Antecedent Dietary Glutamine Supplementation Benefits Modulation of Liver Pyroptosis in Mice with Polymicrobial Sepsis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Study Procedures
2.3. Measurements of Plasma Biochemical Parameters
2.4. RNA Extraction and Quantitative Reverse-Transcription Polymerase Chain Reaction (RT-qPCR) of Liver Tissues
2.5. Western Blotting Analysis for Protein Expressions in the Liver
2.6. Histopathology of the Liver
2.7. Statistical Analysis
3. Results
3.1. Plasma Biochemical Markers and Inflammatory Cytokine Concentrations
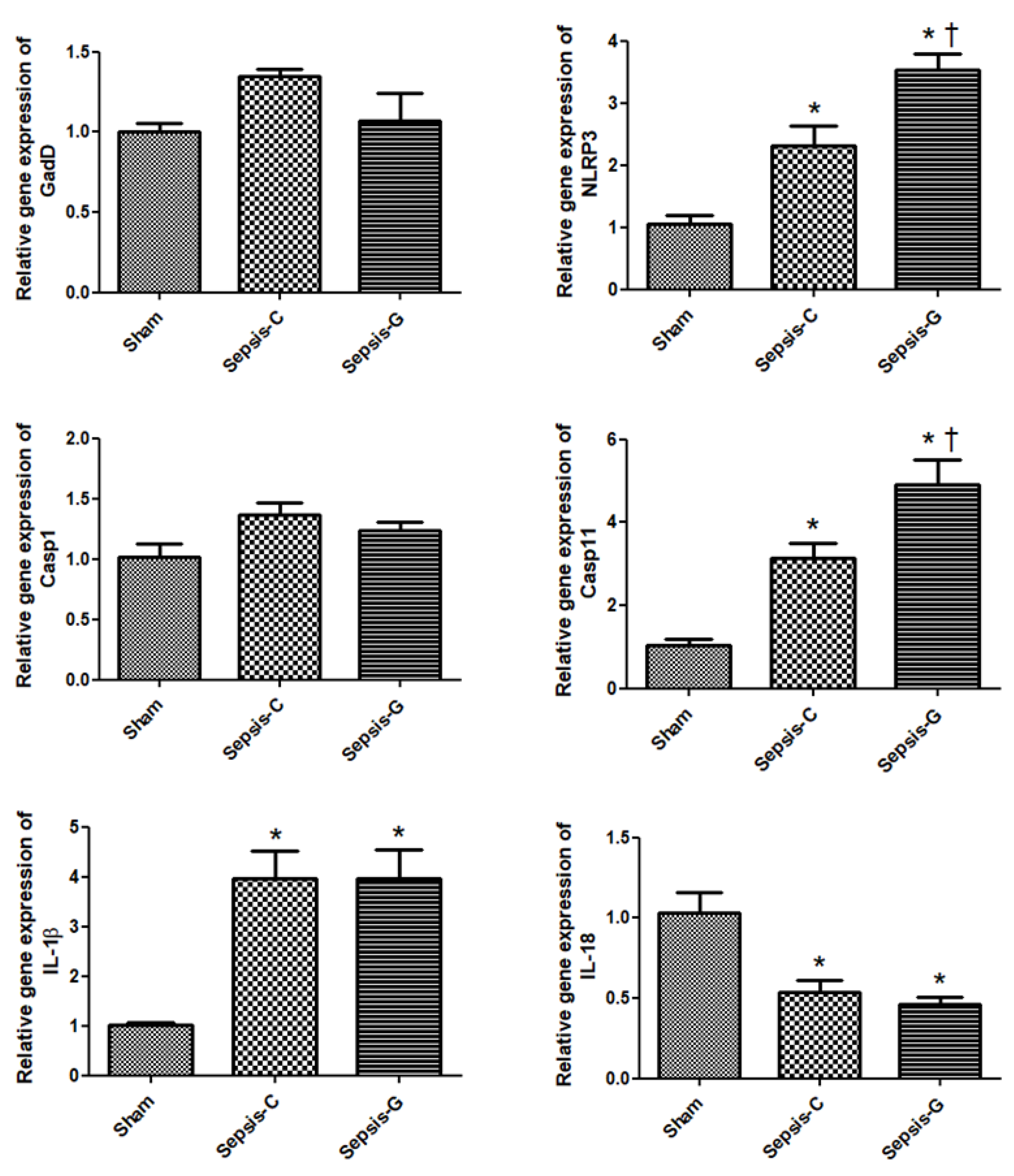
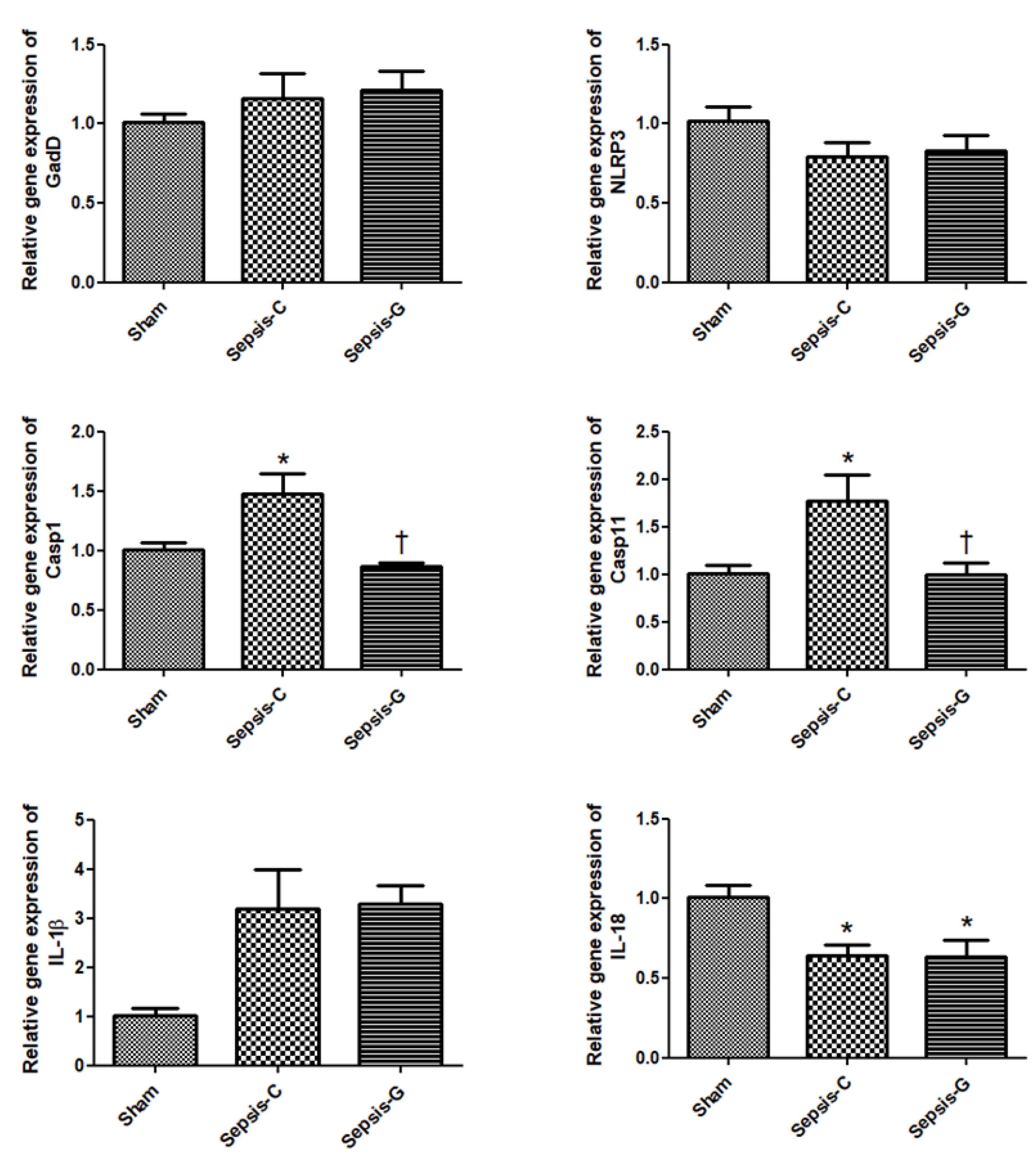
3.2. Pyroptosis-Related Gene Expressions in the Liver after CLP
3.3. Inflammatory-Related Gene Expressions in the Liver after CLP
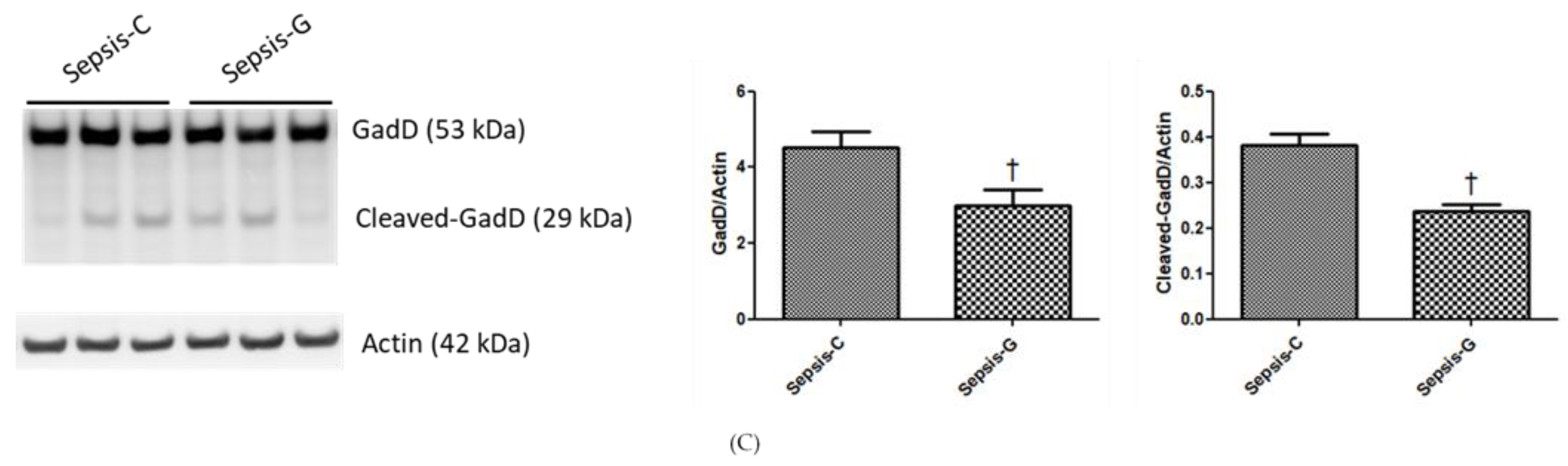
3.4. Protein Expression Levels of Caspase-1/11 and GadD in the Liver at 72 h after CLP
3.5. Histopathological Aspects of the Liver
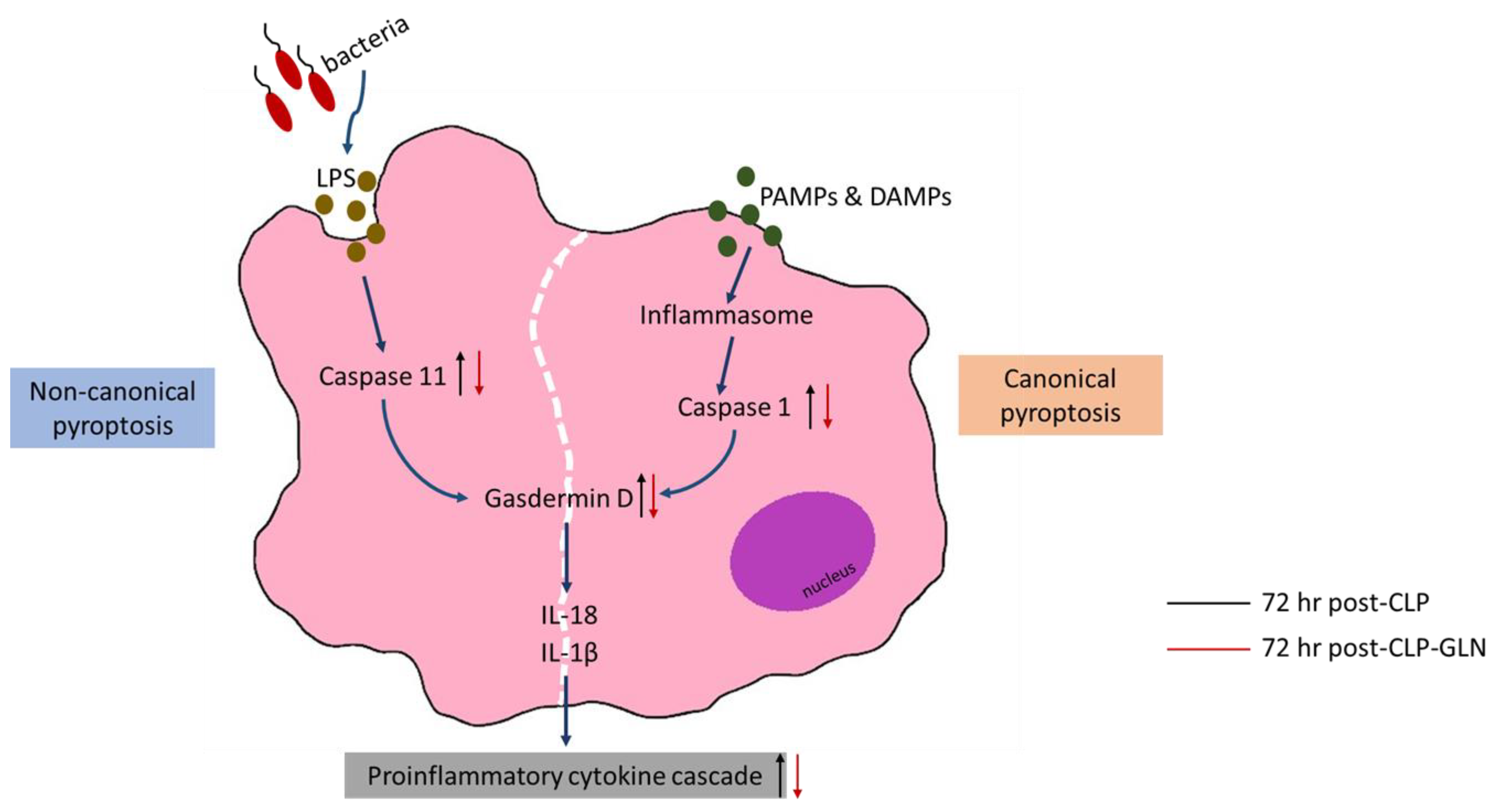
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The third international consensus definitions for sepsis and septic shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.; Vincent, J.L.; Adhikari, N.K.; Machado, F.R.; Angus, D.C.; Calandra, T.; Jaton, K.; Giulieri, S.; Delaloye, J.; Opal, S.; et al. Sepsis: A roadmap for future research. Lancet Infect. Dis. 2015, 15, 581–614. [Google Scholar] [CrossRef]
- Trefts, E.; Gannon, M.; Wasserman, D.H. The liver. Curr. Biol. 2017, 27, R1147–R1151. [Google Scholar] [CrossRef] [PubMed]
- Heymann, F.; Tacke, F. Immunology in the liver—From homeostasis to disease. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 88–110. [Google Scholar] [CrossRef]
- Yan, J.; Li, S.; Li, S. The role of the liver in sepsis. Int. Rev. Immunol. 2014, 33, 498–510. [Google Scholar] [CrossRef] [Green Version]
- Mira, J.C.; Gentile, L.F.; Mathias, B.J.; Efron, P.A.; Brakenridge, S.C.; Mohr, A.M.; Moore, F.A.; Moldawer, L.L. Sepsis pathophysiology, chronic critical illness, and persistent inflammation-immunosuppression and catabolism syndrome. Crit. Care Med. 2017, 45, 253–262. [Google Scholar] [CrossRef]
- Kramer, L.; Jordan, B.; Druml, W.; Bauer, P.; Metnitz, P.G. Incidence and prognosis of early hepatic dysfunction in critically ill patients—A prospective multicenter study. Crit. Care Med. 2007, 35, 1099–1104. [Google Scholar] [CrossRef]
- Canabal, J.M.; Kramer, D.J. Management of sepsis in patients with liver failure. Curr. Opin. Crit. Care 2008, 14, 189–197. [Google Scholar] [CrossRef]
- Wu, J.; Lin, S.; Wan, B.; Velani, B.; Zhu, Y. Pyroptosis in liver disease: New insights into disease mechanisms. Aging Dis. 2019, 10, 1094–1108. [Google Scholar] [CrossRef] [Green Version]
- Vande Walle, L.; Lamkanfi, M. Pyroptosis. Curr. Biol. 2016, 26, R568–R572. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.L.; Xu, G.; Liang, X.; Wei, J.; Luo, J.; Chen, G.N.; Yan, X.D.; Wen, X.P.; Zhong, M.; Lv, X. Inhibition of hepatic cells pyroptosis attenuates CLP-induced acute liver injury. Am. J. Transl. Res. 2016, 8, 5685–5695. [Google Scholar] [PubMed]
- Fink, S.L.; Cookson, B.T. Caspase-1-dependent pore formation during pyroptosis leads to osmotic lysis of infected host macrophages. Cell Microbiol. 2006, 8, 1812–1825. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.S. Caspase-11 non-canonical inflammasome: A critical sensor of intracellular lipopolysaccharide in macrophage-mediated inflammatory responses. Immunology 2017, 152, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Man, S.M.; Karki, R.; Kanneganti, T.D. Molecular mechanisms and functions of pyroptosis, inflammatory caspases and inflammasomes in infectious diseases. Immunol. Rev. 2017, 277, 61–75. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, A.; Sohail, M.A.; Gomes, D.A.; Hashmi, A.; Nagata, J.; Sutterwala, F.S.; Mahmood, S.; Jhandier, M.N.; Shi, Y.; Flavell, R.A.; et al. Inflammasome-mediated regulation of hepatic stellate cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1248–G1257. [Google Scholar] [CrossRef] [Green Version]
- Dixon, L.J.; Berk, M.; Thapaliya, S.; Papouchado, B.G.; Feldstein, A.E. Caspase-1-mediated regulation of fibrogenesis in diet-induced steatohepatitis. Lab. Invest. 2012, 92, 713–723. [Google Scholar] [CrossRef]
- Yan, W.; Chang, Y.; Liang, X.; Cardinal, J.S.; Huang, H.; Thorne, S.H.; Monga, S.P.; Geller, D.A.; Lotze, M.T.; Tsung, A. High-mobility group box 1 activates caspase-1 and promotes hepatocellular carcinoma invasiveness and metastases. Hepatology 2012, 55, 1863–1875. [Google Scholar] [CrossRef]
- Wilmore, D.W. The effect of glutamine supplementation in patients following elective surgery and accidental injury. J. Nutr. 2001, 131, 2543S–2549S. [Google Scholar] [CrossRef]
- Wischmeyer, P.E. Glutamine: Role in critical illness and ongoing clinical trials. Curr. Opin. Gastroenterol. 2008, 24, 190–197. [Google Scholar] [CrossRef]
- Kessel, A.; Toubi, E.; Pavlotzky, E.; Mogilner, J.; Coran, A.G.; Lurie, M.; Karry, R.; Sukhotnik, I. Treatment with glutamine is associated with down-regulation of Toll-like receptor-4 and myeloid differentiation factor 88 expression and decrease in intestinal mucosal injury caused by lipopolysaccharide endotoxaemia in a rat. Clin. Exp. Immunol. 2008, 151, 341–347. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, G.P.; Oliveira, M.B.; Santos, R.S.; Lima, L.D.; Dias, C.M.; Ab’ Saber, A.M.; Teodoro, W.R.; Capelozzi, V.L.; Gomes, R.N.; Bozza, P.T.; et al. Intravenous glutamine decreases lung and distal organ injury in an experimental model of abdominal sepsis. Crit. Care 2009, 13, R74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.M.; Hsiung, Y.C.; Pai, M.H.; Yeh, S.L. Glutamine administration in early or late septic phase downregulates lymphocyte PD-1/PD-L1 expression and the inflammatory response in mice with polymicrobial sepsis. J. Parenter. Enteral. Nutr. 2018, 42, 538–549. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.C.; Wu, J.M.; Chen, K.Y.; Chen, P.D.; Lei, C.S.; Yeh, S.L.; Lin, M.T. Effects of prophylactic administration of glutamine on CD4(+) T cell polarisation and kidney injury in mice with polymicrobial sepsis. Br. J. Nutr. 2019, 122, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.S.; Wu, J.M.; Lee, P.C.; Kuo, T.C.; Chen, P.D.; Hou, Y.C.; Yeh, S.L.; Lin, M.T. Antecedent administration of glutamine benefits the homeostasis of CD4(+) T Cells and attenuates lung injury in mice with gut-derived polymicrobial sepsis. J. Parenter. Enteral. Nutr. 2019, 43, 927–936. [Google Scholar] [CrossRef]
- Wischmeyer, P.E.; Kahana, M.; Wolfson, R.; Ren, H.; Musch, M.M.; Chang, E.B. Glutamine reduces cytokine release, organ damage, and mortality in a rat model of endotoxemia. Shock 2001, 16, 398–402. [Google Scholar] [CrossRef]
- Hubbard, W.J.; Choudhry, M.; Schwacha, M.G.; Kerby, J.D.; Rue, L.W., III; Bland, K.I.; Chaudry, I.H. Cecal ligation and puncture. Shock 2005, 24, 52–57. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Halbach, J.L.; Wang, A.W.; Hawisher, D.; Cauvi, D.M.; Lizardo, R.E.; Rosas, J.; Reyes, T.; Escobedo, O.; Bickler, S.W.; Coimbra, R.; et al. Why antibiotic treatment is not enough for sepsis resolution: An evaluation in an experimental animal model. Infect. Immun. 2017, 85, e00664–e00717. [Google Scholar] [CrossRef] [Green Version]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]
- Howison, J.; Bullard, J. Software in the scientific literature: Problems with seeing, finding, and using software mentioned in the biology literature. J. Assoc. Inform. Sci. Technol. 2016, 67, 2137–2155. [Google Scholar] [CrossRef]
- Jenne, C.N.; Kubes, P. Immune surveillance by the liver. Nat. Immunol. 2013, 14, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Strnad, P.; Tacke, F.; Koch, A.; Trautwein, C. Liver—Guardian, modifier and target of sepsis. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, J.; Nagata, T.; Uchijima, M.; Shibata, K.; Koide, Y. Intracytosolic Listeria monocytogenes induces cell death through caspase-1 activation in murine macrophages. Cell Microbiol. 2008, 10, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Sozen, T.; Hasegawa, Y.; Chen, W.; Zhang, J.H. Caspase-1 inhibitor prevents neurogenic pulmonary edema after subarachnoid hemorrhage in mice. Stroke 2009, 40, 3872–3875. [Google Scholar] [CrossRef] [Green Version]
- Duewell, P.; Kono, H.; Rayner, K.J.; Sirois, C.M.; Vladimer, G.; Bauernfeind, F.G.; Abela, G.S.; Franchi, L.; Nuñez, G.; Schnurr, M.; et al. NLRP3 inflammasomes are required for atherogenesis and activated by cholesterol crystals. Nature 2010, 464, 1357–1361. [Google Scholar] [CrossRef] [Green Version]
- Kayagaki, N.; Stowe, I.B.; Lee, B.L.; O’Rourke, K.; Anderson, K.; Warming, S.; Cuellar, T.; Haley, B.; Roose-Girma, M.; Phung, Q.T.; et al. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature 2015, 526, 666–671. [Google Scholar] [CrossRef]
- Couper, K.N.; Blount, D.G.; Riley, E.M. IL-10: The master regulator of immunity to infection. J. Immunol. 2008, 180, 5771–5777. [Google Scholar] [CrossRef]
- Dinarello, C.A. Interleukin-18. Methods 1999, 19, 121–132. [Google Scholar] [CrossRef]
- Heyland, D.; Muscedere, J.; Wischmeyer, P.E.; Cook, D.; Jones, G.; Albert, M.; Elke, G.; Berger, M.M.; Day, A.G. A randomized trial of glutamine and antioxidants in critically ill patients. N. Engl. J. Med. 2013, 368, 1489–1497. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, T.R.; May, A.K.; Hebbar, G.; Easley, K.A.; Griffith, D.P.; Dave, N.; Collier, B.R.; Cotsonis, G.A.; Hao, L.; Leong, T.; et al. Efficacy and safety of glutamine-supplemented parenteral nutrition in surgical ICU patients: An American multicenter randomized controlled trial. Ann. Surg. 2016, 263, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Wischmeyer, P.E.; Dhaliwal, R.; McCall, M.; Ziegler, T.R.; Heyland, D.K. Parenteral glutamine supplementation in critical illness: A systematic review. Crit. Care 2014, 18, R76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singleton, K.D.; Beckey, V.E.; Wischmeyer, P.E. Glutamine prevents activation of NF-kappaB and stress kinase pathways, attenuates inflammatory cytokine release, and prevents acute respiratory distress syndrome (ARDS) following sepsis. Shock 2005, 24, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Coeffier, M.; Marion, R.; Ducrotte, P.; Dechelotte, P. Modulating effect of glutamine on IL-1beta-induced cytokine production by human gut. Clin. Nutr. 2003, 22, 407–413. [Google Scholar] [CrossRef]
- Tschopp, J.; Schroder, K. NLRP3 inflammasome activation: The convergence of multiple signalling pathways on ROS production? Nat. Rev. Immunol. 2010, 10, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Roth, E.; Oehler, R.; Manhart, N.; Exner, R.; Wessner, B.; Strasser, E.; Spittler, A. Regulative potential of glutamine--relation to glutathione metabolism. Nutrition 2002, 18, 217–221. [Google Scholar] [CrossRef]
- Tsai, P.H.; Liu, J.J.; Yeh, C.L.; Chiu, W.C.; Yeh, S.L. Effects of glutamine supplementation on oxidative stress-related gene expression and antioxidant properties in rats with streptozotocin-induced type 2 diabetes. Br. J. Nutr. 2012, 107, 1112–1118. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Control Diet | Glutamine Diet |
|---|---|---|
| Soybean oil | 70 | 70 |
| Casein | 200 | 150 |
| Glutamine | 0 | 41.7 |
| Sucrose | 100 | 100 |
| Mineral mixture † | 35 | 35 |
| Vitamin mixture ‡ | 10 | 10 |
| Fiber | 50 | 50 |
| Choline bitartrate | 2.5 | 2.5 |
| L-Cysteine | 3 | 3 |
| Corn starch | 529.5 | 537.8 |
| 24 h Post-CLP | 72 h Post-CLP | |||||
|---|---|---|---|---|---|---|
| Sham | Sepsis-C | Sepsis-G | Sham | Sepsis-C | Sepsis-G | |
| ALT (U/L) | 6.04 ± 2.15 | 10.08 ± 1.08 | 25.18 ± 8.13 * | 1.47 ± 0.20 | 9.20 ± 3.80 | 2.48 ± 0.42 |
| AST (U/L) | 31.45 ± 9.15 | 72.67 ± 4.12 * | 72.33 ± 11.23 * | 17.38 ± 4.82 | 49.69 ± 10.66 * | 25.96 ± 2.70 |
| IL-1β (pg/mL) | 5.10 ± 0.75 | 7.77 ± 1.58 | 12.53 ± 1.86 * | 7.18 ± 1.27 | 10.67 ± 1.35 | 8.92 ± 1.55 |
| IL-18 (pg/mL) | 14.42 ± 3.83 | 46.70 ± 6.71 * | 38.87 ± 8.04 * | 23.97 ± 6.38 | 20.91 ± 3.58 | 24.60 ± 4.56 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pai, M.-H.; Wu, J.-M.; Yang, P.-J.; Lee, P.-C.; Huang, C.-C.; Yeh, S.-L.; Lin, M.-T. Antecedent Dietary Glutamine Supplementation Benefits Modulation of Liver Pyroptosis in Mice with Polymicrobial Sepsis. Nutrients 2020, 12, 1086. https://doi.org/10.3390/nu12041086
Pai M-H, Wu J-M, Yang P-J, Lee P-C, Huang C-C, Yeh S-L, Lin M-T. Antecedent Dietary Glutamine Supplementation Benefits Modulation of Liver Pyroptosis in Mice with Polymicrobial Sepsis. Nutrients. 2020; 12(4):1086. https://doi.org/10.3390/nu12041086
Chicago/Turabian StylePai, Man-Hui, Jin-Ming Wu, Po-Jen Yang, Po-Chu Lee, Chun-Chieh Huang, Sung-Ling Yeh, and Ming-Tsan Lin. 2020. "Antecedent Dietary Glutamine Supplementation Benefits Modulation of Liver Pyroptosis in Mice with Polymicrobial Sepsis" Nutrients 12, no. 4: 1086. https://doi.org/10.3390/nu12041086
APA StylePai, M. -H., Wu, J. -M., Yang, P. -J., Lee, P. -C., Huang, C. -C., Yeh, S. -L., & Lin, M. -T. (2020). Antecedent Dietary Glutamine Supplementation Benefits Modulation of Liver Pyroptosis in Mice with Polymicrobial Sepsis. Nutrients, 12(4), 1086. https://doi.org/10.3390/nu12041086