Are Supplements Safe? Effects of Gallic and Ferulic Acids on In Vitro Cell Models

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. HPLC-MS/MS Analysis
2.3. Cell Model Systems
2.4. MTT Viability Assay
2.5. Cell Viability
2.6. Caspase3/7 Detection
2.7. Wound-Healing Assay
2.8. Monocyte Migration
2.9. Intestinal Equivalents
2.10. Immunohistochemical Analysis
2.11. Statistical Analysis
3. Results
3.1. Gallic and Ferulic Acid Content in Commercially Available Dietary Supplements
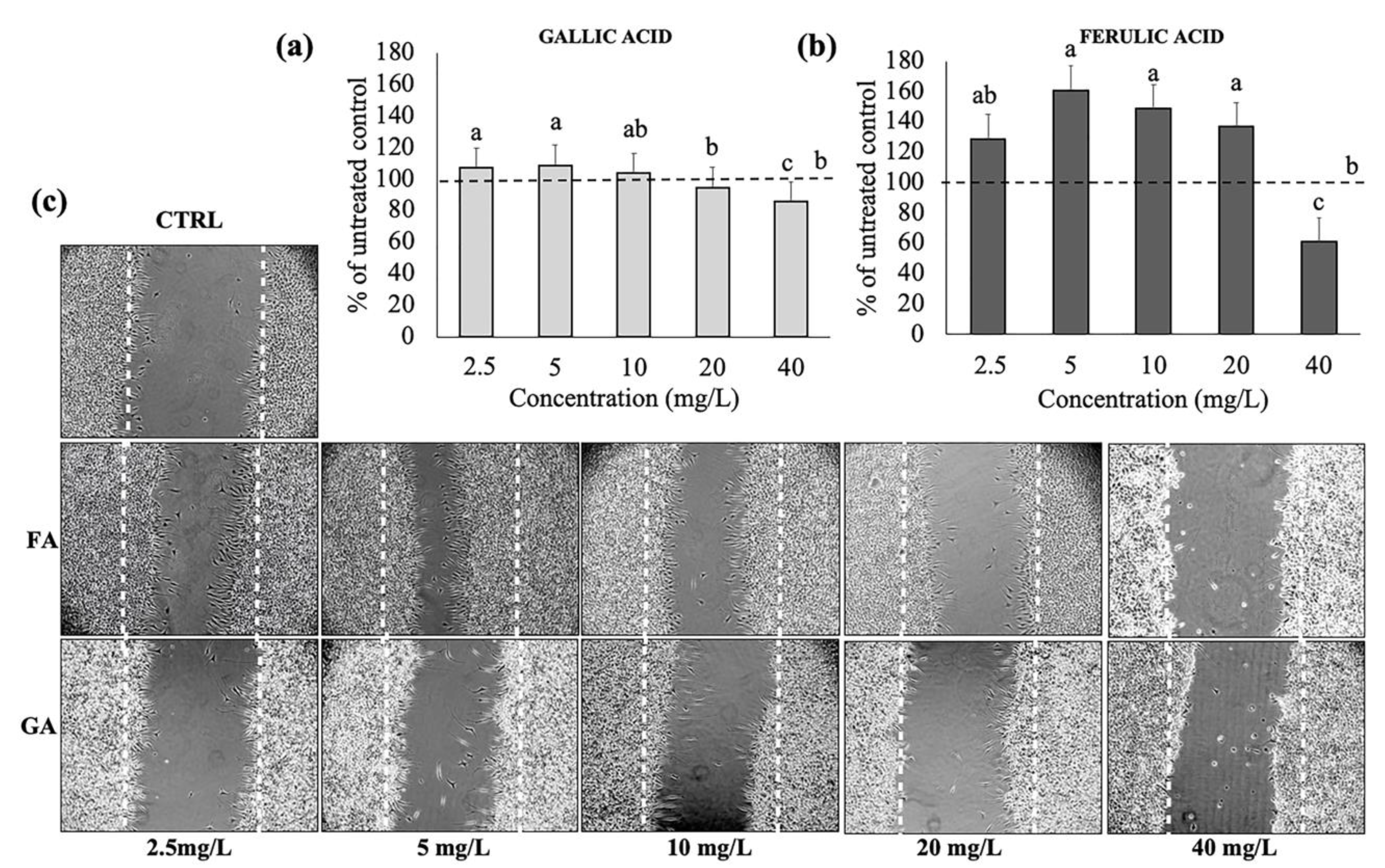
3.2. Effects of Gallic and Ferulic Acids on Mouse Fibroblast Proliferation and Migration
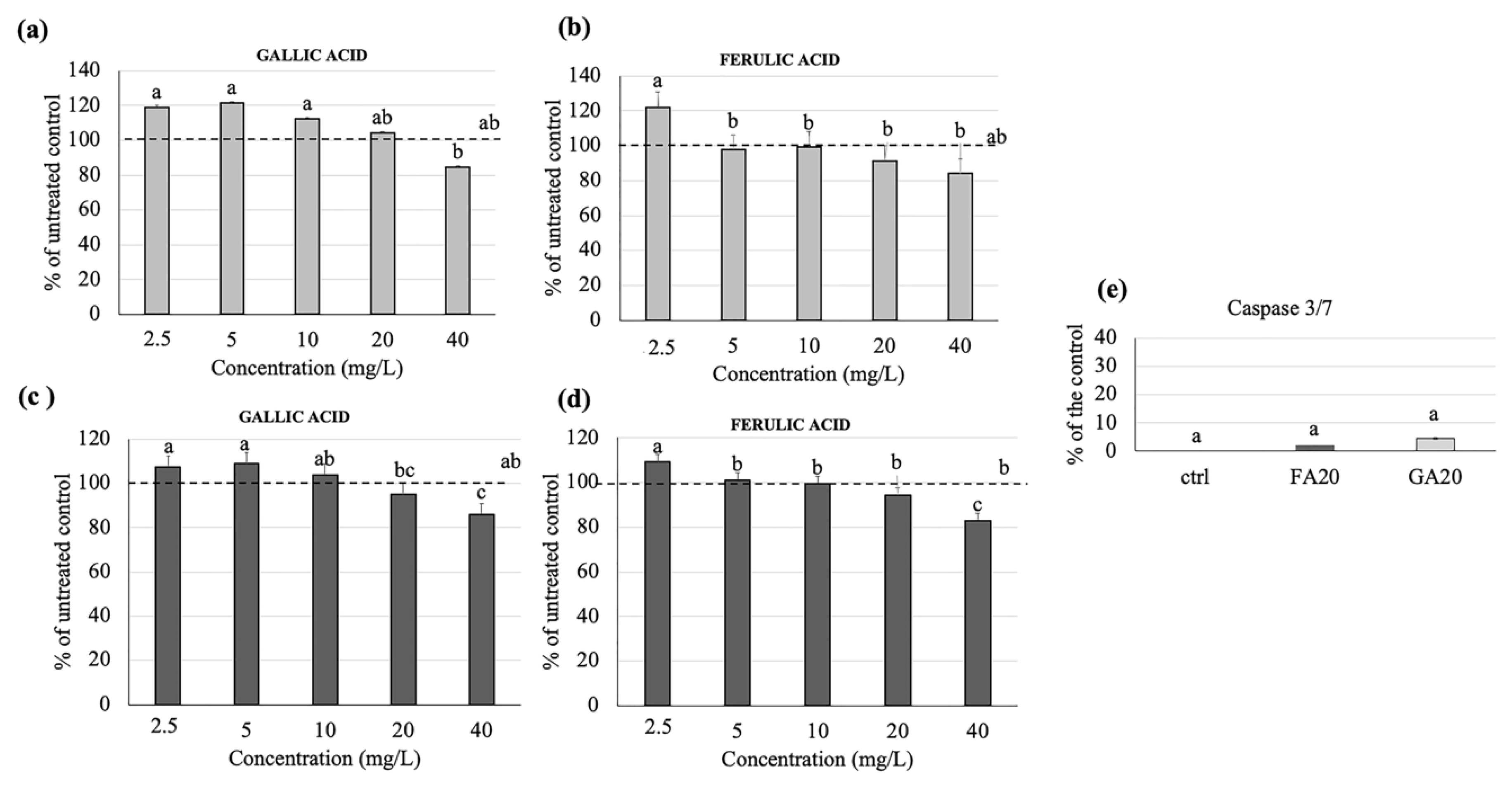
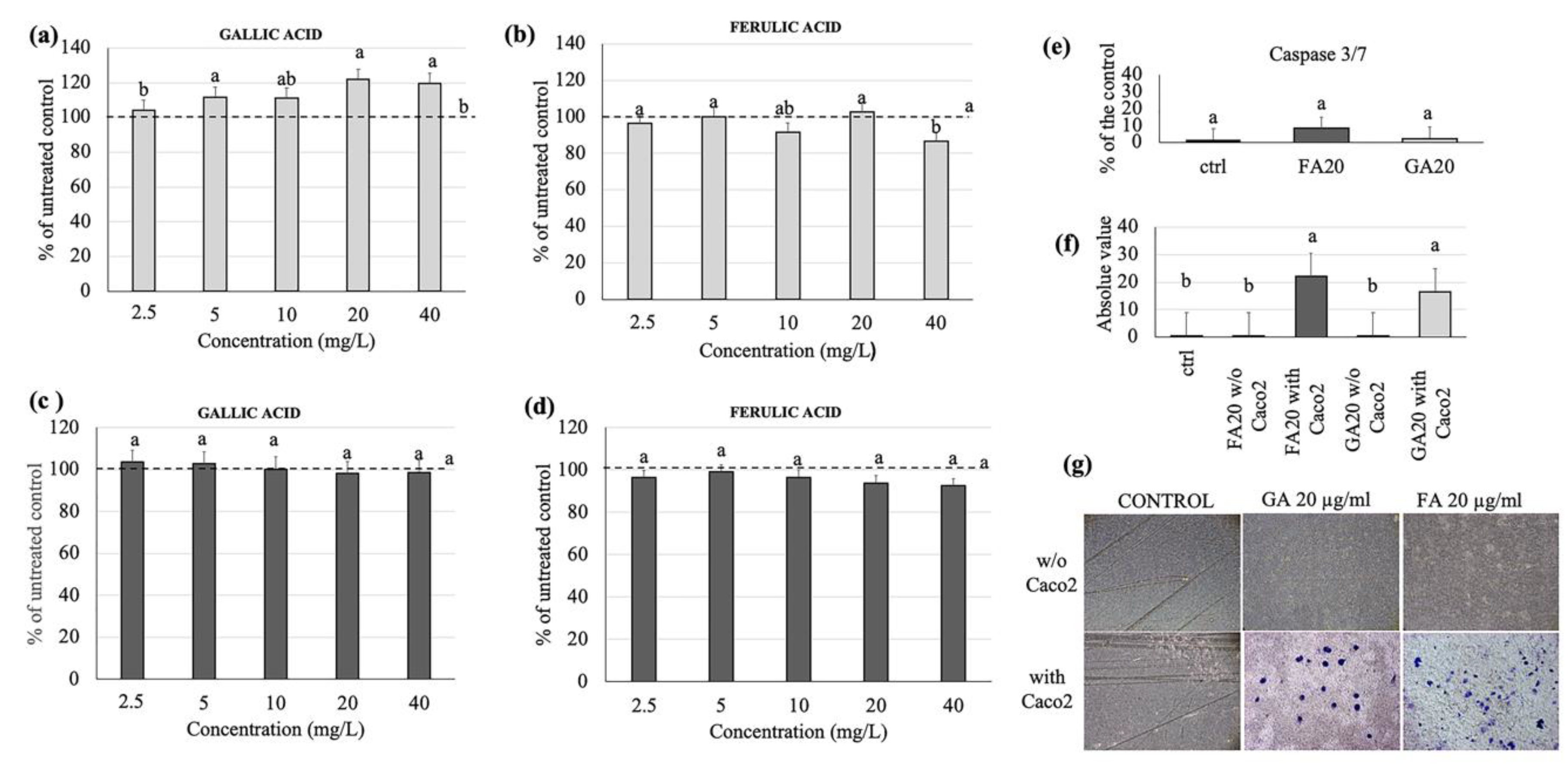
3.3. Effects of Gallic and Ferulic Acid on Human Intestinal Cell Viability
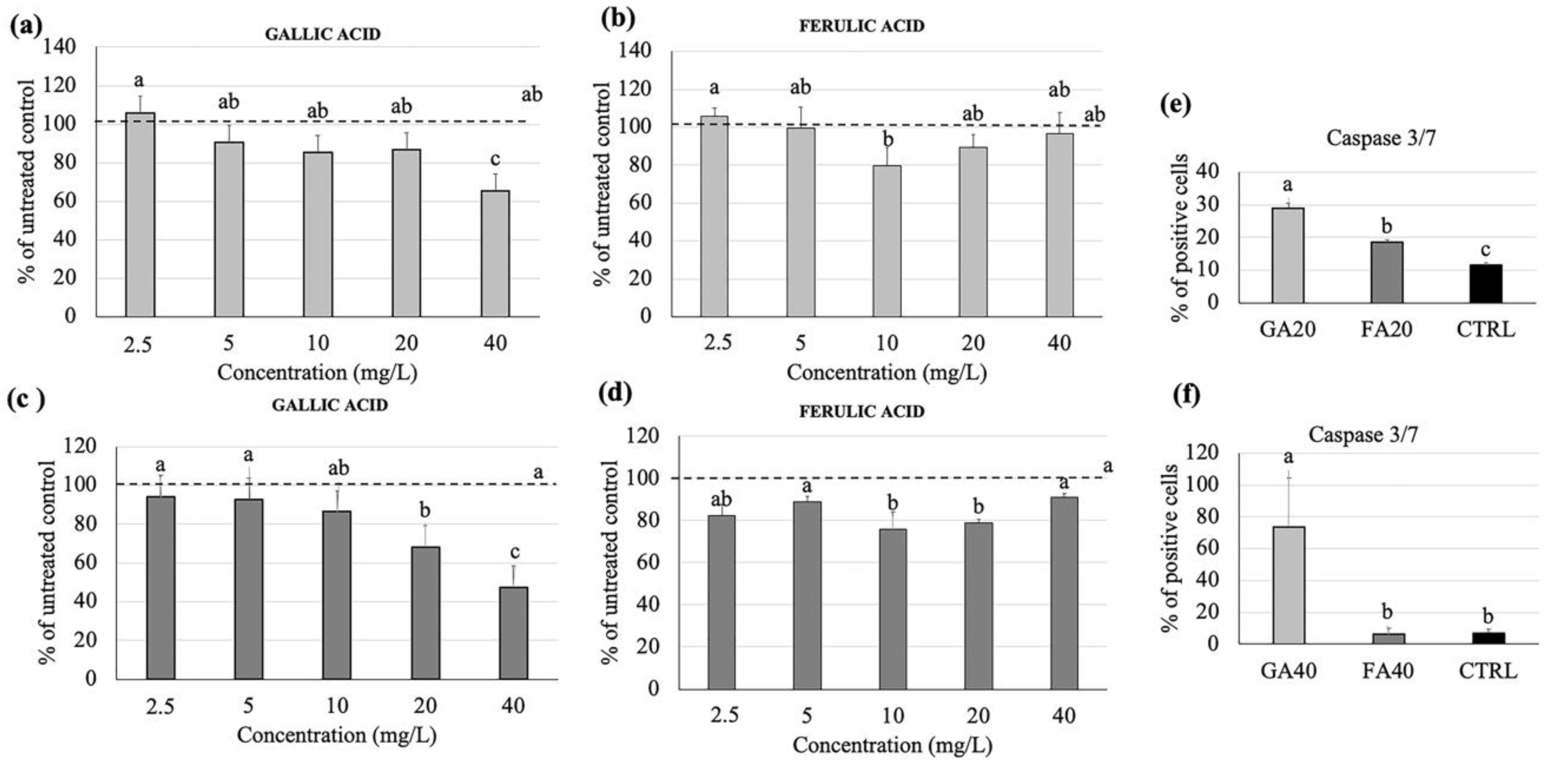
3.4. Effects of Gallic and Ferulic Acids on Human Monocyte Cell Viability
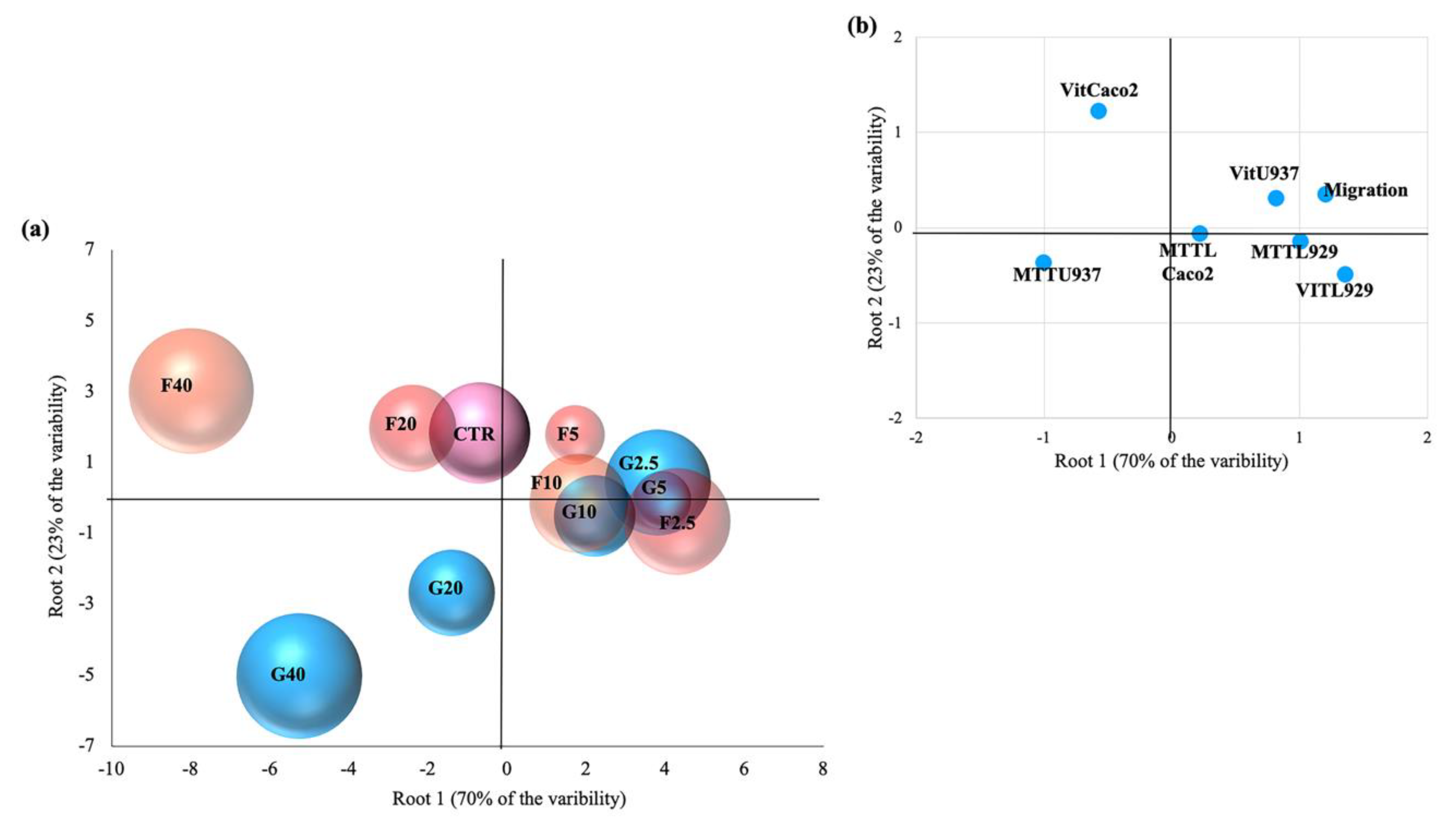
3.5. Linear Discriminant Analysis (LDA)
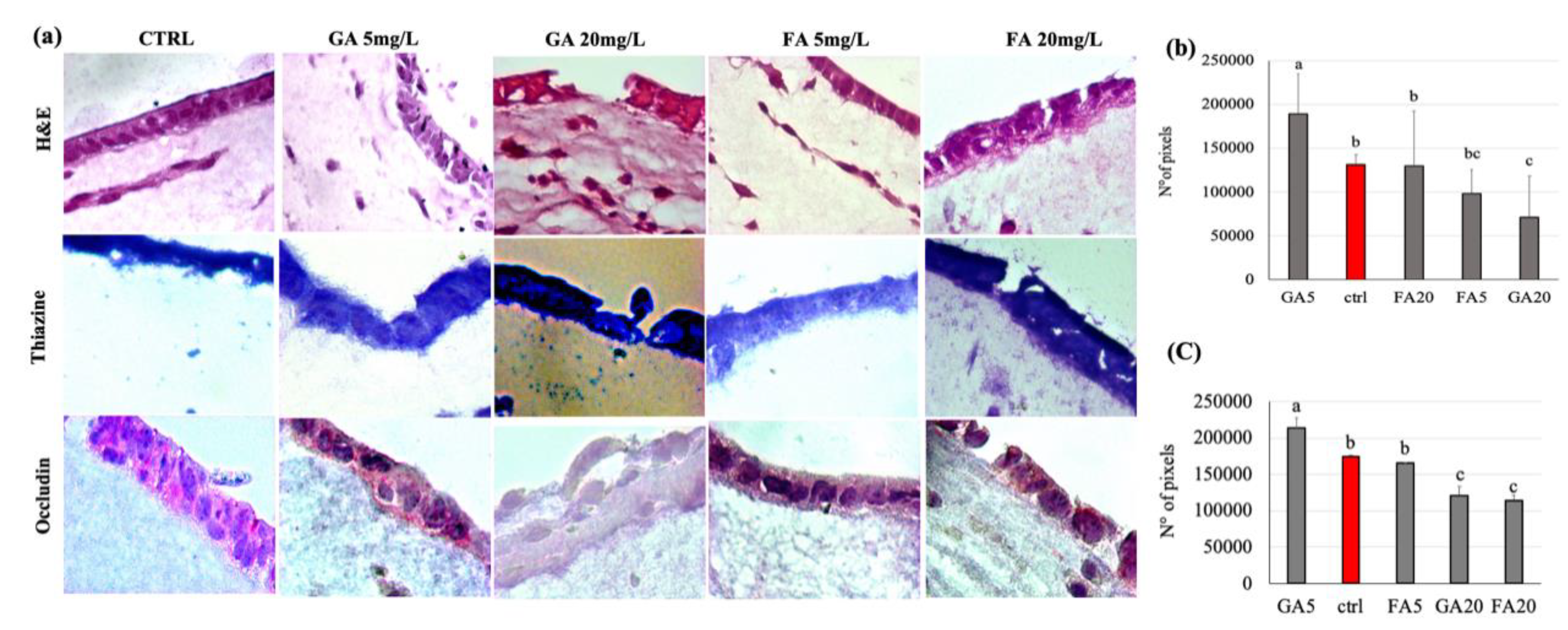
3.6. Effect of Gallic and Ferulic Acids on Intestinal Tridimensional Models
4. Discussion
4.1. Gallic and Ferulic Acids and Intestinal Tissue Cell Lines
4.2. Gallic and Ferulic Acids and Tridimensional Intestinal Equivalent Model
4.3. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef] [PubMed]
- Forni, C.; Facchiano, F.; Bartoli, M.; Pieretti, S.; Facchiano, A.; D’Arcangelo, D.; Norelli, S.; Valle, G.; Nisini, R.; Beninati, S.; et al. Beneficial Role of Phytochemicals on Oxidative Stress and Age-Related Diseases. Biomed Res. Int. 2019, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozdal, T.; Ceylan, F.D.; Eroglu, N.; Kaplan, M.; Olgun, E.O.; Capanoglu, E. Investigation of antioxidant capacity, bioaccessibility and LC-MS/MS phenolic profile of Turkish propolis. Food Res. Int. 2019, 122, 528–536. [Google Scholar] [CrossRef] [PubMed]
- D’Archivio, M.; Filesi, C.; Varì, R.; Scazzocchio, B.; Masella, R. Bioavailability of the polyphenols: Status and controversies. Int. J. Mol. Sci. 2010, 11, 1321–1342. [Google Scholar] [CrossRef]
- Fraga, C.G.; Galleano, M.; Verstraeten, S.V.; Oteiza, P.I. Basic biochemical mechanisms behind the health benefits of polyphenols. Mol. Aspects Med. 2010, 31, 435–445. [Google Scholar] [CrossRef]
- Goszcz, K.; Duthie, G.G.; Stewart, D.; Leslie, S.J.; Megson, I.L. Bioactive polyphenols and cardiovascular disease: Chemical antagonists, pharmacological agents or xenobiotics that drive an adaptive response? Br. J. Pharmacol. 2017, 174, 1209–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Keogh, J.B.; Clifton, P.M. Polyphenols and Glycemic Control. Nutrients 2016, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.S.; Satsu, H.; Bae, M.J.; Zhao, Z.; Ogiwara, H.; Totsuka, M.; Shimizu, M. Anti-inflammatory effect of chlorogenic acid on the IL-8 production in Caco-2 cells and the dextran sulphate sodium-induced colitis symptoms in C57BL/6 mice. Food Chem. 2015, 168, 167–175. [Google Scholar] [CrossRef]
- Zhao, Z.; Shin, H.S.; Satsu, H.; Totsuka, M.; Shimizu, M. 5-caffeoylquinic acid and caffeic acid down-regulate the oxidative stress- and TNF-alpha-induced secretion of interleukin-8 from Caco-2 cells. J. Agric. Food Chem. 2008, 28, 3863–3868. [Google Scholar] [CrossRef]
- Tulyathan, V.; Boulton, R.B.; Singleton, V.L. Oxygen uptake by gallic acid as a model for similar reactions in wines. J. Agric. Food Chem. 1989, 37, 844–849. [Google Scholar] [CrossRef]
- Gil-Longo, J.; Gonzalez-Vazquez, C. Vascular pro-oxidant effects secondary to the autoxidation of gallic acid in rat aorta. J. Nutr. Biochem. 2010, 21, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Babich, H.; Gottesman, R.T.; Liebling, E.J.; Schuck, A.G. Theaflavin- 3-gallate and theaflavin-30-gallate, polyphenols in black tea with prooxidant properties. Basic. Clin. Pharmacol. Toxicol. 2008, 103, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Bellion, P.; Olk, M.; Will, F.; Dietrich, H.; Baum, M.; Eisenbrand, G.; Janzowski, C. Formation of hydrogen peroxide in cell culture media by apple polyphenols and its effect on antioxidant biomarkers in the colon cell line HT-29. Mol. Nutr. Food Res. 2009, 53, 1226–1236. [Google Scholar] [CrossRef] [PubMed]
- Di Loreto, A.; Bosi, S.; Montero, L.; Bregola, V.; Marotti, I.; Sferrazza, R.E.; Dinelli, G.; Herrero, M.; Cifuentes, A. Determination of phenolic compounds in ancient and modern durum wheat genotypes. Electrophoresis 2018, 39. [Google Scholar] [CrossRef]
- Ayaz, F.A.; Hayirlioglu-Ayaz, S.; Gruz, J.; Novak, O.; Strnad, M. Separation, characterization, and quantitation of phenolic acids in a little-known blueberry (Vaccinium arctostaphylos L.) fruit by HPLC-MS. J. Agric. Food Chem. 2005, 53, 8116–8122. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Delgado, M.A.; Gonzalez-Hernandez, G.; Condegonzalez, J.E.; Perez-Trujillo, J.P. Principal component analysis of the polyphenols content in young red wines. Food Chem. 2002, 78, 523–532. [Google Scholar] [CrossRef]
- Zuo, T.; Ng, S.C. The Gut Microbiota in the Pathogenesis and Therapeutics of Inflammatory Bowel Disease. Front Microbiol. 2018, 25, 2247. [Google Scholar] [CrossRef]
- Li, J.; Chen, J.; Kirsner, R. Pathophysiology of acute wound healing. Clin. Dermatol. 2007, 25, 9–18. [Google Scholar] [CrossRef]
- Yang, D.J.; Moh, S.H.; Son, D.H.; You, S.; Kinyua, A.W.; Ko, C.M.; Song, M.; Yeo, J.; Choi, Y.; Kim, K.W. Gallic acid promotes wound healing in normal and hyperglucidic conditions. Molecules 2016, 21, 899. [Google Scholar] [CrossRef] [Green Version]
- Ceriotti, L.; Meloni, M. La valutazione dell’assorbimento intestinale in vitro. L’integratore Nutr. 2014, 17, 62–65. [Google Scholar]
- Hirayama, D.; Iida, T.; Nakase, H. The Phagocytic Function of Macrophage-Enforcing Innate Immunity and Tissue Homeostasis. Int. J. Mol. Sci. 2017, 19, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junger, W.G. Immune cell regulation by autocrine purinergic signaling. Rev. Immunol. 2011, 11, 201–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoni, D.; Burckel, H.; Josset, E.; Noel, G. Three-dimensional cell culture: A breakthrough in vivo. Int. J. Mol. Sci. 2015, 16, 5517–5527. [Google Scholar] [CrossRef] [PubMed]
- Sosulski, F.; Krygier, K.; Hogge, L. Free, esterified, and insoluble bound phenolic acids. 3. Composition of phenolic acids in cereal and potato flours. J. Agric. Food Chem. 1982, 30, 337–340. [Google Scholar] [CrossRef]
- Keskin-Šašić, I.A.; Tahirović, I.A.; Topčagić, A.A.; Klepo, L.A.; Salihović, M.B.; Ibragić, S.A.; Toromanović, J.C.; Ajanović, A.D.; Velispahić, E.A. Total Phenolic Content and Antioxidant Capacity of Fruit Juices. Bull. Chem. Technol. Bosnia Herzeg. 2012, 39, 25–28. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gallic Acid (mg/g) ± s.d. | Ferulic Acid (mg/g) ± s.d. | |
|---|---|---|
| Blueberry Supplement | 0.3252 ± 0.030 | 1.3695 ± 0.140 |
| Supplement A | 0.6354 ± 0.035 | 1.8494 ± 0.180 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truzzi, F.; Valerii, M.C.; Tibaldi, C.; Zhang, Y.; Abduazizova, V.; Spisni, E.; Dinelli, G. Are Supplements Safe? Effects of Gallic and Ferulic Acids on In Vitro Cell Models. Nutrients 2020, 12, 1591. https://doi.org/10.3390/nu12061591
Truzzi F, Valerii MC, Tibaldi C, Zhang Y, Abduazizova V, Spisni E, Dinelli G. Are Supplements Safe? Effects of Gallic and Ferulic Acids on In Vitro Cell Models. Nutrients. 2020; 12(6):1591. https://doi.org/10.3390/nu12061591
Chicago/Turabian StyleTruzzi, Francesca, Maria Chiara Valerii, Camilla Tibaldi, Yanxin Zhang, Veronika Abduazizova, Enzo Spisni, and Giovanni Dinelli. 2020. "Are Supplements Safe? Effects of Gallic and Ferulic Acids on In Vitro Cell Models" Nutrients 12, no. 6: 1591. https://doi.org/10.3390/nu12061591
APA StyleTruzzi, F., Valerii, M. C., Tibaldi, C., Zhang, Y., Abduazizova, V., Spisni, E., & Dinelli, G. (2020). Are Supplements Safe? Effects of Gallic and Ferulic Acids on In Vitro Cell Models. Nutrients, 12(6), 1591. https://doi.org/10.3390/nu12061591