Extending the Overnight Fast: Sex Differences in Acute Metabolic Responses to Breakfast

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Study Design
2.3. Data Analysis
3. Results
3.1. Participant Characteristics
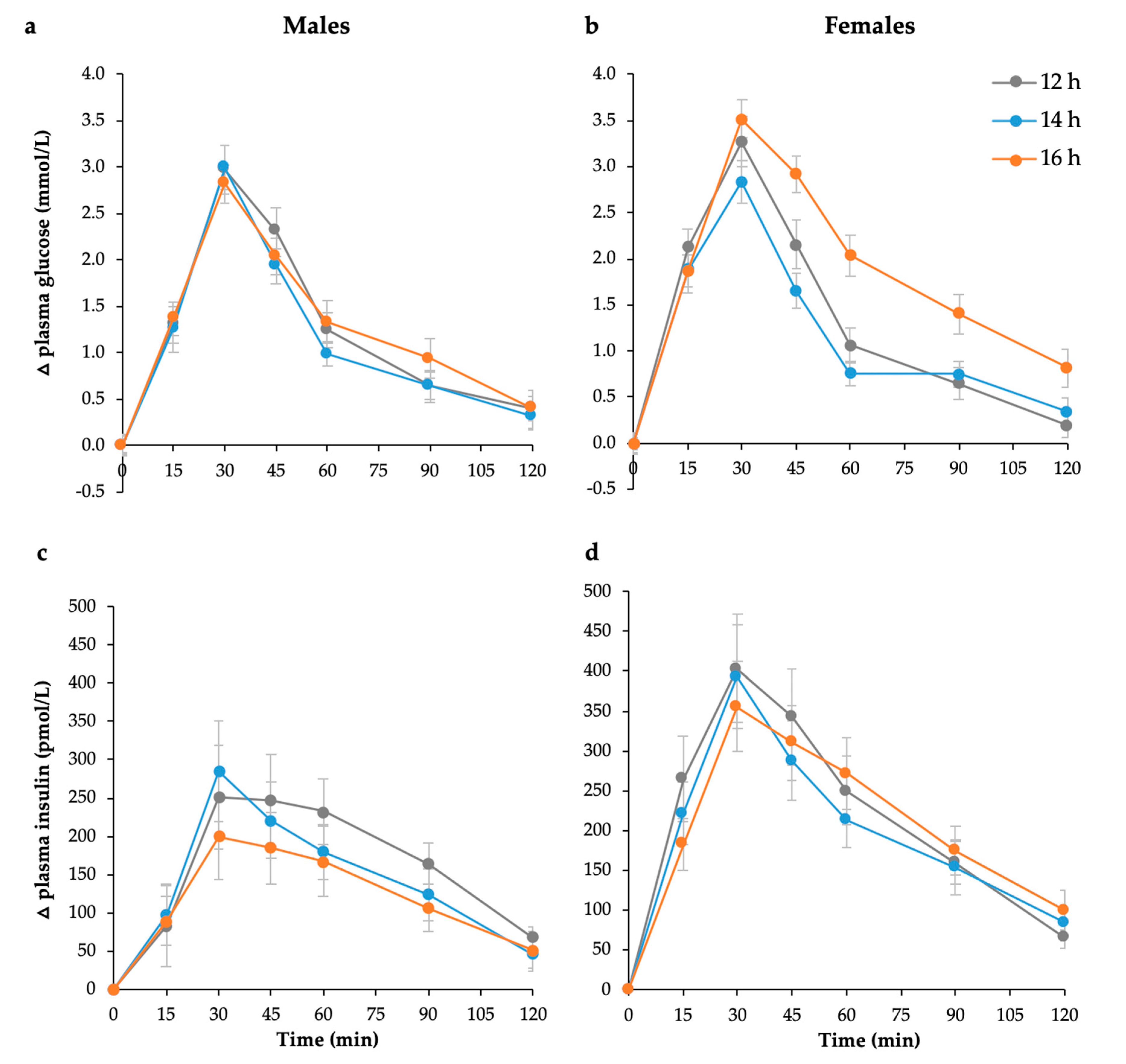
3.2. Postprandial Glycaemia, Insulinemia, and Measures of Insulin Sensitivity
3.3. Subsequent Meal Energy Intake
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wyatt, H.R.; Grunwald, G.K.; Mosca, C.L.; Klem, M.L.; Wing, R.R.; Hill, J.O. Long-term weight loss and breakfast in subjects in the National Weight Control Registry. Obes. Res. 2002, 10, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Schlundt, D.G.; Hill, J.O.; Sbrocco, T.; Pope-Cordle, J.; Sharp, T. The role of breakfast in the treatment of obesity: A randomized clinical trial. Am. J. Clin. Nutr. 1992, 55, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Kleinman, R.E.; Hall, S.; Green, H.; Korzec-Ramirez, D.; Patton, K.; Pagano, M.E.; Murphy, J.M. Diet, breakfast, and academic performance in children. Ann. Nutr. Metab. 2002, 46 (Suppl. 1), 24–30. [Google Scholar] [CrossRef] [Green Version]
- Haines, P.S.; Guilkey, D.K.; Popkin, B.M. Trends in breakfast consumption of US adults between 1965 and 1991. J. Am. Diet. Assoc. 1996, 96, 464–470. [Google Scholar] [CrossRef]
- Siega-Riz, A.M.; Popkin, B.M.; Carson, T. Trends in breakfast consumption for children in the United States from 1965–1991. Am. J. Clin. Nutr. 1998, 67, 748S–756S. [Google Scholar] [CrossRef] [PubMed]
- Keski-Rahkonen, A.; Kaprio, J.; Rissanen, A.; Virkkunen, M.; Rose, R.J. Breakfast skipping and health-compromising behaviors in adolescents and adults. Eur. J. Clin. Nutr. 2003, 57, 842–853. [Google Scholar] [CrossRef] [Green Version]
- Song, W.O.; Chun, O.K.; Obayashi, S.; Cho, S.; Chung, C.E. Is consumption of breakfast associated with body mass index in US adults? J. Am. Diet. Assoc. 2005, 105, 1373–1382. [Google Scholar] [CrossRef]
- Ma, Y.; Bertone, E.R.; Stanek, E.J.; Reed, G.W.; Hebert, J.R.; Cohen, N.L.; Merriam, P.A.; Ockene, I.S. Association between eating patterns and obesity in a free-living US adult population. Am. J. Epidemiol. 2003, 158, 85–92. [Google Scholar] [CrossRef]
- Ma, X.; Chen, Q.; Pu, Y.; Guo, M.; Jiang, Z.; Huang, W.; Long, Y.; Xu, Y. Skipping breakfast is associated with overweight and obesity: A systematic review and meta-analysis. Obes. Res. Clin. Pract. 2020, 14, 1–8. [Google Scholar] [CrossRef]
- Mekary, R.A.; Giovannucci, E.; Willett, W.C.; van Dam, R.M.; Hu, F.B. Eating patterns and type 2 diabetes risk in men: Breakfast omission, eating frequency, and snacking. Am. J. Clin. Nutr. 2012, 95, 1182–1189. [Google Scholar] [CrossRef] [Green Version]
- Mekary, R.A.; Giovannucci, E.; Cahill, L.; Willett, W.C.; van Dam, R.M.; Hu, F.B. Eating patterns and type 2 diabetes risk in older women: Breakfast consumption and eating frequency. Am. J. Clin. Nutr. 2013, 98, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Morgan, K.J.; Zabik, M.E.; Stampley, G.L. The role of breakfast in diet adequacy of the U.S. adult population. J. Am. Coll. Nutr. 1986, 5, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Gabel, K.; Hoddy, K.K.; Haggerty, N.; Song, J.; Kroeger, C.M.; Trepanowski, J.F.; Panda, S.; Varady, K.A. Effects of 8-hour time restricted feeding on body weight and metabolic disease risk factors in obese adults: A pilot study. Nutr. Healthy Aging 2018, 4, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, M.; Cioffi, I.; Evangelista, A.; Ponzo, V.; Goitre, I.; Ciccone, G.; Ghigo, E.; Bo, S. Effects of time-restricted feeding on body weight and metabolism. A systematic review and meta-analysis. Rev. Endocr. Metab. Disord. 2020, 21, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Groop, L.C.; Bonadonna, R.; DelPrato, S.; Ratheiser, K.; DeFronzo, R.A. Effect of prolonged overnight fasting on energy metabolism in non-insulin-dependent diabetic and non-diabetic subjects. Acta Endocrinol. 1990, 123, 30–36. [Google Scholar] [CrossRef]
- Horton, T.J.; Hill, J.O. Prolonged fasting significantly changes nutrient oxidation and glucose tolerance after a normal mixed meal. J. Appl. Physiol. 2001, 90, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Fery, F.; Plat, L.; Balasse, E.O. Mechanisms of whole-body glycogen deposition after oral glucose in normal subjects. Influence of the nutritional status. J. Clin. Endocrinol. Metab. 1998, 83, 2810–2816. [Google Scholar] [CrossRef]
- Goschke, H. Mechanism of glucose intolerance during fasting: Differences between lean and obese subjects. Metab. Clin. Exp. 1977, 26, 1147–1153. [Google Scholar] [CrossRef]
- Mansell, P.I.; Macdonald, I.A. The effect of starvation on insulin-induced glucose disposal and thermogenesis in humans. Metab. Clin. Exp. 1990, 39, 502–510. [Google Scholar] [CrossRef]
- Dickinson, S.; Hancock, D.; Petocz, P.; Ceriello, A.; Brand-Miller, J. High glycemic index carbohydrate increases NF-kB activation in mononuclear cells of young lean healthy subjects. Am. J. Clin. Nutr. 2008, 87, 1188–1193. [Google Scholar] [CrossRef]
- Holt, S.; Brand, J.; Soveny, C.; Hansky, J. Relationship of satiety to postprandial glycaemic, insulin and cholecystokinin responses. Appetite 1992, 18, 129–141. [Google Scholar] [CrossRef]
- Food and Agriculture Organization/World Health Organization. Carbohydrates in Human Nutrition: Report of a Joint FAO/WHO Expert Consultation; WHO: Rome, Italy, 1998; pp. 25–37. [Google Scholar]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, J.C.; Matthews, D.R.; Hermans, M.P. Correct homeostasis model assessment (HOMA) evaluation uses the computer program. Diabetes Care 1998, 21, 2191–2192. [Google Scholar] [CrossRef] [PubMed]
- Stanton, J.L., Jr.; Keast, D.R. Serum cholesterol, fat intake, and breakfast consumption in the United States adult population. J. Am. Coll. Nutr. 1989, 8, 567–572. [Google Scholar] [CrossRef]
- Kral, T.V.E.; Whiteford, L.M.; Heo, M.; Faith, M.S. Effects of eating breakfast compared with skipping breakfast on ratings of appetite and intake at subsequent meals in 8- to 10-y-old children. Am. J. Clin. Nutr. 2011, 93, 284–291. [Google Scholar] [CrossRef] [Green Version]
- Levitsky, D.A.; Pacanowski, C.R. Effect of skipping breakfast on subsequent energy intake. Physiol. Behav. 2013, 119, 9–16. [Google Scholar] [CrossRef]
- Antuna-Puente, B.; Disse, E.; Rabasa-Lhoret, R.; Laville, M.; Capeau, J.; Bastard, J.P. How can we measure insulin sensitivity/resistance? Diabetes Metab. 2011, 37, 179–188. [Google Scholar] [CrossRef]
- Randle, P.J.; Garland, P.B.; Hales, C.N.; Newsholme, E.A. The glucose fatty-acid cycle. Its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus. Lancet 1963, 1, 785–789. [Google Scholar] [CrossRef]
- Shepherd, P.R.; Kahn, B.B. Glucose transporters and insulin action--implications for insulin resistance and diabetes mellitus. N. Engl. J. Med. 1999, 341, 248–257. [Google Scholar] [CrossRef]
- Pfeiffer, A.F.H.; Keyhani-Nejad, F. High glycemic index metabolic damage—A pivotal role of GIP and GLP-1. Trends Endocrinol. Metab. 2018, 29, 289–299. [Google Scholar] [CrossRef] [Green Version]
- DeFronzo, R.A. Pathogenesis of type 2 (non-insulin dependent) diabetes mellitus: A balanced overview. Diabetologia 1992, 35, 389–397. [Google Scholar] [CrossRef]
- Ahmed, M.; Gannon, M.C.; Nuttall, F.Q. Postprandial plasma glucose, insulin, glucagon and triglyceride responses to a standard diet in normal subjects. Diabetologia 1976, 12, 61–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedrington, M.S.; Davis, S.N. Sexual dimorphism in glucose and lipid metabolism during fasting, hypoglycemia, and exercise. Front. Endocrinol. 2015, 6, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepe, G.J.; Albrecht, E.D. Actions of Placental and Fetal Adrenal Steroid Hormones in Primate Pregnancy. Endocr. Rev. 1995, 16, 608–648. [Google Scholar] [CrossRef]
- Godsland, I.F. The influence of female sex steroids on glucose metabolism and insulin action. J. Intern. Med. 1996, 738, 1–60. [Google Scholar]
- Boden, G. Fuel metabolism in pregnancy and in gestational diabetes mellitus. Obstet. Gynecol. Clin. 1996, 23, 1–10. [Google Scholar] [CrossRef]
- Homko, C.J.; Sivan, E.; Reece, E.A.; Boden, G. Fuel metabolism during pregnancy. Semin. Reprod. Endocrinol. 1999, 17, 119–125. [Google Scholar] [CrossRef]
- Metzger, B.E.; Lowe, L.P.; Dyer, A.R.; Trimble, E.R.; Chaovarindr, U.; Coustan, D.R.; Hadden, D.R.; McCance, D.R.; Hod, M.; McIntyre, H.D.; et al. Hyperglycemia and adverse pregnancy outcomes. N. Engl. J. Med. 2008, 358, 1991–2002. [Google Scholar] [CrossRef] [Green Version]
- Wolever, T.M.; Jenkins, D.J.; Jenkins, A.L.; Josse, R.G. The glycemic index: Methodology and clinical implications. Am. J. Clin. Nutr. 1991, 54, 846–854. [Google Scholar] [CrossRef]
- International Standards Organisation. Food Products—Determination of the Glycaemic Index (GI) and Recommendation for Food Classification; ISO 26642–2010; International Standards Organisation: Geneva, Switzerland, 2010. [Google Scholar]
- Wallace, T.M.; Levy, J.C.; Matthews, D.R. Use and abuse of HOMA modeling. Diabetes Care 2004, 27, 1487–1495. [Google Scholar] [CrossRef] [Green Version]
- Ellison, J.M.; Stegmann, J.M.; Colner, S.L.; Michael, R.H.; Sharma, M.K.; Ervin, K.R.; Horwitz, D.L. Rapid changes in postprandial blood glucose produce concentration differences at finger, forearm, and thigh sampling sites. Diabetes Care 2002, 25, 961–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeevi, D.; Korem, T.; Zmora, N.; Israeli, D.; Rothschild, D.; Weinberger, A.; Ben-Yacov, O.; Lador, D.; Avnit-Sagi, T.; lotan-Pompan, M.; et al. Personalized nutrition by prediction of glycemic responses. Cell 2015, 163, 1079–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Variables | 12 h | 14 h | 16 h | p-Value 2 | p-Value 3 |
|---|---|---|---|---|---|
| Fasting glucose (mmol/L) | 5.2 ± 0.06 | 5.2 ± 0.06 | 5.0 ± 0.06 | 0.058 | - |
| 2-h glucose (mmol/L) | 5.4 ± 0.10 | 5.5 ± 0.10 | 5.8 ± 0.10 | 0.021 * | - |
| Glucose iAUC (mmol/L·120 min) | 162.9 ± 11.3 | 143.0 ± 11.3 | 225.5 ± 11.3 | <0.001 ** | - |
| Fasting insulin (pmol/L) | 28.0 ± 2.0 | 24.1 ± 2.0 | 18.6 ± 2.0 | 0.056 | 0.069 |
| 2-h insulin (pmol/L) | 94.2 ± 11.9 | 107.8 ± 11.9 | 118.2 ± 11.9 | 0.255 | 0.217 |
| Insulin iAUC (pmol/L·120 min) | 26,582 ± 2283 | 24,176 ± 2283 | 25,574 ± 2283 | 0.294 | - |
| Log insulin iAUC (pmol/L·120 min) 4 | 21,456 | 19,570 | 21,955 | 0.342 | - |
| HOMA-IR | 0.53 ± 0.03 | 0.46 ± 0.03 | 0.35 ± 0.03 | 0.052 | 0.064 |
| Log of HOMA-IR 4 | 0.44 | 0.42 | 0.32 | 0.024 * | 0.031 * |
| HOMA (%S) | 265.5 ± 19.6 | 269.9 ± 19.6 | 359.9 ± 19.6 | 0.016 * | - |
| Log of HOMA %S 4 | 226.1 | 239.8 | 317.3 | 0.023 * | 0.030 * |
| HOMA (%B) | 55.0 ± 2.2 | 51.2 ± 2.2 | 46.3 ± 2.2 | 0.181 | 0.205 |
| Log of HOMA %B 4 | 50.4 | 48.9 | 44.0 | 0.254 | 0.267 |
| Variables | 12 h | 14 h | 16 h | p-Value 2 | p-Value 3 |
|---|---|---|---|---|---|
| Fasting glucose (mmol/L) | 5.4 ± 0.05 | 5.3 ± 0.05 | 5.2 ± 0.05 | 0.024 | - |
| 2-h glucose (mmol/L) | 5.8 ± 0.11 | 5.6 ± 0.11 | 5.6 ± 0.11 | 0.394 | 0.410 |
| Glucose iAUC (mmol/L·120 min) | 168.8 ± 6.4 | 149.9 ± 6.4 | 165.6 ± 6.4 | 0.326 | 0.362 |
| Fasting insulin (pmol/L) | 24.2 ± 2.1 | 24.3 ± 2.1 | 21.2 ± 2.1 | 0.577 | 0.450 |
| 2-h insulin (pmol/L) | 95.2 ± 8.0 | 78.0 ± 8.0 | 71.8 ± 8.0 | 0.239 | - |
| Insulin iAUC (pmol/L·120 min) | 21,739 ± 2481 | 20,255 ± 2481 | 16,303 ± 2481 | 0.053 | 0.080 |
| Log insulin iAUC (pmol/L·120 min) 4 | 15,408 | 15,287 | 12,642 | 0.025 | - |
| HOMA-IR | 0.47 ± 0.029 | 0.46 ± 0.029 | 0.40 ± 0.029 | 0.529 | 0.390 |
| Log of HOMA-IR 4 | 0.40 | 0.39 | 0.33 | 0.232 | 0.107 |
| HOMA (%S) | 289.5 ± 30.0 | 320.8 ± 30.0 | 375.8 ± 30.0 | 0.100 | - |
| Log of HOMA %S 4 | 250.1 | 258.5 | 307.0 | 0.231 | 0.107 |
| HOMA (%B) | 46.7 ± 2.0 | 47.9 ± 2.0 | 46.3 ± 2.0 | 0.941 | 0.951 |
| Log of HOMA %B 4 | 43.7 | 43.6 | 41.4 | 0.759 | 0.670 |
| Variables | 12 h | 14 h | 16 h | p-Value 2 |
|---|---|---|---|---|
| Energy intake (kJ) | 2293 ± 96 | 2700 ± 91 | 2936 ± 88 | <0.001 ** |
| Protein (g) | 18.1 ± 1.3 | 24.4 ± 1.3 | 27.9 ± 1.2 | 0.002 ** |
| Total fat (g) | 17.8 ± 1.2 | 24.8 ± 1.2 | 27.1 ± 1.2 | <0.001 ** |
| Saturated fat (g) | 7.1 ± 0.7 | 9.9 ± 0.6 | 11.5 ± 0.6 | 0.001 ** |
| Carbohydrates—total (g) | 74.4 ± 3.4 | 78.4 ± 3.2 | 83.8 ± 3.1 | 0.038 * |
| Carbohydrates—sugars (g) | 34.3 ± 2.3 | 34.9 ± 2.2 | 36.7 ± 2.2 | 0.41 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
S. Atkinson, F.; A. Heruc, G.; M. H. Tan, V.; Petocz, P.; C. Brand-Miller, J. Extending the Overnight Fast: Sex Differences in Acute Metabolic Responses to Breakfast. Nutrients 2020, 12, 2173. https://doi.org/10.3390/nu12082173
S. Atkinson F, A. Heruc G, M. H. Tan V, Petocz P, C. Brand-Miller J. Extending the Overnight Fast: Sex Differences in Acute Metabolic Responses to Breakfast. Nutrients. 2020; 12(8):2173. https://doi.org/10.3390/nu12082173
Chicago/Turabian StyleS. Atkinson, Fiona, Gabriella A. Heruc, Verena M. H. Tan, Peter Petocz, and Jennie C. Brand-Miller. 2020. "Extending the Overnight Fast: Sex Differences in Acute Metabolic Responses to Breakfast" Nutrients 12, no. 8: 2173. https://doi.org/10.3390/nu12082173
APA StyleS. Atkinson, F., A. Heruc, G., M. H. Tan, V., Petocz, P., & C. Brand-Miller, J. (2020). Extending the Overnight Fast: Sex Differences in Acute Metabolic Responses to Breakfast. Nutrients, 12(8), 2173. https://doi.org/10.3390/nu12082173