In Vitro and In Vivo Antitumor Efficacy of Hizikia fusiforme Celluclast Extract against Bladder Cancer

 , ,
, ,  , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Sulfated Polysaccharides from Hizikia fusiforme (SPHF)
2.3. CHF and SPHF Chemical Analysis
2.4. HF and HFPS Antioxidant Assay
2.5. Cell Culture and Treatment
2.6. Viability Assays
2.7. Cell Cycle Analysis
2.8. Immunoblots and Immunoprecipitation
2.9. Wound-Healing Migration Assays
2.10. Invasion Assays
2.11. Zymography
2.12. Nuclear Extracts and Electrophoretic Mobility Shift Assay
2.13. Animals
2.14. Mouse Xenograft Generation
2.15. Immunohistochemistry
2.16. Plasma Preparation and Biochemical Analysis
2.17. Statistical Analysis
3. Results
3.1. Total Phenolic Content, Sulfated Polysaccharide Content, Fucose Content, and Free Radical Scavenging Activities of CHF and SPHF
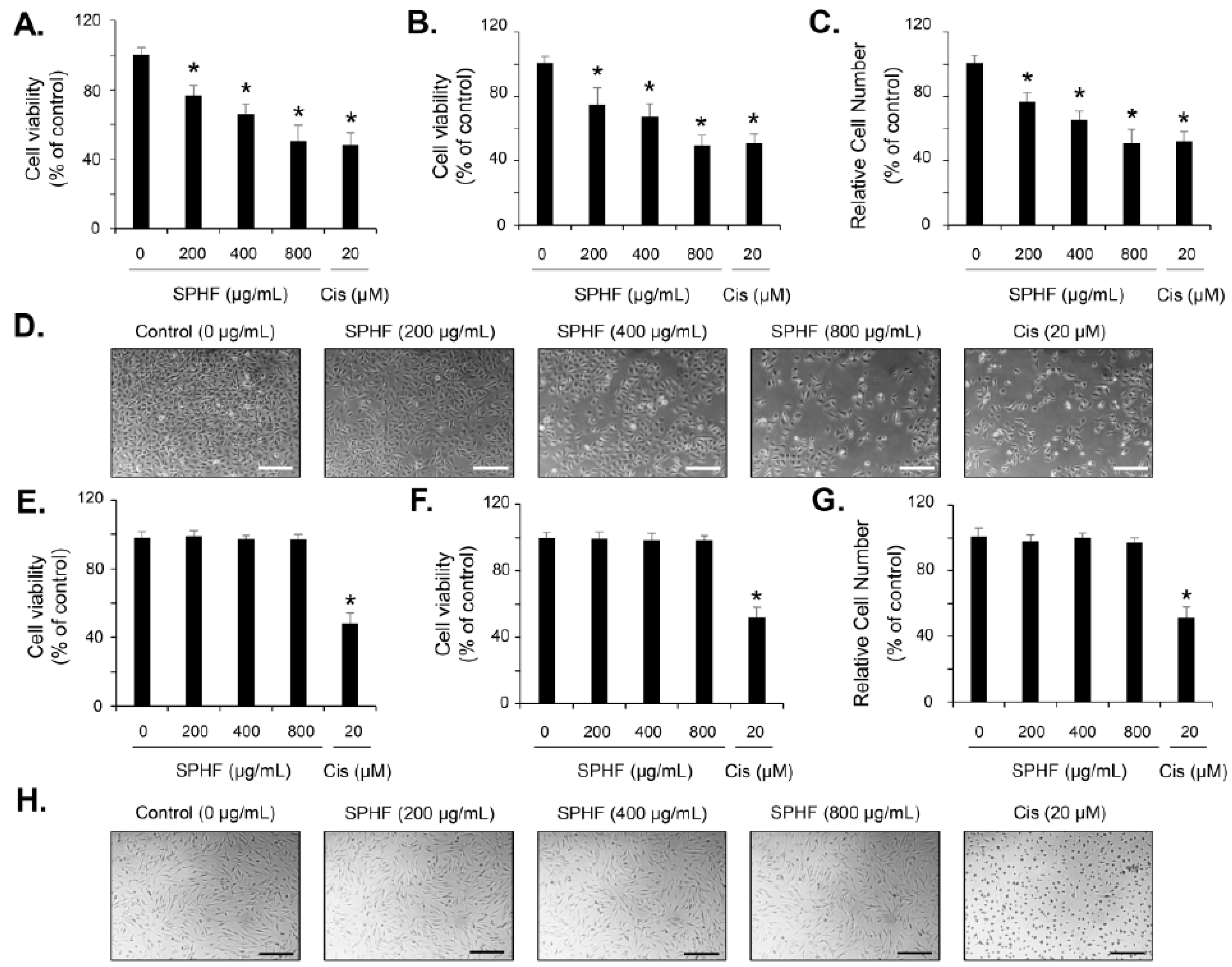
3.2. SPHF Inhibits Bladder Cancer EJ Cell Proliferation via Induction of G1-Phase Cell Cycle Arrest
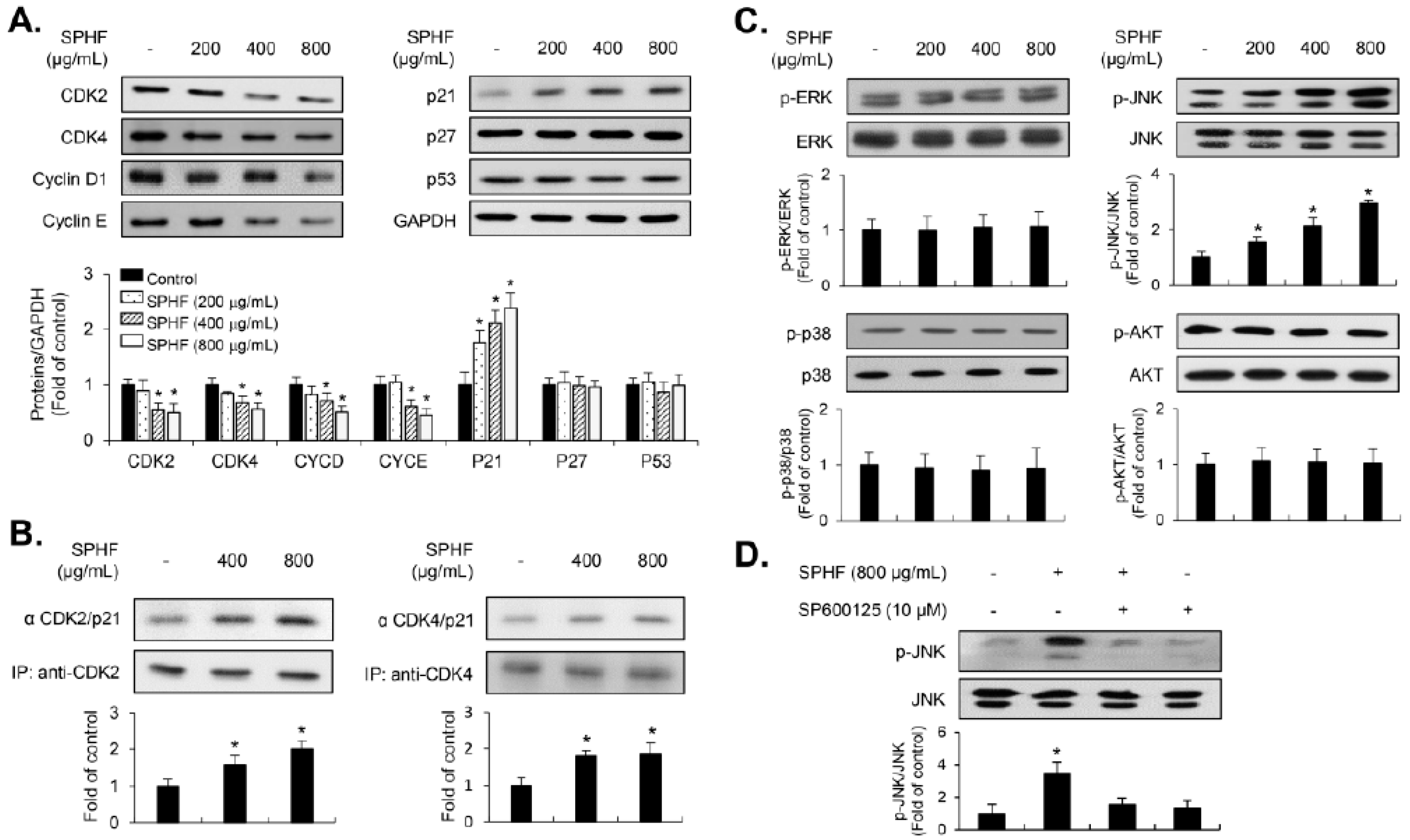
3.3. SPHF-Induced Inhibition of Cell Proliferation was Involved in the p21WAF1-Mediated G1-Phase Cell Cycle Arrest via Decreased Expression of CDKs and Cyclins
3.4. JNK Signaling is Associated with SPHF-Induced Inhibition of EJ Cell Proliferation
3.5. SPHF Inhibits Migration and Invasion of EJ Cells via Decreased MMP-9 Expression Mediated by Suppression of Transcription Factor AP-1, Sp-1, and NF-κB Binding Activity
3.6. SPHF Suppressed EJ Bladder Cancer Cell Xenografted Tumors without Apparent Toxicity
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2017. CA Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barocas, D.A.; Clark, P.E. Bladder Cancer. Curr. Opin. Oncol. 2008, 20, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Brawn, P.N. The Origin of Invasive Carcinoma of the Bladder. Cancer 1982, 50, 515–519. [Google Scholar] [CrossRef]
- Hurst, C.D.; Alder, O.; Platt, F.M.; Droop, A.; Stead, L.F.; Burns, J.E.; Burghel, G.J.; Jain, S.; Klimczak, L.J.; Lindsay, H.; et al. Genomic Subtypes of Non-Invasive Bladder Cancer with Distinct Metabolic Profile and Female Gender Bias in KDM6A Mutation Frequency. Cancer Cell 2017, 32, 701–715.e7. [Google Scholar] [CrossRef] [PubMed]
- Droller, M.J. Bladder Cancer: State-of-the-Art Care. CA Cancer J. Clin. 1998, 48, 269–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, A.; Wszolek, M.F.; Niemierko, A.; Clayman, R.H.; Drumm, M.; Rodríguez, D.; Feldman, A.S.; Dahl, D.M.; Heney, N.M.; Shipley, W.U.; et al. Incidence, Clinicopathological Risk Factors, Management and Outcomes of Nonmuscle Invasive Recurrence After Complete Response to Trimodality Therapy for Muscle Invasive Bladder Cancer. J. Urol. 2018, 199, 407–415. [Google Scholar] [CrossRef]
- Black, P.C.; Dinney, C.P.N. Bladder Cancer Angiogenesis and Metastasis–Translation from Murine Model to Clinical Trial. Cancer Metastasis Rev. 2007, 26, 623–634. [Google Scholar] [CrossRef]
- Knowles, M.A.; Hurst, C.D. Molecular Biology of Bladder Cancer: New Insights into Pathogenesis and Clinical Diversity. Nat. Rev. Cancer 2015, 15, 25–41. [Google Scholar] [CrossRef]
- Gerhardt, D.; Bertola, G.; Dietrich, F.; Figueiró, F.; Zanotto-Filho, A.; Moreira Fonseca, J.C.; Morrone, F.B.; Barrios, C.H.; Battastini, A.M.O.; Salbego, C.G. Boldine Induces Cell Cycle Arrest and Apoptosis in T24 Human Bladder Cancer Cell Line Via Regulation of ERK, AKT, and GSK-3β. Urol. Oncol. 2014, 32, e1–e9. [Google Scholar] [CrossRef]
- Mayer, I.A.; Arteaga, C.L. The PI3K/AKT Pathway as a Target for Cancer Treatment. Annu. Rev. Med. 2016, 67, 11–28. [Google Scholar] [CrossRef]
- Chambard, J.; Lefloch, R.; Pouysségur, J.; Lenormand, P. ERK Implication in Cell Cycle Regulation. Biochim. Biophys. Acta 2007, 1773, 1299–1310. [Google Scholar] [CrossRef]
- Otto, T.; Sicinski, P. Cell Cycle Proteins as Promising Targets in Cancer Therapy. Nat. Rev. Cancer 2017, 17, 93–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeggo, P.A.; Pearl, L.H.; Carr, A.M. DNA Repair, Genome Stability and Cancer: A Historical Perspective. Nat. Rev. Cancer 2016, 16, 35–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, A.G.; Kim, J.; Al-Ahmadie, H.; Bellmunt, J.; Guo, G.; Cherniack, A.D.; Hinoue, T.; Laird, P.W.; Hoadley, K.A.; Akbani, R.; et al. Comprehensive Molecular Characterization of Muscle-Invasive Bladder. Cancer Cell 2017, 171, 540–556.e25. [Google Scholar] [CrossRef] [PubMed]
- Bianco, F.J.; Gervasi, D.C.; Tiguert, R.; Grignon, D.J.; Pontes, J.E.; Crissman, J.D.; Fridman, R.; Wood, D.P. Matrix Metalloproteinase-9 Expression in Bladder Washes from Bladder Cancer Patients Predicts Pathological Stage and Grade. Clin. Cancer Res. 1998, 4, 3011–3016. [Google Scholar]
- Davies, B.; Waxman, J.; Wasan, H.; Abel, P.; Williams, G.; Krausz, T.; Neal, D.; Thomas, D.; Hanby, A.; Balkwill, F. Levels of Matrix Metalloproteases in Bladder Cancer Correlate with Tumor Grade and Invasion. Cancer Res. 1993, 53, 5365–5369. [Google Scholar]
- Bond, M.; Fabunmi, R.P.; Baker, A.H.; Newby, A.C. Synergistic Upregulation of Metalloproteinase-9 by Growth Factors and Inflammatory Cytokines: An Absolute Requirement for Transcription Factor NF-Kappa B. FEBS Lett. 1998, 435, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Sato, H.; Seiki, M. Regulatory Mechanism of 92 kDa Type IV Collagenase Gene Expression which is Associated with Invasiveness of Tumor Cells. Oncogene 1993, 8, 395–405. [Google Scholar]
- Lee, S.; Cho, S.; Lee, E.; Kim, S.; Lee, S.; Lim, J.; Choi, Y.H.; Kim, W.; Moon, S. Interleukin-20 Promotes Migration of Bladder Cancer Cells through Extracellular Signal-Regulated Kinase (ERK)-Mediated MMP-9 Protein Expression Leading to Nuclear Factor (NF-κB) Activation by Inducing the Up-Regulation of p21(WAF1) Protein Expression. J. Biol. Chem. 2013, 288, 5539–5552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Lee, W.; Oh, J.Y.; Cui, Y.R.; Ryu, B.; Jeon, Y. Protective Effect of Sulfated Polysaccharides from Celluclast-Assisted Extract of Hizikia Fusiforme Against Ultraviolet B-induced Skin Damage by Regulating NF-κB, AP-1, and MAPKs Signaling Pathways in Vitro in Human Dermal Fibroblasts. Mar. Drugs 2018, 16, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, Y.T.; Baek, S.H.; Jeong, S.C.; Yoon, Y.D.; Kim, O.H.; Oh, B.C.; Jung, J.W.; Kim, J.H. Osteoprotective Effects of Polysaccharide-Enriched Hizikia Fusiforme Processing Byproduct in Vitro and in Vivo Models. J. Med. Food 2016, 19, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Hwang, E.; Shin, Y.; Lee, D.; Yang, J.; Park, J.; Yi, T. Immunostimulatory Effect of Enzyme-Modified Hizikia fusiformein a Mouse Model in Vitro and Ex Vivo. Mar. Biotechnol. 2017, 19, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.; Lim, S. Effect of Hizikia fusiformis Extracts on Reactive Oxygen Species Mediated Oxidative Damage. Int. J. Adv. Res. Biol. Sci. 2017, 4, 120–126. [Google Scholar] [CrossRef]
- Oh, J.; Kim, J.; Lee, Y. Anti-Inflammatory and Anti-Diabetic Effects of Brown Seaweeds in High-Fat Diet-Induced Obese Mice. Nutr. Res. Pract. 2016, 10, 42–48. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Kim, E.; Kang, I.; Lee, M.; Lee, Y. Anti-Diabetic Effects and Anti-Inflammatory Effects of Laminaria japonica and Hizikia fusiforme in Skeletal Muscle: In Vitro and in Vivo Model. Nutrients 2018, 10, 491. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.; Jin, C.; Kim, G.; Choi, I.; Jeong, Y.K.; Nam, T.; Kim, S.; Choi, Y.H. Ethyl Alcohol Extracts of Hizikia Fusiforme Sensitize AGS Human Gastric Adenocarcinoma Cells to Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand-Mediated Apoptosis. J. Med. Food 2009, 12, 782–787. [Google Scholar] [CrossRef]
- Dai, Y.; Jiang, Y.; Lee, H.G.; Jeon, Y.; Kang, M. Characterization and Screening of Anti-Tumor Activity of Fucoidan from Acid-Processed Hijiki (Hizikia fusiforme). Int. J. Biol. Macromol. 2019, 139, 170–180. [Google Scholar] [CrossRef]
- Folin, O.; Denis, W. On Phosphotungstic-Phosphomolybdic Compounds as Color Reagents. J. Biol. Chem. 1912, 12, 239–243. [Google Scholar]
- Kang, S.; Kim, K.; Lee, S.; Ahn, G.; Cha, S.; Kim, A.; Yang, X.; Kang, M.; Jeon, Y. Anti-Inflammatory Activity of Polysaccharide Purified from AMG-Assistant Extract of Ecklonia cava in LPS-Stimulated RAW 264.7 Macrophages. Carbohydr. Polym. 2011, 85, 80–85. [Google Scholar] [CrossRef]
- Lee, J.; Kim, H.; Ko, J.; Jang, J.; Kim, G.; Lee, J.; Nah, J.; Jeon, Y. Rapid Preparation of Functional Polysaccharides from Pyropia yezoensis by Microwave-Assistant Rapid Enzyme Digest System. Carbohydr. Polym. 2016, 153, 512–517. [Google Scholar] [CrossRef]
- Wang, L.; Oh, J.Y.; Kim, H.S.; Lee, W.; Cui, Y.; Lee, H.G.; Kim, Y.; Ko, J.Y.; Jeon, Y. Protective Effect of Polysaccharides from Celluclast-Assisted Extract of Hizikia fusiforme Against Hydrogen Peroxide-Induced Oxidative Stress in Vitro in Vero Cells and in Vivo in Zebrafish. Int J. Biol. Macromol. 2018, 112, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Osborne, C.K.; Boldt, D.H.; Clark, G.M.; Trent, J.M. Effects of Tamoxifen on Human Breast Cancer Cell Cycle Kinetics: Accumulation of Cells in Early G1 Phase. Cancer Res. 1983, 43, 3583–3585. [Google Scholar]
- Lin, H.L.; Chang, Y.F.; Liu, T.Y.; Wu, C.W.; Chi, C.W. Submicromolar Paclitaxel Induces Apoptosis in Human Gastric Cancer Cells at Early G1 Phase. Anticancer Res. 1998, 18, 3443–3449. [Google Scholar] [PubMed]
- Kamiyama, M.; Naguro, I.; Ichijo, H. In Vivo Gene Manipulation Reveals the Impact of Stress-Responsive MAPK Pathways on Tumor Progression. Cancer Sci. 2015, 106, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Vivanco, I.; Sawyers, C.L. The Phosphatidylinositol 3-Kinase AKT Pathway in Human Cancer. Nat. Rev. Cancer 2002, 2, 489–501. [Google Scholar] [CrossRef]
- Kim, S.O.; Choi, Y.H. The Ethyl Alcohol Extract of Hizikia Fusiforme Inhibits Matrix Metalloproteinase Activity and Regulates Tight Junction Related Protein Expression in Hep3B Human Hepatocarcinoma Cells. J. Med. Food 2010, 13, 31–38. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | CHF | SPHF |
|---|---|---|
| Yield (%) | 45.12 | 31.12 |
| Phenolic content (%) | 1.88 | 1.79 |
| Sulfated polysaccharide (%) | 51.32 | 65.23 |
| Proportion of Fucose (%) | 27.31 | 59.63 |
| Sample | Free Radical Scavenging Activity (IC50 mg/mL) | |
|---|---|---|
| DPPH | TBARs | |
| CHF | 0.92 ± 0.03 | 2.2 ± 0.01 |
| SPHF | 0.79 ± 0.04 | 1.5 ± 0.02 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.-H.; Won, S.Y.; Hwang, B.; Jung, S.; Choi, C.; Park, S.-S.; Choi, Y.H.; Kim, W.-J.; Moon, S.-K. In Vitro and In Vivo Antitumor Efficacy of Hizikia fusiforme Celluclast Extract against Bladder Cancer. Nutrients 2020, 12, 2159. https://doi.org/10.3390/nu12072159
Song J-H, Won SY, Hwang B, Jung S, Choi C, Park S-S, Choi YH, Kim W-J, Moon S-K. In Vitro and In Vivo Antitumor Efficacy of Hizikia fusiforme Celluclast Extract against Bladder Cancer. Nutrients. 2020; 12(7):2159. https://doi.org/10.3390/nu12072159
Chicago/Turabian StyleSong, Jun-Hui, Se Yeon Won, Byungdoo Hwang, Soontag Jung, Changsun Choi, Sung-Soo Park, Yung Hyun Choi, Wun-Jae Kim, and Sung-Kwon Moon. 2020. "In Vitro and In Vivo Antitumor Efficacy of Hizikia fusiforme Celluclast Extract against Bladder Cancer" Nutrients 12, no. 7: 2159. https://doi.org/10.3390/nu12072159
APA StyleSong, J. -H., Won, S. Y., Hwang, B., Jung, S., Choi, C., Park, S. -S., Choi, Y. H., Kim, W. -J., & Moon, S. -K. (2020). In Vitro and In Vivo Antitumor Efficacy of Hizikia fusiforme Celluclast Extract against Bladder Cancer. Nutrients, 12(7), 2159. https://doi.org/10.3390/nu12072159