Plausible Biological Interactions of Low- and Non-Calorie Sweeteners with the Intestinal Microbiota: An Update of Recent Studies

 ,
,  ,
,  and
and 
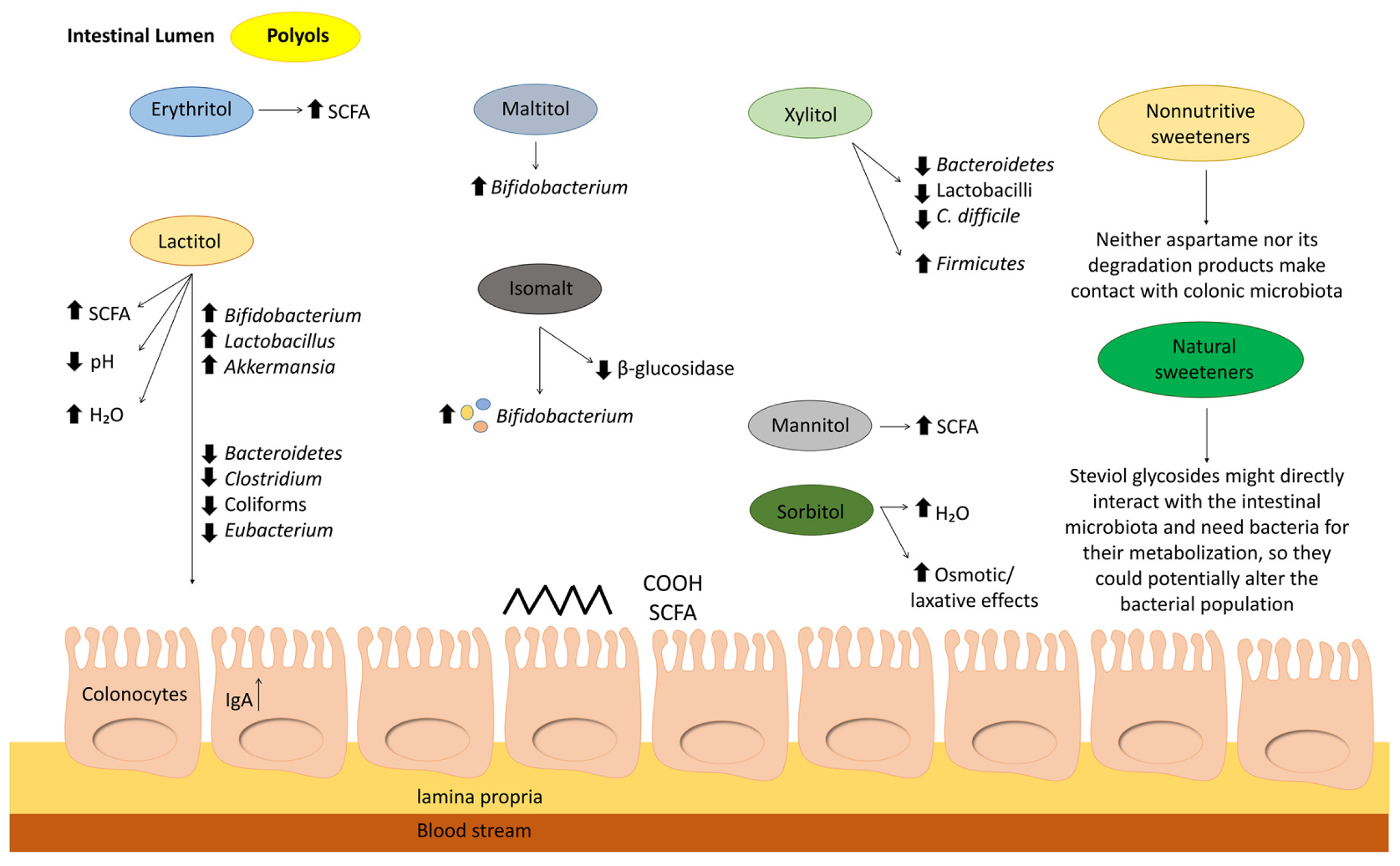{kind=link}
Abstract
:1. Introduction
2. Biological Plausibility: Which Low- and Non-Calorie Sweetener (LNCS) Could Potentially Affect the Colonic Microbiota?
2.1. Effects of Non-Nutritive Low-Calorie Sweeteners on the Gut Microbiota
2.1.1. Aspartame
2.1.2. Potassium Acesulfame (Ace-K)
2.1.3. Cyclamate
2.1.4. Sucralose
2.1.5. Saccharin
2.1.6. Steviol Glycosides
2.2. Effects of Nutritive Low-Calorie Sweeteners on the Gut Microbiota
2.2.1. Polyols
2.2.2. Erythritol
2.2.3. Isomalt
2.2.4. Lactitol
2.2.5. Maltitol
2.2.6. Sorbitol
2.2.7. Mannitol
2.2.8. Xylitol
3. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ruiz-Ojeda, F.J.; Plaza-Diaz, J.; Saez-Lara, M.J.; Gil, A. Effects of Sweeteners on the Gut Microbiota: A Review of Experimental Studies and Clinical Trials. Adv. Nutr. 2019, 10, S31–S48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohner, S.; Toews, I.; Meerpohl, J.J. Health outcomes of non-nutritive sweeteners: Analysis of the research landscape. Nutr. J. 2017, 16, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanhope, K.L. Sugar consumption, metabolic disease and obesity: The state of the controversy. Crit. Rev. Clin. Lab. Sci. 2016, 53, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Suez, J.; Korem, T.; Zilberman-Schapira, G.; Segal, E.; Elinav, E. Non-caloric artificial sweeteners and the microbiome: Findings and challenges. Gut Microbes 2015, 6, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Frankenfeld, C.L.; Sikaroodi, M.; Lamb, E.; Shoemaker, S.; Gillevet, P.M. High-intensity sweetener consumption and gut microbiome content and predicted gene function in a cross-sectional study of adults in the United States. Ann. Epidemiol. 2015, 25, 736–742. [Google Scholar] [CrossRef] [PubMed]
- Fitriakusumah, Y.; Lesmana, C.R.A.; Bastian, W.P.; Jasirwan, C.O.M.; Hasan, I.; Simadibrata, M.; Kurniawan, J.; Sulaiman, A.S.; Gani, R.A. The role of Small Intestinal Bacterial Overgrowth (SIBO) in Non-alcoholic Fatty Liver Disease (NAFLD) patients evaluated using Controlled Attenuation Parameter (CAP) Transient Elastography (TE): A tertiary referral center experience. BMC Gastroenterol. 2019, 19, 43. [Google Scholar] [CrossRef]
- Food Standards Agency, U.K. Food Additives. Different Food Additives and Advice on Regulations and the Safety of Additives in Food. Available online: https://www.food.gov.uk/safety-hygiene/food-additives (accessed on 24 December 2019).
- FDA. High-Intensity Sweeteners. Available online: https://www.fda.gov/food/food-additives-petitions/high-intensity-sweeteners (accessed on 24 December 2019).
- Hill, A.B. The Environment and Disease: Association or Causation? Proc. R. Soc. Med. 1965, 58, 295–300. [Google Scholar] [PubMed]
- Magnuson, B.A.; Carakostas, M.C.; Moore, N.H.; Poulos, S.P.; Renwick, A.G. Biological fate of low-calorie sweeteners. Nutr. Rev. 2016, 74, 670–689. [Google Scholar] [CrossRef] [Green Version]
- Butchko, H.H.; Stargel, W.W.; Comer, C.P.; Mayhew, D.A.; Benninger, C.; Blackburn, G.L.; de Sonneville, L.M.; Geha, R.S.; Hertelendy, Z.; Koestner, A.; et al. Aspartame: Review of safety. Regul. Toxicol. Pharmacol. 2002, 35, S1–S93. [Google Scholar]
- Stegink, L.D.; Wolf-Novak, L.C.; Filer, L.J., Jr.; Bell, E.F.; Ziegler, E.E.; Krause, W.L.; Brummel, M.C. Aspartame-sweetened beverage: Effect on plasma amino acid concentrations in normal adults and adults heterozygous for phenylketonuria. J. Nutr. 1987, 117, 1989–1995. [Google Scholar] [CrossRef] [Green Version]
- Sanchez de Medina Contreras, F.; Suárez Ortega, M.D. Tratado de Nutrición; Metabolismo de los aminoácidos; Editorial Médica Panamericana: Madrid, Spain, 2017; Chapter 8; pp. 173–195. [Google Scholar]
- Nettleton, J.E.; Cho, N.A.; Klancic, T.; Nicolucci, A.C.; Shearer, J.; Borgland, S.L.; Johnston, L.A.; Ramay, H.R.; Noye Tuplin, E.; Chleilat, F.; et al. Maternal low-dose aspartame and stevia consumption with an obesogenic diet alters metabolism, gut microbiota and mesolimbic reward system in rat dams and their offspring. Gut 2020. [Google Scholar] [CrossRef] [Green Version]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobach, A.R.; Roberts, A.; Rowland, I.R. Assessing the in vivo data on low/no-calorie sweeteners and the gut microbiota. Food Chem. Toxicol. 2019, 124, 385–399. [Google Scholar] [CrossRef]
- Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, R.; Shehreen, S.; Shahriar, S.; Rahman, M.S.; Akhteruzzaman, S.; Sajib, A.A. Non-Caloric Artificial Sweeteners Modulate the Expression of Key Metabolic Genes in the Omnipresent Gut Microbe Escherichia coli. J. Mol. Microbiol. Biotechnol. 2019, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Gerasimidis, K.; Bryden, K.; Chen, X.; Papachristou, E.; Verney, A.; Roig, M.; Hansen, R.; Nichols, B.; Papadopoulou, R.; Parrett, A. The impact of food additives, artificial sweeteners and domestic hygiene products on the human gut microbiome and its fibre fermentation capacity. Eur. J. Nutr. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renwick, A.G. The metabolism of intense sweeteners. Xenobiotica Fate Foreign Compd. Biol. Syst. 1986, 16, 1057–1071. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, M.; Ziesenitz, S.C.; Siebert, G. Acesulfame K, cyclamate and saccharin inhibit the anaerobic fermentation of glucose by intestinal bacteria. Z. Ernahr. 1985, 24, 231–235. [Google Scholar] [CrossRef]
- Bian, X.; Chi, L.; Gao, B.; Tu, P.; Ru, H.; Lu, K. The artificial sweetener acesulfame potassium affects the gut microbiome and body weight gain in CD-1 mice. PLoS ONE 2017, 12, e0178426. [Google Scholar] [CrossRef]
- Bueno-Hernández, N.; Vázquez-Frías, R.; Abreu, A.; Almeda-Valdés, P.; Barajas-Nava, L.; Carmona-Sánchez, R.; Chávez-Sáenz, J.; Consuelo-Sánchez, A.; Espinosa-Flores, A.; Hernández-Rosiles, V. Revisión de la evidencia científica y opinión técnica sobre el consumo de edulcorantes no calóricos en enfermedades gastrointestinales. Rev. Gastroenterol. Méx. 2019, 84, 492–510. [Google Scholar]
- Uebanso, T.; Ohnishi, A.; Kitayama, R.; Yoshimoto, A.; Nakahashi, M.; Shimohata, T.; Mawatari, K.; Takahashi, A. Effects of Low-Dose Non-Caloric Sweetener Consumption on Gut Microbiota in Mice. Nutrients 2017, 9, 560. [Google Scholar] [CrossRef] [Green Version]
- Hanawa, Y.; Higashiyama, M.; Sugihara, N.; Wada, A.; Inaba, K.; Horiuchi, K.; Furuhashi, H.; Kurihara, C.; Okada, Y.; Shibuya, N. Su1775–Artificial Sweetener Acesulfame Potassium Enhanced Lymphocyte Migration to Intestinal Microvessels by Enhancing Expression of Adhesion Molecules Through Dysbiosis. Gastroenterology 2019, 156, S-606. [Google Scholar] [CrossRef]
- Beards, E.; Tuohy, K.; Gibson, G. A human volunteer study to assess the impact of confectionery sweeteners on the gut microbiota composition. Br. J. Nutr. 2010, 104, 701–708. [Google Scholar] [CrossRef] [Green Version]
- Stichelen, O.-V.; Rother, K.I.; Hanover, J.A. Maternal exposure to non-nutritive sweeteners impacts progeny’s metabolism and microbiome. Front. Microbiol. 2019, 10, 1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.P.; Browman, D.; Herzog, H.; Neely, G.G. Non-nutritive sweeteners possess a bacteriostatic effect and alter gut microbiota in mice. PLoS ONE 2018, 13, e0199080. [Google Scholar] [CrossRef] [PubMed]
- Price, J.M.; Biava, C.G.; Oser, B.L.; Vogin, E.E.; Steinfeld, J.; Ley, H.L. Bladder tumors in rats fed cyclohexylamine or high doses of a mixture of cyclamate and saccharin. Science 1970, 167, 1131–1132. [Google Scholar] [CrossRef] [PubMed]
- Vamanu, E.; Pelinescu, D.; Gatea, F.; Sarbu, I. Altered in Vitro Metabolomic Response of the Human Microbiota to Sweeteners. Genes 2019, 10, 535. [Google Scholar] [CrossRef] [Green Version]
- Farup, P.G.; Lydersen, S.; Valeur, J. Are Nonnutritive Sweeteners Obesogenic? Associations between Diet, Faecal Microbiota, and Short-Chain Fatty Acids in Morbidly Obese Subjects. J. Obes. 2019, 2019, 4608315. [Google Scholar] [CrossRef]
- Hills, R.D., Jr.; Pontefract, B.A.; Mishcon, H.R.; Black, C.A.; Sutton, S.C.; Theberge, C.R. Gut Microbiome: Profound Implications for Diet and Disease. Nutrients 2019, 11, 1613. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.; Renwick, A.G.; Sims, J.; Snodin, D.J. Sucralose metabolism and pharmacokinetics in man. Food Chem. Toxicol. 2000, 38 (Suppl. 2), S31–S41. [Google Scholar] [CrossRef]
- Wang, X.; Guo, J.; Liu, Y.; Yu, H.; Qin, X. Sucralose Increased Susceptibility to Colitis in Rats. Inflamm. Bowel Dis. 2019, 25, e3–e4. [Google Scholar] [CrossRef]
- Bian, X.; Chi, L.; Gao, B.; Tu, P.; Ru, H.; Lu, K. Gut Microbiome Response to Sucralose and Its Potential Role in Inducing Liver Inflammation in Mice. Front. Physiol. 2017, 8, 487. [Google Scholar] [CrossRef] [Green Version]
- Thomson, P.; Santibanez, R.; Aguirre, C.; Galgani, J.E.; Garrido, D. Short-term impact of sucralose consumption on the metabolic response and gut microbiome of healthy adults. Br. J. Nutr. 2019, 122, 856–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golonka, R.M.; San Yeoh, B.; Vijay-Kumar, M. Dietary Additives and Supplements Revisited: The Fewer, the Safer for Gut and Liver Health. Curr. Pharmacol. Rep. 2019, 5, 303–316. [Google Scholar] [CrossRef]
- Rodriguez-Palacios, A.; Harding, A.; Menghini, P.; Himmelman, C.; Retuerto, M.; Nickerson, K.P.; Lam, M.; Croniger, C.M.; McLean, M.H.; Durum, S.K.; et al. The Artificial Sweetener Splenda Promotes Gut Proteobacteria, Dysbiosis, and Myeloperoxidase Reactivity in Crohn’s Disease-Like Ileitis. Inflamm. Bowel Dis. 2018, 24, 1005–1020. [Google Scholar] [CrossRef]
- Grotz, V.L.; Henry, R.R.; McGill, J.B.; Prince, M.J.; Shamoon, H.; Trout, J.R.; Pi-Sunyer, F.X. Lack of effect of sucralose on glucose homeostasis in subjects with type 2 diabetes. J. Am. Diet. Assoc. 2003, 103, 1607–1612. [Google Scholar] [CrossRef] [PubMed]
- Grotz, V.L.; Pi-Sunyer, X.; Porte, D., Jr.; Roberts, A.; Trout, J.R. A 12-week randomized clinical trial investigating the potential for sucralose to affect glucose homeostasis. Regul. Toxicol. Pharmacol. 2017, 88, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Byard, J.; McChesney, E.; Golberg, L.; Coulston, F. Excretion and metabolism of saccharin in man. II. Studies with 14C-labelled and unlabelled saccharin. Food Cosmet. Toxicol. 1974, 12, 175–184. [Google Scholar] [CrossRef]
- Renwick, A.G. The disposition of saccharin in animals and man—A review. Food Chem. Toxicol. 1985, 23, 429–435. [Google Scholar] [CrossRef]
- Bilan, M.; Lieshchova, M.; Tishkina, N.; Brygadyrenko, V. Combined effect of glyphosate, saccharin and sodium benzoate on the gut microbiota of rats. Regul. Mech. Biosyst. 2019, 10, 228–232. [Google Scholar] [CrossRef]
- Santos, P.; Caria, C.; Gotardo, E.; Ribeiro, M.; Pedrazzoli, J.; Gambero, A. Artificial sweetener saccharin disrupts intestinal epithelial cells’ barrier function in vitro. Food Funct. 2018, 9, 3815–3822. [Google Scholar] [CrossRef]
- Nogueira, J.P.S.; He, F.; Mangian, H.F.; Oba, P.M.; De Godoy, M.R.C. Dietary supplementation of a fiber-prebiotic and saccharin-eugenol blend in extruded diets fed to dogs. J. Anim. Sci 2019, 97, 4519–4531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutapea, A.M.; Toskulkao, C.; Buddhasukh, D.; Wilairat, P.; Glinsukon, T. Digestion of stevioside, a natural sweetener, by various digestive enzymes. J. Clin. Biochem. Nutr. 1997, 23, 177–186. [Google Scholar] [CrossRef]
- Koyama, E.; Kitazawa, K.; Ohori, Y.; Izawa, O.; Kakegawa, K.; Fujino, A.; Ui, M. In Vitro metabolism of the glycosidic sweeteners, stevia mixture and enzymatically modified stevia in human intestinal microflora. Food Chem. Toxicol. 2003, 41, 359–374. [Google Scholar] [CrossRef]
- Gardana, C.; Simonetti, P.; Canzi, E.; Zanchi, R.; Pietta, P. Metabolism of stevioside and rebaudioside A from Stevia rebaudiana extracts by human microflora. J. Agric. Food Chem. 2003, 51, 6618–6622. [Google Scholar] [CrossRef]
- Carakostas, M.C.; Curry, L.L.; Boileau, A.C.; Brusick, D.J. Overview: The history, technical function and safety of rebaudioside A, a naturally occurring steviol glycoside, for use in food and beverages. Food Chem. Toxicol. 2008, 46 (Suppl. 7), S1–S10. [Google Scholar] [CrossRef]
- Renwick, A.G.; Tarka, S.M. Microbial hydrolysis of steviol glycosides. Food Chem. Toxicol. 2008, 46 (Suppl. 7), S70–S74. [Google Scholar] [CrossRef]
- Geuns, J.M.; Buyse, J.; Vankeirsbilck, A.; Temme, E.H.; Compernolle, F.; Toppet, S. Identification of steviol glucuronide in human urine. J. Agric. Food Chem. 2006, 54, 2794–2798. [Google Scholar] [CrossRef]
- Wheeler, A.; Boileau, A.C.; Winkler, P.C.; Compton, J.C.; Prakash, I.; Jiang, X.; Mandarino, D.A. Pharmacokinetics of rebaudioside A and stevioside after single oral doses in healthy men. Food Chem. Toxicol. 2008, 46 (Suppl. 7), S54–S60. [Google Scholar] [CrossRef]
- Bian, X.; Tu, P.; Chi, L.; Gao, B.; Ru, H.; Lu, K. Saccharin induced liver inflammation in mice by altering the gut microbiota and its metabolic functions. Food Chem. Toxicol. 2017, 107, 530–539. [Google Scholar] [CrossRef]
- Additives, E.P.O.F.; Food, N.S.A.T.; Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Dusemund, B.; Filipič, M.; Frutos, M.J.; Galtier, P.; et al. Safety in use of glucosylated steviol glycosides as a food additive in different food categories. EFSA J. 2018, 16, e05181. [Google Scholar]
- Nettleton, J.E.; Klancic, T.; Schick, A.; Choo, A.C.; Shearer, J.; Borgland, S.L.; Chleilat, F.; Mayengbam, S.; Reimer, R.A. Low-Dose Stevia (Rebaudioside A) Consumption Perturbs Gut Microbiota and the Mesolimbic Dopamine Reward System. Nutrients 2019, 11, 1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Carrillo, B.E.; Rosales-Gomez, C.A.; Ramirez-Duran, N.; Resendiz-Albor, A.A.; Escoto-Herrera, J.A.; Mondragon-Velasquez, T.; Valdes-Ramos, R.; Castillo-Cardiel, A. Effect of Chronic Consumption of Sweeteners on Microbiota and Immunity in the Small Intestine of Young Mice. Int. J. Food Sci. 2019, 2019, 9619020. [Google Scholar] [CrossRef] [PubMed]
- Mahalak, K.K.; Firrman, J.; Tomasula, P.M.; Nunez, A.; Lee, J.J.; Bittinger, K.; Rinaldi, W.; Liu, L.S. Impact of Steviol Glycosides and Erythritol on the Human and Cebus apella Gut Microbiome. J. Agric. Food Chem. 2020. [Google Scholar] [CrossRef]
- Grembecka, M. Sugar alcohols—Their role in the modern world of sweeteners: A review. Eur. Food Res. Technol. 2015, 241, 1–14. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Public Consultation on the Draft Protocol for the Assessment of Hazard Identification and Characterisation of the Sweeteners. Available online: http://www.efsa.europa.eu/en/consultations/call/public-consultation-draft-protocol-assessment (accessed on 24 December 2019).
- Oku, T.; Okazaki, M. Laxative threshold of sugar alcohol erythritol in human subjects. Nutr. Res. 1996, 16, 577–589. [Google Scholar] [CrossRef]
- Caballero, B.; Finglas, P.; Toldrá, F. Encyclopedia of Food and Health; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Livesey, G. Health potential of polyols as sugar replacers, with emphasis on low glycaemic properties. Nutr. Res. Rev. 2003, 16, 163–191. [Google Scholar] [CrossRef] [Green Version]
- Gostner, A.; Blaut, M.; Schäffer, V.; Kozianowski, G.; Theis, S.; Klingeberg, M.; Dombrowski, Y.; Martin, D.; Ehrhardt, S.; Taras, D. Effect of isomalt consumption on faecal microflora and colonic metabolism in healthy volunteers. Br. J. Nutr. 2006, 95, 40–50. [Google Scholar] [CrossRef]
- Jiang, L.; Xie, M.; Chen, G.; Qiao, J.; Zhang, H.; Zeng, X. Phenolics and Carbohydrates in Buckwheat Honey Regulate the Human Intestinal Microbiota. Evid. Based Complement. Altern. Med. 2020, 2020, 6432942. [Google Scholar] [CrossRef]
- Schauber, J.; Weiler, F.; Gostner, A.; Melcher, R.; Kudlich, T.; Lührs, H.; Scheppach, W. Human rectal mucosal gene expression after consumption of digestible and non-digestible carbohydrates. Mol. Nutr. Food Res. 2006, 50, 1006–1012. [Google Scholar] [CrossRef]
- Ballongue, J.; Schumann, C.; Quignon, P. Effects of lactulose and lactitol on colonic microflora and enzymatic activity. Scand. J. Gastroenterol. 1997, 32, 41–44. [Google Scholar] [CrossRef]
- Nath, A.; Haktanirlar, G.; Varga, A.; Molnar, M.A.; Albert, K.; Galambos, I.; Koris, A.; Vatai, G. Biological Activities of Lactose-Derived Prebiotics and Symbiotic with Probiotics on Gastrointestinal System. Medicina 2018, 54, 18. [Google Scholar] [CrossRef] [Green Version]
- Finney, M.; Smullen, J.; Foster, H.A.; Brokx, S.; Storey, D.M. Effects of low doses of lactitol on faecal microflora, pH, short chain fatty acids and gastrointestinal symptomology. Eur. J. Nutr. 2007, 46, 307. [Google Scholar] [CrossRef] [PubMed]
- Carocho, M.; Morales, P.; Ferreira, I.C. Sweeteners as food additives in the XXI century: A review of what is known, and what is to come. Food Chem. Toxicol. 2017, 107, 302–317. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.R.; Kang, S.Y.; Kim, S.E.; Lee, S.J.; Lee, Y.C.; Sung, M.K. Prebiotic UG1601 mitigates constipation-related events in association with gut microbiota: A randomized placebo-controlled intervention study. World J. Gastroenterol. 2019, 25, 6129–6144. [Google Scholar] [CrossRef] [PubMed]
- Björklund, M.; Ouwehand, A.C.; Forssten, S.D.; Nikkilä, J.; Tiihonen, K.; Rautonen, N.; Lahtinen, S.J. Gut microbiota of healthy elderly NSAID users is selectively modified with the administration of Lactobacillus acidophilus NCFM and lactitol. Age 2012, 34, 987–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, C.H.; Krych, L.; Nielsen, D.S.; Vogensen, F.K.; Hansen, L.H.; Sorensen, S.J.; Buschard, K.; Hansen, A.K. Early life treatment with vancomycin propagates Akkermansia muciniphila and reduces diabetes incidence in the NOD mouse. Diabetologia 2012, 55, 2285–2294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reunanen, J.; Kainulainen, V.; Huuskonen, L.; Ottman, N.; Belzer, C.; Huhtinen, H.; de Vos, W.M.; Satokari, R. Akkermansia muciniphila Adheres to Enterocytes and Strengthens the Integrity of the Epithelial Cell Layer. Appl. Environ. Microbiol. 2015, 81, 3655–3662. [Google Scholar] [CrossRef] [Green Version]
- Singla, V.; Chakkaravarthi, S. Applications of prebiotics in food industry: A review. Food Sci. Technol. Int. 2017, 23, 649–667. [Google Scholar] [CrossRef]
- Wang, Y.N.; Meng, X.C.; Dong, Y.F.; Zhao, X.H.; Qian, J.M.; Wang, H.Y.; Li, J.N. Effects of probiotics and prebiotics on intestinal microbiota in mice with acute colitis based on 16S rRNA gene sequencing. Chin. Med. J. 2019, 132, 1833–1842. [Google Scholar] [CrossRef]
- Hyams, J.S. Sorbitol intolerance: An unappreciated cause of functional gastrointestinal complaints. Gastroenterology 1983, 84, 30–33. [Google Scholar] [CrossRef]
- Robbins, G.B.; Lewis, K.H. Fermentation of sugar acids by bacteria. J. Bacteriol. 1940, 39, 399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernier, J.; Pascal, G. Valeur énergetique des polyols (sucres-alcools). Méd. Nutr. 1990, 26, 221–238. [Google Scholar]
- Maekawa, M.; Maekawa, M.; Ushida, K.; Hoshi, S.; Kashima, N.; Ajisaka, K.; Maekawa, M.; Ushida, K.; Hoshi, S.; Kashima, N. Butyrate and propionate production from D-mannitol in the large intestine of pig and rat. Microb. Ecol. Health Dis. 2005, 17, 169–176. [Google Scholar] [CrossRef]
- Mäkinen, K.K. History, Safety, and Dental Properties of Xylitol. 2004. Available online: http://www.naturallysweet.com.au/uploads/50072/ufiles/download_info/History_Safety_and_Dental_Properties_of_Xylitol.pdf (accessed on 31 March 2020).
- Dubach, U.; Feiner, E.; Forgó, I. Oral tolerance of Xylit in subjects with normal metabolism. Schweiz. Med. Wochenschr. 1969, 99, 190–194. [Google Scholar]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plaza-Diaz, J.; Pastor-Villaescusa, B.; Rueda-Robles, A.; Abadia-Molina, F.; Ruiz-Ojeda, F.J. Plausible Biological Interactions of Low- and Non-Calorie Sweeteners with the Intestinal Microbiota: An Update of Recent Studies. Nutrients 2020, 12, 1153. https://doi.org/10.3390/nu12041153
Plaza-Diaz J, Pastor-Villaescusa B, Rueda-Robles A, Abadia-Molina F, Ruiz-Ojeda FJ. Plausible Biological Interactions of Low- and Non-Calorie Sweeteners with the Intestinal Microbiota: An Update of Recent Studies. Nutrients. 2020; 12(4):1153. https://doi.org/10.3390/nu12041153
Chicago/Turabian StylePlaza-Diaz, Julio, Belén Pastor-Villaescusa, Ascensión Rueda-Robles, Francisco Abadia-Molina, and Francisco Javier Ruiz-Ojeda. 2020. "Plausible Biological Interactions of Low- and Non-Calorie Sweeteners with the Intestinal Microbiota: An Update of Recent Studies" Nutrients 12, no. 4: 1153. https://doi.org/10.3390/nu12041153
APA StylePlaza-Diaz, J., Pastor-Villaescusa, B., Rueda-Robles, A., Abadia-Molina, F., & Ruiz-Ojeda, F. J. (2020). Plausible Biological Interactions of Low- and Non-Calorie Sweeteners with the Intestinal Microbiota: An Update of Recent Studies. Nutrients, 12(4), 1153. https://doi.org/10.3390/nu12041153