetscher, M.W.; Finegold, M.; Burlingame, T.; Gibson, K.M.; Grompe, M. Maleylacetoacetate isomerase (MAAI/GSTZ)-deficient mice reveal a glutathione-dependent nonenzymatic bypass in tyrosine catabolism. Mol. Cell Biol. 2002, 22, 4943–4951. [Google Scholar] [CrossRef] [Green Version]
- Rindler, P.M.; Plafker, S.M.; Szweda, L.I.; Kinter, M. High dietary fat selectively increases catalase expression within cardiac mitochondria. J. Biol. Chem. 2013, 288, 1979–1990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohr, M.J.; Evangelista, A.M.; Ferlito, M.; Steenbergen, C.; Murphy, E. S-nitrosylation of TRIM72 at cysteine 144 is critical for protection against oxidation-induced protein degradation and cell death. J. Mol. Cell Cardiol. 2014, 69, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Carlson, E.C.; Ye, G.; Xu, M.; Metreveli, N.S.; Epstein, P.N.; Donthi, R.V.; Xia, S. Catalase protects cardiomyocyte function in models of type 1 and type 2 diabetes. Diabetes 2004, 53, 1336–1343. [Google Scholar]
- Kohlhaas, M.; Nickel, A.G.; Maack, C. Mitochondrial energetics and calcium coupling in the heart. J. Physiol. 2017, 595, 3753–3763. [Google Scholar] [CrossRef] [Green Version]
- Hu, F.; Liu, F. Mitochondrial stress: A bridge between mitochondrial dysfunction and metabolic diseases? Cell Signal 2011, 23, 1528–1533. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Chander, V.; Banerjee, P.; Oh, J.G.; Lifirsu, E.; Park, W.J.; Kim, D.H.; Bandyopadhyay, A. Interaction of annexin A6 with alpha actinin in cardiomyocytes. BMC Cell Biol. 2011, 12, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chopra, A.; Tabdanov, E.; Patel, H.; Janmey, P.A.; Kresh, J.Y. Cardiac myocyte remodeling mediated by N-cadherin-dependent mechanosensing. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H1252–H1266. [Google Scholar] [CrossRef] [Green Version]
- Mu, L.; Jing, C.; Guo, Z. Expression of N-cadherin proteins in myocardial hypertrophy in rats. Exp. Ther. Med. 2014, 7, 355–359. [Google Scholar] [CrossRef]
- Kohan, A.B.; Wang, F.; Lo, C.M.; Liu, M.; Tso, P. ApoA-IV: Current and emerging roles in intestinal lipid metabolism, glucose homeostasis, and satiety. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G472–G481. [Google Scholar] [CrossRef] [Green Version]
- Kalogeris, T.J.; Monroe, F.; Demichele, S.J.; Tso, P. Intestinal synthesis and lymphatic secretion of apolipoprotein A-IV vary with chain length of intestinally infused fatty acids in rats. J. Nutr. 1996, 126, 2720–2729. [Google Scholar] [CrossRef] [PubMed]
- Marín-García, J.; Goldenthal, M.J.; Moe, G.W. Post-Genomic Cardiology; Academic Press: Boston, MA, USA, 2007; p. 680. [Google Scholar]
- Ikeda, Y.; Okamura-Ikeda, K.; Tanaka, K. Purification and characterization of short-chain, medium-chain, and long-chain acyl-CoA dehydrogenases from rat liver mitochondria. Isolation of the holo- and apoenzymes and conversion of the apoenzyme to the holoenzyme. J. Biol. Chem. 1985, 260, 1311–1325. [Google Scholar] [PubMed]
- Damian, F.; Van den Mooter, G.; Samyn, C.; Kinget, R. In vitro biodegradation study of acetyl and methyl inulins by Bifidobacteria and inulinase. Eur. J. Pharm. Biopharm. 1999, 47, 275–282. [Google Scholar] [CrossRef]
- Morrison, D.J.; Mackay, W.G.; Edwards, C.A.; Preston, T.; Dodson, B.; Weaver, L.T. Butyrate production from oligofructose fermentation by the human faecal flora: What is the contribution of extracellular acetate and lactate? Br. J. Nutr. 2006, 96, 570–577. [Google Scholar] [CrossRef]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D.; Possemiers, S.; Van de Wiele, T.; Guiot, Y.; Everard, A.; Rottier, O.; Geurts, L.; Naslain, D.; Neyrinck, A.; Lambert, D.M.; et al. Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability. Gut 2009, 58, 1091–1103. [Google Scholar] [CrossRef] [Green Version]
- Yoo, J.Y.; Kim, S.S. Probiotics and prebiotics: Present status and future perspectives on metabolic disorders. Nutrients 2016, 8, 173. [Google Scholar] [CrossRef] [Green Version]
- Bennett, P.M.; Baines, A.J.; Lecomte, M.C.; Maggs, A.M.; Pinder, J.C. Not just a plasma membrane protein: In cardiac muscle cells alpha-II spectrin also shows a close association with myofibrils. J. Muscle Res. Cell Motil. 2004, 25, 119–126. [Google Scholar] [CrossRef]
- Gyorke, I.; Hester, N.; Jones, L.R.; Gyorke, S. The role of calsequestrin, triadin, and junctin in conferring cardiac ryanodine receptor responsiveness to luminal calcium. Biophys. J. 2004, 86, 2121–2128. [Google Scholar] [CrossRef] [Green Version]
- Terentyev, D.; Viatchenko-Karpinski, S.; Gyorke, I.; Volpe, P.; Williams, S.C.; Gyorke, S. Calsequestrin determines the functional size and stability of cardiac intracellular calcium stores: Mechanism for hereditary arrhythmia. Proc. Natl. Acad. Sci. USA 2003, 100, 11759–11764. [Google Scholar] [CrossRef] [Green Version]
- Lima-Leopoldo, A.P.; Leopoldo, A.S.; da Silva, D.C.; do Nascimento, A.F.; de Campos, D.H.; Luvizotto, R.A.; de Deus, A.F.; Freire, P.P.; Medeiros, A.; Okoshi, K.; et al. Long-term obesity promotes alterations in diastolic function induced by reduction of phospholamban phosphorylation at serine-16 without affecting calcium handling. J. Appl. Physiol. 2014, 117, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Ferguson, D.G.; Sako, H.; Dorn, G.W., II; Kadambi, V.J.; Yatani, A.; Hoit, B.D.; Walsh, R.A.; Kranias, E.G. Cardiac-specific overexpression of mouse cardiac calsequestrin is associated with depressed cardiovascular function and hypertrophy in transgenic mice. J. Biol. Chem. 1998, 273, 28470–28477. [Google Scholar] [CrossRef] [Green Version]
- Flashman, E.; Redwood, C.; Moolman-Smook, J.; Watkins, H. Cardiac myosin binding protein C: Its role in physiology and disease. Circ. Res. 2004, 94, 1279–1289. [Google Scholar] [CrossRef]
- Winegrad, S. Cardiac myosin binding protein C. Circ. Res. 1999, 84, 1117–1126. [Google Scholar] [CrossRef] [Green Version]
- Pohlmann, L.; Kroger, I.; Vignier, N.; Schlossarek, S.; Kramer, E.; Coirault, C.; Sultan, K.R.; El-Armouche, A.; Winegrad, S.; Eschenhagen, T.; et al. Cardiac myosin-binding protein C is required for complete relaxation in intact myocytes. Circ. Res. 2007, 101, 928–938. [Google Scholar] [CrossRef]
- Waddingham, M.T.; Edgley, A.J.; Tsuchimochi, H.; Kelly, D.J.; Shirai, M.; Pearson, J.T. Contractile apparatus dysfunction early in the pathophysiology of diabetic cardiomyopathy. World J. Diabetes 2015, 6, 943–960. [Google Scholar] [CrossRef] [PubMed]
- Rundell, V.L.; Geenen, D.L.; Buttrick, P.M.; de Tombe, P.P. Depressed cardiac tension cost in experimental diabetes is due to altered myosin heavy chain isoform expression. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H408–H413. [Google Scholar] [CrossRef]
- Agarkova, I.; Perriard, J.C. The M-band: An elastic web that crosslinks thick filaments in the center of the sarcomere. Trends Cell Biol. 2005, 15, 477–485. [Google Scholar] [CrossRef]
- Pask, H.T.; Jones, K.L.; Luther, P.K.; Squire, J.M. M-band structure, M-bridge interactions and contraction speed in vertebrate cardiac muscles. J. Muscle Res. Cell Motil. 1994, 15, 633–645. [Google Scholar] [CrossRef]
- Rozanski, A.; Takano, A.P.; Kato, P.N.; Soares, A.G.; Lellis-Santos, C.; Campos, J.C.; Ferreira, J.C.; Barreto-Chaves, M.L.; Moriscot, A.S. M-protein is down-regulated in cardiac hypertrophy driven by thyroid hormone in rats. Mol. Endocrinol. 2013, 27, 2055–2065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paudyal, A.; Dewan, S.; Ikie, C.; Whalley, B.J.; de Tombe, P.P.; Boateng, S.Y. Nuclear accumulation of myocyte muscle LIM protein is regulated by heme oxygenase 1 and correlates with cardiac function in the transition to failure. J. Physiol. 2016, 594, 3287–3305. [Google Scholar] [CrossRef]
- Communal, C.; Sumandea, M.; de Tombe, P.; Narula, J.; Solaro, R.J.; Hajjar, R.J. Functional consequences of caspase activation in cardiac myocytes. Proc. Natl. Acad. Sci. USA 2002, 99, 6252–6256. [Google Scholar] [CrossRef] [Green Version]
- Fahien, L.A.; Macdonald, M.J. The complex mechanism of glutamate dehydrogenase in insulin secretion. Diabetes 2011, 60, 2450–2454. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.K.; Thu, V.T.; Heo, H.J.; Kim, N.; Han, J. Cardiac proteomic responses to ischemia-reperfusion injury and ischemic preconditioning. Expert Rev. Proteom. 2011, 8, 241–261. [Google Scholar] [CrossRef]
- De Vadder, F.; Kovatcheva-Datchary, P.; Zitoun, C.; Duchampt, A.; Backhed, F.; Mithieux, G. Microbiota-Produced Succinate Improves Glucose Homeostasis via Intestinal Gluconeogenesis. Cell Metab. 2016, 24, 151–157. [Google Scholar] [CrossRef] [Green Version]
- Brownlee, M. Biochemistry and molecular cell biology of diabetic complications. Nature 2001, 414, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, N.; Thornalley, P.J. Glyoxalase in diabetes, obesity and related disorders. Semin. Cell Dev. Biol. 2011, 22, 309–317. [Google Scholar] [CrossRef]
- Brouwers, O.; de Vos-Houben, J.M.; Niessen, P.M.; Miyata, T.; van Nieuwenhoven, F.; Janssen, B.J.; Hageman, G.; Stehouwer, C.D.; Schalkwijk, C.G. Mild oxidative damage in the diabetic rat heart is attenuated by glyoxalase-1 overexpression. Int. J. Mol. Sci. 2013, 14, 15724–15739. [Google Scholar] [CrossRef] [Green Version]
- Kellow, N.J.; Coughlan, M.T.; Savige, G.S.; Reid, C.M. Effect of dietary prebiotic supplementation on advanced glycation, insulin resistance and inflammatory biomarkers in adults with pre-diabetes: A study protocol for a double-blind placebo-controlled randomised crossover clinical trial. BMC Endocr. Disord. 2014, 14, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hojlund, K.; Wrzesinski, K.; Larsen, P.M.; Fey, S.J.; Roepstorff, P.; Handberg, A.; Dela, F.; Vinten, J.; McCormack, J.G.; Reynet, C.; et al. Proteome analysis reveals phosphorylation of ATP synthase beta -subunit in human skeletal muscle and proteins with potential roles in type 2 diabetes. J. Biol. Chem. 2003, 278, 10436–10442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varma, U.; Koutsifeli, P.; Benson, V.L.; Mellor, K.M.; Delbridge, L.M.D. Molecular mechanisms of cardiac pathology in diabetes—Experimental insights. Biochim. Biophys. Acta Mol. Basis. Dis. 2018, 1864, 1949–1959. [Google Scholar] [CrossRef]
- So, D.; Whelan, K.; Rossi, M.; Morrison, M.; Holtmann, G.; Kelly, J.T.; Shanahan, E.R.; Staudacher, H.M.; Campbell, K.L. Dietary fiber intervention on gut microbiota composition in healthy adults: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2018, 107, 965–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COMPOSITION | CON | HF | HF-FOS |
|---|---|---|---|
| Ingredient (g) | |||
| Casein (protein) | 200 | 190 | 190 |
| L-Cystine | 3 | 3 | 3 |
| Corn Starch | 387.5 | 173.5 | 172.6 |
| Maltodextrin 10 | 122 | 68.3 | 67.6 |
| Sucrose | 100 | 283.3 | 282.6 |
| Cellulose, BW200 | 70 | 70 | 40 |
| Fructooligosaccharide | 0 | 0 | 32.2 |
| Soybean Oil | 70 | 120.3 | 120.3 |
| Milk Fat, Anhydrous | 0 | 44.2 | 44.2 |
| tBHQ | 0.014 | 0.014 | 0.014 |
| Mineral Mix S10022G | 35 | 35 | 35 |
| Nutrient (g) | |||
| Protein | 179.0 | 170.2 | 170.2 |
| Carbohydrate (digestible) | 609.5 | 524.9 | 522.8 |
| Fat | 70.0 | 164.5 | 164.5 |
| Fibre | 70.0 | 70.0 | 72.2 |
| Nutrient (kcal) | |||
| Protein | 716 | 681 | 681 |
| Carbohydrate | 2438 | 2100 | 2091 |
| Fat | 630 | 1481 | 1481 |
| Total | 3784 | 4261 | 4253 |
| Biological Theme * | Gene Names ‡ | GO and KEGG Pathways † | p-Value § |
|---|---|---|---|
| Metabolism (cholesterol) | apolipoprotein A1 (Apoa1) apolipoprotein A4 (Apoa4) apolipoprotein E (Apoe) catalase (Cat) hydroxysteroid (17-beta) dehydrogenase 4 (Hsd17b4) | Cholesterol metabolic process | 3.3 × 10-4 |
| KEGG: | |||
| Fat digestion and absorption | 3.7 × 10-1 | ||
| Peroxisome | 1.6 × 10-1 | ||
| Vitamin digestion and absorption | 3.6 × 10-1 | ||
| Metabolism (lipid) | apolipoprotein A1 (Apoa1) apolipoprotein A4 (Apoa4) | VLDL remodelling | 3.5 × 10-3 |
| apolipoprotein E (Apoe) | Positive regulation of cholesterol esterification | 3.5 × 10-3 | |
| HDL particle assembly | 3.5 × 10-3 | ||
| Peripheral nervous system axon regeneration | 6.5 × 10-3 | ||
| Reverse cholesterol transport | 6.5 × 10-3 | ||
| Phospholipid efflux | 7.5 × 10-3 | ||
| Triglyceride catabolic process | 2.1 × 10-2 | ||
| Triglyceride homeostasis | 2.0 × 10-2 | ||
| Cholesterol Efflux | 1.9 × 10-2 | ||
| Lipoprotein metabolic process | 1.9 × 10-2 | ||
| KEGG: | |||
| Fat digestion and absorption | 3.7 × 10-1 | ||
| Vitamin digestion and absorption | 3.6 × 10-1 | ||
| Differentially Expressed Protein | Protein Accession No. | Log2Fold Change (HF-FOS/HF) * | p-Value † | Gene Name |
|---|---|---|---|---|
| apolipoprotein A-IV (APOA4) | EDL95397.1 | −0.16 | <0.0001 | Apoa4 |
| catalase, isoform CRA_b (CAT) | EDL79667.1 | −0.07 | 0.0011 | Cat |
| creatine kinase, brain, isoform CRA_b (CKB) | EDL97457.1 | −0.14 | <0.0001 | Ckb |
| electron transferring flavoprotein, alpha polypeptide (ETFA) | EDL95569.1 | 0.06 | 0.0010 | Etfa |
| filamin, alpha (predicted), isoform CRA_b (FLNA) | EDL84990.1 (+1) | −0.12 | <0.0001 | Flna |
| four and a half LIM domains 1, isoform CRA_b (FHL1) | EDL75140.1 | −0.13 | 0.0003 | Fhl1 |
| PREDICTED: annexin A6 isoform X1 (ANXA6) | XP_017453031.1 | 0.09 | <0.0001 | Anxa6 |
| preproapolipoprotein A-I (APOA1) | CAA25224.1 | −0.16 | <0.0001 | Apoa1 |
| stress-70 protein, mitochondrial (HSPA9) | NP_001094128.2 | −0.06 | <0.0001 | Hspa9 |
| myosin regulatory light chain 2, ventricular/cardiac muscle isoform (MLY2) | NP_001030329.2 | 0.16 | 0.0001 | Myl2 |
| PREDICTED: guanylate-binding protein 1 (GBP1) | XP_006224340.1 (+1) | 0.21 | 0.0002 | Gbp1 |
| tropomyosin 4 (TPM4) | EDL90830.1 | −0.16 | 0.0002 | Tpm4 |
| glycerol-3-phosphate dehydrogenase 1-like protein (GPD11) | NP_001178814.1 | −0.12 | <0.0001 | Gpd1l |
| heat shock protein, alpha-crystallin-related, B6/ Hsp20 (HSPB6) | EDM07749.1 | −0.11 | 0.0026 | Hspb6 |
| LIM and cysteine-rich domains protein 1 (LMCD1) | NP_001008562.1 | −0.13 | <0.0001 | Lmcd1 |
| myomesin-1 (MYOM1) | NP_001178513.1 | 0.09 | <0.0001 | Myom1 |
| nebulette (NEB1) | NP_001178623.1 | −0.09 | <0.0001 | Nebl |
| nexilin, isoform CRA_c (NEXN) | EDL82495.1 (+1) | −0.09 | 0.0004 | Nexn |
| PREDICTED: cAMP-dependent protein kinase type I-alpha regulatory subunit isoform X1 (PRKAR1a) | XP_017452536.1 | −0.07 | 0.0015 | Prkar1a |
| PREDICTED: cysteine and glycine-rich protein 3 isoform X1/ cardiac LIM protein (CSRP3) | XP_006229299.1 | −0.11 | 0.0005 | Csrp3 |
| PREDICTED: heat shock protein HSP 90-alpha (LOC103692716) | XP_008763191.1 | −0.07 | 0.0013 | LOC103692716 |
| PREDICTED: myomesin-2 isoform X1 (MYOM2) | XP_017455650.1 | 0.06 | <0.0001 | Myom2 |
| actinin alpha 2 (ACTN2) | EDM06982.1 | 0.1 | <0.0001 | Actn2 |
| ROK-alpha (Rho-associated protein expression kinase) (ROCK2) | AAB37540.1 (+1) | −0.1 | 0.0029 | Rock2 |
| sarcoplasmic/endoplasmic reticulum calcium ATPase 2 isoform a (ATP2A2) | NP_001103609.1 | 0.04 | <0.0001 | Atp2a2 |
| vinculin (predicted), isoform CRA_a (VCL) | EDL86257.1 | −0.05 | 0.0002 | Vcl |
| acetyl-Coenzyme A dehydrogenase, short chain, isoform CRA_a (ACADS) | EDM13909.1 | 0.06 | 0.0003 | Acads |
| aldehyde dehydrogenase (ALDH1A1) | AAA96657.1 | 0.25 | 0.0004 | Aldh1a1 |
| kynurenine—oxoglutarate transaminase 1, mitochondrial (KYAT1) | NP_001013182.3 | −0.24 | < 0.0001 | Kyat1 |
| PREDICTED: mimitin, mitochondrial (NDUFAF2) | XP_001073799.1 | −0.18 | 0.0014 | Ndufaf2 |
| prolyl endopeptidase (PREP) | EDL99674.1 (+1) | −0.16 | 0.0005 | Prep |
| pyridine nucleotide-disulfide oxidoreductase domain-containing protein 2 (PYRDOXD2) | NP_001004261.1 (+3) | −0.27 | <0.0001 | Pyroxd2 |
| cytochrome c-1 (predicted), isoform CRA_c (CYC1) | EDM15989.1 | 0.13 | <0.0001 | Cyc1 |
| glutathione S-transferase omega 1 (GSTO1) | ACI32122.1 | −0.12 | 0.0036 | Gsto1 |
| heat shock 70kDa protein 5 (glucose-regulated protein), isoform CRA_a (HSPA5) | EDL93170.1 | −0.08 | <0.0001 | Hspa5 |
| inter alpha-trypsin inhibitor, heavy chain 4, isoform CRA_a (LTIH4) | EDL88978.1 | −0.12 | <0.0001 | Ltih4 |
| kynurenine—oxoglutarate transaminase 3 (KYAT3) | NP_001015037.1 | −0.13 | 0.0006 | Kyat3 |
| leucine-rich PPR motif-containing protein, mitochondrial precursor (LRPPRC) | NP_001008519.1 | −0.06 | <0.0001 | Lrpprc |
| methylcrotonoyl-CoA carboxylase beta chain, mitochondrial (MCCC2) | NP_001012177.1 | −0.07 | <0.0001 | Mccc2 |
| methylcrotonoyl-CoA carboxylase subunit alpha, mitochondrial (MCCC1) | NP_001009653.1 | −0.09 | <0.0001 | Mccc1 |
| muscle glycogen phosphorylase (PYGM) | EDM12601.1 | 0.07 | <0.0001 | Pygm |
| Pgm1 protein, partial (PGM1) | AAI28704.1 | −0.05 | 0.0003 | Pgm1 |
| PREDICTED: isobutyryl-CoA dehydrogenase, mitochondrial isoform X2 (ACAD8) | XP_003754442.1 | −0.07 | 0.0025 | Acad8 |
| PREDICTED: triosephosphate isomerase (LOC100911515) | XP_003750702.2 | −0.06 | <0.0001 | LOC100911515 |
| rCG20653 (ACOT2) | EDL81468.1 | 0.07 | 0.0002 | Acot2 |
| rCG45082 (C3) | EDL83571.1 | −0.05 | <0.0001 | C3 |
| Tu translation elongation factor, mitochondrial (predicted), isoform CRA_c (TUFM) | EDM17456.1 | −0.05 | 0.0001 | Tufm |
| ubiquinol cytochrome c reductase core protein 2, isoform CRA_c (UQCRC2) | EDM17618.1 | 0.05 | 0.0005 | Uqcrc2 |
| glutathione peroxidase 1 (GPX1) | NP_110453.3 | −0.1 | 0.0003 | Gpx1 |
| arginine-tRNA-protein transferase 1 (predicted), isoform CRA_a (ATE1) | EDM17146.1 | −0.21 | 0.0026 | Ate1 |
| DnaJ (Hsp40) homolog, subfamily A, member 2, isoform CRA_b (DNAJ) | EDL87492.1 | −0.15 | <0.0001 | DnaJ |
| rCG22471, isoform CRA_b (EEF1B2) | EDL98899.1 | −0.21 | <0.0001 | Eef1b2 |
| chaperonin subunit 8 (theta) (predicted), isoform CRA_a (CCT8) | EDM10642.1 | −0.06 | 0.0026 | Cct8 |
| crystallin, alpha B, isoform CRA_a (CRYAB) | EDL95479.1 | −0.12 | <0.0001 | Cryab |
| guanosine diphosphate dissociation inhibitor 1, isoform CRA_a (GDI1) | EDL84976.1 | −0.08 | 0.0011 | Gdi1 |
| heat shock protein 27 (HSPB1) | AAA41353.1 | −0.12 | <0.0001 | Hspb1 |
| peptidylprolyl isomerase F (cyclophilin F), isoform CRA_a (PPIF) | EDL75095.1 | −0.07 | 0.0026 | Ppif |
| proteasomal ATPase (rat TBP1) (PSMC3) | BAA11939.1 | −0.11 | 0.0003 | Psmc3 |
| proteasome (prosome, macropain) subunit, alpha type 1, isoform CRA_d (PSMA1) | EDM17781.1 | −0.09 | 0.0037 | Psma1 |
| rCG38543, isoform CRA_a (PSMA7) | EDL88834.1 | −0.07 | 0.0013 | Psma7 |
| drebrin-like, isoform CRA_b (DBN1) | EDM00307.1 | −0.17 | 0.0011 | Dbnl |
| oxidative-stress responsive 1 (predicted) (OXSR1) | EDL76918.1 | −0.16 | 0.0019 | Oxsr1 |
| Ehd1 protein, partial (EHD1) | AAI60908.1 | −0.08 | <0.0001 | Ehd1 |
| EH-domain containing 4 (EHD4) | EDL79933.1 | −0.1 | <0.0001 | Ehd4 |
| heat shock 27kD protein family, member 7 (cardiovascular) (HSPB7) | EDL80988.1 | −0.12 | 0.0038 | Hspb7 |
| nonmuscle myosin heavy chain-A (MYH9) | AAA74950.1 | −0.12 | <0.0001 | Myh9 |
| PDZ and LIM domain 1 (elfin), isoform CRA_a (PDLIM1) | EDL94180.1 | −0.1 | 0.0017 | Pdlim1 |
| PREDICTED: adenylyl cyclase-associated protein 2 isoform X1 (CAP2) | XP_006253816.1 | −0.12 | < 0.0001 | Cap2 |
| PREDICTED: beta-1-syntrophin isoform X1 (SNTB1) | XP_017450243.1 | −0.11 | < 0.0001 | Sntb1 |
| PREDICTED: spectrin alpha chain, non-erythrocytic 1 isoform X1 (SPTAN1) | XP_008759895.1 | 0.06 | 0.0004 | Sptan1 |
| PREDICTED: tubulin-folding cofactor B (LOC103690005) | XP_017444696.1 | −0.2 | 0.0011 | LOC103690005 |
| rCG27764, isoform CRA_a (DYNC1H1) | EDL97508.1 | −0.08 | <0.0001 | |
| acyl-CoA synthetase family member 2, mitochondrial precursor (ACSF2) | NP_001030123.1 | 0.04 | 0.0012 | Acsf2 |
| acyl-Coenzyme A dehydrogenase, very long chain, isoform CRA_c (ACADVL) | EDM04964.1 | 0.06 | <0.0001 | Acadvl |
| arginine--tRNA ligase, cytoplasmic (RARS) | NP_001099247.2 | −0.11 | 0.0002 | Rars |
| catenin beta-1 (CTNNB1) | NP_445809.2 (+1) | −0.08 | 0.0026 | Ctnnb1 |
| calsequestrin 2, isoform CRA_a (CASQ2) | EDL85514.1 | −0.05 | 0.0037 | Casq2 |
| cellular nucleic acid binding protein 1, isoform CRA_b (CNBP) | EDL91294.1 | −0.12 | 0.0036 | Cnbp |
| enoyl coenzyme A hydratase 1, peroxisomal (ECH1) | EDM07870.1 | −0.1 | <0.0001 | Ech1 |
| glutamate dehydrogenase 1, isoform CRA_a (GLUD1) | EDL88881.1 | −0.05 | 0.0019 | Glud1 |
| GrpE-like 1, mitochondrial (GRPEL1) | EDM00047.1 | −0.1 | 0.0007 | Grpel1 |
| Hagh protein, partial (HAGH) | AAH97301.1 | −0.1 | 0.0025 | Hagh |
| Keratin 5 (KRT5) | AAH62086.1 | −0.56 | 0.0018 | Krt5 |
| L-3-hydroxyacyl-Coenzyme A dehydrogenase, short chain, isoform CRA_a (HADH) | EDL82212.1 | 0.09 | 0.0002 | Hadh |
| PREDICTED: glycogen phosphorylase, brain form isoform X1 (PYGB) | XP_017446988.1 | 0.05 | 0.0026 | Pygb |
| PREDICTED: microtubule-associated protein 4 isoform X1 (MAP4) | XP_006243875.1 | −0.1 | 0.001 | Map4 |
| PREDICTED: myosin-binding protein C, cardiac-type isoform X1 (MYBPC3) | XP_006234566.1 | 0.07 | <0.0001 | Mybpc3 |
| thioredoxin 2, isoform CRA_b (TXN2) | EDM15902.1 | −0.27 | 0.0034 | Txn2 |
| TPA_exp: type II keratin Kb1 (KRT1) | DAA02055.1 | −0.43 | 0.0038 | Krt1 |
| transcription factor A, mitochondrial, isoform CRA_a (TFAM) | EDL97257.1 | −0.1 | 0.0023 | Tfam |
| glutamate oxaloacetate transaminase 1, isoform CRA_a (GOT1) | EDL94253. | −0.07 | <0.0001 | Got1 |
| Gpc1 protein (GPC1) | AAH61572.1 | −0.08 | 0.0005 | Gpc1 |
| isocitrate dehydrogenase 3 (NAD+) alpha, isoform CRA_a (IDH3a) | EDL95540.1 | −0.06 | 0.0038 | Idh3a |
| PREDICTED: myosin-7 isoform X2 (MYH7) | XP_006252013.1 | −0.17 | <0.0001 | Myh7 |
| ATPase family, AAA domain containing 3A (ATAD3A) | EDL81320.1 | −0.17 | 0.0009 | Atad3a |
| glyoxylase 1 (GLO1) | EDL96989.1 | −0.14 | 0.0007 | Glo1 |
| PREDICTED: pre-B-cell leukemia transcription factor-interacting protein 1 isoform X4 (PBXIP1) | XP_017446382.1 | −0.14 | 0.0012 | Pbxip1 |
| PREDICTED: solute carrier family 12 member 7 isoform X2 (SLC12A7) | XP_006227851.1 | −0.19 | <0.0001 | Slc12a7 |
| PREDICTED: transportin-1 isoform X1 (TNPO1) | XP_008758890.1 | −0.25 | 0.0029 | Tnpo1 |
| AFG3-like protein 2 (AFG312) | NP_001128336.1 | −0.08 | 0.0002 | Afg3l2 |
| annexin A5 (ANXA5) | NP_037264.1 (+ 4) | −0.12 | <0.0001 | Anxa5 |
| clathrin heavy chain 1 (CLTC) | NP_062172.1 | −0.06 | <0.0001 | Cltc |
| complement inhibitory factor H (CFH) | CAC67513.1 | −0.09 | <0.0001 | Cfh |
| elongation factor 1-delta (EEFLD) | NP_001013122.1 | −0.09 | <0.0001 | Eef1d |
| fibrinogen B beta chain (FGB) | AAA64866.1 | −0.1 | <0.0001 | Fgb |
| inter-alpha-trypsin inhibitor heavy chain H3 precursor (ITIH3) | NP_059047.1 (+ 4) | −0.13 | 0.0033 | Itih3 |
| lumican (LUM) | EDM16832.1 | 0.06 | 0.0029 | Lum |
| PREDICTED: dynamin-like 120 kDa protein, mitochondrial isoform X1 (OPA1) | XP_006248559.1 | −0.07 | <0.0001 | Opa1 |
| PREDICTED: fibrinogen alpha chain isoform X1 (FGA) | XP_006232594.1 | −0.13 | <0.0001 | Fga |
| rat ribosomal protein L13a (RPL13A) | CAA48343.1 | −0.11 | 0.0010 | Rpl13a |
| rCG27551, isoform CRA_b (DSTN) | EDL95182.1 | −0.1 | 0.0017 | Dstn |
| rCG35863 (LAP3) | EDL99928.1 | −0.11 | <0.0001 | Lap3 |
| selenium-binding protein 1 (LOC103689947) | NP_001316822.1 | −0.06 | 0.0023 | LOC103689947 |
| PREDICTED: similar to RIKEN cDNA 2310039E09 (CAVIN4) | EDL78187.1 | −0.13 | 0.0023 | Cavin4 |
| Sucla2 protein, partial; succinate-coA ligase subunit beta (SUCLA2) | AAI66998.1 | −0.06 | 0.0003 | Sucla2 |
| thioredoxin-like 5 (predicted), isoform CRA_b (TXNDC17) | EDM05095.1 | −0.09 | 0.0014 | Txndc17 |
| Biological Theme * | Gene Names ‡ | GO Terms and KEGG Pathways † | p-Value § |
|---|---|---|---|
| Fatty acid homeostasis | acyl-CoA dehydrogenase family, member 8 (Acad8) acyl-CoA dehydrogenase, C-2 to C-3 short chain (Acads) acyl-CoA dehydrogenase, very long chain (Acadvl) apolipoprotein A4 (Apoa4) electron transfer flavoprotein alpha subunit (Etfa) | GO-lipid homeostasis | 3.0 × 10−2 |
| acyl-CoA dehydrogenase family, member 8 (Acad8) acyl-CoA dehydrogenase, C-2 to C-3 short chain (Acads) acyl-CoA dehydrogenase, very long chain (Acadvl) electron transfer flavoprotein alpha subunit (Etfa) | GO-fatty acid beta-oxidation using acyl-CoA dehydrogenase | 3.8 × 10-2 | |
| Cardiac muscle regulation | calsequestrin 2 (Casq2) cysteine and glycine rich protein 3 (Csrp3) myosin binding protein C, cardiac (Mybpc3) myosin heavy chain 7 (Myh7) myosin light chain 2 (Myl2) | GO-cardiac muscle contraction | 3.6 × 10-2 |
| actinin alpha 2 (Actn2) myomesin 1 (Myom1) myomesin 2 (Myom2) myosin binding protein C, cardiac (Mybpc3) myosin heavy chain 7 (Myh7) tropomyosin 4 (Tpm4) | GO-muscle contraction | 6.6 × 10-3 | |
| Carbohydrate metabolism | acyl-CoA dehydrogenase, C-2 to C-3 short chain (Acads) catalase (Cat) glutamate dehydrogenase 1 (Glud1) glutamic-oxaloacetic transaminase 1 (Got1) isocitrate dehydrogenase 3 (NAD+) alpha (Idh3a) succinate-CoA ligase ADP-forming beta subunit t (Sucla2) triosephosphate isomerase-like (LOC100911515) | KEGG-carbon metabolism | 7.3 × 10-2 |
| glycerol-3-phosphate dehydrogenase 1-like (Gpd1l) glyoxalase 1(Glo1) hydroxyacyl glutathione hydrolase (Hagh) phosphoglucomutase 1 (Pgm1) phosphorylase, glycogen, muscle (Pygm) phosphorylase, glycogen; brain (Pygb) | GO-carbohydrate metabolic process | 8.8 × 10-2 | |
| Protein processes | GrpE-like 1, mitochondrial (Grpel1) chaperonin containing TCP1 subunit 8 (Cct8) crystallin, alpha B (Cryab) heat shock protein HSP 90-alpha (LOC103692716) heat shock protein family A member 9 (Hspa9) peptidylprolyl isomerase F (Ppif) thioredoxin 2 (Txn2) | KEGG-Protein folding | 2.1 × 10-2 |
| Adhesion and cytoskeletal regulation | PDZ and LIM domain 1 (Pdlim1) arginyl-tRNA synthetase (Rars) chaperonin containing TCP1 subunit 8 (Cct8) drebrin-like protein (Dbnl) eukaryotic translation elongation factor 1 delta (Eef1d) heat shock protein family A member 5 (Hspa5) spectrin, alpha, non-erythrocytic 1 (Sptan1) | GO-cell-cell adhesion | 4.5 × 10-2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the Her Majesty the Queen in Right of Canada as represented by the Agriculture and Agri-Food Canada; Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarfaraz, S.; Singh, S.; Hawke, A.; Clarke, S.T.; Ramdath, D.D. Effects of High-Fat Diet Induced Obesity and Fructooligosaccharide Supplementation on Cardiac Protein Expression. Nutrients 2020, 12, 3404. https://doi.org/10.3390/nu12113404
Sarfaraz S, Singh S, Hawke A, Clarke ST, Ramdath DD. Effects of High-Fat Diet Induced Obesity and Fructooligosaccharide Supplementation on Cardiac Protein Expression. Nutrients. 2020; 12(11):3404. https://doi.org/10.3390/nu12113404
Chicago/Turabian StyleSarfaraz, Sidra, Shamjeet Singh, Aileen Hawke, Sandra T. Clarke, and D. Dan Ramdath. 2020. "Effects of High-Fat Diet Induced Obesity and Fructooligosaccharide Supplementation on Cardiac Protein Expression" Nutrients 12, no. 11: 3404. https://doi.org/10.3390/nu12113404
APA StyleSarfaraz, S., Singh, S., Hawke, A., Clarke, S. T., & Ramdath, D. D. (2020). Effects of High-Fat Diet Induced Obesity and Fructooligosaccharide Supplementation on Cardiac Protein Expression. Nutrients, 12(11), 3404. https://doi.org/10.3390/nu12113404