Effects of L-Phenylalanine on Energy Intake and Glycaemia—Impacts on Appetite Perceptions, Gastrointestinal Hormones and Gastric Emptying in Healthy Males




Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Study Design
2.3. Study Treatments
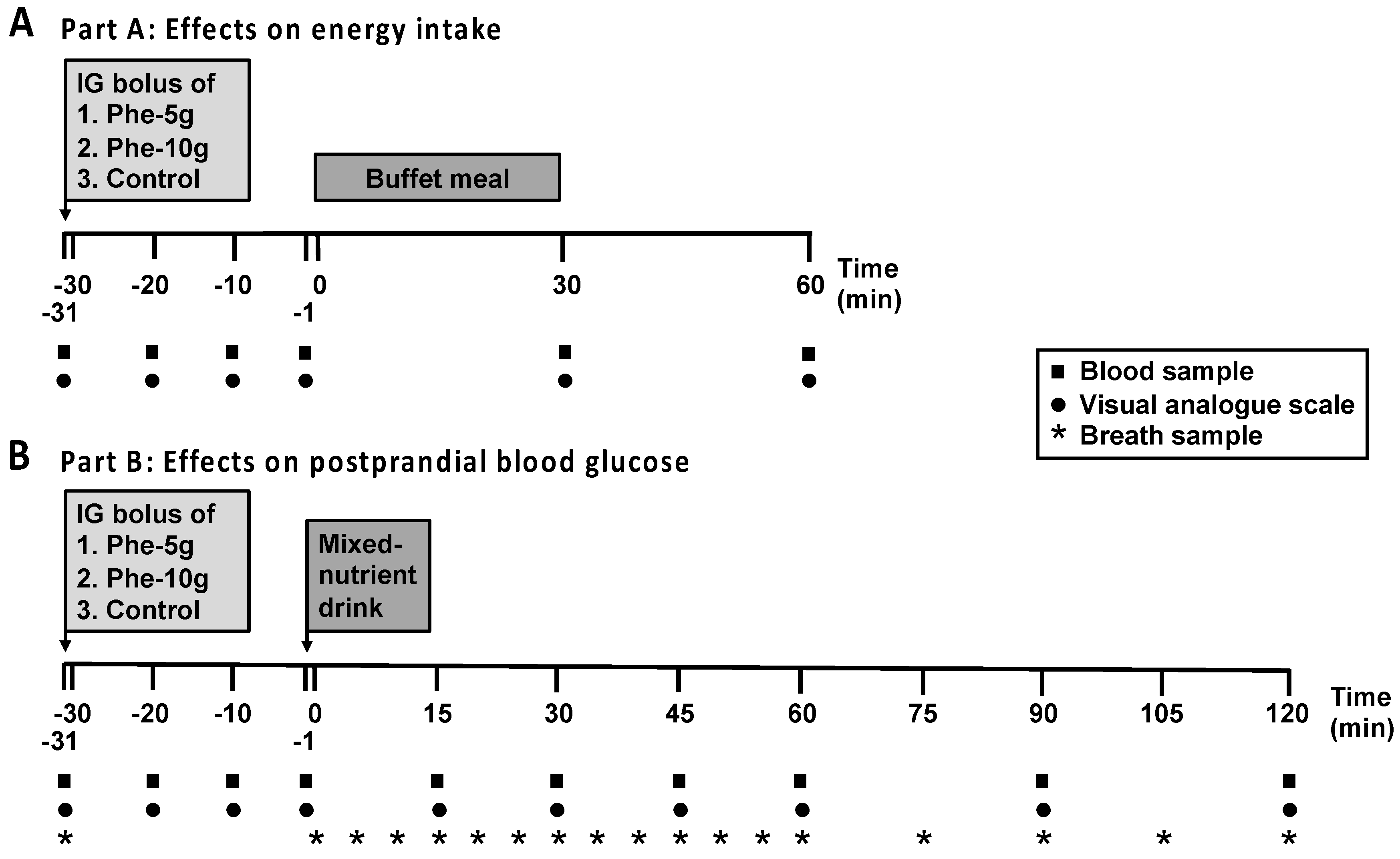
2.4. Study Protocol
2.5. Measurements
2.5.1. Plasma Hormone and Glucose Analyses
2.5.2. Measurement of Gastric Emptying
2.5.3. Appetite Perceptions, GI Symptoms and Energy Intake
2.6. Data and Statistical Analyses
3. Results
3.1. Part A: Effects on Energy Intake
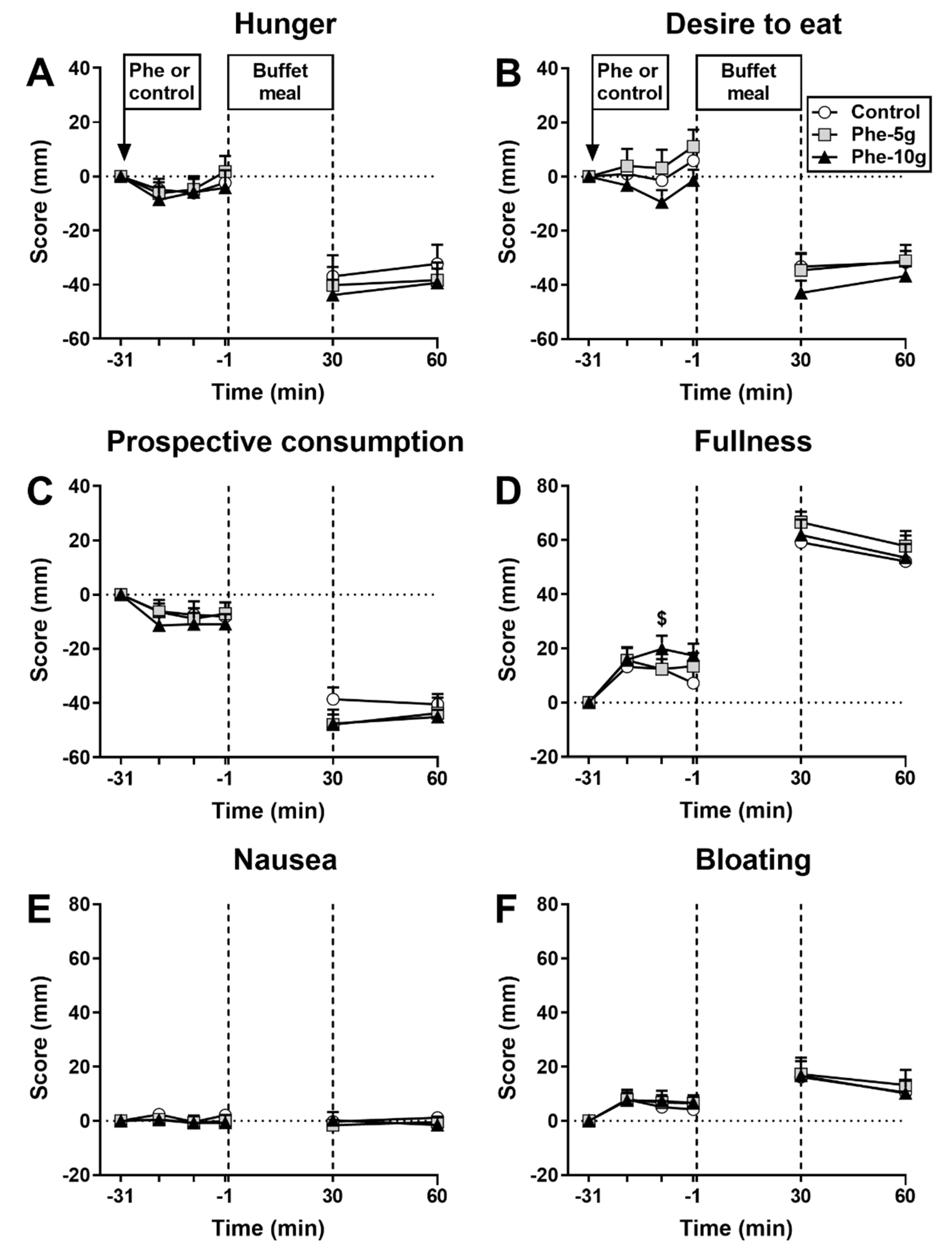
3.1.1. Appetite Perceptions and GI Symptoms
3.1.2. Energy Intake
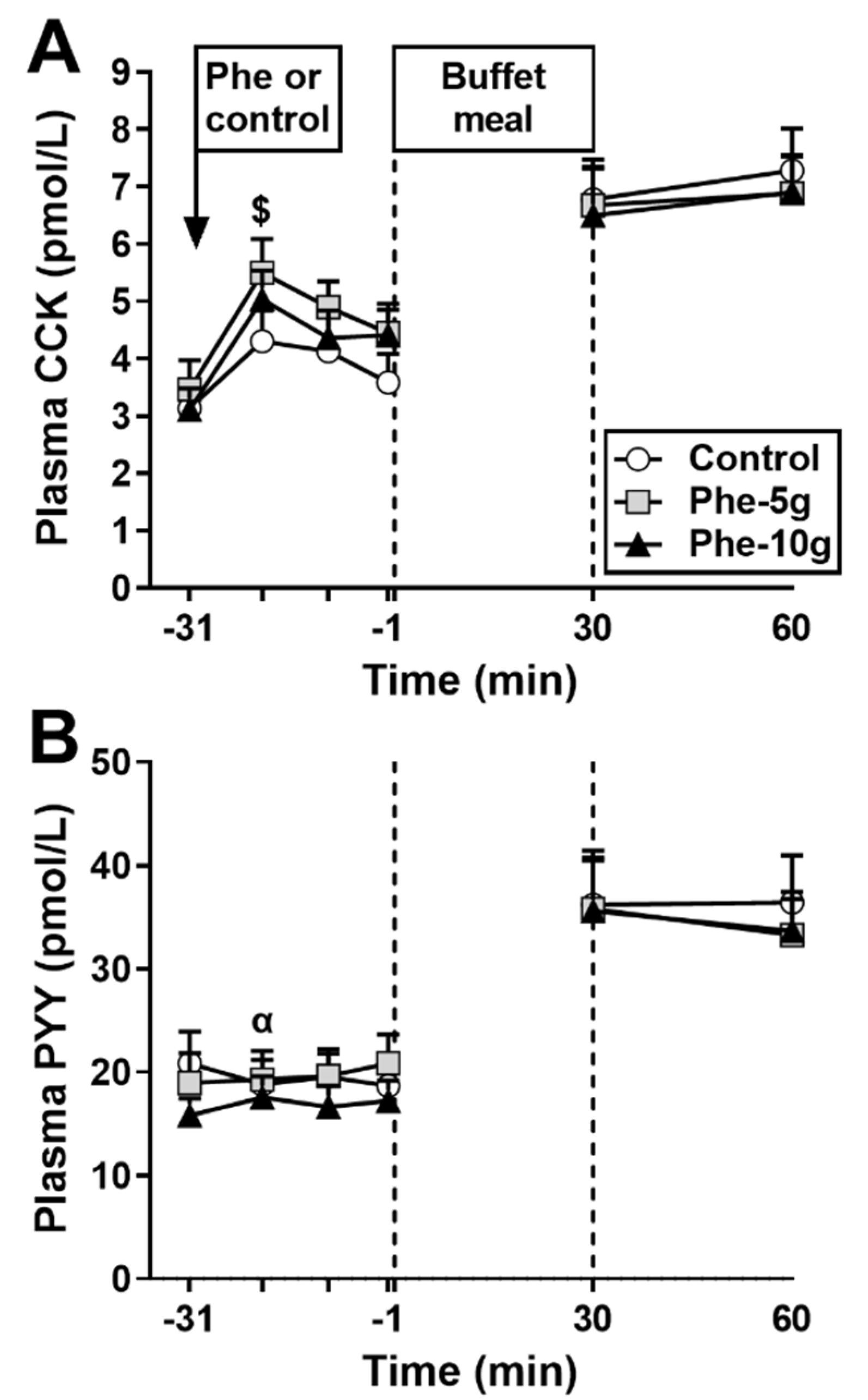
3.1.3. Plasma CCK
3.1.4. Plasma PYY
3.1.5. Relationships between Dose of Phenylalanine, Energy Intake, Plasma CCK, Plasma PYY and Appetite Perceptions
3.2. Part B: Effects on Glycaemia
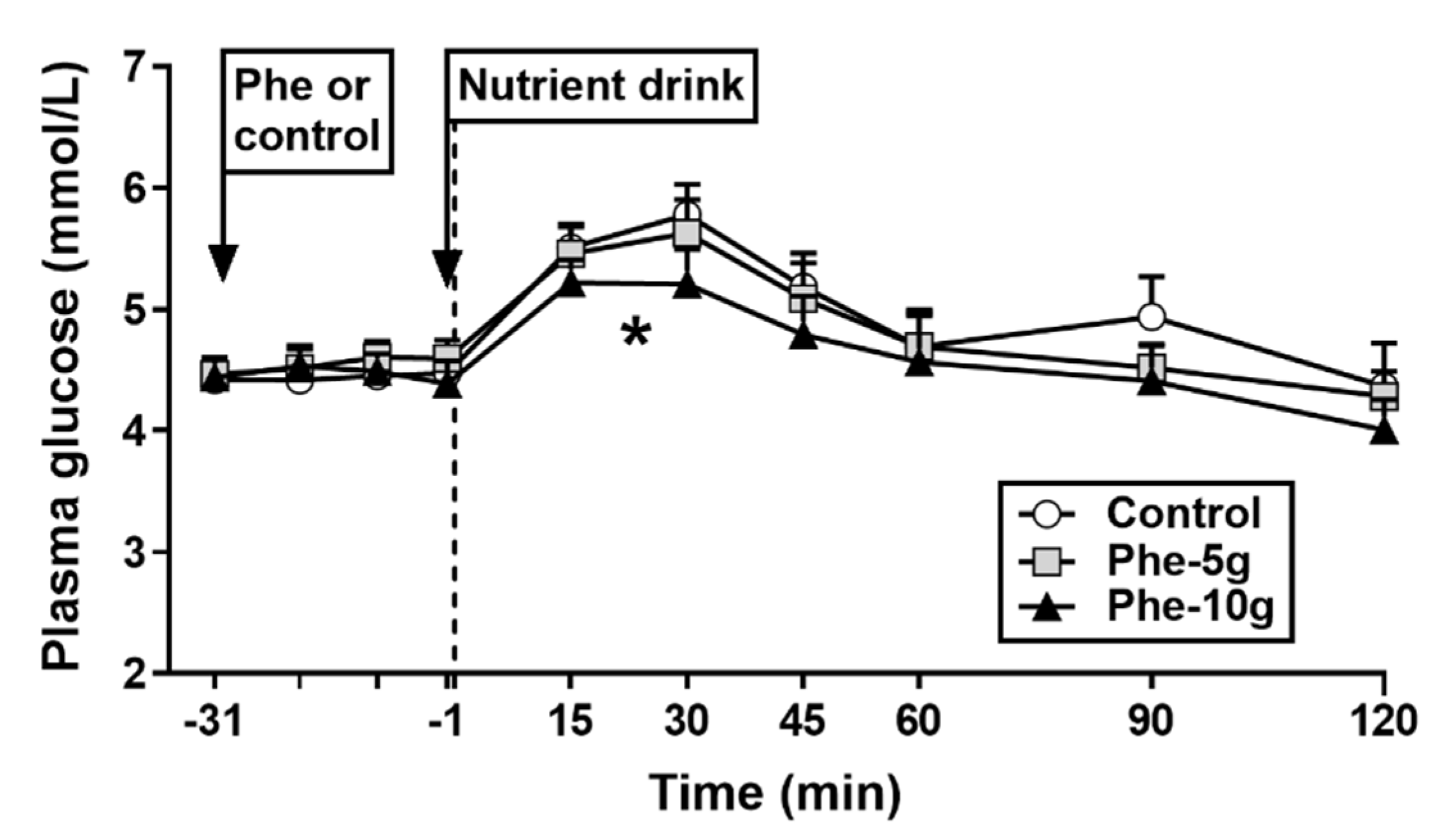
3.2.1. Plasma Glucose
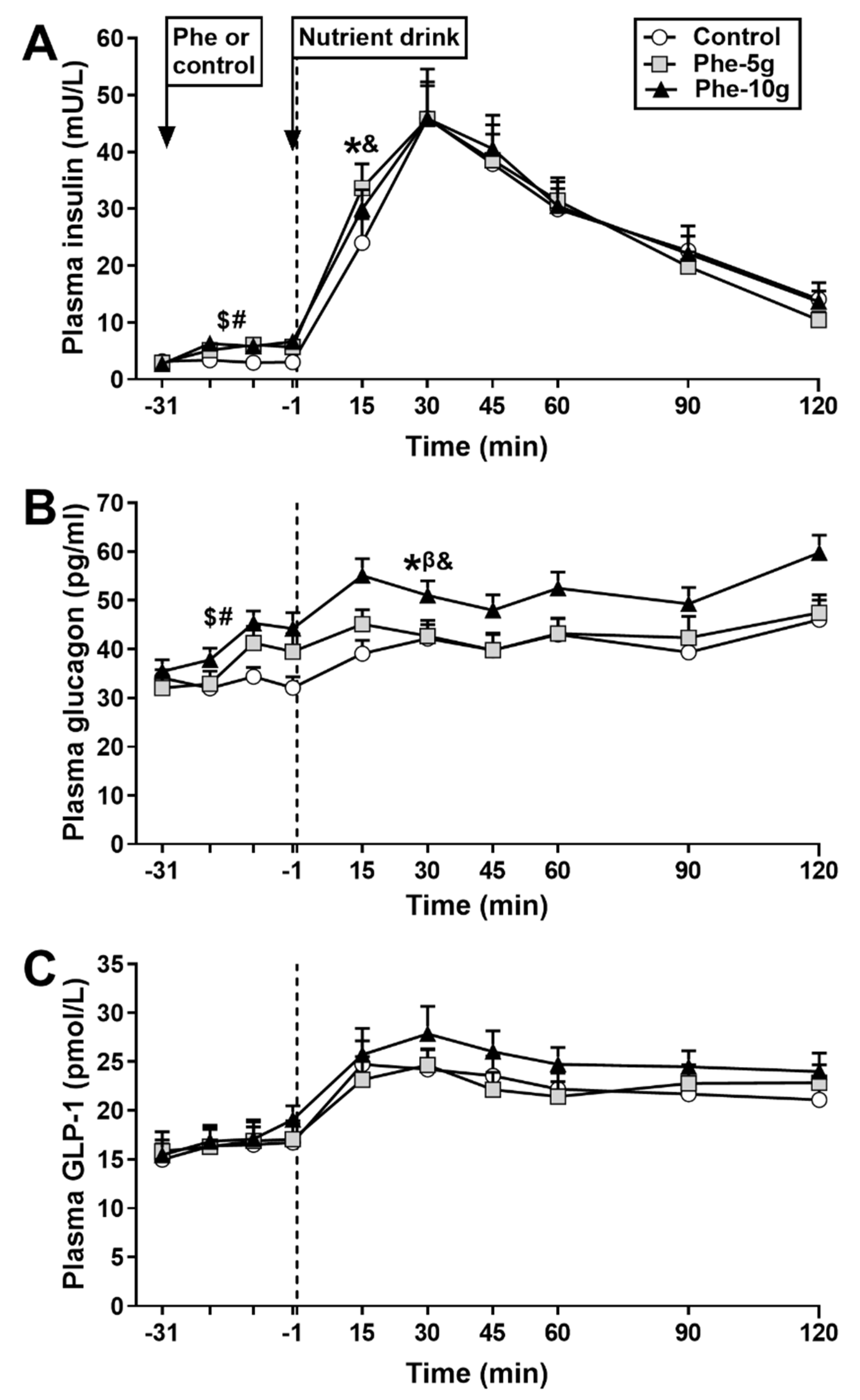
3.2.2. Plasma Insulin
3.2.3. Plasma Glucagon
3.2.4. Plasma GLP-1
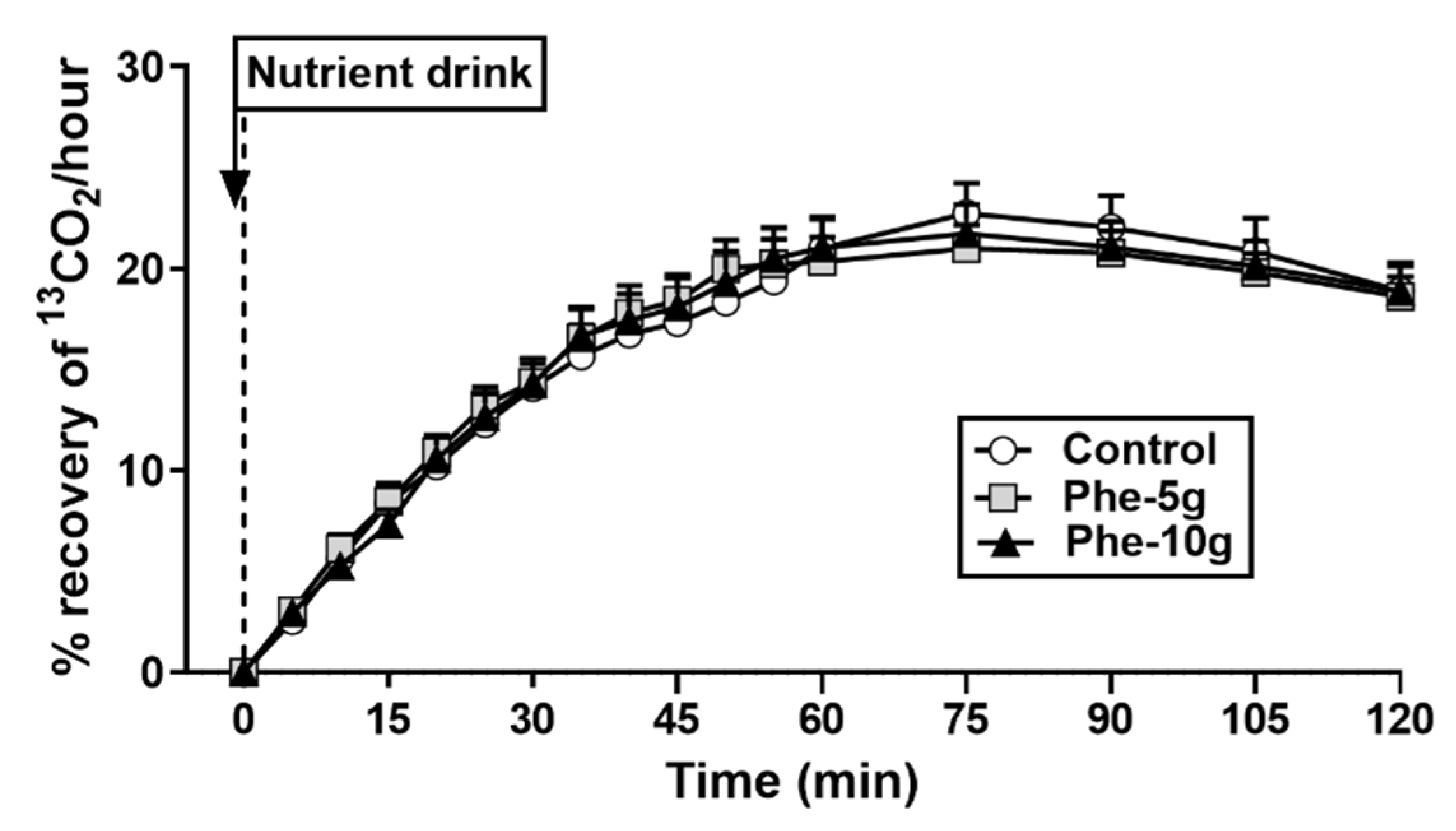
3.2.5. Gastric Emptying
3.2.6. Relationships between Dose of Phenylalanine, Peak Plasma Glucose, Insulin, Glucagon and GLP-1
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Steinert, R.E.; Feinle-Bisset, C.; Asarian, L.; Horowitz, M.; Beglinger, C.; Geary, N. Ghrelin, CCK, GLP-1, and PYY(3–36): Secretory controls and physiological roles in eating and glycemia in health, obesity, and after RYGB. Physiol. Rev. 2017, 97, 411–463. [Google Scholar] [CrossRef] [Green Version]
- Seimon, R.V.; Lange, K.; Little, T.J.; Brennan, I.M.; Pilichiewicz, A.N.; Feltrin, K.L.; Smeets, A.J.; Horowitz, M.; Feinle-Bisset, C. Pooled-data analysis identifies pyloric pressures and plasma cholecystokinin concentrations as major determinants of acute energy intake in healthy, lean men. Am. J. Clin. Nutr. 2010, 92, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Liddle, R.A.; Morita, E.T.; Conrad, C.K.; Williams, J.A. Regulation of gastric emptying in humans by cholecystokinin. J. Clin. Investig. 1986, 77, 992–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajishafiee, M.; Bitarafan, V.; Feinle-Bisset, C. Gastrointestinal sensing of meal-related signals in humans, and dysregulations in eating-related disorders. Nutrients 2019, 11, 1298. [Google Scholar] [CrossRef] [Green Version]
- Drucker, D.J. The biology of incretin hormones. Cell Metab. 2006, 3, 153–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhavan, T.; Luhovyy, B.L.; Brown, P.H.; Cho, C.E.; Anderson, G.H. Effect of premeal consumption of whey protein and its hydrolysate on food intake and postmeal glycemia and insulin responses in young adults. Am. J. Clin. Nutr. 2010, 91, 966–975. [Google Scholar] [CrossRef]
- Ma, J.; Stevens, J.E.; Cukier, K.; Maddox, A.F.; Wishart, J.M.; Jones, K.L.; Clifton, P.M.; Horowitz, M.; Rayner, C.K. Effects of a protein preload on gastric emptying, glycemia, and gut hormones after a carbohydrate meal in diet-controlled type 2 diabetes. Diabetes Care 2009, 32, 1600–1602. [Google Scholar] [CrossRef] [Green Version]
- Ryan, A.T.; Feinle-Bisset, C.; Kallas, A.; Wishart, J.M.; Clifton, P.M.; Horowitz, M.; Luscombe-Marsh, N.D. Intraduodenal protein modulates antropyloroduodenal motility, hormone release, glycemia, appetite, and energy intake in lean men. Am. J. Clin. Nutr. 2012, 96, 474–482. [Google Scholar] [CrossRef] [Green Version]
- Ballinger, A.B.; Clark, M.L. L-phenylalanine releases cholecystokinin (CCK) and is associated with reduced food intake in humans: Evidence for a physiological role of CCK in control of eating. Metabolism 1994, 43, 735–738. [Google Scholar] [CrossRef]
- Steinert, R.E.; Landrock, M.F.; Horowitz, M.; Feinle-Bisset, C. Effects of intraduodenal infusions of L-phenylalanine and L-glutamine on antropyloroduodenal motility and plasma cholecystokinin in healthy men. J. Neurogastroenterol. Motil. 2015, 21, 404–413. [Google Scholar] [CrossRef] [Green Version]
- Nuttall, F.Q.; Schweim, K.J.; Gannon, M.C. Effect of orally administered phenylalanine with and without glucose on insulin, glucagon and glucose concentrations. Horm. Metab. Res. 2006, 38, 518–523. [Google Scholar] [CrossRef]
- Alamshah, A.; Spreckley, E.; Norton, M.; Kinsey-Jones, J.S.; Amin, A.; Ramgulam, A.; Cao, Y.; Johnson, R.; Saleh, K.; Akalestou, E.; et al. L-phenylalanine modulates gut hormone release and glucose tolerance, and suppresses food intake through the calcium-sensing receptor in rodents. Int. J. Obes. 2017, 41, 1693–1701. [Google Scholar] [CrossRef] [Green Version]
- Stunkard, A.J.; Messick, S. The three-factor eating questionnaire to measure dietary restraint, disinhibition and hunger. J. Psychosom. Res. 1985, 29, 71–83. [Google Scholar] [CrossRef]
- Nair, N.; Brennan, I.; Little, T.; Gentilcore, D.; Hausken, T.; Jones, K.; Wishart, J.M.; Horowitz, M.; Feinle-Bisset, C. Reproducibility of energy intake, gastric emptying, blood glucose, plasma insulin and cholecystokinin responses in healthy young males. Br. J. Nutr. 2008, 101, 1094–1102. [Google Scholar] [CrossRef] [Green Version]
- Brennan, I.M.; Feltrin, K.L.; Nair, N.S.; Hausken, T.; Little, T.J.; Gentilcore, D.; Wishart, J.M.; Jones, K.L.; Horowitz, M.; Feinle-Bisset, C. Effects of the phases of the menstrual cycle on gastric emptying, glycemia, plasma GLP-1 and insulin, and energy intake in healthy lean women. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G602–G610. [Google Scholar] [CrossRef] [Green Version]
- Mossi, S.; Meyer-Wyss, B.; Beglinger, C.; Schwizer, W.; Fried, M.; Ajami, A.; Brignoli, R. Gastric emptying of liquid meals measured noninvasively in humans with [13C]Acetate breath test. Dig. Dis. Sci. 1994, 39, 107S–109S. [Google Scholar] [CrossRef]
- Santangelo, A.; Peracchi, M.; Conte, D.; Fraquelli, M.; Porrini, M. Physical state of meal affects gastric emptying, cholecystokinin release and satiety. Br. J. Nutr. 1998, 80, 521–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullrich, S.S.; Fitzgerald, P.C.; Schober, G.; Steinert, R.E.; Horowitz, M.; Feinle-Bisset, C. Intragastric administration of leucine or isoleucine lowers the blood glucose response to a mixed-nutrient drink by different mechanisms in healthy, lean volunteers. Am. J. Clin. Nutr. 2016, 104, 1274–1284. [Google Scholar] [CrossRef] [Green Version]
- Parker, B.A.; Sturm, K.; MacIntosh, C.G.; Feinle, C.; Horowitz, M.; Chapman, I.M. Relation between food intake and visual analogue scale ratings of appetite and other sensations in healthy older and young subjects. Eur. J. Clin. Nutr. 2004, 58, 212–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinert, R.E.; Luscombe-Marsh, N.D.; Little, T.J.; Standfield, S.; Otto, B.; Horowitz, M.; Feinle-Bisset, C. Effects of intraduodenal infusion of L-tryptophan on ad Libitum eating, antropyloroduodenal motility, glycemia, insulinemia, and gut peptide secretion in healthy men. J. Clin. Endocrinol. Metab. 2014, 99, 3275–3284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinert, R.E.; Landrock, M.F.; Ullrich, S.S.; Standfield, S.; Otto, B.; Horowitz, M.; Feinle-Bisset, C. Effects of intraduodenal infusion of the branched-chain amino acid leucine on ad libitum eating, gut motor and hormone functions, and glycemia in healthy men. Am. J. Clin. Nutr. 2015, 102, 820–827. [Google Scholar] [CrossRef]
- Degen, L.; Oesch, S.; Casanova, M.; Graf, S.; Ketterer, S.; Drewe, J.; Beglinger, C. Effect of peptide YY3–36 on food intake in humans. Gastroenterology 2005, 129, 1430–1436. [Google Scholar] [CrossRef] [PubMed]
- Gutzwiller, J.P.; Göke, B.; Drewe, J.; Hildebrand, P.; Ketterer, S.; Handschin, D.; Winterhalder, R.; Conen, D.; Beglinger, C. Glucagon-like peptide-1: A potent regulator of food intake in humans. Gut 1999, 44, 81–86. [Google Scholar] [CrossRef]
- Fried, M.; Mayer, E.A.; Jansen, J.B.M.J.; Lamers, C.B.H.W.; Taylor, I.L.; Bloom, S.R.; Meyer, J.H. Temporal relationships of cholecystokinin release, pancreatobiliary secretion, and gastric emptying of a mixed meal. Gastroenterology 1988, 95, 1344–1350. [Google Scholar] [CrossRef]
- Flint, A.; Gregersen, N.T.; Gluud, L.L.; Møller, B.K.; Raben, A.; Tetens, I.; Verdich, C.; Astrup, A. Associations between postprandial insulin and blood glucose responses, appetite sensations and energy intake in normal weight and overweight individuals: A meta-analysis of test meal studies. Br. J. Nutr. 2007, 98, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Bagger, J.I.; Holst, J.J.; Hartmann, B.; Andersen, B.; Knop, F.K.; Vilsbøll, T. Effect of oxyntomodulin, glucagon, GLP-1, and combined glucagon +GLP-1 infusion on food intake, appetite, and resting energy expenditure. J. Clin. Endocrinol. Metab. 2015, 100, 4541–4552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fried, M.; Erlacher, U.R.S.; Schwizer, W.; Löchner, C.; Koerfer, J.; Beglinger, C.; Jansen, J.B.; Lamers, C.B.; Harder, F.; Bischof-Delaloye, A. Role of cholecystokinin in the regulation of gastric emptying and pancreatic enzyme secretion in humans: Studies with the cholecystokinin-receptor antagonist loxiglumide. Gastroenterology 1991, 101, 503–511. [Google Scholar] [CrossRef]
- Schwanstecher, C.; Meyer, M.; Schwanstecher, M.; Panten, U. Interaction of N-benzoyl-D-phenylalanine and related compounds with the sulphonylurea receptor of β-cells. Br. J. Pharmacol. 1998, 123, 1023–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nauck, M.A.; Kleine, N.; Ørskov, C.; Holst, J.J.; Willms, B.; Creutzfeldt, W. Normalization of fasting hyperglycaemia by exogenous glucagon-like peptide 1 (7-36 amide) in type 2 (non-insulin-dependent) diabetic patients. Diabetologia 1993, 36, 741–744. [Google Scholar] [CrossRef] [Green Version]
- Horowitz, M.; Edelbroek, M.A.L.; Wishart, J.M.; Straathof, J.W. Relationship between oral glucose tolerance and gastric emptying in normal healthy subjects. Diabetologia 1993, 36, 857–862. [Google Scholar] [CrossRef]
- Marathe, C.S.; Horowitz, M.; Trahair, L.G.; Wishart, J.M.; Bound, M.; Lange, K.; Rayner, C.K.; Jones, K.L. Relationships of early and late glycemic responses with gastric emptying during an oral glucose tolerance test. J. Clin. Endocrinol. Metab. 2015, 100, 3565–3571. [Google Scholar] [CrossRef] [Green Version]
- Rolls, B.J.; Kim-Harris, S.; Fischman, M.W.; Foltin, R.W.; Moran, T.H.; Stoner, S.A. Satiety after preloads with different amounts of fat and carbohydrate: Implications for obesity. Am. J. Clin. Nutr. 1994, 60, 476–487. [Google Scholar] [CrossRef]
- Hall, W.L.; Millward, D.J.; Rogers, P.J.; Morgan, L.M. Physiological mechanisms mediating aspartame-induced satiety. Physiol. Behav. 2003, 78, 557–562. [Google Scholar] [CrossRef]
- Hardman, C.A.; Herbert, V.M.; Brunstrom, J.M.; Munafò, M.R.; Rogers, P.J. Dopamine and food reward: Effects of acute tyrosine/phenylalanine depletion on appetite. Physiol. Behav. 2012, 105, 1202–1207. [Google Scholar] [CrossRef]
- Montgomery, A.J.; McTavish, S.F.; Cowen, P.J.; Grasby, P.M. Reduction of brain dopamine concentration with dietary tyrosine plus phenylalanine depletion: An [11C] raclopride PET study. Am. J. Psychiatry. 2003, 160, 1887–1889. [Google Scholar] [CrossRef]
- Meyer-Gerspach, A.C.; Häfliger, S.; Meili, J.; Doody, A.; Rehfeld, J.F.; Drewe, J.; Beglinger, C.; Wölnerhanssen, B. Effect of L-tryptophan and L-leucine on gut hormone secretion, appetite feelings and gastric emptying rates in lean and non-diabetic obese participants: A randomized, double-blind, parallel-group trial. PLoS ONE 2016, 11, e0166758. [Google Scholar] [CrossRef] [Green Version]
- Kalogeropoulou, D.; Lafave, L.; Schweim, K.; Gannon, M.C.; Nuttall, F.Q. Leucine, when ingested with glucose, synergistically stimulates insulin secretion and lowers blood glucose. Metabolism 2008, 57, 1747–1752. [Google Scholar] [CrossRef]
- Gannon, M.C.; Nuttall, F.Q. Amino acid ingestion and glucose metabolism—A review. IUBMB Life 2010, 62, 660–668. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Control | Phe-5 g | Phe-10 g | p |
|---|---|---|---|---|
| Energy intake (kcal) | 1084 ± 76 | 987 ± 82 | 900 ± 78 * | 0.005 |
| Amount consumed (g) | 1096 ± 87 | 1065 ± 93 | 988 ± 78 | NS |
| Variables | Control | Phe-5 g | Phe-10 g | p (Mixed Models Analysis) |
|---|---|---|---|---|
| PART A: Effects on Energy Intake Plasma CCK | ||||
| AUC−31 to −1, pmol/L × min | 121 ± 5 | 137 ± 6 | 135 ± 6 * | 0.002 |
| Plasma PYY | ||||
| AUC−31 to −1, pmol/L × min | 521 ± 24 | 577 ± 16 γ | 578 ± 30 | 0.057 |
| PART B: Effects on glycaemia Plasma glucose | ||||
| AUC−31 to −1, mmol/L × min | 134 ± 2 | 136 ± 2 | 134 ± 3 | NS |
| AUC−1 to 120, mmol/L × min | 601 ± 25 | 581 ± 20 | 556 ± 28 * | 0.027 |
| AUC−1 to 30, mmol/L × min | 160 ± 4 | 158 ± 4 | 150 ± 4 | 0.095 |
| Peak glucose, mmol/L | 6.1 ± 0.2 | 5.9 ± 0.2 | 5.5 ± 0.3 * | 0.021 |
| Plasma insulin | ||||
| AUC−31 to −1, mU/L × min | 89 ± 5 | 155 ± 16 * | 173 ± 18 * | 0.000 |
| AUC−1 to 120, mU/L × min | 3183 ± 375 | 3270 ± 334 | 3372 ± 293 | NS |
| AUC−1 to 30, mU/L × min 2 | 6.4 ± 0.2 | 6.7 ± 0.1 * | 6.6 ± 0.1 * | 0.006 |
| Plasma glucagon | ||||
| AUC−31 to −1, pg/mL × min | 989 ± 38 | 1139 ± 51 * | 1196 ± 39 * | 0.000 |
| AUC −1 to 120, pg/mL × min | 4879 ± 342 | 5282 ± 329 § | 6100 ± 304 * | 0.002 |
| AUC−1 to 30, pg/ mL × min | 1138 ± 61 | 1334 ± 67 * | 1507 ± 76 * | 0.000 |
| Plasma GLP-1 | ||||
| AUC−31 to −1, pmol/L × min | 499 ± 30 | 484 ± 16 | 511 ± 23 | NS |
| AUC−1 to 120, pmol/L × min | 2703 ± 157 | 2662 ± 151 | 2986 ± 163 | 0.087 |
| AUC−1 to 30, pmol/L × min | 688 ± 44 | 651 ± 31 | 737 ± 48 | 0.088 |
| Gastric emptying | ||||
| AUC0 to 120, % recovery of 13CO2 | 2032 ± 138 | 2021 ± 103 | 2021 ± 126 | NS |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fitzgerald, P.C.E.; Manoliu, B.; Herbillon, B.; Steinert, R.E.; Horowitz, M.; Feinle-Bisset, C. Effects of L-Phenylalanine on Energy Intake and Glycaemia—Impacts on Appetite Perceptions, Gastrointestinal Hormones and Gastric Emptying in Healthy Males. Nutrients 2020, 12, 1788. https://doi.org/10.3390/nu12061788
Fitzgerald PCE, Manoliu B, Herbillon B, Steinert RE, Horowitz M, Feinle-Bisset C. Effects of L-Phenylalanine on Energy Intake and Glycaemia—Impacts on Appetite Perceptions, Gastrointestinal Hormones and Gastric Emptying in Healthy Males. Nutrients. 2020; 12(6):1788. https://doi.org/10.3390/nu12061788
Chicago/Turabian StyleFitzgerald, Penelope C. E., Benoit Manoliu, Benjamin Herbillon, Robert E. Steinert, Michael Horowitz, and Christine Feinle-Bisset. 2020. "Effects of L-Phenylalanine on Energy Intake and Glycaemia—Impacts on Appetite Perceptions, Gastrointestinal Hormones and Gastric Emptying in Healthy Males" Nutrients 12, no. 6: 1788. https://doi.org/10.3390/nu12061788
APA StyleFitzgerald, P. C. E., Manoliu, B., Herbillon, B., Steinert, R. E., Horowitz, M., & Feinle-Bisset, C. (2020). Effects of L-Phenylalanine on Energy Intake and Glycaemia—Impacts on Appetite Perceptions, Gastrointestinal Hormones and Gastric Emptying in Healthy Males. Nutrients, 12(6), 1788. https://doi.org/10.3390/nu12061788