The Importance of the Fatty Acid Transporter L-Carnitine in Non-Alcoholic Fatty Liver Disease (NAFLD)




Abstract
:1. Introduction
2. The Importance of L-Carnitine
2.1. Dietary Intake of L-carnitine
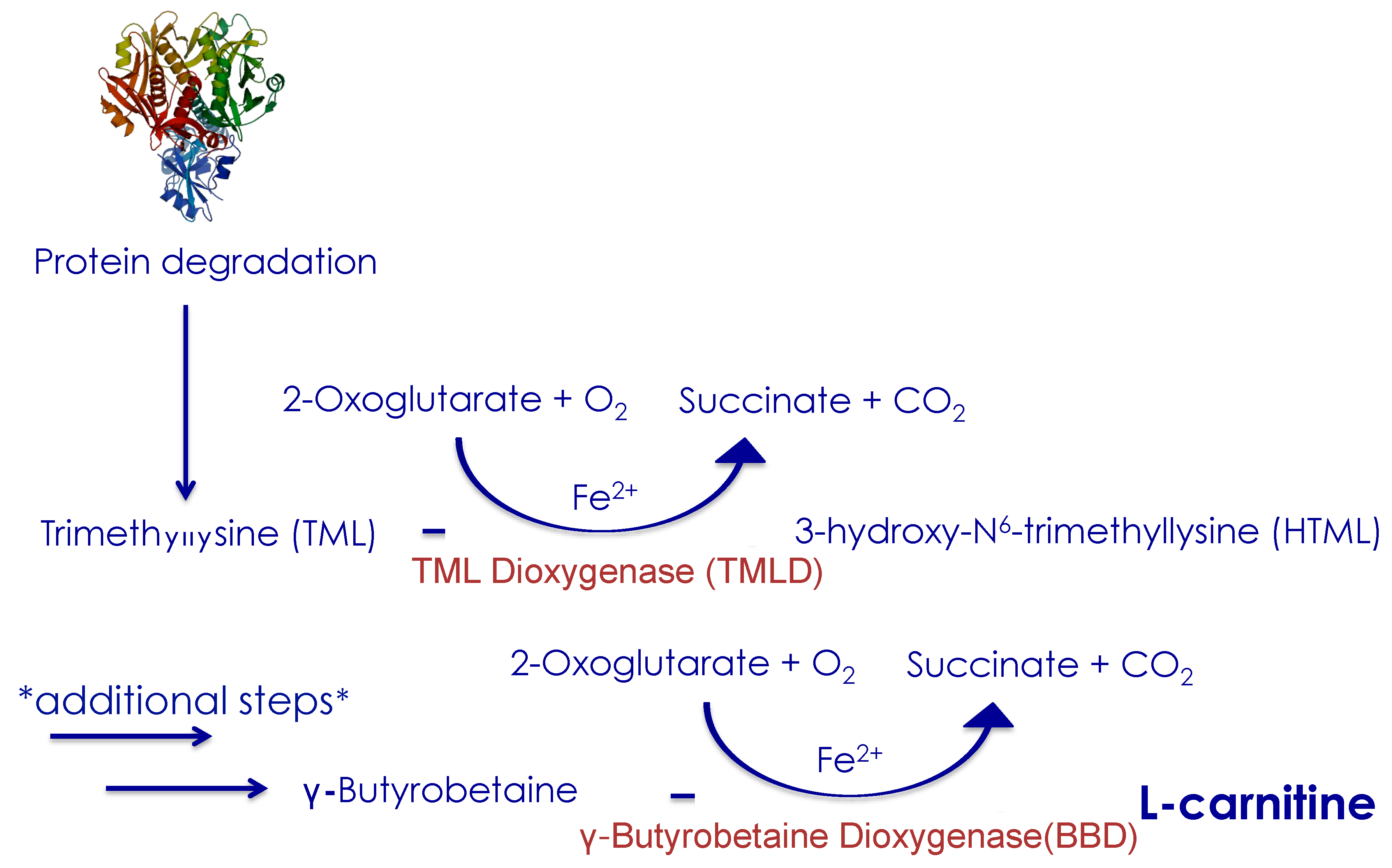
2.2. Endogenous L-carnitine Synthesis
2.3. L-Carnitine Absorption
2.4. Transport of Fatty Acids into Mitochondria
2.5. L-Carnitine as a Buffer for Excess Acetyl-CoA
3. Fatty Liver Disease and the Role of L-Carnitine
3.1. Drivers of Non-Alcoholic Fatty Liver Disease (NAFLD)
3.2. Fatty Liver Disease Is a Feature of Primary Carnitine Deficiency
3.3. Other Evidence Linking L-Carnitine Deficiency to Liver Disease
3.4. Patients with Chronic Liver Disease have Low Levels of L-Carnitine
3.5. Acyl-Carnitine Chain Length can be Associated with Liver Disease
4. The Importance of L-Carnitine Supplementation
4.1. L-Carnitine Supplementation is Beneficial to the Liver
4.2. Effects of L-Carnitine in Ketogenesis
4.3. L-Carnitine has Significant Effect on Insulin and Glucose Levels
5. Summary and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Younossi, Z.M.; Marchesini, G.; Pinto-Cortez, H.; Petta, S. Epidemiology of nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Transplantation 2019, 103, 22–27. [Google Scholar] [CrossRef] [PubMed]
- European Association for the Study of the Liver (EASL); European Association for the Study of Diabetes (EASD); European Association for the Study of Obesity (EASO). EASL-EASD-EASO clinical practice guidelines for the management of non-alcoholic fatty liver disease. Obes. Facts 2016, 9, 65–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angulo, P.; Kleiner, D.E.; Dam-Larsen, S.; Adams, L.A.; Bjornsson, E.S.; Charatcharoenwitthaya, P.; Mills, P.R.; Keach, J.C.; Lafferty, H.D.; Stahler, A.; et al. Liver fibrosis, but no other histologic features, is associated with long-term outcomes of patients with nonalcoholic fatty liver disease. Gastroenterology 2015, 149, 389–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fielding, C.M.; Angulo, P. Hepatic steatosis and steatohepatitis: Are they really two distinct entities? Curr. Hepatol. Rep. 2014, 13, 151–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchesini, G.; Brizi, M.; Bianchi, G.; Tomassetti, S.; Bugianesi, E.; Lenzi, M.; McCullough, A.J.; Natale, S.; Forlani, G.; Melchionda, N. Nonalcoholic fatty liver disease: A feature of the metabolic syndrome. Diabetes 2001, 50, 1844–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamamoğullari, N.; Siliğ, Y.; Içağasioğlu, S.; Atalay, A. Carnitine deficiency in diabetes mellitus complications. J. Diabetes Complicat. 1999, 13, 251–253. [Google Scholar] [CrossRef]
- Pekala, J.; Patkowska-Sokola, B.; Bodkowski, R.; Jamroz, D.; Nowakowski, P.; Lochynski, S.; Librowski, T. L-Carnitine-Metabolic functions and meaning in humans life. Curr. Drug Metab. 2011, 12, 667–668. [Google Scholar] [CrossRef]
- Rodrigues, B.; Xiang, H.; McNeill, J.H. Effect of L-Carnitine treatment on lipid metabolism and cardiac performance in chronically diabetic rats. Diabetes 1988, 37, 1358–1364. [Google Scholar] [CrossRef]
- Cecere, A.; Ciaramella, F.; Tancredi, L.; Romano, C.; Gattoni, A. Efficacy of L-Carnitine in reducing hyperammonaemia and improving neuropsychological test performance in patients with hepatic cirrhosis results of a randomised Trial. Clin. Drug Investig. 2002, 22, 7–14. [Google Scholar] [CrossRef]
- Malaguarnera, M.; Gargante, M.P.; Russo, C.; Antic, T.; Vacante, M.; Malaguarnera, M.; Avitabile, T.; Li Volti, G.; Galvano, F. L-Carnitine supplementation to diet: A new tool in treatment of nonalcoholic steatohepatitis—A randomized and controlled clinical trial. Am. J. Gastroenterol. 2010, 105, 1338–1345. [Google Scholar] [CrossRef]
- Bae, J.C.; Lee, W.Y.; Yoon, K.H.; Park, J.Y.; Son, H.S.; Han, K.A.; Lee, K.W.; Woo, J.T.; Ju, Y.C.; Lee, W.J.; et al. Improvement of nonalcoholic fatty liver disease with carnitine-orotate complex in type 2 diabetes (CORONA): A randomized controlled trial. Diabetes Care 2015, 38, 1245–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, A.; Tsuda, Y.; Asai, A.; Yokohama, K.; Nakamura, K.; Sujishi, T.; Ohama, H.; Tsuchimoto, Y.; Fukunishi, S.; Abdelaal, U.M.; et al. Effects of oral L-carnitine on liver functions after transarterial chemoembolization in intermediate-stage HCC patients. Mediat. Inflamm. 2015, 2015, 608216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulewitsch, W.; Krimberg, R. Zur kenntnis der extraktivstoffe der muskeln: II. mitteilung: Über das carnitin. Hoppe Seyler’s Z. Physiol. Chem. 1905, 45, 326–330. [Google Scholar] [CrossRef] [Green Version]
- Kutscher, F. Zur kenntnis des novains. Hoppe Seyler’s Z. Physiol. Chem. 1906, 49, 47–49. [Google Scholar] [CrossRef]
- Harmeyer, J. The physiological role of L-carnitine. Lohmann Inf. 2002, 27, 15–21. [Google Scholar]
- Noland, R.C.; Koves, T.R.; Seiler, S.E.; Lum, H.; Lust, R.M.; Ilkayeva, O.; Stevens, R.D.; Hegardt, F.G.; Muoio, D.M. Carnitine insufficiency caused by aging and overnutrition compromises mitochondrial performance and metabolic control. J. Biol. Chem. 2009, 284, 22840–22852. [Google Scholar] [CrossRef] [Green Version]
- Adeva-Andany, M.M.; Calvo-Castro, I.; Fernández-Fernández, C.; Donapetry-García, C.; Pedre-Piñeiro, A.M. Significance of L-carnitine for human health. IUBMB Life 2017, 69, 578–594. [Google Scholar] [CrossRef] [Green Version]
- Stephens, F.B.; Marimuthu, K.; Cheng, Y.; Patel, N.; Constantin, D.; Simpson, E.J.; Greenhaff, P.L. Vegetarians have a reduced skeletal muscle carnitine transport capacity. Am. J. Clin. Nutr. 2011, 94, 938–944. [Google Scholar] [CrossRef] [Green Version]
- Lombard, K.A.; Olson, A.L.; Nelson, S.E.; Rebouche, C.J. Carnitine status of lactoovovegetarians and strict vegetarian adults and children. Am. J. Clin. Nutr. 1989, 50, 301–306. [Google Scholar] [CrossRef]
- Krajčovičová-Kudláčková, M.; Šimončič, R.; Béderová, A.; Babinská, K.; Béder, I. Correlation of carnitine levels to methionine and lysine intake. Physiol. Res. 2000, 49, 399–402. [Google Scholar]
- Rosell, M.S.; Lloyd-Wright, Z.; Appleby, P.N.; Sanders, T.A.; Allen, N.E.; Key, T.J. Long-chain n-3 polyunsaturated fatty acids in plasma in british meat-eating, vegetarian, and vegan men. Am. J. Clin. Nutr. 2005, 82, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Evans, A.M.; Fornasini, G. Pharmacokinetics of L-Carnitine. Clin. Pharmacokinet. 2003, 42, 941–967. [Google Scholar] [CrossRef] [PubMed]
- Levocarnitine. Drugs and Lactation Database (LactMed); National Library of Medicine (US): Bethesda, MD, USA, 2006. Available online: https://www.ncbi.nlm.nih.gov/books/NBK501864/ (accessed on 19 June 2020).
- Rebouche, C.J.; Paulson, D.J. Carnitine Metabolism and Function in Humans. Annu. Rev. Nutr. 1986, 6, 41–66. [Google Scholar] [CrossRef] [PubMed]
- Sahajwalla, C.G.; Helton, E.D.; Purich, E.D.; Hoppel, C.L.; Cabana, B.E. Multiple-dose pharmacokinetics and bioequivalence of L-carnitine 330-mg tablet versus 1-g chewable tablet versus enteral solution in healthy adult male volunteers. J. Pharm. Sci. 1995, 84, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Segre, G.; Bianchi, E.; Corsi, M.; D’Iddio, S.; Ghirardi, O.; Maccari, F. Plasma and urine pharmacokinetics of free and of short-chain carnitine after administration of carnitine in man. Arzneim. Forsch. Drug Res. 1988, 38, 1830. [Google Scholar]
- Rebouche, C.J.; Chenard, C.A. Metabolic fate of dietary carnitine in human adults: Identification and quantification of urinary and fecal metabolites. J. Nutr. 1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strijbis, K.; Vaz, F.M.; Distel, B. Enzymology of the carnitine biosynthesis pathway. IUBMB Life 2010. [Google Scholar] [CrossRef]
- Englard, S. Hydroxylation of γ-butyrobetaine to carnitine in human and monkey tissues. FEBS Lett. 1979. [Google Scholar] [CrossRef] [Green Version]
- Priore, P.; Stanca, E.; Gnoni, G.V.; Siculella, L. Dietary fat types differently modulate the activity and expression of mitochondrial carnitine/acylcarnitine translocase in rat liver. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2012. [Google Scholar] [CrossRef]
- Longo, N.; Frigeni, M.; Pasquali, M. Carnitine transport and fatty acid oxidation. Biochim. Biophys. Acta 2016, 1863, 2422–2435. [Google Scholar] [CrossRef]
- Schönfeld, P.; Wojtczak, L. Short- and medium-chain fatty acids in energy metabolism: The cellular perspective. J. Lipid Res. 2016, 57, 943–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, S.; Zhan, Z.-Y.; Cao, H.-Y.; Wu, C.; Bian, Y.-Q.; Li, J.-Y.; Cheng, G.-H.; Liu, P.; Sun, M.-Y. Urinary metabolomics analysis identifies key biomarkers of different stages of nonalcoholic fatty liver disease. World J. Gastroenterol. 2017, 23, 2771–2784. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Hajhossein Talasaz, A.; Alidoosti, M. Preventive effect of L-carnitine and its derivatives on endothelial dysfunction and platelet aggregation. Clin. Nutr. ESPEN 2016, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rinaldo, P.; Cowan, T.M.; Matern, D. Acylcarnitine profile analysis. Genet. Med. 2008, 10, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Bae, J.C.; Cho, Y.K.; Lee, W.Y.; Seo, H.I.; Rhee, E.J.; Park, S.E.; Park, C.Y.; Oh, K.W.; Sung, K.C.; Kim, B.I. Impact of nonalcoholic fatty liver disease on insulin resistance in relation to HbA1c levels in nondiabetic subjects. Am. J. Gastroenterol. 2010, 105, 2389–2395. [Google Scholar] [CrossRef] [PubMed]
- Rector, R.S.; Thyfault, J.P.; Wei, Y.; Ibdah, J.A. Non-alcoholic fatty liver disease and the metabolic syndrome: An update. World J. Gastroenterol. 2008, 14, 185. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.C.; Rhee, E.J.; Lee, W.Y.; Park, S.E.; Park, C.Y.; Oh, K.W.; Park, S.W.; Kim, S.W. Combined effect of nonalcoholic fatty liver disease and impaired fasting glucose on the development of type 2 diabetes: A 4-year retrospective longitudinal study. Diabetes Care 2011, 34, 727–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepanova, M.; Younossi, Z.M. Independent association between nonalcoholic fatty liver disease and cardiovascular disease in the US population. Clin. Gastroenterol. Hepatol. 2012, 10, 646–650. [Google Scholar] [CrossRef]
- Gruben, N.; Shiri-Sverdlov, R.; Koonen, D.P.Y.; Hofker, M.H. Nonalcoholic fatty liver disease: A main driver of insulin resistance or a dangerous liaison? Biochim. Biophys. Acta Mol. Basis Dis. 2014, 1842, 2329–2343. [Google Scholar] [CrossRef] [Green Version]
- Cheung, O.; Sanyal, A. Abnormalities of lipid metabolism in nonalcoholic fatty liver disease. Semin. Liver Dis. 2008, 28, 351–359. [Google Scholar] [CrossRef]
- Baffy, G. Kupffer cells in non-alcoholic fatty liver disease: The emerging view. J. Hepatol. 2009. [Google Scholar] [CrossRef] [Green Version]
- El-Sheikh, A.A.; Rifaai, R.A. Peroxisome proliferator activator receptor (PPAR)-γ ligand, but not PPAR-α, ameliorates cyclophosphamide-induced oxidative stress and inflammation in rat liver. PPAR Res. 2014, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mamoulakis, D.; Galanakis, E.; Dionyssopoulou, E.; Evangeliou, A.; Sbyrakis, S. Carnitine deficiency in children and adolescents with type 1 diabetes. J. Diabetes Complicat. 2004, 18, 271–274. [Google Scholar] [CrossRef]
- la Marca, G.; Malvagia, S.; Toni, S.; Piccini, B.; Di Ciommo, V.; Bottazzo, G.F. Children who develop type 1 diabetes early in life show low levels of carnitine and amino acids at birth: Does this finding shed light on the etiopathogenesis of the disease? Nutr. Diabetes 2013, 3, e94. [Google Scholar] [CrossRef] [PubMed]
- Fucho, R.; Casals, N.; Serra, D.; Herrero, L. Ceramides and mitochondrial fatty acid oxidation in obesity. FASEB J. 2017, 31, 1263–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, C.C.G.; De Almeida, I.T.; Jakobs, C.; Poll-The, B.-T.; Duran, M. Dynamic changes of plasma acylcarnitine levels induced by fasting and sunflower oil challenge test in children. Pediatr. Res. 1999, 46, 440. [Google Scholar] [CrossRef] [Green Version]
- Chapoy, P.R.; Angelini, C.; Brown, W.J.; Stiff, J.E.; Shug, A.L.; Cederbaum, S.D. Systemic carnitine deficiency—A treatable inherited lipid-storage disease presenting as Reye’s Syndrome. N. Engl. J. Med. 1980, 303, 1389–1394. [Google Scholar] [CrossRef]
- CA, S. Carnitine deficiency disorders in children. Ann. N. Y. Acad. Sci. 2004, 1033, 42–51. [Google Scholar]
- Longo, N.; Amat di San Filippo, C.; Pasquali, M. Disorders of carnitine transport and the carnitine cycle. In American Journal of Medical Genetics Part C: Seminars in Medical Genetics; Wiley: Hoboken, NJ, USA, 2006; Volume 142, pp. 77–85. [Google Scholar]
- Jun, J.S.; Lee, E.J.; Park, H.D.; Kim, H.S. Systemic primary carnitine deficiency with hypoglycemic encephalopathy. Ann. Pediatr. Endocrinol. Metab. 2016, 21, 226. [Google Scholar] [CrossRef] [Green Version]
- Pike, L.S.; Smift, A.L.; Croteau, N.J.; Ferrick, D.A.; Wu, M. Inhibition of fatty acid oxidation by etomoxir impairs NADPH production and increases reactive oxygen species resulting in ATP depletion and cell death in human glioblastoma cells. Biochim. Biophys. Acta Bioenerg. 2011, 1807, 726–734. [Google Scholar] [CrossRef] [Green Version]
- Deswal, S.; Bijarnia-Mahay, S.; Manocha, V.; Hara, K.; Shigematsu, Y.; Saxena, R.; Verma, I.C. Primary carnitine deficiency—A rare treatable cause of cardiomyopathy and massive hepatomegaly. Indian J. Pediatr. 2017, 84, 83–85. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, J.; Nielsen, O.W.; Lund, A.M.; Køber, L.; Djurhuus, H. Primary carnitine deficiency and pivalic acid exposure causing encephalopathy and fatal cardiac events. J. Inherit. Metab. Dis. 2013, 36, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Wang, F.; Wang, Y.; Ye, J.; Qiu, W.; Zhang, H.; Gao, X.; Gong, Z.; Gu, X. Analysis of genetic mutations in Chinese patients with systemic primary carnitine deficiency. Eur. J. Med. Genet. 2014. [Google Scholar] [CrossRef] [PubMed]
- Treem, W.A.; Stanley, C.A. Massive hepatomegaly, steatosis, and secondary plasma carnitine deficiency in an infant with cystic fibrosis. Pediatrics 1989, 83, 993–997. [Google Scholar] [PubMed]
- Ravindranath, A.; Pai, G.; Srivastava, A.; Poddar, U.; Yachha, S.K. Infant with hepatomegaly and hypoglycemia: A setting for fatty acid oxidation defects. Indian J. Gastroenterol. 2017, 36, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Hinderling, V.B.; Schrauwen, P.; Langhans, W.; Westerterp-Plantenga, M.S. The effect of etomoxir on 24-h substrate oxidation and satiety in humans. Am. J. Clin. Nutr. 2002, 76, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Horn, C.C.; Ji, H.; Friedman, M.I. Etomoxir, a fatty acid oxidation inhibitor, increases food intake and reduces hepatic energy status in rats. Physiol. Behav. 2004, 81, 157–162. [Google Scholar] [CrossRef]
- Lundsgaard, A.M.; Fritzen, A.M.; Nicolaisen, T.S.; Carl, C.S.; Sjøberg, K.A.; Raun, S.H.; Klein, A.B.; Sanchez-Quant, E.; Langer, J.; Ørskov, C.; et al. Glucometabolic consequences of acute and prolonged inhibition of fatty acid oxidation. J. Lipid Res. 2020. [Google Scholar] [CrossRef]
- Coort, S.; Niessen, H.; Koonen, D.; Coumans, W.; Bonen, A.; Vandervusse, G.; Glatz, J.; Luiken, J. 204 effects of the hypertrophy-inducing agent etomoxir on fatty acid utilization and fatty acid transporters in rat heart and liver. Eur. J. Hear. Fail. Suppl. 2003, 2, 42. [Google Scholar] [CrossRef]
- Selimoglu, M.A.; Aydogdu, S.; Yagci, R.V.; Huseyinov, A. Plasma and liver carnitine status of children with chronic liver disease and cirrhosis. Pediatr. Int. 2001, 43, 391–395. [Google Scholar] [CrossRef]
- Rudman, D.; Sewell, C.W.; Ansley, J.D. Deficiency of carnitine in cachectic cirrhotic patients. J. Clin. Investig. 1977, 60, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Enooku, K.; Nakagawa, H.; Fujiwara, N.; Kondo, M.; Minami, T.; Hoshida, Y.; Shibahara, J.; Tateishi, R.; Koike, K. Altered serum acylcarnitine profile is associated with the status of nonalcoholic fatty liver disease (NAFLD) and NAFLD-related hepatocellular carcinoma. Sci. Rep. 2019, 9, 10663. [Google Scholar] [CrossRef] [PubMed]
- Lieber, C.S.; Lefèvre, A.; Spritz, N.; Feinman, L.; DeCarli, L.M. Difference in hepatic metabolism of long- and medium-chain fatty acids: The role of fatty acid chain length in the production of the alcoholic fatty liver. J. Clin. Invest. 1967, 46, 1451–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuller, R.K.; Hoppel, C.L. Elevated plasma carnitine in hepatic cirrhosis. Hepatology 2007, 3, 554–558. [Google Scholar] [CrossRef] [PubMed]
- Krahenbuhl, S.; Reichen, J. Carnitine metabolism in patients with chronic liver disease. Hepatology 1997, 25, 148–153. [Google Scholar] [CrossRef]
- Su, C.C.; Chang, C.S.; Chou, C.H.; Wu, Y.H.S.; Yang, K.T.; Tseng, J.K.; Chang, Y.Y.; Chen, Y.C. L-carnitine ameliorates dyslipidemic and hepatic disorders induced by a high-fat diet via regulating lipid metabolism, self-antioxidant capacity, and inflammatory response. J. Funct. Foods 2015. [Google Scholar] [CrossRef]
- Karlic, H.; Lohninger, S.; Koeck, T.; Lohninger, A. Dietary L-carnitine stimulates carnitine acyltransferases in the liver of aged rats. J. Histochem. Cytochem. 2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.-W.; Addy, C.; Kusunoki, J.; Anderson, N.N.; Deja, S.; Fu, X.; Burgess, S.C.; Li, C.; Ruddy, M.; Chakravarthy, M.; et al. Acetyl CoA carboxylase inhibition reduces hepatic steatosis but elevates plasma triglycerides in mice and humans: A bedside to bench investigation. Cell Metab. 2017, 26, 394–406. [Google Scholar] [CrossRef]
- Somi, M.H.; Fatahi, E.; Panahi, J.; Havasian, M.R.; Judaki, A. Data from a randomized and controlled trial of LCarnitine prescription for the treatment for Non-Alcoholic Fatty Liver Disease. Bioinformation 2014, 10, 575–579. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Liang, K.; Zhao, S.; Jia, W.; Liu, Y.; Wu, H.; Lv, J.; Cao, C.; Chen, T.; Zhuang, S.; et al. Chemoproteomics reveals baicalin activates hepatic CPT1 to ameliorate diet-induced obesity and hepatic steatosis. Proc. Natl. Acad. Sci. USA 2018. [Google Scholar] [CrossRef] [Green Version]
- Rinella, M.; Alonso, E.; Rao, S.; Whitington, P.; Fryer, J.; Abecassis, M.; Superina, R.; Flamm, S.L.; Blei, A.T. Body mass index as a predictor of hepatic steatosis in living liver donors. Liver Transplant. 2001, 7, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Angulo, P.; Keach, J.C.; Batts, K.P.; Lindor, K.D. Independent predictors of liver fibrosis in patients with nonalcoholic steatohepatitis. Hepatology 1999, 30, 1356–1362. [Google Scholar] [CrossRef] [PubMed]
- LaBrecque, D.R.; Abbas, Z.; Anania, F.; Ferenci, P.; Khan, A.G.; Goh, K.-L.; Hamid, S.S.; Isakov, V.; Lizarzabal, M.; Peñaranda, M.M.; et al. World gastroenterology organisation global guidelines. J. Clin. Gastroenterol. 2014, 48, 467–473. [Google Scholar] [CrossRef]
- Savic, D.; Ball, V.; Pavlides, M.; Heather, L.C.; Tyler, D.J. Linking diabetic cardiovascular disease with non-alcoholic fatty liver disease through L-carnitine: A hyperpolarized MRS study. In Proceedings of the International Liver Conference, Vienna, Austria, 14 April 2019. [Google Scholar]
- Malaguarnera, M.; Pistone, G.; Elvira, R.; Leotta, C.; Scarpello, L.; Liborio, R. Effects of L-carnitine in patients with hepatic encephalopathy. World J. Gastroenterol. 2005, 11, 7197–7202. [Google Scholar] [CrossRef] [PubMed]
- Hanai, T.; Shiraki, M.; Nishimura, K.; Imai, K.; Suetsugu, A.; Takai, K.; Shimizu, M.; Naiki, T.; Moriwaki, H. Free fatty acid as a marker of energy malnutrition in liver cirrhosis. Hepatol. Res. 2014. [Google Scholar] [CrossRef]
- Alavinejad, P.; Alavinejad, P.; Zakerkish, M.; Hajiani, E.; Hashemi, S.J.; Chobineh, M.; Moghaddam, E.K. Evaluation of L-carnitine efficacy in the treatment of non-alcoholic fatty liver disease among diabetic patients: A randomized double blind pilot study. J. Gastroenterol. Hepatol. Res. 2016, 5, 2191–2195. [Google Scholar] [CrossRef]
- Lim, C.Y.; Jun, D.W.; Jang, S.S.; Cho, W.K.; Chae, J.D.; Jun, J.H. Effects of carnitine on peripheral blood mitochondrial DNA copy number and liver function in non-alcoholic fatty liver disease. Korean J. Gastroenterol. 2010, 55, 384–389. [Google Scholar] [CrossRef] [Green Version]
- Sakai, Y.; Nishikawa, H.; Enomoto, H.; Yoh, K.; Iwata, Y.; Hasegawa, K.; Nakano, C.; Kishino, K.; Shimono, Y.; Takata, R.; et al. Effect of L-carnitine in patients with liver cirrhosis on energy metabolism using indirect calorimetry: A pilot study. J. Clin. Med. Res. 2016. [Google Scholar] [CrossRef] [Green Version]
- Schwenk, W.F.; Hale, D.E.; Haymond, M.W. Decreased fasting free fatty acids with L-carnitine in children with carnitine deficiency. Pediatr. Res. 1988, 23, 491–494. [Google Scholar] [CrossRef] [Green Version]
- Isaeva, A.P.; Gapparova, K.M. The effect of L-carnitine on lipid metabolism in patients with obesity. Clin. Nutr. 2018, 37, S38–S39. [Google Scholar] [CrossRef]
- Zillikens, M.C.; Swart, G.R.; Wilson, J.H.P. Metabolic Effects of L-Carnitine in Alcoholic Cirrhosis. J. Hepatol. 1988, 7, S200. [Google Scholar] [CrossRef]
- Xia, Y.; Li, Q.; Zhong, W.; Dong, J.; Wang, Z.; Wang, C. L-carnitine ameliorated fatty liver in high-calorie diet/STZ-induced type 2 diabetic mice by improving mitochondrial function. Diabetol. Metab. Syndr. 2011. [Google Scholar] [CrossRef] [Green Version]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, T.; Horiuchi, M.; Yamanaka, H.; Kizaki, Z.; Inoue, F.; Kodo, N.; Kinugasa, A.; Saheki, T.; Sawada, T. The effect of carnitine on ketogenesis in perfused livers from juvenile visceral steatosis mice with systemic carnitine deficiency. Pediatr. Res. 1997, 42, 108–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcgarry, J.D.; Robles-Valdes, C.; Foster, D.W. Role of carnitine in hepatic ketogenesis. Proc. Natl. Acad. Sci. USA 1975, 72, 4385–4388. [Google Scholar] [CrossRef] [Green Version]
- Karpati, G.; Carpenter, S.; Engel, A.G.; Watters, G.; Allen, J.; Rothman, S.; Klassen, G.; Mamer, O.A. The syndrome of systemic carnitine deficiency. Clinical, morphologic, biochemical, and pathophysiologic features. Neurology 1975, 25, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Gravina, E.; Gravina-Sanvitale, G. Effect of carnitine on blood acetoacetate in fasting children. Clin. Chim. Acta 1969, 23, 376–377. [Google Scholar] [CrossRef]
- Blanchi, P.B.; Lehotay, D.C.; Davis, A.T. Carnitine supplementation ameliorates the steatosis and ketosis induced by pivalate in rats. J. Nutr. 1996, 126, 2873–2879. [Google Scholar]
- Paulson, D.J.; Hoganson, G.E.; Traxler, J.; Sufit, R.; Peters, H.; Shug, A.L. Ketogenic effects of carnitine in patients with muscular dystrophy and cytochrome oxidase deficiency. Biochem. Med. Metab. Biol. 1988. [Google Scholar] [CrossRef]
- Waber, L.J.; Valle, D.; Neill, C.; DiMauro, S.; Shug, A. Carnitine deficiency presenting as familial cardiomyopathy: A treatable defect in carnitine transport. J. Pediatr. 1982. [Google Scholar] [CrossRef]
- Chalasani, N.; Deeg, M.A.; Persohn, S.; Crabb, D.W. Metabolic and anthropometric evaluation of insulin resistance in nondiabetic patients with nonalcoholic steatohepatitis. Am. J. Gastroenterol. 2003, 98, 1849–1855. [Google Scholar] [CrossRef]
- Bugianesi, E.; Gastaldelli, A.; Vanni, E.; Gambino, R.; Cassader, M.; Baldi, S.; Ponti, V.; Pagano, G.; Ferrannini, E.; Rizzetto, M. Insulin resistance in non-diabetic patients with non-alcoholic fatty liver disease: Sites and mechanisms. Diabetologia 2005, 48, 634–642. [Google Scholar] [CrossRef] [Green Version]
- Broderick, T.L.; Quinney, H.A.; Lopaschuk, G.D. Carnitine stimulation of glucose oxidation in the fatty acid perfused isolated working rat heart. J. Biol. Chem. 1992, 267, 3758–3763. [Google Scholar]
- Randle, P.J.; Garland, P.B.; Hales, C.N.; Newsholme, E.A. The glucose fatty-acid cycle its role in insulin sensitivity and the metabolic disturbances of diabetes mellituS. Lancet 1963, 281, 785–789. [Google Scholar] [CrossRef]
- Hue, L.; Taegtmeyer, H. The Randle cycle revisited: A new head for an old hat. AJP Endocrinol. Metab. 2009, 297, E578–E591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruggenenti, P.; Cattaneo, D.; Loriga, G.; Ledda, F.; Motterlini, N.; Gherardi, G.; Orisio, S.; Remuzzi, G. Ameliorating hypertension and insulin resistance in subjects at increased cardiovascular risk. Hypertension 2009, 54, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Grandi, M.; Pederzoli, S.; Sacchetti, C. Effect of acute carnitine administration on glucose insulin metabolism in healthy subjects. Int. J. Clin. Pharmacol. Res. 1997, 17, 143–147. [Google Scholar] [PubMed]
- Uziel, G.; Garavaglia, B.; Di Donato, S. Carnitine stimulation of pyruvate dehydrogenase complex (PDHC) in isolated human skeletal muscle mitochondria. Muscle Nerve 1988. [Google Scholar] [CrossRef] [PubMed]
- Keller, K.; Ringseis, R.; Priebe, S.; Guthke, R.; Kluge, H.; Eder, K. Transcript profiling in the liver of piglets fed L-carnitine. Nutr. Metab. 2011, 8, 76. [Google Scholar] [CrossRef] [Green Version]
- Molfino, A.; Cascino, A.; Ramaccini, C.; Conte, C.; Fanelli, F.R.; Laviano, A. L-carnitine administration improves insulin sensitivity in patients with impaired glucose metabolism. Eur. J. Intern. Med. 2008, 19, S48. [Google Scholar] [CrossRef]
- Angelini, A.; Imparato, L.; Landi, C.; Porfido, F.A.; Ciarimboli, M.; Marro, A. Variation in levels of glycaemia and insulin after infusion of glucose solutions with or without added L-carnitine. Drugs Exp. Clin. Res. 1993, 19, 219–222. [Google Scholar] [PubMed]
- Stephens, F.B.; Constantin-Teodosiu, D.; Laithwaite, D.; Simpson, E.J.; Greenhaff, P.L. Insulin stimulates L-carnitine accumulation in human skeletal muscle. FASEB J. 2006, 20, 377–379. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Ohn, J.H.; Jung, H.S.; Park, Y.J.; Jang, H.C.; Chung, S.S.; Park, K.S. Carnitine induces autophagy and restores high-fat diet-induced mitochondrial dysfunction. Metabolism. Clin. Exp. 2018. [Google Scholar] [CrossRef] [PubMed]
- de Luca, C.; Olefsky, J.M. Inflammation and insulin resistance. FEBS Lett. 2008, 582, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mingrone, G.; Greco, A.V.; Capristo, E.; Benedetti, G.; Giancaterini, A.; De Gaetano, A.; Gasbarrini, G. L-carnitine improves glucose disposal in type 2 diabetic patients. J. Am. Coll. Nutr. 1999. [Google Scholar] [CrossRef] [PubMed]
- Paulson, D.J.; Schmidt, M.J.; Traxler, J.S.; Ramacci, M.T.; Shug, A.L. Improvement of myocardial function in diabetic rats after treatment with L-carnitine. Metabolism 1984, 33, 358–363. [Google Scholar] [CrossRef]
- Aguer, C.; McCoin, C.S.; Knotts, T.A.; Thrush, A.B.; Ono-Moore, K.; McPherson, R.; Dent, R.; Hwang, D.H.; Adams, S.H.; Harper, M.-E. Acylcarnitines: Potential implications for skeletal muscle insulin resistance. FASEB J. 2015, 29, 336–345. [Google Scholar] [CrossRef] [Green Version]
- Schooneman, M.G.; Vaz, F.M.; Houten, S.M.; Soeters, M.R. Acylcarnitines: Reflecting or inflicting insulin resistance? Diabetes 2013, 62, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Mihalik, S.J.; Goodpaster, B.H.; Kelley, D.E.; Chace, D.H.; Vockley, J.; Toledo, F.G.S.; DeLany, J.P. Increased levels of plasma acylcarnitines in obesity and type 2 diabetes and identification of a marker of glucolipotoxicity. Obesity 2010, 18, 1695–1700. [Google Scholar] [CrossRef] [Green Version]
- Koo, S.-H. Nonalcoholic fatty liver disease: Molecular mechanisms for the hepatic steatosis. Clin. Mol. Hepatol. 2013, 19, 210–215. [Google Scholar] [CrossRef]
- Liu, Q.; Bengmark, S.; Qu, S. The role of hepatic fat accumulation in pathogenesis of non-alcoholic fatty liver disease (NAFLD). Lipids Health Dis. 2010, 9, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanyal, D.; Mukherjee, P.; Raychaudhuri, M.; Ghosh, S.; Mukherjee, S.; Chowdhury, S. Profile of liver enzymes in non-alcoholic fatty liver disease in patients with impaired glucose tolerance and newly detected untreated type 2 diabetes. Indian J. Endocrinol. Metab. 2015, 19, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Go, Y.; Jeong, J.Y.; Jeoung, N.H.; Jeon, J.-H.; Park, B.-Y.; Kang, H.-J.; Ha, C.-M.; Choi, Y.-K.; Lee, S.J.; Ham, H.J.; et al. Inhibition of pyruvate dehydrogenase kinase 2 protects against hepatic steatosis through modulation of tricarboxylic acid cycle anaplerosis and ketogenesis. Diabetes 2016, 65, 2876–2887. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Author | Type of Study | Patients | Duration | Dose | Disease | Results |
|---|---|---|---|---|---|---|
| Alavinejadet al. [79] | Randomized double blind pilot study | 60 | 3 months | 750 mg of L-carnitine tablets | NAFLD + Diabetes | Reduced ALT levels |
| Somi et al. [71] | Placebo controlled trial | 80 | 24 weeks | 500 mg of L-carnitine twice/day | NAFLD | Reduced ALT, AST, BW Improved sonographic grade |
| Lim et al. [80] | Pilot Study | 45 | 3 months | 600 mg/day of L-carnitine | NAFLD | Reduced ALT, AST, bilirubin. Elevated peripheral mitochondrial copy number |
| Bae et al. [11] | Randomized control trial | 78 | 12 weeks | 150 mg of carnitine-orotate | NAFLD + diabetes | Reduced liver attenuation index on CT Reduced ALT, AST and HbA1c levels |
| Malaguarna et al. [10] | Randomized double-blind placebo-controlled trial | 74 | 24 weeks | 1 g of oral L-carnitine twice/day | NASH | Reduced liver inflammation (biopsies) Reduced serum TNF-alpha & CRP Reduced insulin, LDL, cholesterol, TG. |
| Cecere et al. [9] | Randomized controlled clinical trial | 31 | 4 weeks | 6 g/day of L-carnitine | Cirrhosis | Reduced ammonia levels, improved psychometric test. Improved neurological function |
| Sakai et al. [81] | Pilot Study | 13 | 4 weeks | 1800 mg/day | Reduced Free FA Elevated whole-body carbohydrate oxidation. Decreased FA and protein oxidation |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savic, D.; Hodson, L.; Neubauer, S.; Pavlides, M. The Importance of the Fatty Acid Transporter L-Carnitine in Non-Alcoholic Fatty Liver Disease (NAFLD). Nutrients 2020, 12, 2178. https://doi.org/10.3390/nu12082178
Savic D, Hodson L, Neubauer S, Pavlides M. The Importance of the Fatty Acid Transporter L-Carnitine in Non-Alcoholic Fatty Liver Disease (NAFLD). Nutrients. 2020; 12(8):2178. https://doi.org/10.3390/nu12082178
Chicago/Turabian StyleSavic, Dragana, Leanne Hodson, Stefan Neubauer, and Michael Pavlides. 2020. "The Importance of the Fatty Acid Transporter L-Carnitine in Non-Alcoholic Fatty Liver Disease (NAFLD)" Nutrients 12, no. 8: 2178. https://doi.org/10.3390/nu12082178
APA StyleSavic, D., Hodson, L., Neubauer, S., & Pavlides, M. (2020). The Importance of the Fatty Acid Transporter L-Carnitine in Non-Alcoholic Fatty Liver Disease (NAFLD). Nutrients, 12(8), 2178. https://doi.org/10.3390/nu12082178