High Variability in Erythrocyte, Plasma and Whole Blood EPA and DHA Levels in Response to Supplementation



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Study Design
2.3. Clinic Visits
2.4. Dietary Intake Analysis
2.5. Blood Sample Collection and Processing
2.6. Fatty Acid Analysis
2.7. Statistical Analysis
3. Results
3.1. Study Participants Characteristics
3.2. Dietary Intakes and Compliance to Treatment
3.3. Variability of Baseline Dietary EPA Plus DHA (g/day) and Erythrocyte EPA Plus DHA (% of Total Fatty Acids)
3.4. Fatty Acid Response to n-3 LCPUFA Supplementation
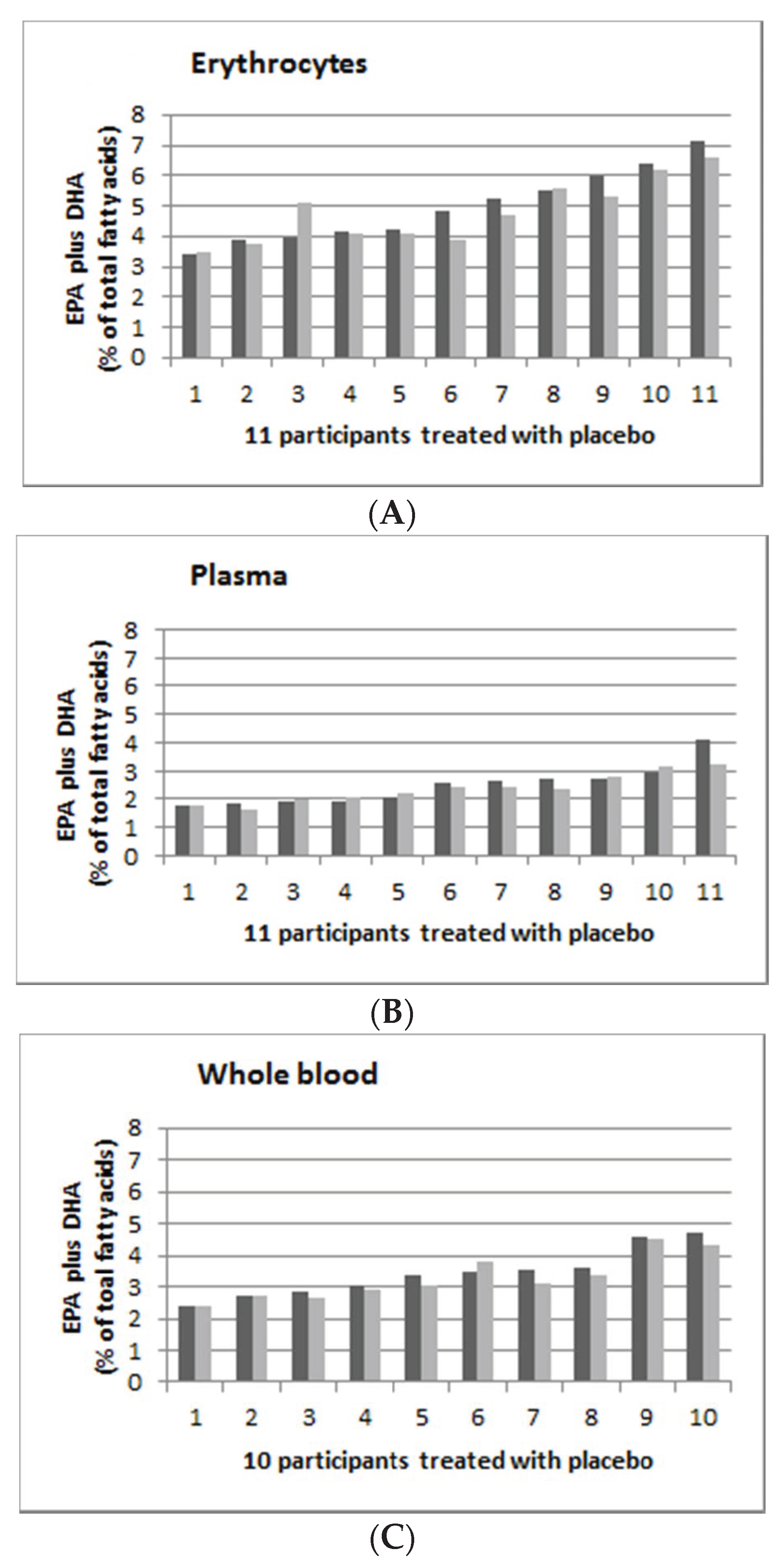
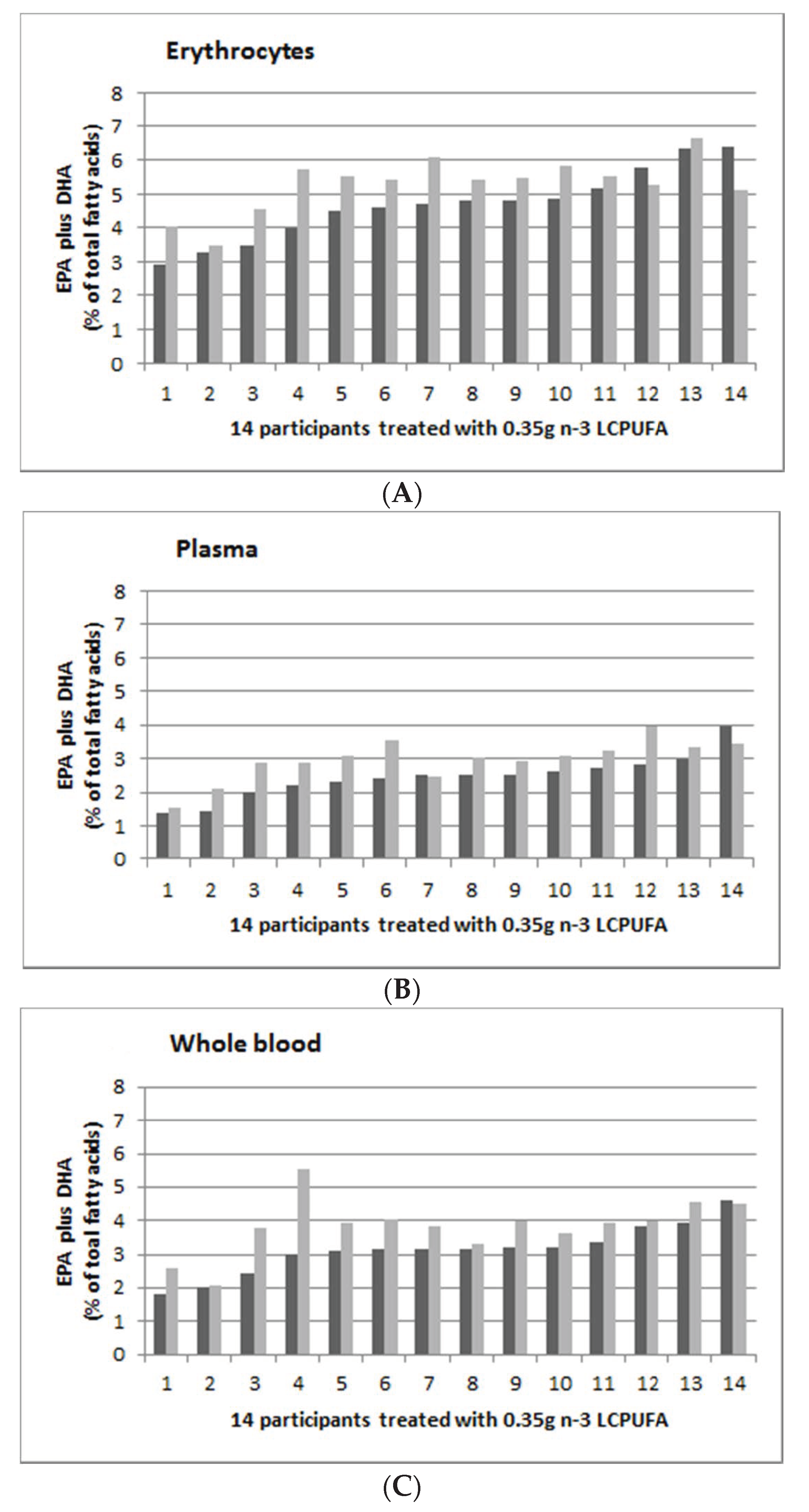
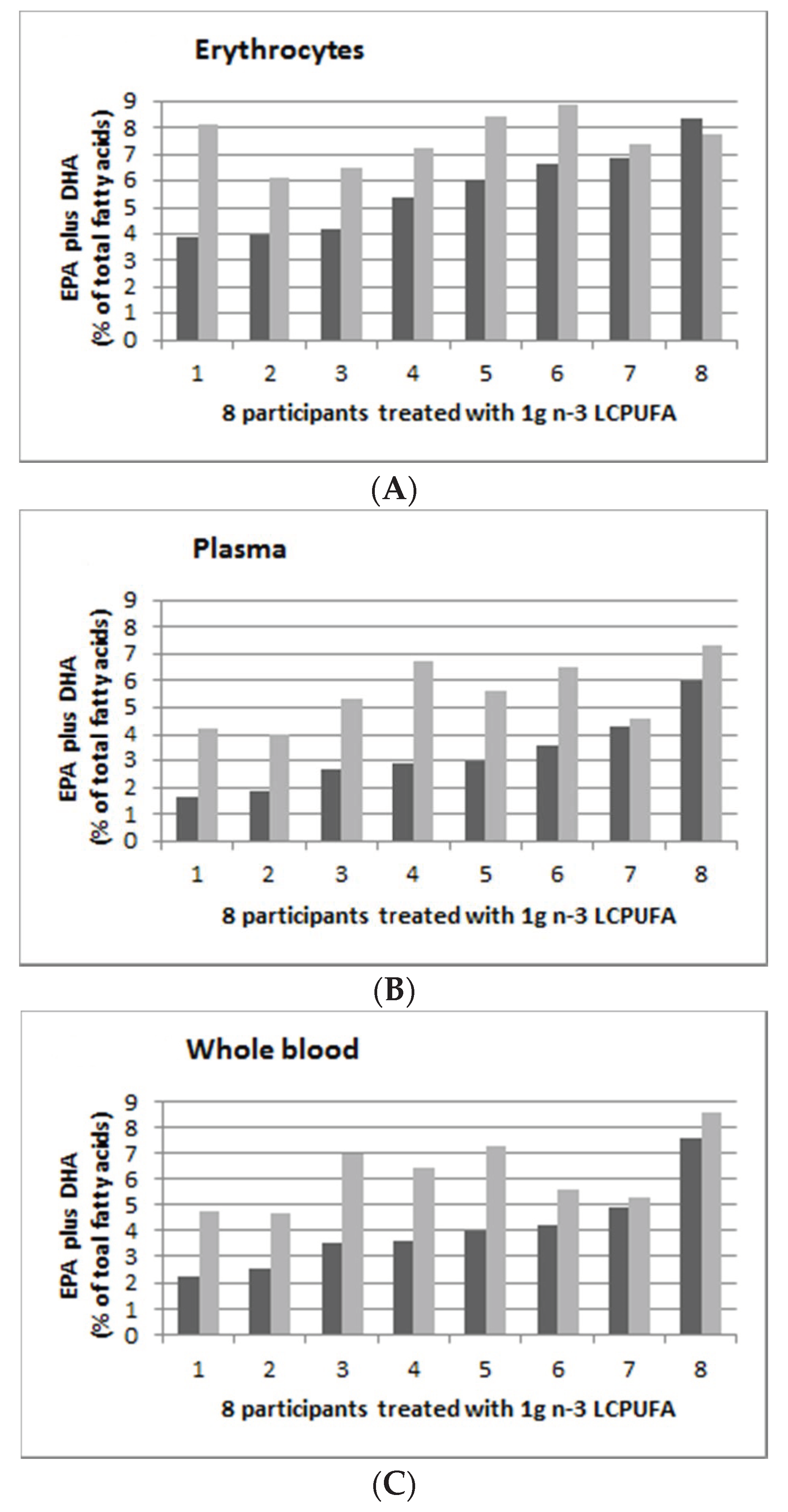
3.5. Individual Response to n-3 LCPUFA Supplementation
4. Discussion
Strengths and Limitations
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| n-3 LCPUFA | Omega-3 long chain polyunsaturated fatty acids |
| EPA | eicosapentaenoic acid |
| DPA | docosapentaenoic acid |
| DHA | docosahexaenoic acid |
| LA | Linoleic acid |
| AA | Arachidonic acid |
| ALA | Alpha-linolenic acid |
| EDTA | ethylenediaminetetraacetic acid |
| FAME | fatty acid methyl esters |
| BMI | body mass index |
| SBP | systolic blood pressure |
| DBP | diastolic blood pressure |
| IQR | Interquartile range |
| L MUFA | Monounsaturated fatty acids |
| PUFA | Polyunsaturated fatty acids |
| SFA | Saturated fatty acids |
References
- Makrides, M.; Gibson, R.A. Long-chain polyunsaturated fatty acid requirements during pregnancy and lactation. Am. J. Clin. Nutr. 2000, 71, 307S–311S. [Google Scholar] [CrossRef] [Green Version]
- Meyer, B.J.; Onyiaodike, C.C.; Brown, E.A.; Jordan, F.; Murray, H.; Nibbs, R.J.; Sattar, N.; Lyall, H.; Nelson, S.M.; Freeman, D.J. Maternal plasma dha levels increase prior to 29 days post-lh surge in women undergoing frozen embryo transfer: A prospective, observational study of human pregnancy. J. Clin. Endocrinol. Metab. 2016, 101, 1745–1753. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, A. Docosahexaenoic acid and the brain– what is its role? Asia Pac. J. Clin. Nutr. 2019, 28, 675–688. [Google Scholar] [PubMed]
- GISSI Prevezione Investigators. Dietary supplementation with n-3 polyunsaturated fatty acids and vitamin e after myocardial infarction: Results of the gissi-prevenzione trial. Lancet 1999, 354, 447–455. [Google Scholar] [CrossRef]
- Yokoyama, M.; Origasa, H.; Matsuzaki, M.; Matsuzawa, Y.; Saito, Y.; Ishikawa, Y.; Oikawa, S.; Sasaki, J.; Hishida, H.; Itakura, H.; et al. Effects of eicosapentaenoic acid on major coronary events in hypercholesterolaemic patients (jelis): A randomised open-label, blinded endpoint analysis. Lancet 2007, 369, 1090–1098. [Google Scholar] [CrossRef]
- Hino, A.; Adachi, H.; Toyomasu, K.; Yoshida, N.; Enomoto, M.; Hiratsuka, A.; Hirai, Y.; Satoh, A.; Imaizumi, T. Very long chain n-3 fatty acids intake and carotid atherosclerosis: An epidemiological study evaluated by ultrasonography. Atherosclerosis 2004, 176, 145–149. [Google Scholar] [CrossRef]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta 2015, 1851, 469–484. [Google Scholar] [CrossRef]
- Proudman, S.M.; Cleland, L.G.; Metcalf, R.G.; Sullivan, T.R.; Spargo, L.D.; James, M.J. Plasma n-3 fatty acids and clinical outcomes in recent-onset rheumatoid arthritis. Br. J. Nutr. 2015, 114, 885–890. [Google Scholar] [CrossRef] [Green Version]
- Parletta, N.; Zarnowiecki, D.; Cho, J.; Wilson, A.; Procter, N.; Gordon, A.; Bogomolova, S.; O’Dea, K.; Strachan, J.; Ballestrin, M.; et al. People with schizophrenia and depression have a low omega-3 index. Prost. Leukot. Essent. Fat. Acids 2016, 110, 42–47. [Google Scholar] [CrossRef] [Green Version]
- Parletta, N.; Zarnowiecki, D.; Cho, J.; Wilson, A.; Bogomolova, S.; Villani, A.; Itsiopoulos, C.; Niyonsenga, T.; Blunden, S.; Meyer, B.; et al. A mediterranean-style dietary intervention supplemented with fish oil improves diet quality and mental health in people with depression: A randomized controlled trial (helfimed). Nutr. Neurosci. 2019, 22, 474–487. [Google Scholar] [CrossRef] [Green Version]
- Alqarni, A.; Mitchell, T.W.; McGorry, P.D.; Nelson, B.; Markulev, C.; Yuen, H.P.; Schafer, M.R.; Berger, M.; Mossaheb, N.; Schlogelhofer, M.; et al. Comparison of erythrocyte omega-3 index, fatty acids and molecular phospholipid species in people at ultra-high risk of developing psychosis and healthy people. Schizophr. Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.Y.; Mischoulon, D.; Freeman, M.P.; Matsuoka, Y.; Hibbeln, J.; Belmaker, R.H.; Su, K.P. Are omega-3 fatty acids antidepressants or just mood-improving agents? The effect depends upon diagnosis, supplement preparation, and severity of depression. Mol. Psychiatr. 2012, 17, 1161–1163. [Google Scholar] [CrossRef]
- Harris, W.S.; Von Schacky, C. The omega-3 index: A new risk factor for death from coronary heart disease? Prev. Med. 2004, 39, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Cadenhead, K.S.; Minichino, A.; Kelsven, S.; Addington, J.; Bearden, C.; Cannon, T.D.; Cornblatt, B.A.; Mathalon, D.; McGlashan, T.H.; Perkins, D.O.; et al. Metabolic abnormalities and low dietary omega 3 are associated with symptom severity and worse functioning prior to the onset of psychosis: Findings from the north american prodrome longitudinal studies consortium. Schizophr. Res. 2019, 204, 96–103. [Google Scholar] [CrossRef] [PubMed]
- De Groot, R.H.M.; Emmett, R.; Meyer, B.J. Non-dietary factors associated with n-3 long-chain pufa levels in humans—A systematic literature review. Br. J. Nutr. 2019, 121, 793–808. [Google Scholar] [CrossRef] [Green Version]
- De Groot, R.H.M.; Meyer, B.J. Issfal official statement number 6 the importance of measuring blood omega-3 long chain polyunsaturated fatty acid levels in research. Prost. Leukot. Essent. Fat. Acids 2019. [Google Scholar] [CrossRef]
- Köhler, A.; Bittner, D.; Löw, A.; Schacky, C. Effects of a convenience drink fortified with n-3 fatty acids on the n-3 index. Br. J. Nutr. 2010, 104, 729–736. [Google Scholar] [CrossRef] [Green Version]
- Kohler, A.; Heinrich, J.; Von Schacky, C. Bioavailability of dietary omega-3 fatty acids added to a variety of sausages in healthy individuals. Nutrients 2017, 9, 629. [Google Scholar] [CrossRef] [Green Version]
- Kohler, A.; Sarkkinen, E.; Tapola, N.; Niskanen, T.; Bruheim, I. Bioavailability of fatty acids from krill oil, krill meal and fish oil in healthy subjects--a randomized, single-dose, cross-over trial. Lipids Health Dis. 2015, 14, 19. [Google Scholar] [CrossRef] [Green Version]
- Flock, M.; Skulas-Ray, A.; Harris, W.; Etherton, T.; Fleming, J.; Kris-Etherton, P. Determinants of erythrocyte omega-3 fatty acid content in response to fish oil supplementation: A dose-response randomized controlled trial. J. Am. Heart Assoc. 2013, 2, e000513. [Google Scholar] [CrossRef] [Green Version]
- Sparkes, C.; Gibson, R.; Sinclair, A.; Else, P.L.; Meyer, B.J. Effect of low dose docosahexaenoic acid-rich fish oil on plasma lipids and lipoproteins in pre-menopausal women: A dose(-)response randomized placebo-controlled trial. Nutrients 2018, 10, 1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swierk, M.; Williams, P.G.; Wilcox, J.; Russell, K.G.; Meyer, B.J. Validation of an australian electronic food frequency questionnaire to measure polyunsaturated fatty acid intake. Nutrition 2011, 27, 641–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, B.L.; Williams, P.G.; Meyer, B.J. Biomarker validation of a long-chain omega-3 polyunsaturated fatty acid food frequency questionnaire. Lipids 2006, 41, 845–850. [Google Scholar] [CrossRef] [Green Version]
- Lepage, G.; Roy, C. Direct transesterificaton of all classes of lipids in one-step reaction. J. Lipid Res. 1986, 27, 114–120. [Google Scholar] [PubMed]
- Von Schacky, C.; Weber, P. Metabolism and effects on platelet function of the purified eicosapentaenoic and docosahexaenoic acids in humans. J. Clin. Investig. 1985, 76, 2446–2450. [Google Scholar] [CrossRef]
- Von Schacky, C.; Fahrer, C.; Fischer, S. Catabolism of leukotriene b5 in humans. J. Lipid Res. 1990, 31, 1831–1838. [Google Scholar]
- Arab, L. Biomarker of fat and fatty acid intakes. J. Nutr. 2003, 133, 925S–932S. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Ma, J.; Campos, H.; Hankinson, S.E.; Hu, F.B. Comparison between plasma and erythrocyte fatty acid content as biomarkers of fatty acid intake in us women. Am. J. Clin. Nutr. 2007, 86, 74–81. [Google Scholar] [CrossRef]
- Skeaff, C.M.; Hodson, L.; McKenzie, J.E. Dietary-induced changes in fatty acid composition of human plasma, platelet, and erythrocyte lipids follow a similar time course. J. Nutr. 2006, 136, 565–569. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.F.; Tong, W.F.; Ruan, Y.; Sinclair, A.J.; Li, D. Different metabolism of epa, dpa and dha in humans: A double blind corss-over study. Prost. Leukot. Essent. Fat. Acids 2019. [Google Scholar] [CrossRef]
- Katan, M.B.; Deslypere, J.P.; Van Birgelen, A.P.; Penders, M.M.Z. Kinetics of the incorporation of dietary fatty acids into serum cholesteryl esters, erythrocyte membranes, and adipose tissue: An 18-month controlled study. J. Lipid Res. 1997, 38, 2012–2022. [Google Scholar] [PubMed]
- Harris, W. The omega-3 index as a risk factor for coronary heart disease. Am. J. Clin. Nutr. 2008, 87, 1997S–2002S. [Google Scholar] [CrossRef] [PubMed]
- Ghasemifard, S.; Turchini, G.M.; Sinclair, A.J. Omega-3 long chain fatty acid "bioavailability": A review of evidence and methodological considerations. Prog. Lipid Res. 2014, 56, 92–108. [Google Scholar] [CrossRef] [PubMed]
- Dushianthan, A.; Cusack, R.; Koster, G.; Grocott, M.P.W.; Postie, A.D. Insight into erythrocyte phospholipid molecular flux in healthy humans and in patients with acute respiratory distress syndrome. PLoS ONE 2019, 14, e0221595. [Google Scholar] [CrossRef]
- Arduini, A.; Mancinelli, G.; Radatti, G.L.; Dottori, S.; Molajoni, F.; Ramsay, R.R. Role of carnitine and carnitine palmitoyltransferase as integral components of the pathway for membrane phospholipid fatty acid turnover in intact human erythrocytes. J. Biol. Chem. 1992, 267, 12673–12681. [Google Scholar]
- Keenan, A.H.; Pedersen, T.L.; Fillaus, K.; Larson, M.K.; Shearer, G.C.; Newman, J.W. Basal omega-3 fatty acid status affects fatty acid and oxylipin responses to high-dose n3-hufa in healthy volunteers. J. Lipid Res. 2012, 53, 1662–1669. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, I.; Sasaki, E.; Yasunami, H.; Nomiyama, S.; Nakayama, M.; Sugano, M.; Imaizumi, K.K.Y. Digestion and lymphatic transport of eicosapentaenoic acid and docosaheaenoic acids given in the form of triacylglycerol, free acid and ethyl esters in rats. Biochim. Biophys. Acta 1995, 1259, 297–304. [Google Scholar] [CrossRef]
- Nelson, G.J.; Ackman, R.G. Absorption and transport of fat in mammals with emphasis on n−3 polyunsaturated fatty acids. Lipids 1988, 23, 1005–1014. [Google Scholar] [CrossRef]
- Lawson, L.D.; Hughes, B.G. Absorption of eicosapentaenoic acid and docosahexaenoic acid from fish oil triacylglycerols or fish oil ethyl esters co-ingested with a high-fat meal. Biochem. Biophys. Res. Commun. 1988, 156, 960–963. [Google Scholar] [CrossRef]
- Rice, H.B.; Bernasconi, A.; Maki, K.C.; Harris, W.S.; Von Schacky, C.; Calder, P.C. Conducting omega-3 clinical trials with cardiovascular outcomes: Proceedings of a workshop held at issfal 2014. Prost. Leukot. Essent. Fat. Acids 2016, 107, 30–42. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Schwichtenberg, K.A.; Hanson, N.Q.; Tsai, M.Y. Incorporation and clearance of omega-3 fatty acids in erythrocyte membranes and plasma phospholipids. Clin. Chem. 2006, 52, 2265–2272. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Whole Group (n = 45) | 0 g/day (n = 11) | 0.35 g/day (n = 14) | 0.7 g/day (n = 12) | 1 g/day (n = 8) | p Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | ||
| Age (years) | 26.5 | 6.4 | 26.8 | 6.8 | 26.8 | 6.7 | 24.7 | 5.3 | 28.5 | 7.4 | 0.63 |
| SBP (mmHg) | 114 | 9.1 | 113 | 7.0 | 116 | 8.8 | 116 | 12.0 | 109 | 7.0 | 0.35 |
| DBP (mmHg) | 71 | 8.4 | 70 | 7.2 | 72 | 8.5 | 72 | 11.2 | 71 | 5.3 | 0.91 |
| BMI | 25 | 5.1 | 23 | 3.1 | 25 | 5.3 | 25 | 6.2 | 25 | 5.4 | 0.68 |
| Whole Group | Placebo Group | Doses of DHA | p Value | |||
|---|---|---|---|---|---|---|
| (n = 45) | 0 g/day (n = 11) | 0.35 g/day (n = 14) | 0.7 g/day (n = 12) | 1 g/day (n = 8) | ||
| Median (IQR) | Median (IQR) | Median (IQR) | Median (IQR) | Median (IQR) | ||
| Energy (kJ/day) | 6165 (4979, 8021) | 6412 (5173, 8029) | 5862 (4568, 7618) | 7109 (5245, 8969) | 5187 (3774, 6731) | 0.28 |
| Protein (g/day) | 79 (59, 96) | 86 (64, 101) | 72 (57, 95) | 88 (58, 112) | 64 (55, 87) | 0.50 |
| Carbohydrate (g/day) | 163 (134, 201) | 172 (156, 195) | 146 (117, 200) | 192 (141, 241) | 149 (102, 176) | 0.12 |
| Total fat (g/day) | 59 (44, 77) | 61 (44, 81) | 59 (45, 78) | 67 (45, 77) | 43 (31, 62) | 0.43 |
| SFA (g/day) | 24 (16, 28) | 24 (20, 28) | 24 (18, 31) | 24 (18, 32) | 15 (12, 24) | 0.38 |
| MUFA (g/day) | 22 (15, 27) | 23 (14, 33) | 21 (17, 28) | 25 (15, 27) | 16 (11, 23) | 0.47 |
| PUFA (g/day) # | 9.4 (7.4, 13.3) | 7.5 (5.3, 10.5) * | 13.3 (8.3, 18.4) | 9.6 (7.6, 11.3) | 8.8 (6.3, 13.4) | 0.01 |
| LA (g/day) # | 7.5 (6.3, 11.3) | 6.4 (4.8, 8.3) * | 11.2 (7.3, 15.8) | 7.9 (6.2, 9.3) | 6.91 (5.25, 11.1) | 0.01 |
| ALA (g/day) # | 0.86 (0.64, 1.44) | 0.77 (0.59, 0.86) | 1.00 (0.62, 1.73) | 0.91 (0.69, 1.39) | 0.84 (0.59, 1.43) | 0.35 |
| AA (g/day) # | 0.15 (0.12, 0.23) | 0.15 (0.09, 0.20) | 0.16 (0.13, 0.33) | 0.15 (0.09, 0.21) | 0.16 (0.11, 0.24) | 0.08 |
| EPA (mg/day) # | 120 (51, 178) | 80 (27, 122) | 123 (65, 195) | 90 (55, 212) | 153 (55, 207) | 0.60 |
| DPA (mg/day) # | 89 (54, 149) | 87 (41, 113) | 87 (58, 178) | 93 (52, 150) | 13 (56, 178) | 0.18 |
| DHA (mg/day) # | 117 (56, 242) | 117 (21, 186) | 110 (56, 249) | 136 (77, 237) | 145 (40, 392) | 0.28 |
| n-3 LCPUFA (mg/day) # | 338 (186, 546) | 255 (96, 425) | 368 (197, 600) | 279 (203, 628) | 445 (160, 700) | 0.21 |
| 0 g/day (n = 11) | 0.35 g/day (n = 14) | 0.7 g/day (n = 12) | 1 g/day (n = 8) | |||||
|---|---|---|---|---|---|---|---|---|
| 0 Weeks | 8 Weeks | 0 Weeks | 8 Weeks | 0 Weeks | 8 Weeks | 0 Weeks | 8 Weeks | |
| Erythrocyte | ||||||||
| EPA | 0.48 (0.40, 0.58) | 0.50 a (0.33, 0.71) | 0.52 (0.44, 0.72) | 0.54 a (0.43, 0.64) | 0.63 (0.59, 0.70) | 0.71 a (0.67, 0.81) | 0.89 (0.40, 1.19) | 1.14 b (0.76, 1.37) |
| DHA | 4.38 (3.57, 5.54) | 3.97 a (3.46, 4.95) | 4.12 (3.48, 4.78) | 4.94 a (4.48, 5.25) | 4.47 (4.03, 4.82) | 5.70 b (5.55, 6.24) | 4.80 (3.63, 5.64) | 6.27 b (5.93, 7.15) |
| EPA+DHA | 4.82 (3.97, 5.99) | 4.68 a (3.86, 5.57) | 4.74 (3.85, 5.32) | 5.44 a (4.98, 5.75) | 5.10 (4.62, 5.60) | 6.47 b (6.22, 6.87) | 5.69 (4.00, 6.83) | 7.59 c (6.70, 8.38) |
| Plasma | ||||||||
| EPA | 0.54 (0.49, 0.76) | 0.65 a (0.47, 0.85) | 0.71 (0.56, 0.81) | 0.71 a (0.57, 0.84) | 0.73 (0.62, 0.86) | 1.07 b (0.87, 1.33) | 0.82 (0.57, 1.44) | 1.64 c (1.10, 2.45) |
| DHA | 1.92 (1.33, 2.15) | 1.54 a (1.39, 1.91) | 1.82 (1.47, 2.00) | 2.32 a (1.98, 2.47) | 1.61 (1.43, 2.12) | 3.14 b (2.71, 3.41) | 2.24 (1.52, 2.70) | 3.76 c (3.18, 4.45) |
| EPA+DHA | 2.59 (1.88, 2.70) | 2.33 a (1.97, 2.76) | 2.52 (2.14, 2.74) | 3.06 a (2.79, 3.35) | 2.24 (2.10, 3.15) | 4.30 b (3.60, 4.88) | 2.94 (2.07, 4.13) | 5.48 c (4.34, 6.68) |
| Whole blood | ||||||||
| EPA | 0.59 (0.48, 0.68) | 0.63 a (0.47, 0.84) | 0.66 (0.50, 0.77) | 0.68 a (0.54, 0.89) | 0.72 (0.60, 0.85) | 0.90 a (0.74, 1.14) | 0.88 (0.59, 1.36) | 1.42 b (1.03, 2.17) |
| DHA | 2.78 (2.3, 3.2) | 2.37 a (2.07, 2.96) | 2.56 (2.16, 2.87) | 3.28 a,b (2.80, 3.41) | 2.57 (2.13, 2.84) | 4.05 b (3.61, 4.53) | 3.05 (2.10, 3.40) | 4.41 b,c (3.88, 5.56) |
| EPA+DHA | 3.42 (2.80, 3.87) | 3.04 a (2.71, 3.81) | 3.17 (2.86, 3.50) | 3.94 a,b (3.55, 4.17) | 3.32 (2.79, 3.57) | 4.79 b (4.52, 5.67) | 3.81 (2.78, 4.73) | 6.03 c (4.91, 7.21) |
| Dietary Intake of EPA Plus DHA (g/day) Baseline | Dietary Intake of EPA Plus DHA (g/day) Post-Intervention | |||
|---|---|---|---|---|
| Model 1 | R2 adjusted | p value | R2 adjusted | p value |
| EPA plus DHA in erythrocytes (% of total fatty acids) | 0.25 | 0.0003 | 0.48 | <0.0001 |
| EPA plus DHA in plasma (% of total fatty acids) | 0.21 | 0.0009 | 0.50 | <0.0001 |
| EPA plus DHA in whole blood (% of total fatty acids) | 0.20 | 0.0016 | 0.48 | <0.0001 |
| Model 2 (includes baseline BMI) | R2 adjusted | p value | R2 adjusted | p value |
| EPA plus DHA in erythrocytes (% of total fatty acids) | 0.28 | 0.0003 | 0.55 | <0.0001 |
| EPA plus DHA in plasma (% of total fatty acids) | 0.32 | <0.0001 | 0.58 | <0.0001 |
| EPA plus DHA in whole blood (% of total fatty acids) | 0.26 | 0.0010 | 0.58 | <0.0001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sparkes, C.; Sinclair, A.J.; Gibson, R.A.; Else, P.L.; Meyer, B.J. High Variability in Erythrocyte, Plasma and Whole Blood EPA and DHA Levels in Response to Supplementation. Nutrients 2020, 12, 1017. https://doi.org/10.3390/nu12041017
Sparkes C, Sinclair AJ, Gibson RA, Else PL, Meyer BJ. High Variability in Erythrocyte, Plasma and Whole Blood EPA and DHA Levels in Response to Supplementation. Nutrients. 2020; 12(4):1017. https://doi.org/10.3390/nu12041017
Chicago/Turabian StyleSparkes, Cassandra, Andrew J. Sinclair, Robert A. Gibson, Paul L. Else, and Barbara J. Meyer. 2020. "High Variability in Erythrocyte, Plasma and Whole Blood EPA and DHA Levels in Response to Supplementation" Nutrients 12, no. 4: 1017. https://doi.org/10.3390/nu12041017
APA StyleSparkes, C., Sinclair, A. J., Gibson, R. A., Else, P. L., & Meyer, B. J. (2020). High Variability in Erythrocyte, Plasma and Whole Blood EPA and DHA Levels in Response to Supplementation. Nutrients, 12(4), 1017. https://doi.org/10.3390/nu12041017