Lipotoxic Injury Differentially Regulates Brain Microvascular Gene Expression in Male Mice

 , ,
, ,
Abstract
:1. Introduction
2. Methods
2.1. Experimental Animals
2.2. Blood Metabolic and Hormone Assays
2.3. Isolation and Cryosection of Murine Brain Hippocampus
2.4. Laser Capture Microdissection (LCM) of Hippocampal Microvessels
2.5. RNA Extraction from Laser Captured Brain Microvessels
2.6. Microarray Hybridization and Transcriptome Analysis
2.7. qRT-PCR Analysis of Gene Expression in Murine Hippocampal Microvessels
2.8. Bioinformatic Analysis
2.9. Statistical Methods
3. Results
3.1. Model of Hyperlipidemia
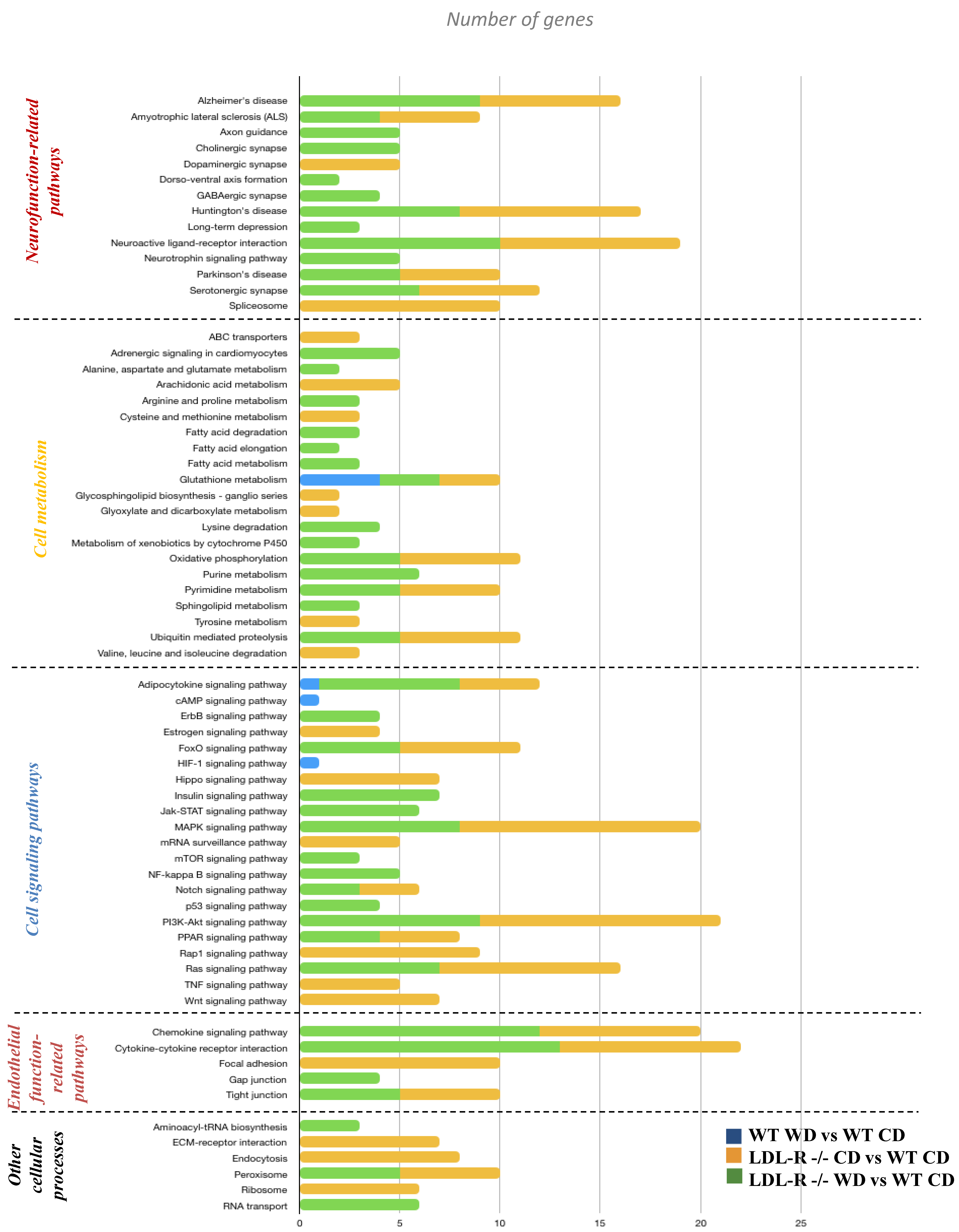
3.2. Effect of the Western Diet on Brain Hippocampal Microvessel Gene Expression
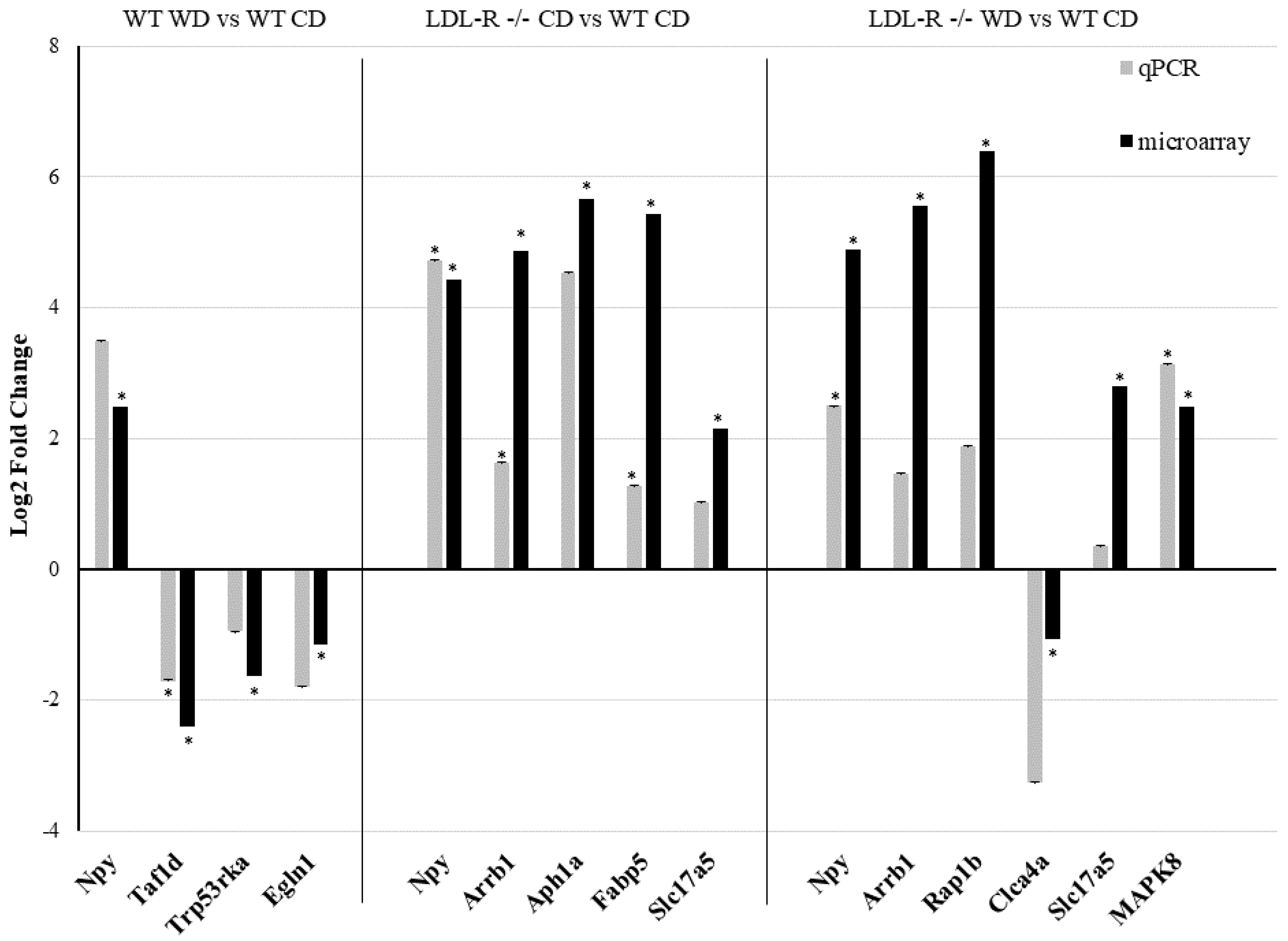
3.3. Effect of the Western Diet on Expression of Protein-coding Genes in Brain Hippocampal Microvessels
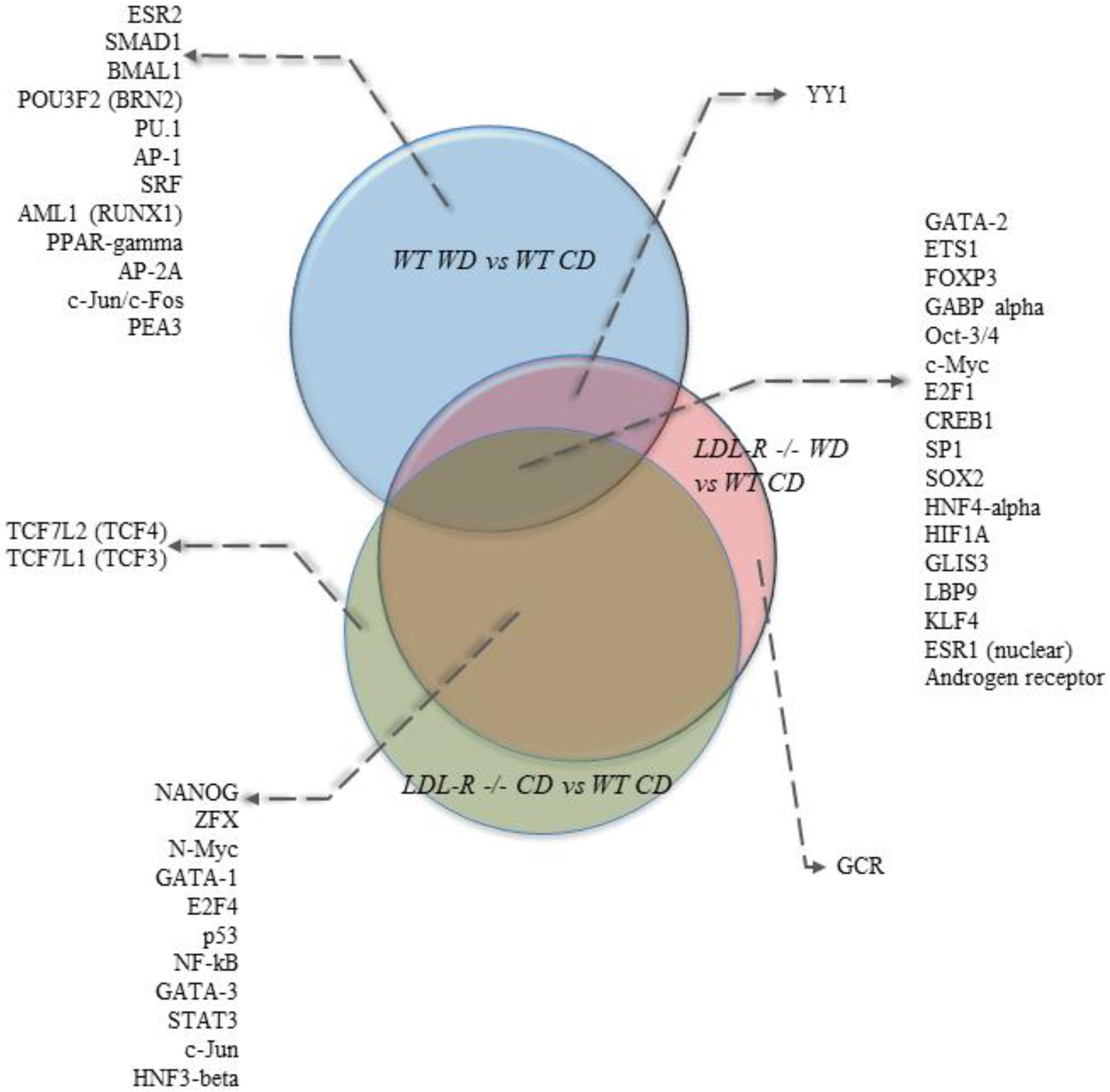
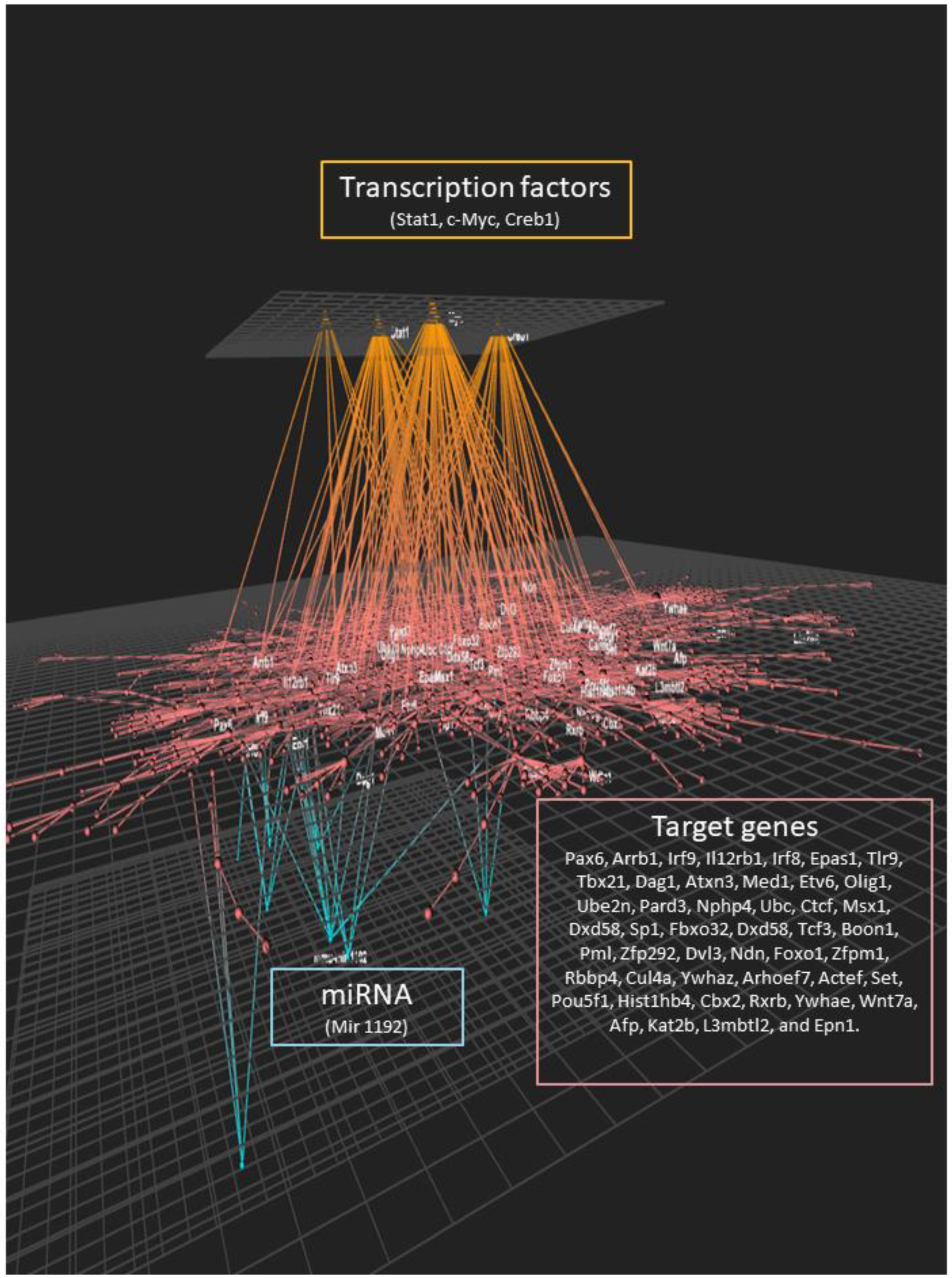
3.4. Potential Transcription Factors Involved in the Genomic Effects of the Western Diet on Brain Hippocampal Microvessels
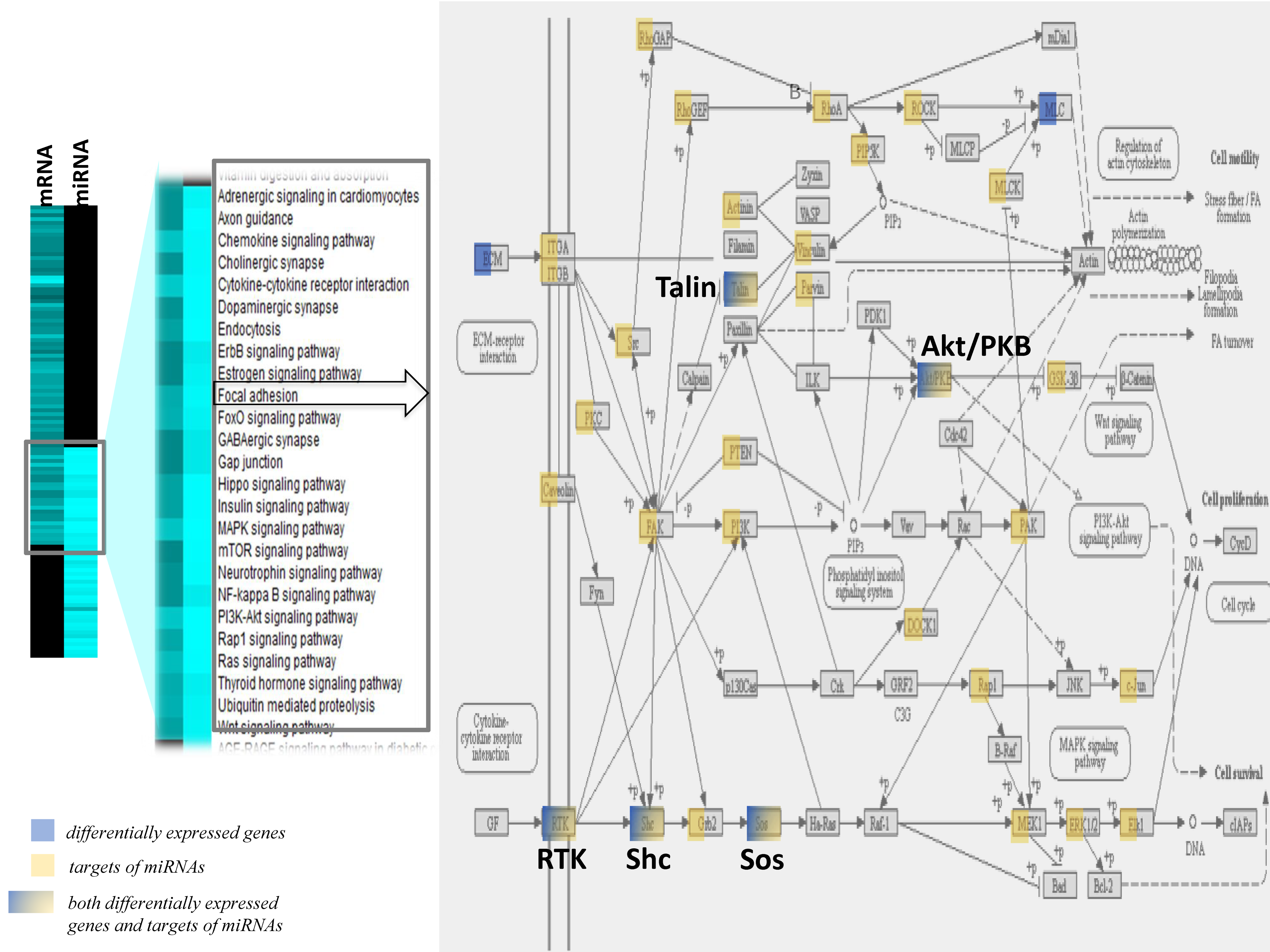
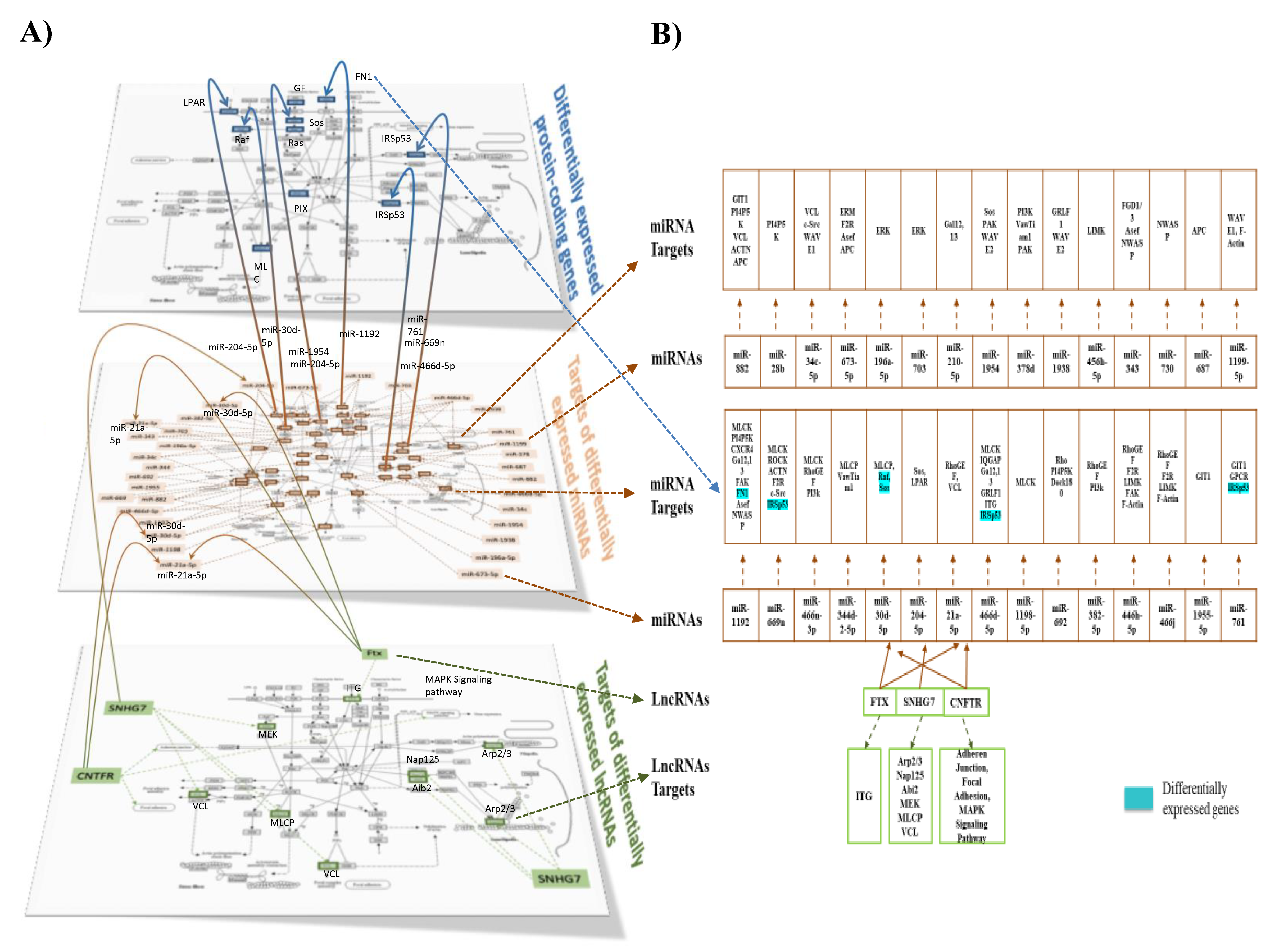
3.5. Impact of the Western Diet on Expression of miRNA, their Targets and Pathways in Brain Microvessels
3.6. Impact of the Western Diet on Expression of snoRNAs and lncRNAs in Brain Microvessels
3.7. Integration of Multiomics Data
4. Discussion
- ➢
- The WD resulted in the differential expression (primarily up-regulation) of a large number of genes (1972), representing 5.7% of the genome of microvessels in the hippocampus of male mice;
- ➢
- Overall, the differential gene expression was associated with the differential regulation of cell signaling proteins and their transcription factors, with complex mechanisms of action for genes that regulate increased endothelial dysfunction following lipid stress as the main disruption, particularly via pathways that serve to increase permeability, consistent with the previously reported increase in BBB permeability following the WD;
- ➢
- There were some differences in the differential gene expression for diet and genotype. Differentially expressed genes involved in focal adhesion, ECM–receptor interaction, and signaling pathways (such as PI3K-Akt, TNF, Jak-STAT, and Ras) were up-regulated with lipid injury in the LR genotype while down-regulated by the WD in the wild type mice;
- ➢
- Most of the differential gene expression was attributable to protein-coding genes (85%), but approximately 4% was due to the differential expression of miRNAs, and 10% was due to other non-protein-coding RNAs not previously found to be affected by the WD, including mostly long non-coding RNAs (lncRNAs) and small nucleolar RNAs (snoRNAs). The targets of lncRNAs included genes involved in NF-kB signaling, Ras/Rap signaling, focal adhesion, actin cytoskeleton organization, cell adhesion, chemokine signaling, tight junctions, and adherent junctions;
- ➢
- Lipotoxic injury resulted in previously unreported complex and multilevel molecular regulation of the hippocampal microvasculature involving transcriptional and post-transcriptional regulation. Post-transcriptional regulation accounted for up to a third of the differential gene expression;
- ➢
- Specific detailed examples of this complex regulation for the representative genes, pathways, transcription factors, and non-coding RNAs are provided below.
4.1. Lipotoxic Injury Up-Regulates Hippocampal Microvascular Gene Expression
4.2. Regulation of Gene Networks and Pathways for Endothelial Permeability, Neurofunction and Serotonergic Pathways
4.3. Differential Regulation of Transcription Factors for Endothelial Dysfunction: HNF4a, KLF4, CREB1
4.4. Novel Lipotoxic Injury-Mediated Differential Expression of non-Coding RNAs (miRNA, sno RNA, lncRNA), and their Targets: Implications for Cognitive Dysfunction
4.5. An Integrated Multi-omics Approach Reveals a Complex and Integrated Transcriptional and Post Transcriptional Response to Lipid Injury in Brain Microvessels
4.6. Summary and Implications for Future Research
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Reitz, C.; Brayne, C.; Mayeux, R. Epidemiology of Alzheimer disease. Nat. Rev. Neurol. 2011, 7, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Kalaria, R.N.; Erkinjuntti, T. Small vessel disease and subcortical vascular dementia. J. Clin. Neurol. 2006, 2, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, A.D. Distinct pathways for cognitive decline in the presence of Alzheimer’s disease pathology or cerebrovascular disease. Brain A J. Neurol. 2016, 139, 2340–2341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dye, L.; Boyle, N.B.; Champ, C.; Lawton, C. The relationship between obesity and cognitive health and decline. Proc. Nutr. Soc. 2017, 76, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schilling, S.; Tzourio, C.; Soumaré, A.; Kaffashian, S.; Dartigues, J.F.; Ancelin, M.L.; Samieri, C.; Dufouil, C.; Debette, S. Differential associations of plasma lipids with incident dementia and dementia subtypes in the 3C Study: A longitudinal, population-based prospective cohort study. PLoS Med. 2017, 14, e1002265. [Google Scholar] [CrossRef] [PubMed]
- Francis, H.; Stevenson, R. The longer-term impacts of Western diet on human cognition and the brain. Appetite 2013, 63, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Rutkowsky, J.M.; Lee, L.L.; Puchowicz, M.; Golub, M.S.; Befroy, D.E.; Wilson, D.W.; Anderson, S.; Cline, G.; Bini, J.; Borkowski, K.; et al. Reduced cognitive function, increased blood-brain-barrier transport and inflammatory responses, and altered brain metabolites in LDLr-/-and C57BL/6 mice fed a western diet. PLoS ONE 2018, 13, e0191909. [Google Scholar] [CrossRef] [Green Version]
- Palmer, A.M. The role of the blood brain barrier in neurodegenerative disorders and their treatment. J. Alzheimers Dis. 2011, 24, 643–656. [Google Scholar] [CrossRef]
- Montagne, A.; Barnes, S.R.; Sweeney, M.D.; Halliday, M.R.; Sagare, A.P.; Zhao, Z.; Toga, A.W.; Jacobs, R.E.; Liu, C.Y.; Amezcua, L.; et al. Blood-brain barrier breakdown in the aging human hippocampus. Neuron 2015, 85, 296–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aung, H.H.; Altman, R.; Nyunt, T.; Kim, J.; Nuthikattu, S.; Budamagunta, M.; Voss, J.C.; Wilson, D.; Rutledge, J.C.; Villablanca, A.C. Lipotoxic brain microvascular injury is mediated by activating transcription factor 3-dependent inflammatory and oxidative stress pathways. J. Lipid Res. 2016, 57, 955–968. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Shi, H.; Zhang, J.; Shen, Z.; Zhou, G.; Hu, M. Effects of the duration of hyperlipidemia on cerebral lipids, vessels and neurons in rats. Lipids Health Dis. 2017, 16, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, K.N.; Mounier, A.; Wolfe, C.M.; Fitz, N.F.; Carter, A.Y.; Castranio, E.L.; Kamboh, H.I.; Reeves, V.L.; Wang, J.; Han, X.; et al. Effect of high fat diet on phenotype, brain transcriptome and lipidome in Alzheimer’s model mice. Sci. Rep. 2017, 7, 4307. [Google Scholar] [CrossRef]
- Labouesse, M.A.; Polesel, M.; Clementi, E.; Muller, F.; Markkanen, E.; Mouttet, F.; Cattaneo, A.; Richetto, J. MicroRNA Expression Profiling in the Prefrontal Cortex: Putative Mechanisms for the Cognitive Effects of Adolescent High Fat Feeding. Sci. Rep. 2018, 8, 8344. [Google Scholar] [CrossRef]
- Yoon, G.; Cho, K.A.; Song, J.; Kim, Y.-K. Transcriptomic Analysis of High Fat Diet Fed Mouse Brain Cortex. Front. Genet. 2019, 10, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, R.E.; Lim, D.A. Forging our understanding of lncRNAs in the brain. Cell Tissue Res. 2018, 371, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Bieghs, V.; Van Gorp, P.J.; Wouters, K.; Hendrikx, T.; Gijbels, M.J.; van Bilsen, M.; Bakker, J.; Binder, C.J.; Lutjohann, D.; Staels, B.; et al. LDL receptor knock-out mice are a physiological model particularly vulnerable to study the onset of inflammation in non-alcoholic fatty liver disease. PLoS ONE 2012, 7, e30668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuthikattu, S.; Milenkovic, D.; Rutledge, J.; Villablanca, A. The Western Diet Regulates Hippocampal Microvascular Gene Expression: An Integrated Genomic Analyses in Female Mice. Sci. Rep. 2019, 9, 19058. [Google Scholar] [CrossRef] [Green Version]
- Kochanek, K.D.; Murphy, S.L.; Xu, J.; Tejada-Vera, B. Deaths: Final Data for 2014. Cent. Dis. Control Prev. 2016, 65, 1–122. [Google Scholar]
- Ball, H.J.; McParland, B.; Driussi, C.; Hunt, N.H. Isolating vessels from the mouse brain for gene expression analysis using laser capture microdissection. Brain Res. Brain Res. Protoc. 2002, 9, 206–213. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3--new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Supek, F.; Bosnjak, M.; Skunca, N.; Smuc, T. REVIGO summarizes and visualizes long lists of gene ontology terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stockel, D.; Kehl, T.; Trampert, P.; Schneider, L.; Backes, C.; Ludwig, N.; Gerasch, A.; Kaufmann, M.; Gessler, M.; Graf, N.; et al. Multi-omics enrichment analysis using the GeneTrail2 web service. Bioinformatics 2016, 32, 1502–1508. [Google Scholar] [CrossRef] [PubMed]
- Dweep, H.; Sticht, C.; Pandey, P.; Gretz, N. miRWalk--database: Prediction of possible miRNA binding sites by “walking” the genes of three genomes. J. Biomed. Inform. 2011, 44, 839–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caraux, G.; Pinloche, S. PermutMatrix: A graphical environment to arrange gene expression profiles in optimal linear order. Bioinformatics 2005, 21, 1280–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [Green Version]
- Li, J.H.; Liu, S.; Zhou, H.; Qu, L.H.; Yang, J.H. starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014, 42, D92–D97. [Google Scholar] [CrossRef] [Green Version]
- Jeggari, A.; Marks, D.S.; Larsson, E. miRcode: A map of putative microRNA target sites in the long non-coding transcriptome. Bioinformatics 2012, 28, 2062–2063. [Google Scholar] [CrossRef]
- RNAcentral: A hub of information for non-coding RNA sequences. Nucleic Acids Res. 2019, 47, D1250–D1251. [CrossRef] [PubMed] [Green Version]
- Gruben, N.; Funke, A.; Kloosterhuis, N.J.; Schreurs, M.; Sheedfar, F.; Havinga, R.; Houten, S.M.; Shiri-Sverdlov, R.; van de Sluis, B.; Kuivenhoven, J.A.; et al. Cholesterol-induced hepatic inflammation does not underlie the predisposition to insulin resistance in dyslipidemic female LDL receptor knockout mice. J. Diabetes Res. 2015, 2015, 956854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngai, Y.F.; Quong, W.L.; Glier, M.B.; Glavas, M.M.; Babich, S.L.; Innis, S.M.; Kieffer, T.J.; Gibson, W.T. Ldlr-/-mice display decreased susceptibility to Western-type diet-induced obesity due to increased thermogenesis. Endocrinology 2010, 151, 5226–5236. [Google Scholar] [CrossRef] [PubMed]
- Mehus, A.A.; Dickey, A.M.; Smith, T.P.L.; Yeater, K.M.; Picklo, M.J. Next-Generation Sequencing Identifies Polyunsaturated Fatty Acid Responsive Genes in the Juvenile Rat Cerebellum. Nutrients 2019, 11, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukriti, S.; Tauseef, M.; Yazbeck, P.; Mehta, D. Mechanisms Regulating Endothelial Permeability. Pulm. Circ. 2014, 4, 535–551. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, G.A. Neurological Diseases in Relation to the Blood–Brain Barrier. J. Cereb. Blood Flow Metab. 2012, 32, 1139–1151. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Cong, W.; Zhou, S.; Shi, Y.; Dai, W.; Zhang, H.; Wang, X.; He, B.; Zhang, Q. Small GTPases: Structure, biological function and its interaction with nanoparticles. Asian J. Pharm. Sci. 2019, 14, 30–39. [Google Scholar] [CrossRef]
- Li, Q.-F.; Decker-Rockefeller, B.; Bajaj, A.; Pumiglia, K. Activation of Ras in the Vascular Endothelium Induces Brain Vascular Malformations and Hemorrhagic Stroke. Cell Rep. 2018, 24, 2869–2882. [Google Scholar] [CrossRef] [Green Version]
- Sturza, A.; Popoiu, C.M.; Ionica, M.; Duicu, O.M.; Olariu, S.; Muntean, D.M.; Boia, E.S. Monoamine Oxidase-Related Vascular Oxidative Stress in Diseases Associated with Inflammatory Burden. Oxidative Med. Cell. Longev. 2019, 2019, 8954201. [Google Scholar] [CrossRef]
- Kataoka, H.; Ariyama, Y.; Deushi, M.; Osaka, M.; Nitta, K.; Yoshida, M. Inhibitory Effect of Serotonin Antagonist on Leukocyte-Endothelial Interactions In Vivo and In Vitro. PLoS ONE 2016, 11, e0147929. [Google Scholar] [CrossRef]
- Morgan, J.A.; Singhal, G.; Corrigan, F.; Jaehne, E.J.; Jawahar, M.C.; Breen, J.; Pederson, S.; Baune, B.T. Ceasing exercise induces depression-like, anxiety-like, and impaired cognitive-like behaviours and altered hippocampal gene expression. Brain Res. Bull. 2019, 148, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, V.; Nocentini, G.; Riccardi, C.; Ayroldi, E.; Di Nardo, P.; Roberti, R.; Binaglia, L.; Vecchini, A. Effect of dietary saturated fatty acids on HNF-4alpha DNA binding activity and ApoCIII mRNA in sedentary rat liver. Mol. Cell. Biochem. 2011, 347, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Yamanishi, K.; Doe, N.; Sumida, M.; Watanabe, Y.; Yoshida, M.; Yamamoto, H.; Xu, Y.; Li, W.; Yamanishi, H.; Okamura, H.; et al. Hepatocyte nuclear factor 4 alpha is a key factor related to depression and physiological homeostasis in the mouse brain. PLoS ONE 2015, 10, e0119021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, P.; Zhou, Z.; Natarelli, L.; Wei, Y.; Nazari-Jahantigh, M.; Zhu, M.; Grommes, J.; Steffens, S.; Weber, C.; Schober, A. Endothelial Dicer promotes atherosclerosis and vascular inflammation by miRNA-103-mediated suppression of KLF4. Nat. Commun. 2016, 7, 10521. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; Zou, X.; Jin, Y.; Gao, S.; Lv, J.; Li, B.; Cui, R. The Role of KLF4 in Alzheimer’s Disease. Front. Cell. Neurosci. 2018, 12, 12. [Google Scholar] [CrossRef] [Green Version]
- Han, X.-R.; Wen, X.; Wang, Y.-J.; Wang, S.; Shen, M.; Zhang, Z.-F.; Fan, S.-H.; Shan, Q.; Wang, L.; Li, M.-Q.; et al. Effects of CREB1 gene silencing on cognitive dysfunction by mediating PKA-CREB signaling pathway in mice with vascular dementia. Mol. Med. 2018, 24, 18. [Google Scholar] [CrossRef]
- Bao, M.H.; Luo, H.Q.; Chen, L.H.; Tang, L.; Ma, K.F.; Xiang, J.; Dong, L.P.; Zeng, J.; Li, G.Y.; Li, J.M. Impact of high fat diet on long non-coding RNAs and messenger RNAs expression in the aortas of ApoE (-/-) mice. Sci. Rep. 2016, 6, 34161. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.; Yu, J.; Wu, Z.; Si, W.; Li, X.; Liu, Y.; Zhou, J.; Deng, R.; Chen, D. MicroRNA-210-5p Contributes to Cognitive Impairment in Early Vascular Dementia Rat Model Through Targeting Snap25. Front. Mol. Neurosci. 2018, 11, 388. [Google Scholar] [CrossRef]
- Fasanaro, P.; D’Alessandra, Y.; Di Stefano, V.; Melchionna, R.; Romani, S.; Pompilio, G.; Capogrossi, M.C.; Martelli, F. MicroRNA-210 modulates endothelial cell response to hypoxia and inhibits the receptor tyrosine kinase ligand Ephrin-A3. J. Biol. Chem. 2008, 283, 15878–15883. [Google Scholar] [CrossRef] [Green Version]
- McNay, E.C.; Recknagel, A.K. Brain insulin signaling: A key component of cognitive processes and a potential basis for cognitive impairment in type 2 diabetes. Neurobiol. Learn. Mem. 2011, 96, 432–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rask-Madsen, C.; Li, Q.; Freund, B.; Feather, D.; Abramov, R.; Wu, I.H.; Chen, K.; Yamamoto-Hiraoka, J.; Goldenbogen, J.; Sotiropoulos, K.B.; et al. Loss of insulin signaling in vascular endothelial cells accelerates atherosclerosis in apolipoprotein E null mice. Cell Metab. 2010, 11, 379–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporali, A.; Pani, E.; Horrevoets, A.J.; Kraenkel, N.; Oikawa, A.; Sala-Newby, G.B.; Meloni, M.; Cristofaro, B.; Graiani, G.; Leroyer, A.S.; et al. Neurotrophin p75 receptor (p75NTR) promotes endothelial cell apoptosis and inhibits angiogenesis: Implications for diabetes-induced impaired neovascularization in ischemic limb muscles. Circ. Res. 2008, 103, e15–e26. [Google Scholar] [CrossRef] [Green Version]
- Henriksson, B.G.; Söderström, S.; Gower, A.J.; Ebendal, T.; Winblad, B.; Mohammed, A.H. Hippocampal nerve growth factor levels are related to spatial learning ability in aged rats. Behav. Brain Res. 1992, 48, 15–20. [Google Scholar] [CrossRef]
- Wimmer, I.; Tietz, S.; Nishihara, H.; Deutsch, U.; Sallusto, F.; Gosselet, F.; Lyck, R.; Muller, W.A.; Lassmann, H.; Engelhardt, B. PECAM-1 Stabilizes Blood-Brain Barrier Integrity and Favors Paracellular T-Cell Diapedesis Across the Blood-Brain Barrier During Neuroinflammation. Front. Immunol. 2019, 10, 711. [Google Scholar] [CrossRef] [Green Version]
- Stepanov, G.A.; Filippova, J.A.; Komissarov, A.B.; Kuligina, E.V.; Richter, V.A.; Semenov, D.V. Regulatory role of small nucleolar RNAs in human diseases. BioMed Res. Int. 2015, 2015, 206849. [Google Scholar] [CrossRef] [Green Version]
- Rogelj, B.; Hartmann, C.E.; Yeo, C.H.; Hunt, S.P.; Giese, K.P. Contextual fear conditioning regulates the expression of brain-specific small nucleolar RNAs in hippocampus. Eur. J. Neurosci. 2003, 18, 3089–3096. [Google Scholar] [CrossRef]
- Kim, C.; Kang, D.; Lee, E.K.; Lee, J.S. Long Noncoding RNAs and RNA-Binding Proteins in Oxidative Stress, Cellular Senescence, and Age-Related Diseases. Oxidative Med. Cell. Longev. 2017, 2017, 2062384. [Google Scholar] [CrossRef] [Green Version]
- Montes, M.; Lund, A.H. Emerging roles of lncRNAs in senescence. FEBS J. 2016, 283, 2414–2426. [Google Scholar] [CrossRef]
- Li, W.; Shen, W.; Zhang, B.; Tian, K.; Li, Y.; Mu, L.; Luo, Z.; Zhong, X.; Wu, X.; Liu, Y.; et al. Long Non-Coding RNA LncKdm2b Regulates Cortical Neuronal Differentiation by Cis-Activating Kdm2b. bioRxiv 2018, 11, 161–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labonne, J.D.J.; Lee, K.-H.; Iwase, S.; Kong, I.-K.; Diamond, M.P.; Layman, L.C.; Kim, C.-H.; Kim, H.-G. An atypical 12q24.31 microdeletion implicates six genes including a histone demethylase KDM2B and a histone methyltransferase SETD1B in syndromic intellectual disability. Hum. Genet. 2016, 135, 757–771. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Yang, W.; Qiu, Y.; Chen, H.Z. Identification of novel quantitative traits-associated susceptibility loci for APOE epsilon 4 non-carriers of Alzheimer’s disease. Curr. Alzheimer Res. 2015, 12, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, J.; Chen, X.; Xu, X.; Cao, G.; Li, H.; Wu, T. LncRNA FTX sponges miR-215 and inhibits phosphorylation of vimentin for promoting colorectal cancer progression. Gene Ther. 2018, 25, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Dave, J.M.; Bayless, K.J. Vimentin as an integral regulator of cell adhesion and endothelial sprouting. Microcirculation 2014, 21, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.; Mackenzie, I.R.; Pickering-Brown, S.M.; Gass, J.; Rademakers, R.; Lindholm, C.; Snowden, J.; Adamson, J.; Sadovnick, A.D.; Rollinson, S.; et al. Mutations in progranulin cause tau-negative frontotemporal dementia linked to chromosome 17. Nature 2006, 442, 916–919. [Google Scholar] [CrossRef] [PubMed]
- Vercellino, M.; Grifoni, S.; Romagnolo, A.; Masera, S.; Mattioda, A.; Trebini, C.; Chiavazza, C.; Caligiana, L.; Capello, E.; Mancardi, G.L.; et al. Progranulin expression in brain tissue and cerebrospinal fluid levels in multiple sclerosis. Mult. Scler. 2011, 17, 1194–1201. [Google Scholar] [CrossRef]
- Pickford, F.; Marcus, J.; Camargo, L.M.; Xiao, Q.; Graham, D.; Mo, J.R.; Burkhardt, M.; Kulkarni, V.; Crispino, J.; Hering, H.; et al. Progranulin is a chemoattractant for microglia and stimulates their endocytic activity. Am. J. Pathol. 2011, 178, 284–295. [Google Scholar] [CrossRef]
- Mullin, N.K.; Mallipeddi, N.V.; Hamburg-Shields, E.; Ibarra, B.; Khalil, A.M.; Atit, R.P. Wnt/beta-catenin Signaling Pathway Regulates Specific lncRNAs That Impact Dermal Fibroblasts and Skin Fibrosis. Front. Genet. 2017, 8, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, C.X.; Yin, J.Y.; Zhang, Y.; Wang, Z.B.; Yang, Z.Q.; He, Z.W.; Li, X.M.; Mao, X.Y.; Cui, R.T.; Li, X.J.; et al. The molecular classification of astrocytic tumors. Oncotarget 2017, 8, 96340–96350. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Ma, C.; Chen, C.; Lu, Q.; Shi, W.; Liu, Z.; Wang, H.; Guo, H. Integration of lncRNA-miRNA-mRNA reveals novel insights into oviposition regulation in honey bees. PeerJ 2017, 5, e3881. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SnoRNAs | WTWD vs. WTCD | LDL-R−/− CD vs. WTCD | LDL-R−/− CD vs. WTCD | lncRNAs | WTWD vs. WTCD | LDL-R−/− CD vs. WTCD | LDL-R−/− CD vs. WTCD |
|---|---|---|---|---|---|---|---|
| Snord61 | −39.34 | AI504432 | 56.21 | ||||
| Gm25443 | −2.32 | Cep83os | 49.06 | ||||
| AF357355 | 123.27 | Zfp91Cntf | 19.36 | 27.1 | |||
| Gm25635 | 90.8 | D4Ertd617e | 12.6 | ||||
| Gm25856 | 55.53 | 2810049E08Rik | 11.51 | ||||
| Gm22289 | 45.23 | Mir124-1 | 10.4 | ||||
| Snord95 | 27.98 | Ftx * | 6.43 | ||||
| Gm25607 | 12.59 | F420014N23Rik | 5.55 | 6.86 | |||
| Snora17 | 11.93 | 64.98 | 1700110C19Rik | 4.07 | |||
| Gm23123 | 9.93 | 6.21 | AI314278 | 2.98 | |||
| Snora23 * | 8.96 | 2310069G16Rik | 2.4 | ||||
| Snord88c | 7.89 | 1700027J07Rik | 2.27 | 2.5 | |||
| Gm24013 | 6.77 | 1700009J07Rik | 2.23 | ||||
| Gm24336 | 6.38 | 33.24 | 1700024B18Rik | 2.23 | |||
| Gm25125 | 6.24 | D130017N08Rik | 2.08 | ||||
| Gm24770 | 5.9 | 6.8 | C130071C03Rik * | 2.87 | |||
| Gm23456 | 5.32 | 11.45 | Gm10010 | 4.65 | |||
| Gm24844 | 5.3 | Gm13411 | 3.58 | 3.32 | |||
| Gm25092 | 3.89 | Gm15409 | 2.92 | ||||
| Snord107 | 3.77 | Gm26593 | 2.84 | ||||
| Snord16a * | 3.73 | Gm26643 | 2.68 | ||||
| Gm22546 | 3.59 | Gm14254 | 2.06 | ||||
| Gm24284 | 3.07 | AU022754 | 2.22 | ||||
| Snord72 | 2.99 | 1700016L21Rik | 2.2 | ||||
| Gm26070 | 2.72 | Snhg7os | 2.1 | ||||
| Gm23722 | 2.4 | 1700027H10Rik | 2.08 | ||||
| Snora5c | 2.38 | Pcsk2os2 | 2.11 | 2.03 | |||
| Gm25410 | 2.24 | Plet1os | 2.15 | ||||
| Gm22531 | 2.21 | Gldnos | 2.15 | ||||
| Gm24429 | 2.09 | Sp3os | 2.31 | ||||
| Gm26272 | 2.09 | 4921534H16Rik | 2.24 | ||||
| Gm25860 | 2.05 | 4930405O22Rik | 2.21 | ||||
| Gm26387 | 2.04 | 5330413P13Rik | 2.19 | 2.3 | |||
| Snord61 | −27.1 | −29.61 | 6330415B21Rik | 5.62 | |||
| Gm23199 | 32.34 | Mhrt * | 3.72 | ||||
| Gm24400 | 20.1 | Med9os | 3.15 | ||||
| Gm23121 | 17.62 | 9530082P21Rik | 2.75 | ||||
| Gm22935 | 11.9 | A930024E05Rik * | 2.52 | ||||
| Gm24878 | 9.74 | 9430083A17Rik | 2.45 | ||||
| Gm25401 | 9.05 | Gm26583 | 2.22 | ||||
| Gm25720 | 6.28 | 67.3 | Gm14061 | 2.13 | |||
| Gm22378 | 6.25 | Gm26777 | 15.18 | ||||
| Gm24449 | 5.46 | Gm15323 | 3.28 | ||||
| Gm22485 | 3.66 | Gm15322 | 3.28 | ||||
| Gm25371 | 3.12 | Gm12121 | 2.7 | ||||
| Gm24504 | 2.57 | Gm22 | 2.03 | ||||
| Gm25982 | 2.14 | Atcayos | 5.91 | ||||
| Gm24678 | 2.03 | 3.88 | Gm10790 | 5.16 | |||
| Gm24682 | 3.65 | Chn1os3 | 3.42 | ||||
| Snhg7 | 11.93 | 5730420D15Rik | 3.27 | ||||
| Snord66 * | 70.95 | Gm16793 | 3.2 | ||||
| Gm23546 | 66.56 | Gm10390 | 3.07 | ||||
| Gm25777 | 36.23 | Arhgap33os | 3 | ||||
| Gm25788 | 29.72 | B230312C02Rik | 2.98 | ||||
| Gm23734 | 18.87 | 4632428C04Rik | 2.96 | ||||
| Gm24916 | 11.99 | 4933424G05Rik | 2.65 | ||||
| Gm22144 | 11.18 | Gm19784 | 2.59 | ||||
| Gm24771 | 11.03 | Gm12603 * | 2.59 | ||||
| Gm22504 | 8.42 | D5Ertd605e | 2.54 | ||||
| Snord14a * | 7.54 | 9330102E08Rik | 2.51 | ||||
| Gm23527 | 5.66 | 9230105E05Rik | 2.43 | ||||
| Gm26148 | 4.58 | 9630013K17Rik | 2.43 | ||||
| Gm25376 | 4.27 | 4933432I03Rik | 2.36 | ||||
| Gm25432 | 3.18 | 4930565D16Rik | 2.35 | ||||
| Gm25715 | 3.1 | Gm16548 | 2.33 | ||||
| Gm23534 | 2.94 | Gm13985 | 2.31 | ||||
| Gm26047 | 2.88 | Gm10007 | 2.24 | ||||
| Gm25526 | 2.81 | Gm10619 | 2.23 | ||||
| Rnu3a | 2.76 | 4930568E12Rik | 2.2 | ||||
| Snord53 * | 2.72 | C130080G10Rik * | 2.19 | ||||
| Gm24127 | 2.67 | 4930522O17Rik | 2.19 | ||||
| Gm24581 | 2.26 | Gm13003 | 2.14 | ||||
| Gm24648 | 2.23 | 4930444M15Rik | 2.12 | ||||
| Gm25945 | 2.17 | 4931403G20Rik | 2.11 | ||||
| Gm23136 | 2.16 | 4930488L21Rik | 2.11 | ||||
| Gm23129 | 2.13 | 1700113A16Rik | 2.1 | ||||
| Gm24252 | 2.13 | Gm5144 | 2.09 | ||||
| Gm22840 | 2.11 | Lincred1 * | 2.09 | ||||
| Gm24313 | 2.07 | 4933433G08Rik | 2.08 | ||||
| Gm22269 | 2.05 | A930019D19Rik | 2.07 | ||||
| AF357425 * | −22.59 | 1700047A11Rik | 2.07 | ||||
| ScaRNA15 | 8.54 | 2500002B13Rik | 2.07 | ||||
| Gm24668 | 167.27 | 4930455F16Rik | 2.05 | ||||
| Gm22940 | 75.5 | Hoxaas3 * | 2.05 | ||||
| Gm23970 | 14.37 | 1700045H11Rik | 2.04 | ||||
| Gm23000 | 4.21 | Gm15413 | 2.04 | ||||
| Gm22883 | 2.56 | 1700064J06Rik | 2.02 | ||||
| Gm23031 | 2.21 | 5430434I15Rik | 2.02 | ||||
| Gm24524 | 2.03 | 1700066N21Rik | 2.01 | ||||
| Gm23119 | 63.09 | C530044C16Rik | 2.01 | ||||
| Snhg7 * | 64.98 | ||||||
| Gm12590 | 28.43 | ||||||
| Gm26906 | 4.55 | ||||||
| Gm26675 | 4.4 | ||||||
| Gm6410 | 3.92 | ||||||
| Gm9898 | 2.58 | ||||||
| Gm10425 | 2.45 | ||||||
| Gm13790 | 2.42 | ||||||
| Gm28890 | 2.36 | ||||||
| Gm26542 | 2.26 | ||||||
| Gm16295 | 2.19 | ||||||
| Gm14684 | 2.17 | ||||||
| Gm26656 | 2.1 | ||||||
| Gm15556 | 2.07 | ||||||
| Gm12148 | 2.05 | ||||||
| Gm12637 | 2.05 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nuthikattu, S.; Milenkovic, D.; Rutledge, J.C.; Villablanca, A.C. Lipotoxic Injury Differentially Regulates Brain Microvascular Gene Expression in Male Mice. Nutrients 2020, 12, 1771. https://doi.org/10.3390/nu12061771
Nuthikattu S, Milenkovic D, Rutledge JC, Villablanca AC. Lipotoxic Injury Differentially Regulates Brain Microvascular Gene Expression in Male Mice. Nutrients. 2020; 12(6):1771. https://doi.org/10.3390/nu12061771
Chicago/Turabian StyleNuthikattu, Saivageethi, Dragan Milenkovic, John C. Rutledge, and Amparo C. Villablanca. 2020. "Lipotoxic Injury Differentially Regulates Brain Microvascular Gene Expression in Male Mice" Nutrients 12, no. 6: 1771. https://doi.org/10.3390/nu12061771
APA StyleNuthikattu, S., Milenkovic, D., Rutledge, J. C., & Villablanca, A. C. (2020). Lipotoxic Injury Differentially Regulates Brain Microvascular Gene Expression in Male Mice. Nutrients, 12(6), 1771. https://doi.org/10.3390/nu12061771