Probiotic Supplementation during the Perinatal and Infant Period: Effects on gut Dysbiosis and Disease

 , ,
, ,  and
and 
Abstract
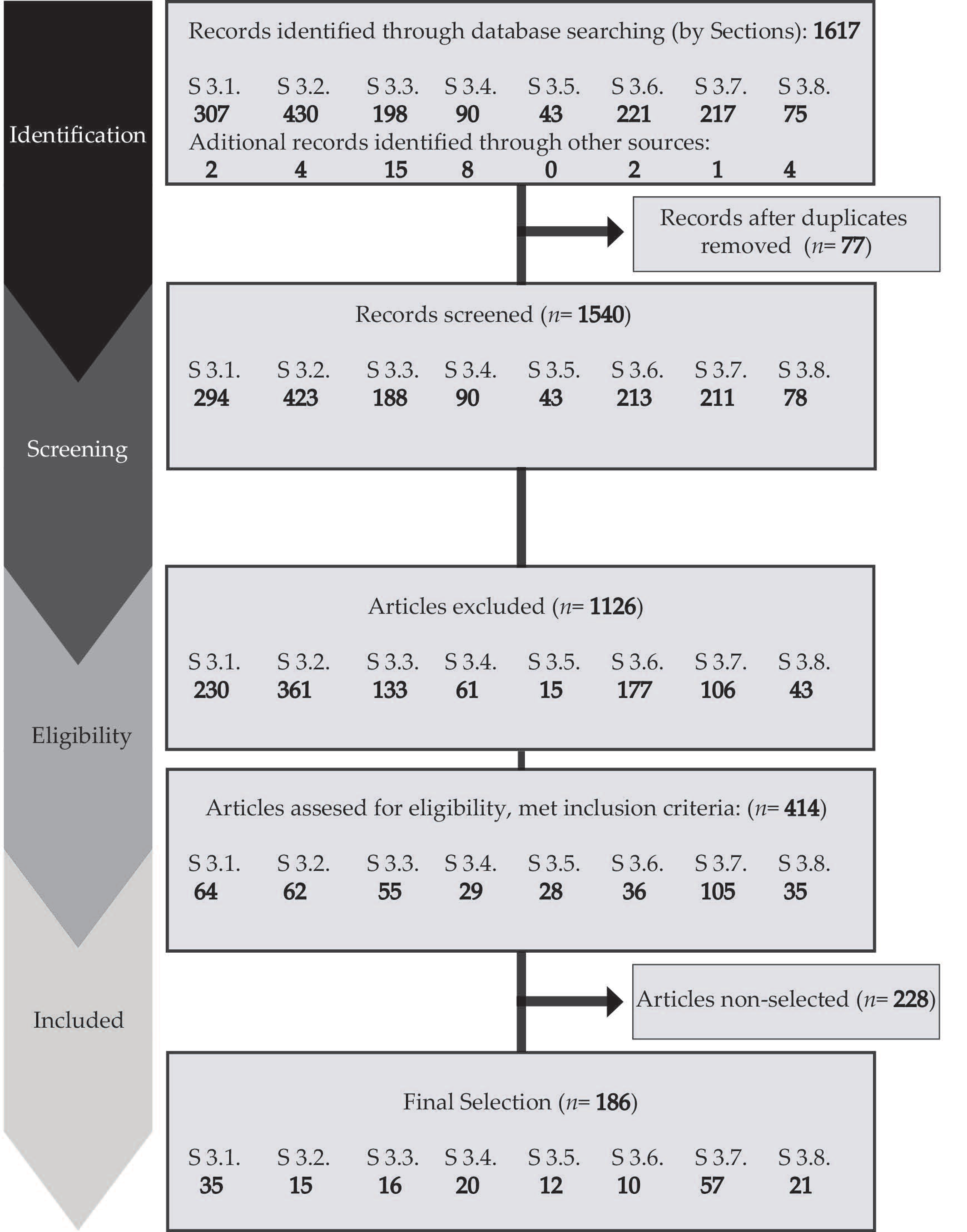
:1. Introduction
2. Materials and Methods
3. Results
3.1. Prenatal Development of the Microbiome and Early Colonization
3.1.1. Gut Colonization
3.1.2. Respiratory Colonization
3.1.3. Skin
3.2. Gut Dysbiosis Induced by Antibiotic and Nonantibiotic Medications
3.3. Early Aberrant Microbiota and Its Effect on Pediatric Diseases
3.4. Transfer of Probiotic Bacteria from Mother to Child
3.5. Probiotics for the Prevention of Food Sensitization in Infants: Administration to Mothers Versus Infants
3.6. Probiotics for Prevention of Asthma/Wheezing and Rhinitis: Administration to Mothers Versus Infants
3.7. The Use of Probiotics and Paraprobiotics in Preterm Neonates
3.8. Safety of Probiotics in Pregnancy and Neonatal Period
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Al Alam, D.; Danopoulos, S.; Grubbs, B.; Ali, N.A.t.B.M.; MacAogain, M.; Chotirmall, S.H.; Warburton, D.; Gaggar, A.; Ambalavanan, N.; Lal, C.V. Human fetal lungs harbor a microbiome signature. Am. J. Respir. Crit. Care Med. 2020, 201, 1002–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prince, A.L.; Chu, D.M.; Seferovic, M.D.; Antony, K.M.; Ma, J.; Aagaard, K.M. The perinatal microbiome and pregnancy: Moving beyond the vaginal microbiome. Cold Spring Harb. Perspect. Med. 2015, 5, a023051. [Google Scholar] [CrossRef]
- Satokari, R.; Grönroos, T.; Laitinen, K.; Salminen, S.; Isolauri, E. Bifidobacterium and Lactobacillus DNA in the human placenta. Lett. Appl. Microbiol. 2009, 48, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, E.; Marín, M.L.; Martín, R.; Odriozola, J.M.; Olivares, M.; Xaus, J.; Fernández, L.; Rodríguez, J.M. Is meconium from healthy newborns actually sterile? Res. Microbiol. 2008, 159, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Rautava, S.; Aakko, J.; Isolauri, E.; Salminen, S. Human gut colonisation may be initiated in utero by distinct microbial communities in the placenta and amniotic fluid. Sci. Rep. 2016, 6, 23129. [Google Scholar] [CrossRef] [Green Version]
- Bearfield, C.; Davenport, E.S.; Sivapathasundaram, V.; Allaker, R.P. Possible association between amniotic fluid micro-organism infection and microflora in the mouth. Bjog Int. J. Obstet. Gynaecol. 2002, 109, 527–533. [Google Scholar] [CrossRef]
- Mai, V.; Young, C.M.; Ukhanova, M.; Wang, X.; Sun, Y.; Casella, G.; Theriaque, D.; Li, N.; Sharma, R.; Hudak, M. Fecal microbiota in premature infants prior to necrotizing enterocolitis. PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Levy, I.; Comarsca, J.; Davidovits, M.; Klinger, G.; Sirota, L.; Linder, N. Urinary tract infection in preterm infants: the protective role of breastfeeding. Pediatric Nephrol. 2009, 24, 527–531. [Google Scholar] [CrossRef]
- Ward, D.V.; Scholz, M.; Zolfo, M.; Taft, D.H.; Schibler, K.R.; Tett, A.; Segata, N.; Morrow, A.L. Metagenomic sequencing with strain-level resolution implicates uropathogenic E. coli in necrotizing enterocolitis and mortality in preterm infants. Cell Rep. 2016, 14, 2912–2924. [Google Scholar] [CrossRef] [Green Version]
- Greenwood, C.; Morrow, A.L.; Lagomarcino, A.J.; Altaye, M.; Taft, D.H.; Yu, Z.; Newburg, D.S.; Ward, D.V.; Schibler, K.R. Early empiric antibiotic use in preterm infants is associated with lower bacterial diversity and higher relative abundance of Enterobacter. J. Pediatrics 2014, 165, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Sjögren, Y.M.; Jenmalm, M.C.; Böttcher, M.F.; Björkstén, B.; Sverremark-Ekström, E. Altered early infant gut microbiota in children developing allergy up to 5 years of age. Clin. Exp. Allergy 2009, 39, 518–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azad, M.B.; Konya, T.; Guttman, D.S.; Field, C.; Sears, M.; HayGlass, K.; Mandhane, P.; Turvey, S.; Subbarao, P.; Becker, A. Infant gut microbiota and food sensitization: Associations in the first year of life. Clin. Exp. Allergy 2015, 45, 632–643. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.H. Revisiting the hygiene hypothesis for allergy and asthma. J. Allergy Clin. Immunol. 2015, 136, 860–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björkstén, B.; Sepp, E.; Julge, K.; Voor, T.; Mikelsaar, M. Allergy development and the intestinal microflora during the first year of life. J. Allergy Clin. Immunol. 2001, 108, 516–520. [Google Scholar] [CrossRef]
- Owaga, E.E.; Elbakkoush, A.; KS MS, L.R. Antiallergic effects of probiotic lactobacilli–cellular and molecular mechanisms. J Microbiol Res 2014, 4, 92–97. [Google Scholar]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. Ann. Intern. Med. 2009, 151, W-65–W-94. [Google Scholar] [CrossRef] [Green Version]
- Shamseer, L.; Moher, D.; Clarke, M.; Ghersi, D.; Liberati, A.; Petticrew, M.; Shekelle, P.; Stewart, L.A. Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015: Elaboration and explanation. BMJ 2015, 349. [Google Scholar] [CrossRef] [Green Version]
- Higgins, J.; Altman, D.; Gøtzsche, P.; Jüni, P.; Moher, D.; Oxman, A.; Savovic, J.; Schulz, K.; Weeks, L.; Sterne, J. The Cochrane Collaboration’s tool for assessing risk of bias in randomised trials. BMJ 2011, 343, d5928. [Google Scholar]
- Nagpal, R.; Mainali, R.; Ahmadi, S.; Wang, S.; Singh, R.; Kavanagh, K.; Kitzman, D.W.; Kushugulova, A.; Marotta, F.; Yadav, H. Gut microbiome and aging: Physiological and mechanistic insights. Nutr. Healthy Aging 2018, 4, 267–285. [Google Scholar] [CrossRef] [Green Version]
- Avershina, E.; Storrø, O.; Øien, T.; Johnsen, R.; Pope, P.; Rudi, K. Major faecal microbiota shifts in composition and diversity with age in a geographically restricted cohort of mothers and their children. Fems Microbiol. Ecol. 2014, 87, 280–290. [Google Scholar] [CrossRef]
- Tapiainen, T.; Paalanne, N.; Tejesvi, M.V.; Koivusaari, P.; Korpela, K.; Pokka, T.; Salo, J.; Kaukola, T.; Pirttilä, A.M.; Uhari, M. Maternal influence on the fetal microbiome in a population-based study of the first-pass meconium. Pediatric Res. 2018, 84. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Nomura, Y.; Bashir, A.; Fernandez-Hernandez, H.; Itzkowitz, S.; Pei, Z.; Stone, J.; Loudon, H.; Peter, I. Diversified microbiota of meconium is affected by maternal diabetes status. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, D.M.; Antony, K.M.; Ma, J.; Prince, A.L.; Showalter, L.; Moller, M.; Aagaard, K.M. The early infant gut microbiome varies in association with a maternal high-fat diet. Genome Med. 2016, 8, 77. [Google Scholar] [CrossRef] [Green Version]
- Lundgren, S.N.; Madan, J.C.; Emond, J.A.; Morrison, H.G.; Christensen, B.C.; Karagas, M.R.; Hoen, A.G. Maternal diet during pregnancy is related with the infant stool microbiome in a delivery mode-dependent manner. Microbiome 2018, 6, 109. [Google Scholar] [CrossRef] [Green Version]
- Zijlmans, M.A.; Korpela, K.; Riksen-Walraven, J.M.; de Vos, W.M.; de Weerth, C. Maternal prenatal stress is associated with the infant intestinal microbiota. Psychoneuroendocrinology 2015, 53, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Isolauri, E.; Laitinen, K.; Salminen, S. Effect of mother’s weight on infant’s microbiota acquisition, composition, and activity during early infancy: A prospective follow-up study initiated in early pregnancy. Am. J. Clin. Nutr. 2010, 92, 1023–1030. [Google Scholar] [CrossRef]
- Stanislawski, M.A.; Dabelea, D.; Wagner, B.D.; Sontag, M.K.; Lozupone, C.A.; Eggesbø, M. Pre-pregnancy weight, gestational weight gain, and the gut microbiota of mothers and their infants. Microbiome 2017, 5, 113. [Google Scholar] [CrossRef]
- de Goffau, M.C.; Lager, S.; Sovio, U.; Gaccioli, F.; Cook, E.; Peacock, S.J.; Parkhill, J.; Charnock-Jones, D.S.; Smith, G.C. Human placenta has no microbiome but can contain potential pathogens. Nature 2019, 572, 329–334. [Google Scholar] [CrossRef]
- Korpela, K.; Costea, P.; Coelho, L.P.; Kandels-Lewis, S.; Willemsen, G.; Boomsma, D.I.; Segata, N.; Bork, P. Selective maternal seeding and environment shape the human gut microbiome. Genome Res. 2018, 28, 561–568. [Google Scholar] [CrossRef] [Green Version]
- Makino, H.; Kushiro, A.; Ishikawa, E.; Kubota, H.; Gawad, A.; Sakai, T.; Oishi, K.; Martin, R.; Ben-Amor, K.; Knol, J. Mother-to-infant transmission of intestinal bifidobacterial strains has an impact on the early development of vaginally delivered infant’s microbiota. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Biasucci, G.; Benenati, B.; Morelli, L.; Bessi, E.; Boehm, G.n. Cesarean delivery may affect the early biodiversity of intestinal bacteria. J. Nutr. 2008, 138, 1796S–1800S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fallani, M.; Young, D.; Scott, J.; Norin, E.; Amarri, S.; Adam, R.; Aguilera, M.; Khanna, S.; Gil, A.; Edwards, C.A. Intestinal microbiota of 6-week-old infants across Europe: Geographic influence beyond delivery mode, breast-feeding, and antibiotics. J. Pediatric Gastroenterol. Nutr. 2010, 51, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Tuominen, H.; Collado, M.C.; Rautava, J.; Syrjänen, S.; Rautava, S. Composition and maternal origin of the neonatal oral cavity microbiota. J. Oral Microbiol. 2019, 11, 1663084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Chen, S.; Wu, L.; Wang, H.; Xiao, K.; Gao, Y.; Li, Y.; Li, H.; Xiao, B.; Zhu, Y. The effects of perineal disinfection on infant’s oral microflora after transvaginal examination during delivery. BMC Pregnancy Childbirth 2019, 19, 213. [Google Scholar] [CrossRef] [PubMed]
- Hesla, H.M.; Stenius, F.; Jäderlund, L.; Nelson, R.; Engstrand, L.; Alm, J.; Dicksved, J. Impact of lifestyle on the gut microbiota of healthy infants and their mothers–the ALADDIN birth cohort. Fems Microbiol. Ecol. 2014, 90, 791–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huurre, A.; Kalliomäki, M.; Rautava, S.; Rinne, M.; Salminen, S.; Isolauri, E. Mode of delivery–effects on gut microbiota and humoral immunity. Neonatology 2008, 93, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Korpela, K.; Salonen, A.; Hickman, B.; Kunz, C.; Sprenger, N.; Kukkonen, K.; Savilahti, E.; Kuitunen, M.; de Vos, W.M. Fucosylated oligosaccharides in mother’s milk alleviate the effects of caesarean birth on infant gut microbiota. Sci. Rep. 2018, 8, 1–7. [Google Scholar]
- Brumbaugh, D.E.; Arruda, J.; Robbins, K.; Ir, D.; Santorico, S.A.; Robertson, C.E.; Frank, D.N. Mode of delivery determines neonatal pharyngeal bacterial composition and early intestinal colonization. J. Pediatric Gastroenterol. Nutr. 2016, 63, 320–328. [Google Scholar] [CrossRef]
- Hill, C.J.; Lynch, D.B.; Murphy, K.; Ulaszewska, M.; Jeffery, I.B.; O’Shea, C.A.; Watkins, C.; Dempsey, E.; Mattivi, F.; Tuohy, K. Evolution of gut microbiota composition from birth to 24 weeks in the INFANTMET Cohort. Microbiome 2017, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Lif Holgerson, P.; Harnevik, L.; Hernell, O.; Tanner, A.; Johansson, I. Mode of birth delivery affects oral microbiota in infants. J. Dent. Res. 2011, 90, 1183–1188. [Google Scholar] [CrossRef] [Green Version]
- Martin, R.; Makino, H.; Yavuz, A.C.; Ben-Amor, K.; Roelofs, M.; Ishikawa, E.; Kubota, H.; Swinkels, S.; Sakai, T.; Oishi, K. Early-life events, including mode of delivery and type of feeding, siblings and gender, shape the developing gut microbiota. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Stokholm, J.; Thorsen, J.; Chawes, B.L.; Schjørring, S.; Krogfelt, K.A.; Bønnelykke, K.; Bisgaard, H. Cesarean section changes neonatal gut colonization. J. Allergy Clin. Immunol. 2016, 138, 881–889. e882. [Google Scholar] [CrossRef] [PubMed]
- Gregory, K.E.; LaPlante, R.D.; Shan, G.; Kumar, D.V.; Gregas, M. Mode of birth influences preterm infant intestinal colonization with bacteroides over the early neonatal period. Adv. Neonatal Care Off. J. Natl. Assoc. Neonatal Nurses 2015, 15, 386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahl, C.; Stigum, H.; Valeur, J.; Iszatt, N.; Lenters, V.; Peddada, S.; Bjørnholt, J.V.; Midtvedt, T.; Mandal, S.; Eggesbø, M. Preterm infants have distinct microbiomes not explained by mode of delivery, breastfeeding duration or antibiotic exposure. Int. J. Epidemiol. 2018, 47, 1658–1669. [Google Scholar] [CrossRef] [PubMed]
- Fouhy, F.; Watkins, C.; Hill, C.J.; O’Shea, C.-A.; Nagle, B.; Dempsey, E.M.; O’Toole, P.W.; Ross, R.P.; Ryan, C.A.; Stanton, C. Perinatal factors affect the gut microbiota up to four years after birth. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grier, A.; McDavid, A.; Wang, B.; Qiu, X.; Java, J.; Bandyopadhyay, S.; Yang, H.; Holden-Wiltse, J.; Kessler, H.A.; Gill, A.L. Neonatal gut and respiratory microbiota: Coordinated development through time and space. Microbiome 2018, 6, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Chernikova, D.A.; Madan, J.C.; Housman, M.L.; Lundgren, S.N.; Morrison, H.G.; Sogin, M.L.; Williams, S.M.; Moore, J.H.; Karagas, M.R.; Hoen, A.G. The premature infant gut microbiome during the first 6 weeks of life differs based on gestational maturity at birth. Pediatric Res. 2018, 84, 71–79. [Google Scholar] [CrossRef]
- Forsgren, M.; Isolauri, E.; Salminen, S.; Rautava, S. Late preterm birth has direct and indirect effects on infant gut microbiota development during the first six months of life. Acta Paediatr. 2017, 106, 1103–1109. [Google Scholar] [CrossRef] [Green Version]
- Shilts, M.H.; Rosas-Salazar, C.; Tovchigrechko, A.; Larkin, E.K.; Torralba, M.; Akopov, A.; Halpin, R.; Peebles, R.S.; Moore, M.L.; Anderson, L.J. Minimally invasive sampling method identifies differences in taxonomic richness of nasal microbiomes in young infants associated with mode of delivery. Microb. Ecol. 2016, 71, 233–242. [Google Scholar] [CrossRef] [Green Version]
- Bosch, A.A.; Levin, E.; van Houten, M.A.; Hasrat, R.; Kalkman, G.; Biesbroek, G.; de Steenhuijsen Piters, W.A.; de Groot, P.-K.C.; Pernet, P.; Keijser, B.J. Development of upper respiratory tract microbiota in infancy is affected by mode of delivery. EBioMedicine 2016, 9, 336–345. [Google Scholar] [CrossRef] [Green Version]
- Lohmann, P.; Luna, R.A.; Hollister, E.B.; Devaraj, S.; Mistretta, T.-A.; Welty, S.E.; Versalovic, J. The airway microbiome of intubated premature infants: Characteristics and changes that predict the development of bronchopulmonary dysplasia. Pediatric Res. 2014, 76, 294–301. [Google Scholar] [CrossRef] [Green Version]
- Marchini, G.; Nelson, A.; Edner, J.; Lonne-Rahm, S.; Stavréus-Evers, A.; Hultenby, K. Erythema toxicum neonatorum is an innate immune response to commensal microbes penetrated into the skin of the newborn infant. Pediatric Res. 2005, 58, 613–616. [Google Scholar] [CrossRef] [Green Version]
- Nelson, A.; Hultenby, K.; Hell, É.; Riedel, H.M.; Brismar, H.; Flock, J.-I.; Lundahl, J.; Giske, C.G.; Marchini, G. Staphylococcus epidermidis isolated from newborn infants express pilus-like structures and are inhibited by the cathelicidin-derived antimicrobial peptide LL37. Pediatric Res. 2009, 66, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Soeorg, H.; Metsvaht, T.; Eelmäe, I.; Merila, M.; Treumuth, S.; Huik, K.; Jürna-Ellam, M.; Ilmoja, M.-L.; Lutsar, I. The role of breast milk in the colonization of neonatal gut and skin with coagulase-negative staphylococci. Pediatric Res. 2017, 82, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Stokholm, J.; Schjørring, S.; Pedersen, L.; Bischoff, A.L.; Følsgaard, N.; Carson, C.G.; Chawes, B.L.; Bønnelykke, K.; Mølgaard, A.; Krogfelt, K.A. Prevalence and predictors of antibiotic administration during pregnancy and birth. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Alexander, V.N.; Northrup, V.; Bizzarro, M.J. Antibiotic exposure in the newborn intensive care unit and the risk of necrotizing enterocolitis. J. Pediatrics 2011, 159, 392–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuppala, V.S.; Meinzen-Derr, J.; Morrow, A.L.; Schibler, K.R. Prolonged initial empirical antibiotic treatment is associated with adverse outcomes in premature infants. J. Pediatrics 2011, 159, 720–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weintraub, A.; Ferrara, L.; Deluca, L.; Moshier, E.; Green, R.; Oakman, E.; Lee, M.; Rand, L. Antenatal antibiotic exposure in preterm infants with necrotizing enterocolitis. J. Perinatol. 2012, 32, 705–709. [Google Scholar] [CrossRef] [Green Version]
- Benjamin, D.K.; DeLong, E.R.; Steinbach, W.J.; Cotton, C.M.; Walsh, T.J.; Clark, R.H. Empirical therapy for neonatal candidemia in very low birth weight infants. Pediatrics 2003, 112, 543–547. [Google Scholar] [CrossRef]
- Aloisio, I.; Quagliariello, A.; De Fanti, S.; Luiselli, D.; De Filippo, C.; Albanese, D.; Corvaglia, L.T.; Faldella, G.; Di Gioia, D. Evaluation of the effects of intrapartum antibiotic prophylaxis on newborn intestinal microbiota using a sequencing approach targeted to multi hypervariable 16S rDNA regions. Appl. Microbiol. Biotechnol. 2016, 100, 5537–5546. [Google Scholar] [CrossRef]
- Corvaglia, L.; Tonti, G.; Martini, S.; Aceti, A.; Mazzola, G.; Aloisio, I.; Di Gioia, D.; Faldella, G. Influence of intrapartum antibiotic prophylaxis for group B streptococcus on gut microbiota in the first month of life. J. Pediatric Gastroenterol. Nutr. 2016, 62, 304–308. [Google Scholar] [CrossRef]
- Mazzola, G.; Murphy, K.; Ross, R.P.; Di Gioia, D.; Biavati, B.; Corvaglia, L.T.; Faldella, G.; Stanton, C. Early gut microbiota perturbations following intrapartum antibiotic prophylaxis to prevent group B streptococcal disease. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Nogacka, A.; Salazar, N.; Suárez, M.; Milani, C.; Arboleya, S.; Solís, G.; Fernández, N.; Alaez, L.; Hernández-Barranco, A.M.; Clara, G. Impact of intrapartum antimicrobial prophylaxis upon the intestinal microbiota and the prevalence of antibiotic resistance genes in vaginally delivered full-term neonates. Microbiome 2017, 5, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stearns, J.C.; Simioni, J.; Gunn, E.; McDonald, H.; Holloway, A.C.; Thabane, L.; Mousseau, A.; Schertzer, J.D.; Ratcliffe, E.M.; Rossi, L. Intrapartum antibiotics for GBS prophylaxis alter colonization patterns in the early infant gut microbiome of low risk infants. Sci. Rep. 2017, 7, 1–9. [Google Scholar]
- Azad, M.B.; Konya, T.; Persaud, R.R.; Guttman, D.S.; Chari, R.S.; Field, C.J.; Sears, M.R.; Mandhane, P.; Turvey, S.; Subbarao, P. Impact of maternal intrapartum antibiotics, method of birth and breastfeeding on gut microbiota during the first year of life: A prospective cohort study. Bjog Int. J. Obstet. Gynaecol. 2016, 123, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Dardas, M.; Gill, S.R.; Grier, A.; Pryhuber, G.S.; Gill, A.L.; Lee, Y.-H.; Guillet, R. The impact of postnatal antibiotics on the preterm intestinal microbiome. Pediatric Res. 2014, 76, 150–158. [Google Scholar] [CrossRef]
- Fouhy, F.; Guinane, C.M.; Hussey, S.; Wall, R.; Ryan, C.A.; Dempsey, E.M.; Murphy, B.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. High-throughput sequencing reveals the incomplete, short-term recovery of infant gut microbiota following parenteral antibiotic treatment with ampicillin and gentamicin. Antimicrob. Agents Chemother. 2012, 56, 5811–5820. [Google Scholar] [CrossRef] [Green Version]
- Zwittink, R.D.; Renes, I.B.; van Lingen, R.A.; van Zoeren-Grobben, D.; Konstanti, P.; Norbruis, O.F.; Martin, R.; Jebbink, L.J.G.; Knol, J.; Belzer, C. Association between duration of intravenous antibiotic administration and early-life microbiota development in late-preterm infants. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 475–483. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Xiao, S.; Yu, J.; Ai, Q.; He, Y.; Cheng, C.; Zhang, Y.; Pan, Y. Effects of one-week empirical antibiotic therapy on the early development of gut microbiota and metabolites in preterm infants. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Zou, Z.-H.; Liu, D.; Li, H.-D.; Zhu, D.-P.; He, Y.; Hou, T.; Yu, J.-L. Prenatal and postnatal antibiotic exposure influences the gut microbiota of preterm infants in neonatal intensive care units. Ann. Clin. Microbiol. Antimicrob. 2018, 17, 9. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Kobayashi, T.; Songjinda, P.; Tateyama, A.; Tsubouchi, M.; Kiyohara, C.; Shirakawa, T.; Sonomoto, K.; Nakayama, J. Influence of antibiotic exposure in the early postnatal period on the development of intestinal microbiota. FEMS Immunol. Med. Microbiol. 2009, 56, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Arboleya, S.; Sánchez, B.; Milani, C.; Duranti, S.; Solís, G.; Fernández, N.; Clara, G.; Ventura, M.; Margolles, A.; Gueimonde, M. Intestinal microbiota development in preterm neonates and effect of perinatal antibiotics. J. Pediatrics 2015, 166, 538–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neu, J.; Pammi, M. Pathogenesis of NEC: Impact of an altered intestinal microbiome. Semin. Perinatol. 2017, 41, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Manges, A.R.; Labbe, A.; Loo, V.G.; Atherton, J.K.; Behr, M.A.; Masson, L.; Tellis, P.A.; Brousseau, R. Comparative metagenomic study of alterations to the intestinal microbiota and risk of nosocomial Clostridum difficile-associated disease. J. Infect. Dis. 2010, 202, 1877–1884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pamer, E.G. Resurrecting the intestinal microbiota to combat antibiotic-resistant pathogens. Science 2016, 352, 535–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aloisio, I.; Mazzola, G.; Corvaglia, L.T.; Tonti, G.; Faldella, G.; Biavati, B.; Di Gioia, D. Influence of intrapartum antibiotic prophylaxis against group B Streptococcus on the early newborn gut composition and evaluation of the anti-Streptococcus activity of Bifidobacterium strains. Appl. Microbiol. Biotechnol. 2014, 98, 6051–6060. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Le Bastard, Q.; Al-Ghalith, G.; Grégoire, M.; Chapelet, G.; Javaudin, F.; Dailly, E.; Batard, E.; Knights, D.; Montassier, E. Systematic review: human gut dysbiosis induced by non-antibiotic prescription medications. Aliment. Pharmacol. Ther. 2018, 47, 332–345. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.S.; Springston, E.E.; Warrier, M.R.; Smith, B.; Kumar, R.; Pongracic, J.; Holl, J.L. The prevalence, severity, and distribution of childhood food allergy in the United States. Pediatrics 2011, 128, e9–e17. [Google Scholar] [CrossRef] [Green Version]
- Bruno, G.; Cantani, A.; Ragno, V.; Milita, O.; Ziruolo, G.; Businco, L. Natural history of IgE antibodies in children at risk for atopy. Ann. Allergyasthma Immunol. Off. Publ. Am. Coll. Allergy Asthma Immunol. 1995, 74, 431–436. [Google Scholar]
- Jakobsson, H.E.; Abrahamsson, T.R.; Jenmalm, M.C.; Harris, K.; Quince, C.; Jernberg, C.; Björkstén, B.; Engstrand, L.; Andersson, A.F. Decreased gut microbiota diversity, delayed Bacteroidetes colonisation and reduced Th1 responses in infants delivered by caesarean section. Gut 2014, 63, 559–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stokholm, J.; Schjørring, S.; Eskildsen, C.; Pedersen, L.; Bischoff, A.; Følsgaard, N.; Carson, C.; Chawes, B.; Bønnelykke, K.; Mølgaard, A. Antibiotic use during pregnancy alters the commensal vaginal microbiota. Clin. Microbiol. Infect. 2014, 20, 629–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, H.; Pamp, S.J.; Hill, J.A.; Surana, N.K.; Edelman, S.M.; Troy, E.B.; Reading, N.C.; Villablanca, E.J.; Wang, S.; Mora, J.R. Gut immune maturation depends on colonization with a host-specific microbiota. Cell 2012, 149, 1578–1593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahenzli, J.; Köller, Y.; Wyss, M.; Geuking, M.B.; McCoy, K.D. Intestinal microbial diversity during early-life colonization shapes long-term IgE levels. Cell Host Microbe 2013, 14, 559–570. [Google Scholar] [CrossRef] [Green Version]
- Forchielli, M.L.; Walker, W.A. The role of gut-associated lymphoid tissues and mucosal defence. Br. J. Nutr. 2005, 93, S41–S48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kachikis, A.; Englund, J.A. Maternal immunization: Optimizing protection for the mother and infant. J. Infect. 2016, 72, S83–S90. [Google Scholar] [CrossRef] [PubMed]
- Goldman, A.S.; Garza, C.; Nichols, B.L.; Goldblum, R.M. Immunologic factors in human milk during the first year of lactation. J. Pediatrics 1982, 100, 563–567. [Google Scholar] [CrossRef]
- Ximenez, C.; Torres, J. Development of microbiota in infants and its role in maturation of gut mucosa and immune system. Arch. Med Res. 2017, 48, 666–680. [Google Scholar] [CrossRef]
- Dzidic, M.; Abrahamsson, T.R.; Artacho, A.; Björkstén, B.; Collado, M.C.; Mira, A.; Jenmalm, M.C. Aberrant IgA responses to the gut microbiota during infancy precede asthma and allergy development. J. Allergy Clin. Immunol. 2017, 139, 1017–1025. e1014. [Google Scholar] [CrossRef] [Green Version]
- Gauhe, A.; Gyorgy, P.; Hoover, J.R.; Kuhn, R.; Rose, C.S.; Ruelius, H.W.; Zilliken, F. Bifidus factor. 4. Preparations obtained from human milk. Arch. Biochem. 1954, 48, 214–224. [Google Scholar] [CrossRef]
- Lawson, M.A.; O’Neill, I.J.; Kujawska, M.; Javvadi, S.G.; Wijeyesekera, A.; Flegg, Z.; Chalklen, L.; Hall, L.J. Breast milk-derived human milk oligosaccharides promote Bifidobacterium interactions within a single ecosystem. ISME J. 2020, 14, 635–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drago, L.; De Vecchi, E.; Gabrieli, A.; De Grandi, R.; Toscano, M. Immunomodulatory effects of Lactobacillus salivarius LS01 and Bifidobacterium breve BR03, alone and in combination, on peripheral blood mononuclear cells of allergic asthmatics. Allergy Asthma Immunol. Res. 2015, 7, 409–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, Y.; Iwabuchi, N.; Xiao, J.-z.; Yaeshima, T.; Iwatsuki, K. Suppressive effects of Bifidobacterium breve strain M-16V on T-helper type 2 immune responses in a murine model. Biol. Pharm. Bull. 2009, 32, 760–763. [Google Scholar] [CrossRef] [Green Version]
- Toh, Z.Q.; Anzela, A.; Tang, M.L.; Licciardi, P.V. Probiotic therapy as a novel approach for allergic disease. Front. Pharmacol. 2012, 3, 171. [Google Scholar] [CrossRef] [Green Version]
- Rusu, E.; Enache, G.; Cursaru, R.; Alexescu, A.; Radu, R.; Onila, O.; Cavallioti, T.; Rusu, F.; Posea, M.; Jinga, M. Prebiotics and probiotics in atopic dermatitis. Exp. Ther. Med. 2019, 18, 926–931. [Google Scholar] [CrossRef] [PubMed]
- Majamaa, H.; Isolauri, E. Evaluation of the gut mucosal barrier: Evidence for increased antigen transfer in children with atopic eczema. J. Allergy Clin. Immunol. 1996, 97, 985–990. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Lee, E.; Park, Y.M.; Hong, S.-J. Microbiome in the gut-skin axis in atopic dermatitis. Allergy Asthma Immunol. Res. 2018, 10, 354–362. [Google Scholar] [CrossRef]
- Reddel, S.; Del Chierico, F.; Quagliariello, A.; Giancristoforo, S.; Vernocchi, P.; Russo, A.; Fiocchi, A.; Rossi, P.; Putignani, L.; El Hachem, M. Gut microbiota profile in children affected by atopic dermatitis and evaluation of intestinal persistence of a probiotic mixture. Sci. Rep. 2019, 9, 1–10. [Google Scholar]
- Pan, H.-H.; Lue, K.-H.; Sun, H.-L.; Ku, M.-S. Gastroenteritis during infancy is a novel risk factor for allergic disease. Medicine 2019, 98. [Google Scholar] [CrossRef]
- Patrick, D.M.; Sbihi, H.; Dai, D.L.Y.; Al Mamun, A.; Rasali, D.; Rose, C.; Marra, F.; Boutin, R.C.T.; Petersen, C.; Stiemsma, L.T.; et al. Decreasing antibiotic use, the gut microbiota, and asthma incidence in children: Evidence from population-based and prospective cohort studies. Lancet. Respir. Med. 2020. [Google Scholar] [CrossRef]
- Vuillermin, P.J.; O’Hely, M.; Collier, F.; Allen, K.J.; Tang, M.L.; Harrison, L.C.; Carlin, J.B.; Saffery, R.; Ranganathan, S.; Sly, P.D. Maternal carriage of Prevotella during pregnancy associates with protection against food allergy in the offspring. Nat. Commun. 2020, 11, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biagi, E.; Quercia, S.; Aceti, A.; Beghetti, I.; Rampelli, S.; Turroni, S.; Faldella, G.; Candela, M.; Brigidi, P.; Corvaglia, L. The bacterial ecosystem of mother’s milk and infant’s mouth and gut. Front. Microbiol. 2017, 8, 1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, K.M.; Foster, J.A.; Forney, L.J.; Schütte, U.M.E.; Beck, D.L.; Abdo, Z.; Fox, L.K.; Williams, J.E.; McGuire, M.K.; McGuire, M.A. Characterization of the Diversity and Temporal Stability of Bacterial Communities in Human Milk. PLoS ONE 2011, 6, e21313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez, E.; de Andres, J.; Manrique, M.; Pareja-Tobes, P.; Tobes, R.; Martinez-Blanch, J.F.; Codoner, F.M.; Ramon, D.; Fernandez, L.; Rodriguez, J.M. Metagenomic Analysis of Milk of Healthy and Mastitis-Suffering Women. J. Hum Lact 2015, 31, 406–415. [Google Scholar] [CrossRef]
- Jost, T.; Lacroix, C.; Braegger, C.; Chassard, C. Assessment of bacterial diversity in breast milk using culture-dependent and culture-independent approaches. Br. J. Nutr. 2013, 110, 1253–1262. [Google Scholar] [CrossRef] [Green Version]
- Tuzun, F.; Kumral, A.; Duman, N.; Ozkan, H. Breast milk jaundice: effect of bacteria present in breast milk and infant feces. J Pediatr Gastroenterol Nutr 2013, 56, 328–332. [Google Scholar] [CrossRef]
- Cabrera-Rubio, R.; Collado, M.C.; Laitinen, K.; Salminen, S.; Isolauri, E.; Mira, A. The human milk microbiome changes over lactation and is shaped by maternal weight and mode of delivery. Am. J. Clin. Nutr. 2012, 96, 544–551. [Google Scholar] [CrossRef] [Green Version]
- Collado, M.C.; Delgado, S.; Maldonado, A.; Rodriguez, J.M. Assessment of the bacterial diversity of breast milk of healthy women by quantitative real-time PCR. Lett. Appl. Microbiol. 2009, 48, 523–528. [Google Scholar] [CrossRef]
- Martin, R.; Heilig, H.G.; Zoetendal, E.G.; Jimenez, E.; Fernandez, L.; Smidt, H.; Rodriguez, J.M. Cultivation-independent assessment of the bacterial diversity of breast milk among healthy women. Res. Microbiol. 2007, 158, 31–37. [Google Scholar] [CrossRef]
- Khodayar-Pardo, P.; Mira-Pascual, L.; Collado, M.C.; Martinez-Costa, C. Impact of lactation stage, gestational age and mode of delivery on breast milk microbiota. J. Perinatol. 2014, 34, 599–605. [Google Scholar] [CrossRef]
- Newburg, D.S.; Walker, W.A. Protection of the Neonate by the Innate Immune System of Developing Gut and of Human Milk. Pediatric Res. 2007, 61, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Perez, P.; Dore, J.; Leclerc, M.; Florence, L.; Benyacoub, J.; Serrant, P.; Segura-Roggero, I.; Schiffrin, E.; Donnet-Hughes, A. Bacterial Imprinting of the Neonatal Immune System: Lessons From Maternal Cells? Pediatrics 2007, 119, e724–e732. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, E.; Fernández, L.; Maldonado, A.; Martín, R.; Olivares, M.; Xaus, J.; Rodríguez, J.M. Oral administration of Lactobacillus strains isolated from breast milk as an alternative for the treatment of infectious mastitis during lactation. Appl. Env. Microbiol. 2008, 74, 4650–4655. [Google Scholar] [CrossRef] [Green Version]
- Arroyo, R.; Martin, V.; Maldonado, A.; Jimenez, E.; Fernandez, L.; Rodriguez, J.M. Treatment of infectious mastitis during lactation: Antibiotics versus oral administration of Lactobacilli isolated from breast milk. Clin. Infect Dis. 2010, 50, 1551–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, L.; Cardenas, N.; Arroyo, R.; Manzano, S.; Jimenez, E.; Martin, V.; Rodriguez, J.M. Prevention of Infectious Mastitis by Oral Administration of Lactobacillus salivarius PS2 During Late Pregnancy. Clin. Infect Dis. 2016, 62, 568–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrahamsson, T.R.; Sinkiewicz, G.; Jakobsson, T.; Fredrikson, M.; Bjorksten, B. Probiotic lactobacilli in breast milk and infant stool in relation to oral intake during the first year of life. J. Pediatr. Gastroenterol. Nutr. 2009, 49, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Simpson, M.R.; Avershina, E.; Storrø, O.; Johnsen, R.; Rudi, K.; Øien, T. Breastfeeding-associated microbiota in human milk following supplementation with Lactobacillus rhamnosus GG, Lactobacillus acidophilus La-5, and Bifidobacterium animalis ssp. lactis Bb-12. J. Dairy Sci. 2018, 101, 889–899. [Google Scholar] [CrossRef]
- Mastromarino, P.; Capobianco, D.; Miccheli, A.; Pratico, G.; Campagna, G.; Laforgia, N.; Capursi, T.; Baldassarre, M.E. Administration of a multistrain probiotic product (VSL#3) to women in the perinatal period differentially affects breast milk beneficial microbiota in relation to mode of delivery. Pharm. Res. 2015, 95–96, 63–70. [Google Scholar] [CrossRef]
- Malamitsi-Puchner, A.; Protonotariou, E.; Boutsikou, T.; Makrakis, E.; Sarandakou, A.; Creatsas, G. The influence of the mode of delivery on circulating cytokine concentrations in the perinatal period. Early Hum. Dev. 2005, 81, 387–392. [Google Scholar] [CrossRef]
- Rodríguez, J.M. The origin of human milk bacteria: is there a bacterial entero-mammary pathway during late pregnancy and lactation? Adv. Nutr. 2014, 5, 779–784. [Google Scholar] [CrossRef] [Green Version]
- Eigenmann, P.A.; Sicherer, S.H.; Borkowski, T.A.; Cohen, B.A.; Sampson, H.A. Prevalence of IgE-mediated food allergy among children with atopic dermatitis. Pediatrics 1998, 101, e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koletzko, S.; Niggemann, B.; Arato, A.; Dias, J.A.; Heuschkel, R.; Husby, S.; Mearin, M.L.; Papadopoulou, A.; Ruemmele, F.M.; Staiano, A.; et al. Diagnostic approach and management of cow’s-milk protein allergy in infants and children: ESPGHAN GI Committee practical guidelines. J. Pediatr. Gastroenterol. Nutr. 2012, 55, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.L.; Dunstan, J.A.; Prescott, S.L. Probiotic supplementation for the first 6 months of life fails to reduce the risk of atopic dermatitis and increases the risk of allergen sensitization in high-risk children: A randomized controlled trial. J. Allergy Clin. Immunol. 2007, 119, 184–191. [Google Scholar] [CrossRef] [PubMed]
- West, C.E.; Hammarström, M.L.; Hernell, O. Probiotics in primary prevention of allergic disease–follow-up at 8–9 years of age. Allergy 2013, 68, 1015–1020. [Google Scholar] [CrossRef]
- Viljanen, M.; Kuitunen, M.; Haahtela, T.; Juntunen-Backman, K.; Korpela, R.; Savilahti, E. Probiotic effects on faecal inflammatory markers and on faecal IgA in food allergic atopic eczema/dermatitis syndrome infants. Pediatr. Allergy Immunol. 2005, 16, 65–71. [Google Scholar] [CrossRef]
- Hol, J.; van Leer, E.H.; Schuurman, B.E.E.; de Ruiter, L.F.; Samsom, J.N.; Hop, W.; Neijens, H.J.; de Jongste, J.C.; Nieuwenhuis, E.E. The acquisition of tolerance toward cow’s milk through probiotic supplementation: A randomized, controlled trial. J. Allergy Clin. Immunol. 2008, 121, 1448–1454. [Google Scholar] [CrossRef]
- Morisset, M.; Aubert-Jacquin, C.; Soulaines, P.; Moneret-Vautrin, D.A.; Dupont, C. A non-hydrolyzed, fermented milk formula reduces digestive and respiratory events in infants at high risk of allergy. Eur. J. Clin. Nutr. 2011, 65, 175–183. [Google Scholar] [CrossRef]
- Sato, A.; Hashiguchi, M.; Toda, E.; Iwasaki, A.; Hachimura, S.; Kaminogawa, S. CD11b+ Peyer’s patch dendritic cells secrete IL-6 and induce IgA secretion from naive B cells. J. Immunol. 2003, 171, 3684–3690. [Google Scholar] [CrossRef] [Green Version]
- Allen, S.J.; Jordan, S.; Storey, M.; Thornton, C.A.; Gravenor, M.B.; Garaiova, I.; Plummer, S.F.; Wang, D.; Morgan, G. Probiotics in the prevention of eczema: A randomised controlled trial. Arch. Dis. Child 2014, 99, 1014–1019. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Kwon, J.H.; Ahn, S.H.; Lee, S.I.; Han, Y.S.; Choi, Y.O.; Lee, S.Y.; Ahn, K.M.; Ji, G.E. Effect of probiotic mix (Bifidobacterium bifidum, Bifidobacterium lactis, Lactobacillus acidophilus) in the primary prevention of eczema: A double-blind, randomized, placebo-controlled trial. Pediatr. Allergy Immunol. 2010, 21, e386–393. [Google Scholar] [CrossRef]
- Kuitunen, M.; Kukkonen, K.; Juntunen-Backman, K.; Korpela, R.; Poussa, T.; Tuure, T.; Haahtela, T.; Savilahti, E. Probiotics prevent IgE-associated allergy until age 5 years in cesarean-delivered children but not in the total cohort. J. Allergy Clin. Immunol. 2009, 123, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Abrahamsson, T.R.; Jakobsson, T.; Böttcher, M.F.; Fredrikson, M.; Jenmalm, M.C.; Björkstén, B.; Oldaeus, G. Probiotics in prevention of IgE-associated eczema: Double-blind, randomized, placebo-controlled trial. J. Allergy Clin. Immunol. 2007, 119, 1174–1180. [Google Scholar] [CrossRef] [PubMed]
- Boyle, R.J.; Ismail, I.H.; Kivivuori, S.; Licciardi, P.V.; Robins-Browne, R.M.; Mah, L.J.; Axelrad, C.; Moore, S.; Donath, S.; Carlin, J.B.; et al. Lactobacillus GG treatment during pregnancy for the prevention of eczema: A randomized controlled trial. Allergy 2011, 66, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Akinbami, O.J. Trends in Asthma Prevalence, Health Care Use, and Mortality in the United States, 2001-2010; US Department of Health and Human Services, Centers for Disease Control and Prevention, National Center for Health Statistics: Hyattsville, MD, USA, 2012.
- Wallace, D.V.; Dykewicz, M.S.; Bernstein, D.I.; Blessing-Moore, J.; Cox, L.; Khan, D.A.; Lang, D.M.; Nicklas, R.A.; Oppenheimer, J.; Portnoy, J.M. The diagnosis and management of rhinitis: An updated practice parameter. J. Allergy Clin. Immunol. 2008, 122, S1–S84. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, E.J. Pediatric allergy: A brief review of risk factors associated with developing allergic disease in childhood. Ann. Allergyasthma Immunol. 2003, 90, 53–58. [Google Scholar] [CrossRef]
- Zuccotti, G.; Meneghin, F.; Aceti, A.; Barone, G.; Callegari, M.L.; Di Mauro, A.; Fantini, M.; Gori, D.; Indrio, F.; Maggio, L. Probiotics for prevention of atopic diseases in infants: Systematic review and meta-analysis. Allergy 2015, 70, 1356–1371. [Google Scholar] [CrossRef] [Green Version]
- Elazab, N.; Mendy, A.; Gasana, J.; Vieira, E.R.; Quizon, A.; Forno, E. Probiotic administration in early life, atopy, and asthma: A meta-analysis of clinical trials. Pediatrics 2013, 132, e666–e676. [Google Scholar] [CrossRef] [Green Version]
- Cabana, M.D.; McKean, M.; Caughey, A.B.; Fong, L.; Lynch, S.; Wong, A.; Leong, R.; Boushey, H.A.; Hilton, J.F. Early probiotic supplementation for eczema and asthma prevention: A randomized controlled trial. Pediatrics 2017, 140, e20163000. [Google Scholar] [CrossRef] [Green Version]
- Loo, E.X.; Llanora, G.V.; Lu, Q.; Aw, M.M.; Lee, B.W.; Shek, L.P. Supplementation with probiotics in the first 6 months of life did not protect against eczema and allergy in at-risk Asian infants: A 5-year follow-up. Int. Arch. Allergy Immunol. 2014, 163, 25–28. [Google Scholar] [CrossRef]
- Kapoor, R.; Menon, C.; Hoffstad, O.; Bilker, W.; Leclerc, P.; Margolis, D.J. The prevalence of atopic triad in children with physician-confirmed atopic dermatitis. J. Am. Acad. Dermatol. 2008, 58, 68–73. [Google Scholar] [CrossRef]
- Schneider, L.; Hanifin, J.; Boguniewicz, M.; Eichenfield, L.F.; Spergel, J.M.; Dakovic, R.; Paller, A.S. Study of the atopic march: Development of atopic comorbidities. Pediatric Dermatol. 2016, 33, 388–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickens, K.; Barthow, C.; Mitchell, E.A.; Kang, J.; van Zyl, N.; Purdie, G.; Stanley, T.; Fitzharris, P.; Murphy, R.; Crane, J. Effects of Lactobacillus rhamnosus HN001 in early life on the cumulative prevalence of allergic disease to 11 years. Pediatric Allergy Immunol. 2018, 29, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Wickens, K.; Black, P.; Stanley, T.; Mitchell, E.; Barthow, C.; Fitzharris, P.; Purdie, G.; Crane, J. A protective effect of L actobacillus rhamnosus HN 001 against eczema in the first 2 years of life persists to age 4 years. Clin. Exp. Allergy 2012, 42, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Davies, G.; Jordan, S.; Brooks, C.J.; Thayer, D.; Storey, M.; Morgan, G.; Allen, S.; Garaiova, I.; Plummer, S.; Gravenor, M. Long term extension of a randomised controlled trial of probiotics using electronic health records. Sci. Rep. 2018, 8, 1–8. [Google Scholar]
- Dotterud, C.; Storrø, O.; Johnsen, R.; Øien, T. Probiotics in pregnant women to prevent allergic disease: A randomized, double-blind trial. Br. J. Dermatol. 2010, 163, 616–623. [Google Scholar] [CrossRef]
- Simpson, M.R.; Dotterud, C.K.; Storrø, O.; Johnsen, R.; Øien, T. Perinatal probiotic supplementation in the prevention of allergy related disease: 6 year follow up of a randomised controlled trial. BMC Dermatol. 2015, 15, 13. [Google Scholar] [CrossRef] [Green Version]
- Abrahamsson, T.R.; Jakobsson, T.; Björkstén, B.; Oldaeus, G.; Jenmalm, M.C. No effect of probiotics on respiratory allergies: A seven-year follow-up of a randomized controlled trial in infancy. Pediatric Allergy Immunol. 2013, 24, 556–561. [Google Scholar] [CrossRef] [Green Version]
- Abrahamsson, T.; Sandberg Abelius, M.; Forsberg, A.; Björkstén, B.; Jenmalm, M. A Th1/Th2-associated chemokine imbalance during infancy in children developing eczema, wheeze and sensitization. Clin. Exp. Allergy 2011, 41, 1729–1739. [Google Scholar] [CrossRef] [Green Version]
- Kukkonen, A.K.; Kuitunen, M.; Savilahti, E.; Pelkonen, A.; Malmberg, P.; Mäkelä, M. Airway inflammation in probiotic-treated children at 5 years. Pediatric Allergy Immunol. 2011, 22, 249–251. [Google Scholar] [CrossRef]
- Schmidt, R.M.; Pilmann Laursen, R.; Bruun, S.; Larnkjær, A.; Mølgaard, C.; Michaelsen, K.F.; Høst, A. Probiotics in late infancy reduce the incidence of eczema: A randomized controlled trial. Pediatric Allergy Immunol. 2019, 30, 335–340. [Google Scholar] [CrossRef]
- Puhan, M.A.; Schünemann, H.J.; Murad, M.H.; Li, T.; Brignardello-Petersen, R.; Singh, J.A.; Kessels, A.G.; Guyatt, G.H. A GRADE Working Group approach for rating the quality of treatment effect estimates from network meta-analysis. Bmj 2014, 349, g5630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahlgren, A.F.; Pan, A.; Lam, V.; Gouthro, K.C.; Simpson, P.M.; Salzman, N.H.; Nghiem-Rao, T.H. Longitudinal changes in the gut microbiome of infants on total parenteral nutrition. Pediatric Res. 2019, 86, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Berrington, J.E.; Stewart, C.J.; Cummings, S.P.; Embleton, N.D. The neonatal bowel microbiome in health and infection. Curr. Opin. Infect. Dis. 2014, 27, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Morowitz, M.J.; Poroyko, V.; Caplan, M.; Alverdy, J.; Liu, D.C. Redefining the role of intestinal microbes in the pathogenesis of necrotizing enterocolitis. Pediatrics 2010, 125, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Mihatsch, W.A.; Braegger, C.P.; Decsi, T.; Kolacek, S.; Lanzinger, H.; Mayer, B.; Moreno, L.A.; Pohlandt, F.; Puntis, J.; Shamir, R. Critical systematic review of the level of evidence for routine use of probiotics for reduction of mortality and prevention of necrotizing enterocolitis and sepsis in preterm infants. Clin. Nutr. 2012, 31, 6–15. [Google Scholar] [CrossRef]
- Bonsante, F.; Iacobelli, S.; Gouyon, J.-B. Routine probiotic use in very preterm infants: Retrospective comparison of two cohorts. Am. J. Perinatol. 2013, 30, 041–046. [Google Scholar] [CrossRef] [Green Version]
- Hunter, C.; Dimaguila, M.A.V.; Gal, P.; Wimmer, J.E.; Ransom, J.L.; Carlos, R.Q.; Smith, M.; Davanzo, C.C. Effect of routine probiotic, Lactobacillus reuteri DSM 17938, use on rates of necrotizing enterocolitis in neonates with birthweight<1000 grams: Sequential analysis. Bmc Pediatrics 2012, 12, 142. [Google Scholar]
- Janvier, A.; Malo, J.; Barrington, K.J. Cohort study of probiotics in a North American neonatal intensive care unit. J. Pediatrics 2014, 164, 980–985. [Google Scholar] [CrossRef]
- Robertson, C.; Savva, G.M.; Clapuci, R.; Jones, J.; Maimouni, H.; Brown, E.; Minocha, A.; Hall, L.J.; Clarke, P. Incidence of necrotising enterocolitis before and after introducing routine prophylactic Lactobacillus and Bifidobacterium probiotics. Arch. Dis. Child. Fetal Neonatal Ed. 2020, 105, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Härtel, C.; Pagel, J.; Rupp, J.; Bendiks, M.; Guthmann, F.; Rieger-Fackeldey, E.; Heckmann, M.; Franz, A.; Schiffmann, J.-H.; Zimmermann, B. Prophylactic use of Lactobacillus acidophilus/Bifidobacterium infantis probiotics and outcome in very low birth weight infants. J. Pediatrics 2014, 165, 285–289. e281. [Google Scholar]
- Li, D.; Rosito, G.; Slagle, T. Probiotics for the prevention of necrotizing enterocolitis in neonates: An 8-year retrospective cohort study. J. Clin. Pharm. Ther. 2013, 38, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Luoto, R.; Matomäki, J.; Isolauri, E.; Lehtonen, L. Incidence of necrotizing enterocolitis in very-low-birth-weight infants related to the use of Lactobacillus GG. Acta Paediatr. 2010, 99, 1135–1138. [Google Scholar] [CrossRef] [PubMed]
- Dilli, D.; Aydin, B.; Fettah, N.D.; Özyazıcı, E.; Beken, S.; Zenciroğlu, A.; Okumuş, N.; Özyurt, B.M.; İpek, M.Ş.; Akdağ, A. The propre-save study: Effects of probiotics and prebiotics alone or combined on necrotizing enterocolitis in very low birth weight infants. J. Pediatrics 2015, 166, 545–551. e541. [Google Scholar] [CrossRef]
- Fernández-Carrocera, L.A.; Solis-Herrera, A.; Cabanillas-Ayón, M.; Gallardo-Sarmiento, R.B.; García-Pérez, C.S.; Montaño-Rodríguez, R.; Echániz-Aviles, M.O.L. Double-blind, randomised clinical assay to evaluate the efficacy of probiotics in preterm newborns weighing less than 1500 g in the prevention of necrotising enterocolitis. Arch. Dis. Child. Fetal Neonatal Ed. 2013, 98, F5–F9. [Google Scholar]
- Oncel, M.Y.; Sari, F.N.; Arayici, S.; Guzoglu, N.; Erdeve, O.; Uras, N.; Oguz, S.S.; Dilmen, U. Lactobacillus reuteri for the prevention of necrotising enterocolitis in very low birthweight infants: A randomised controlled trial. Arch. Dis. Child. Fetal Neonatal Ed. 2014, 99, F110–F115. [Google Scholar] [CrossRef]
- Lambæk, I.D.; Fonnest, G.; Gormsen, M.; Brok, J.; Greisen, G. Probiotics to prevent necrotising enterocolitis in very preterm infants. Dan. Med. J. 2016, 63, A5203. [Google Scholar]
- Braga, T.D.; da Silva, G.A.P.; de Lira, P.I.C.; de Carvalho Lima, M. Efficacy of Bifidobacterium breve and Lactobacillus casei oral supplementation on necrotizing enterocolitis in very-low-birth-weight preterm infants: a double-blind, randomized, controlled trial–. Am. J. Clin. Nutr. 2011, 93, 81–86. [Google Scholar] [CrossRef]
- Dang, S.; Shook, L.; Garlitz, K.; Hanna, M.; Desai, N. Nutritional outcomes with implementation of probiotics in preterm infants. J. Perinatol. 2015, 35, 447–450. [Google Scholar] [CrossRef]
- Demirel, G.; Erdeve, O.; Celik, I.H.; Dilmen, U. Saccharomyces boulardii for prevention of necrotizing enterocolitis in preterm infants: A randomized, controlled study. Acta Paediatr. 2013, 102, e560–e565. [Google Scholar] [CrossRef]
- Ree, I.M.; Smits-Wintjens, V.E.; Rijntjes-Jacobs, E.G.; Pelsma, I.C.; Steggerda, S.J.; Walther, F.J.; Lopriore, E. Necrotizing enterocolitis in small-for-gestational-age neonates: A matched case-control study. Neonatology 2014, 105, 74–78. [Google Scholar] [CrossRef]
- Houghteling, P.D.; Walker, W.A. Why is initial bacterial colonization of the intestine important to the infant’s and child’s health? J. Pediatric Gastroenterol. Nutr. 2015, 60, 294. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, I.; Schofield, Z.; Hall, L.J. Exploring the role of the microbiota member Bifidobacterium in modulating immune-linked diseases. Emerg. Top. Life Sci. 2017, 1, 333–349. [Google Scholar]
- Costeloe, K.; Hardy, P.; Juszczak, E.; Wilks, M.; Millar, M.R. Bifidobacterium breve BBG-001 in very preterm infants: A randomised controlled phase 3 trial. Lancet 2016, 387, 649–660. [Google Scholar] [CrossRef]
- Liu, D.; Shao, L.; Zhang, Y.; Kang, W. Safety and efficacy of Lactobacillus for preventing necrotizing enterocolitis in preterm infants: A systematic review and meta-analysis. Int. J. Surg. 2020. [Google Scholar] [CrossRef]
- Deshpande, G.; Jape, G.; Rao, S.; Patole, S. Benefits of probiotics in preterm neonates in low-income and medium-income countries: A systematic review of randomised controlled trials. BMJ Open 2017, 7, e017638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, L.-w.; Yan, B.-l.; Yang, Q.-y.; Li, M.-m.; Cui, H.-l. Which is the best probiotic treatment strategy to prevent the necrotizing enterocolitis in premature infants: A network meta-analysis revealing the efficacy and safety. Medicine 2019, 98, e17521. [Google Scholar] [CrossRef]
- Van den Akker, C.H.P.; van Goudoever, J.B.; Shamir, R.; Domellöf, M.; Embleton, N.D.; Hojsak, I.; Lapillonne, A.; Mihatsch, W.A.; Berni Canani, R.; Bronsky, J.; et al. Probiotics and Preterm Infants: A Position Paper by the European Society for Paediatric Gastroenterology Hepatology and Nutrition Committee on Nutrition and the European Society for Paediatric Gastroenterology Hepatology and Nutrition Working Group for Probiotics and Prebiotics. J. Pediatric Gastroenterol. Nutr. 2020, 70, 664–680. [Google Scholar] [CrossRef]
- Ohishi, A.; Takahashi, S.; Ito, Y.; Ohishi, Y.; Tsukamoto, K.; Nanba, Y.; Ito, N.; Kakiuchi, S.; Saitoh, A.; Morotomi, M. Bifidobacterium septicemia associated with postoperative probiotic therapy in a neonate with omphalocele. J. Pediatrics 2010, 156, 679–681. [Google Scholar] [CrossRef]
- Zheng, M.; Zhang, R.; Tian, X.; Zhou, X.; Pan, X.; Wong, A. Assessing the risk of probiotic dietary supplements in the context of antibiotic resistance. Front. Microbiol. 2017, 8, 908. [Google Scholar] [CrossRef]
- Deshpande, G.; Athalye-Jape, G.; Patole, S. Para-probiotics for preterm neonates—The next frontier. Nutrients 2018, 10, 871. [Google Scholar] [CrossRef] [Green Version]
- Serce, O.; Benzer, D.; Gursoy, T.; Karatekin, G.; Ovali, F. Efficacy of Saccharomyces boulardii on necrotizing enterocolitis or sepsis in very low birth weight infants: A randomised controlled trial. Early Hum. Dev. 2013, 89, 1033–1036. [Google Scholar] [CrossRef] [PubMed]
- De Simone, C. The unregulated probiotic market. Clin. Gastroenterol. Hepatol. 2019, 17, 809–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Directorate-General, S.C.o.F.E.C.H.a.C.P. Report of the Scientific Committee on Food on the Revision of Essential Requirements of Infant Formulae and Follow-on Formulae; European Commission: Brussels, Belgium, 18 May 2003; p. 213. [Google Scholar]
- Topcuoglu, S.; Gursoy, T.; Ovalı, F.; Serce, O.; Karatekin, G. A new risk factor for neonatal vancomycin-resistant Enterococcus colonisation: bacterial probiotics. J. Matern. Fetal Neonatal Med. 2015, 28, 1491–1494. [Google Scholar] [CrossRef]
- Jenke, A.; Ruf, E.-M.; Hoppe, T.; Heldmann, M.; Wirth, S. Bifidobacterium septicaemia in an extremely low-birthweight infant under probiotic therapy. Arch. Dis. Child. Fetal Neonatal Ed. 2012, 97, F217–F218. [Google Scholar] [CrossRef]
- Mantaring, J.; Benyacoub, J.; Destura, R.; Pecquet, S.; Vidal, K.; Volger, S.; Guinto, V. Effect of maternal supplement beverage with and without probiotics during pregnancy and lactation on maternal and infant health: a randomized controlled trial in the Philippines. BMC Pregnancy Childbirth 2018, 18, 193. [Google Scholar] [CrossRef]
- Luoto, R.; Laitinen, K.; Nermes, M.; Isolauri, E. Impact of maternal probiotic-supplemented dietary counselling on pregnancy outcome and prenatal and postnatal growth: A double-blind, placebo-controlled study. Br. J. Nutr. 2010, 103, 1792–1799. [Google Scholar] [CrossRef] [Green Version]
- Pellonperä, O.; Mokkala, K.; Houttu, N.; Vahlberg, T.; Koivuniemi, E.; Tertti, K.; Rönnemaa, T.; Laitinen, K. Efficacy of fish oil and/or probiotic intervention on the incidence of gestational diabetes mellitus in an at-risk group of overweight and obese women: A randomized, placebo-controlled, double-blind clinical trial. Diabetes Care 2019, 42, 1009–1017. [Google Scholar] [CrossRef] [PubMed]
- Baldassarre, M.E.; Di Mauro, A.; Mastromarino, P.; Fanelli, M.; Martinelli, D.; Urbano, F.; Capobianco, D.; Laforgia, N. Administration of a multi-strain probiotic product to women in the perinatal period differentially affects the breast milk cytokine profile and may have beneficial effects on neonatal gastrointestinal functional symptoms. A randomized clinical trial. Nutrients 2016, 8, 677. [Google Scholar] [CrossRef] [Green Version]
- Baldassarre, M.E.; Di Mauro, A.; Tafuri, S.; Rizzo, V.; Gallone, M.S.; Mastromarino, P.; Capobianco, D.; Laghi, L.; Zhu, C.; Capozza, M. Effectiveness and safety of a probiotic-mixture for the treatment of infantile colic: A double-blind, randomized, placebo-controlled clinical trial with fecal real-time PCR and NMR-based metabolomics analysis. Nutrients 2018, 10, 195. [Google Scholar] [CrossRef] [Green Version]
- Cekola, P.L.; Czerkies, L.A.; Storm, H.M.; Wang, M.H.; Roberts, J.; Saavedra, J.M. Growth and tolerance of term infants fed formula with probiotic Lactobacillus reuteri. Clin. Pediatrics 2015, 54, 1175–1184. [Google Scholar] [CrossRef]
- Escribano, J.; Ferré, N.; Gispert-Llaurado, M.; Luque, V.; Rubio-Torrents, C.; Zaragoza-Jordana, M.; Polanco, I.; Codoñer, F.M.; Chenoll, E.; Morera, M. Bifidobacterium longum subsp infantis CECT7210-supplemented formula reduces diarrhea in healthy infants: A randomized controlled trial. Pediatric Res. 2018, 83, 1120–1128. [Google Scholar] [CrossRef] [PubMed]
- Lundelin, K.; Poussa, T.; Salminen, S.; Isolauri, E. Long-term safety and efficacy of perinatal probiotic intervention: Evidence from a follow-up study of four randomized, double-blind, placebo-controlled trials. Pediatric Allergy Immunol. 2017, 28, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Pärtty, A.; Luoto, R.; Kalliomäki, M.; Salminen, S.; Isolauri, E. Effects of early prebiotic and probiotic supplementation on development of gut microbiota and fussing and crying in preterm infants: A randomized, double-blind, placebo-controlled trial. J. Pediatrics 2013, 163, 1272–1277. e1272. [Google Scholar]
- Radke, M.; Picaud, J.-C.; Loui, A.; Cambonie, G.; Faas, D.; Lafeber, H.N.; de Groot, N.; Pecquet, S.S.; Steenhout, P.G.; Hascoet, J.-M. Starter formula enriched in prebiotics and probiotics ensures normal growth of infants and promotes gut health: A randomized clinical trial. Pediatric Res. 2017, 81, 622–631. [Google Scholar] [CrossRef] [Green Version]
- Savino, F.; Cordisco, L.; Tarasco, V.; Palumeri, E.; Calabrese, R.; Oggero, R.; Roos, S.; Matteuzzi, D. Lactobacillus reuteri DSM 17938 in infantile colic: A randomized, double-blind, placebo-controlled trial. Pediatrics 2010, 126, e526–e533. [Google Scholar] [CrossRef]
- Scalabrin, D.; Harris, C.; Johnston, W.; Berseth, C. Long-term safety assessment in children who received hydrolyzed protein formulas with Lactobacillus rhamnosus GG: A 5-year follow-up. Eur. J. Pediatrics 2017, 176, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Benor, S.; Marom, R.; Tov, A.B.; Domany, K.A.; Zaidenberg-Israeli, G.; Dollberg, S. Probiotic supplementation in mothers of very low birth weight infants. Am. J. Perinatol. 2014, 31, 497–504. [Google Scholar]
- Fatheree, N.Y.; Liu, Y.; Taylor, C.M.; Hoang, T.K.; Cai, C.; Rahbar, M.H.; Hessabi, M.; Ferris, M.; McMurtry, V.; Wong, C. Lactobacillus reuteri for infants with colic: A double-blind, placebo-controlled, randomized clinical trial. J. Pediatrics 2017, 191, 170–178. e172. [Google Scholar] [CrossRef]
- Jacobs, S.E.; Tobin, J.M.; Opie, G.F.; Donath, S.; Tabrizi, S.N.; Pirotta, M.; Morley, C.J.; Garland, S.M. Probiotic effects on late-onset sepsis in very preterm infants: A randomized controlled trial. Pediatrics 2013, 132, 1055–1062. [Google Scholar] [CrossRef] [Green Version]
- Luoto, R.; Ruuskanen, O.; Waris, M.; Kalliomäki, M.; Salminen, S.; Isolauri, E. Prebiotic and probiotic supplementation prevents rhinovirus infections in preterm infants: A randomized, placebo-controlled trial. J. Allergy Clin. Immunol. 2014, 133, 405–413. [Google Scholar] [CrossRef]
- Smilowitz, J.T.; Moya, J.; Breck, M.A.; Cook, C.; Fineberg, A.; Angkustsiri, K.; Underwood, M.A. Safety and tolerability of Bifidobacterium longum subspecies infantis EVC001 supplementation in healthy term breastfed infants: A phase I clinical trial. BMC Pediatrics 2017, 17, 133. [Google Scholar]
- Wang, Y.; Gao, L.; Zhang, Y.-h.; Shi, C.-s.; Ren, C.-m. Efficacy of probiotic therapy in full-term infants with critical illness. Asia Pac. J. Clin. Nutr. 2014, 23, 575. [Google Scholar] [PubMed]
- Mi, G.L.; Zhao, L.; Qiao, D.D.; Kang, W.Q.; Tang, M.Q.; Xu, J.K. Effectiveness of Lactobacillus reuteri in infantile colic and colicky induced maternal depression: A prospective single blind randomized trial. Antonie Van Leeuwenhoek 2015, 107, 1547–1553. [Google Scholar] [CrossRef] [PubMed]
- Rautava, S.; Kainonen, E.; Salminen, S.; Isolauri, E. Maternal probiotic supplementation during pregnancy and breast-feeding reduces the risk of eczema in the infant. J. Allergy Clin. Immunol. 2012, 130, 1355–1360. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Cernada, M.; Baüerl, C.; Vento, M.; Pérez-Martínez, G. Microbial ecology and host-microbiota interactions during early life stages. Gut Microbes 2012, 3, 352–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vangay, P.; Ward, T.; Gerber, J.S.; Knights, D. Antibiotics, pediatric dysbiosis, and disease. Cell Host Microbe 2015, 17, 553–564. [Google Scholar] [CrossRef] [Green Version]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef] [Green Version]
- Taverniti, V.; Guglielmetti, S. The immunomodulatory properties of probiotic microorganisms beyond their viability (ghost probiotics: proposal of paraprobiotic concept). Genes Nutr. 2011, 6, 261. [Google Scholar] [CrossRef] [Green Version]
- Fiocchi, A.; Pawankar, R.; Cuello-Garcia, C.; Ahn, K.; Al-Hammadi, S.; Agarwal, A.; Beyer, K.; Burks, W.; Canonica, G.W.; Ebisawa, M. World allergy organization-McMaster university guidelines for allergic disease prevention (GLAD-P): Probiotics. World Allergy Organ. J. 2015, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Morgan, A.R.; Han, D.Y.; Wickens, K.; Barthow, C.; Mitchell, E.A.; Stanley, T.V.; Dekker, J.; Crane, J.; Ferguson, L.R. Differential modification of genetic susceptibility to childhood eczema by two probiotics. Clin. Exp. Allergy 2014, 44, 1255–1265. [Google Scholar] [CrossRef]
- Huang, L.; Chen, Q.; Zhao, Y.; Wang, W.; Fang, F.; Bao, Y. Is elective cesarean section associated with a higher risk of asthma? A meta-analysis. J. Asthma 2015, 52, 16–25. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Author (Year) | Antibiotic Exposure | Objectives | n | Population | Key Results | Conclusions |
|---|---|---|---|---|---|---|
| Studies in Preterm Infants | ||||||
| Zou et al. (2018) [70] | Prenatal/ Postnatal | Determine the effects of prenatal antibiotic therapy (PAT) versus prenatal antibiotic free (PAF) group and effects of antibiotic exposure intensity (before and after delivery) on gut microbiota in preterm infants. | 24 | PAT group (n = 12) and PAF group (n = 12). Fecal samples on day 7 and day 14. Treatment duration before and after delivery: H-group (>7d) (n = 11) versus L-group (<7d) (n =11). Fecal samples on day 14. | Phylum level: d7 Proteobacteria (PAF 79.75% vs PAT 92.35%) and Firmicutes (PAF 9.73% vs PAT 4.69%); d14 Proteobacteria (PAF 74.78% vs PAT 87.22%) and Firmicutes (PAF 11.19% vs PAT 10.61%). Bacteroidetes (PAF 5.75% vs PAT 0.38%). Genus level: d7 Klebsiella (PAF 52.17% vs 48.96%), d14 (PAF 45.03% vs PAT 45.81%). Bifidobacterium d7 PAT 5% vs PAF 12%. d14 PAT 9% vs PAF 12%. H-group/L-group: Phylum level: Proteobacteria (H-group 79.35% vs L-group 70.66%); Firmicutes (H-group 19.33% vs L-group 14.81%) Genus level: Klebsiella (H-group 55.91% vs L-group 36.15%). Enterococcus (H-group 23% vs L-group 34.22%). Bifidobacterium (H-group 5.47% vs L-group 10.24%). | The PAT group showed higher prevalence of Proteobacteria and significant decrease in Bacteroides colonization. Delayed colonization of Bifidobacterium in the PAT and H-group. Pre-postnatal antibiotic exposure may affect early gut microbiota composition in preterm infants. |
| Greenwood et al. (2014) [10] | Postnatal | Determine the impact of empiric ampicillin and gentamicin use in the first week of life on microbial colonization and diversity in preterm infants. | 74 | Empiric ampicillin and gentamicin. 3 groups: no antibiotics (0d), brief administration (1–4d), intensive administration (5–7d) Fecal samples on w1–w3. | No differences in Simpson diversity index in the first week between groups. Significant decrease in diversity at weeks 2 and 3 in both antibiotic groups (p < 0.001 and p < 0.004). w1: 0d: Staphylococcus 41%, Enterococcus 26%, Enterobacter 19%. 1–4d: Enterobacter 40%; 5–7d: Enterococcus 34%, Clostridium 33%. w2: Enterobacter as the most common genus in patients who received antibiotics in w1. w3: Enterobacter (47%), Enterococcus (35%) in infants who received intensive administration. | Sustained effects on the gut microbiota by intensive antibiotic therapy in preterm infants. A brief course of antibiotics suppresses the microbiota diversity temporarily. |
| Arboleya et al. (2015) [72] | Intrapartum/ Postnatal | Assessment of intestinal microbiota in VLBW preterm infants considering perinatal factors as delivery mode and antibiotic use (IPA and postnatal). | 40 | 27 VLBW infants (24–32 WGA) vs. 13 full-term, vaginally delivered, exclusively breast-fed (FTVDBF) neonates without antibiotic exposure. IPA: n=14 VLBW vs n=3 FTVDBF. Postnatal antibiotics: n=12 for 5-8 days after birth, n=5 antibiotics starting at 10-13 days of life. Fecal samples: 24hours–48hours, day 10, day 30, day 90 | VLBW vs FTVDBF: 24–48h: VLBW group: reduced colonization of Bacteroidaceae, Clostridiaceae, unclassified Actinobacteri and increased colonization of Bifidobacteriaceae and Lactobacillales (p < 0.05). d10: VLBW group: reduced colonization of Bacteroidaceae, Bifidobacteriaceae and increased colonization of Enterobacteriaceae (p < 0.05). d30–d90: increased colonization of Enterobacteriaceae and reduced colonization of Bacteroidaceae (p < 0.05). 30 days of age: infants not exposed to antibiotics showed significantly higher percentages of Bifidobacteriaceae, Streptococcaceae, and lower of Enterobacteriaceae than infants whose mothers received IPA (independently on whether or not the infant received antibiotics). | VLBW group showed reduced Bacteroidaceae colonization and increased Lactobacillaceae colonization during the first hours of life, followed by a dominance of Enterobacteriaceae, on the first days and up to 3 months of age. At 1 month of age, infants whose mothers received IPA had an intestinal microbiota different from that of the infants whose mothers had not received IPA IPA has an equal or higher effect than postnatal antibiotics in the first days of life. Importance of minimizing early medication exposure. |
| Zwittink et al. (2018) [68] | Postnatal | Effect of postnatal antibiotic treatment duration on preterm gut microbiota. | 15 | 15 late preterm infants (WGA 35.7 ± 0.9) treated with amoxicillin/ceftazidime 3 groups: Antibiotic free (AF) (control): n = 5; Short term (ST) (<3days): n = 5; Long term (LT) (>5days): n = 5. Fecal samples: birth, week1, week2, week3, week4, week6. | AF: high abundance of Bifidobacterium w1–w6 (average RA of 73% at w6). ST and LT infants showed significantly lower abundance of Bifidobacterium after treatment (p = 0.027, 0.027) and at w1 (p = 0.027, 0.021), w2 (p = 0.016, 0.009) and w3 (p = 0.028,0.028) vs. AF. Enterococcus dominant in ST and LT infants during w1, not observed in AF group. Bifidobacterium abundance significantly decreased until w6 in LT group (p = 0.009). Bifidobacterium negatively correlated to Enterococcus, Veillonella, Clostridium, Escherichia–Shigella, and Enterobacter. | Short- and long-term treatment with amoxicillin/ceftazidime during the first postnatal week drastically disturbs the normal colonization pattern. ST but not LT allows the recovery of Bifidobacterium levels in the first 6 w. Bifidobacterium dominance allows higher richness and diversity in gut microbiota. |
| Dardas et al. (2014) [66] | Postnatal | Determine if the duration of antibiotics within the first 10 or 30 d after birth affects the intestinal microbiome. | 29 | 29 preterm infants (WGA <32) fed with breast milk. G1: 2 days of antibiotic (n = 15); G1: 7–10 days of antibiotics (n = 12). Fecal samples: 10d and 30d feeds as maternal breast milk and two received exclusively formula. | Significantly lower Shannon–Wiener diversity index in G2 from 10 d samples vs. G1. Firmicutes and Bacteroidetes dominated the 10d samples, in the 30d samples, the predominant phylum remained Firmicutes, but there was a relative rise in Actinobacteria and Proteobacteria: Firmicutes was the predominant phylum. | Rectal microbiota diversity increases over time but decreases with antibiotic exposure. Despite antibiotic pressure, it continues to acquire different bacterial genera. |
| Zhu et al. (2017) [69] | Postnatal | To assess the effects of one-week antibacterial treatment on the gut bacterial community in preterm infants during the first week of life. | 36 | 36 preterm infants (WGA: 28–37), formula-fed. 3 groups: Penicillin-moxalactam group (PM): n = 12; Piperacillin-tazobactam group (PT): n = 12; Antibiotic free group (AF): n = 12; Fecal samples: day3, day7 | No statistical difference in Shannon–Wiener index among groups on both d3 and d7. Significantly lower Shannon–Wiener index in PM (p = 0.008) and PT (p = 0.028) groups on d7 compared to d3. Firmicutes and Proteobacteria the most abundant phyla in all groups on d3 and d7. Bacteroidetes and Clostridia were rarely detected. d3: PT group: Enterococcus, Streptococcus, and Pseudomonas > 60% of the microbiota. Lactobacillus significantly higher in PM group (31.57%) than in the other two groups. d7: Higher prevalence of Bacteroidetes in PM and PT than in the AF group (p < 0.05). Significantly higher prevalence of Enterococcus (p = 0.003) in PT vs. AF group. Significantly higher prevalence of Escherichia-Shigella in the PM vs. AF group (p = 0.018). | Prolonged antibiotic therapy affects the early development of gut microbiota in preterm infants. Antibiotic treatment generates a reduction in bacterial diversity and an enrichment of harmful bacteria such as Streptococcus and Pseudomonas. |
| Studies in full-term neonates | ||||||
| Nogacka et al. (2017) [63] | Intrapartum | Impact of IPA on the neonatal gut microbiota. | 40 | IPA group: penicillin (n = 18); No-IPA group (n = 22). All vaginally delivered full-term babies (>37 WGA). Fecal samples: d2, d10, d30, d90. | Relative proportion of Proteobacteria: d2: IPA group: 67% vs. non-IPA 50% 10d: IPA group: 46% vs. non-IPA 35% 90d: IPA group: 34% vs. non-IPA 32% Significantly lower levels of Bifidobacteriaceae and Actinobacteria (p < 0.05) in IPA group. | IPA impacts the establishing neonatal microbiota. The effect remains for at least the first month of life, a very critical time of the development of the microbiota-induced host homeostasis. |
| Aloisio et al. (2016) [60] | Intrapartum | Evaluate IPA on whole microbiome composition of newborns seven days after birth. | 20 | 10 mothers IPA (ampicillin) versus 10 mothers no IPA. Full-term neonates and vaginal delivery. Fecal samples: d6–d7. | Actinobacteria: IPA group 0.4% vs. control 3.8% Bacteroidetes: 16% IPA group vs. control 47.7% Proteobacteria: IPA group 54.7% vs. control 15.5% (p < 0.05) Higher abundance of Gram-negative phyla within the IPA group compared to the control group. | IPA impacts on neonatal gut microbiota reducing microbial biodiversity, allowing colonization of Enterobacteriaceae, and reducing the amount of Actinobacteria. |
| Mazzola et al. (2016) [62] | Intrapartum | Assessment of the impact of maternal IPA on the gut microbiota in the first month of life (neonates). | 26 | 4 study groups: 1: Breast-fed infants /control group (BF-C), Group B Streptococcus (GBS) - 2: Breast-fed infants with IPA (BF-IPA), GBS +. 3: Mixed-fed infantes /control group (MF-C) GBS-. 4: Mixed-fed infants with IPA (MF-IPA), GBS+. Fecal samples: d7, d30. | BF-IPA and BF-C: d7: significantly reduced diversity in BF-IPA based on alpha diversity analysis: Chao1 (p = 0.0122), Simpson (p = 0.035), and Shannon–Wiener (p = 0.0082). Actinobacteria not detected in BF-IPA, 17% in BF-C. BF-IPA dominated by Enterobacteriaceae (E. coli 52%). Bifidobacteria not detected in BF-IPA. BF-C infants also had higher levels of Bacteroides. d30: BF-IPA recovered Bifidobacteria; Enterobacteriaceae still dominate in BF-IPA infants (44%) vs. BF-C (16%). MF-IPA and MF-C No significant difference in diversity. MF-IPA: increased colonization of Proteobacteria (37%) and Firmicutes (41%), compared with MF-C. MF-IPA: increased colonization of Enterobacteriaceae (35%). | IPA had a significant impact on the early gut microbial composition, which could partially be reversed after 30 days of life. |
| Azad et al. (2015) [65] | Intrapartum | Assessment of the impact of IPA on neonatal gut microbiota. Secondary objective: assess the role of breastfeeding in modifying antibiotic-induced gut dysbiosis. | 198 | Full-term neonates, vaginal or C-section birth, and antibiotics. Exposure groups: -no IPA+vaginal delivery; -IPA+vaginal delivery; -IPA+elective CS; -IPA+emergency CS. Fecal samples: m3, m12. Perinatal antibiotics were directly adminis- tered to 8 (4%) infants for suspected sepsis within the first 48 hours after birth, and 69 (37%) of infants received post- natal antibiotics before the 1-year stool collection. Perinatal antibiotics were directly adminis- tered to 8 (4%) infants for suspected sepsis within the first 48 hours after birth, and 69 (37%) of infants received post- natal antibiotics before the 1-year stool collection. | m3: IPA+vaginal delivery was associated with decreased gut microbiota richness (p = 0.005). Phylum level: Decreased colonization of Bacteroides (24%) compared with 46% among unexposed infants (p < 0.05). IPA with CS delivery associated with elevated proportions of Firmicutes (p < 0.01), and Proteobacteria (p < 0.05). Genus level: Enterococcus and Clostridium were predominant. No persistent microbiota differences at one year among infants exposed to IPA with elective CS or vaginal delivery. | IPA in C-section and vaginal delivery are associated with neonatal gut microbiota dysbiosis. IPA was associated with reduced microbiota richness and a depletion of Bacteroidetes and increased colonization of Enterococcus and Clostridium. Breastfeeding modifies some of these effects. |
| Tanaka et al. (2009) [71] | Prenatal/ Postnatal | Impact of antibiotic treatment in neonates or their mothers on the developmental gut microbiota. | 44 | n = 26: 36–41 WGA Control (antibiotic free—AF) group: n = 18 Treatment group (AT): 5 infant subjects were orally administered cefalexin. 3 infants (CD) delivered by C-section: no postnatal antibiotics but their mothers were intravenously injected with cefotiam hydrochloride for 4 days after the delivery. All infants breastfed or given formula. Fecal samples: daily for the first five days and monthly for the first two months. | AT group: diversity decreased from d1 to d3 and remained low until d5. Diversity in AT significantly lower than AF at month 2 (p = 0.02). Colonization by Bifidobacterium attenuated until one month after birth. High detection rate of Enterococcus observed in the AT group since d1 and significantly higher in the first month of life vs. AF (p = 0.01). Enterobacteriaceae significantly higher in months 1 and 2 (p = 0.02) and Bifidobacterium count significantly lower on d3 (p = 0.03) and d5 (p = 0.11) in the AT vs. AF group. AF group: increased colonization during first two months of Bifidobacterium, Clostridium, Bacteroidaceae, and Veillonella. Bifidobacterium increase from 28% to 67% in the first month. Facultative anaerobes (Staphylococcus and Enterococcus) did not show such an increasing trend. CD group: reduced intestinal microbiotal diversity compared to AF group. Reduced colonization of Bifidobacterium and aberrant growth of Enterococcus | Colonization by Bifidobacterium was greatly attenuated in both the AT and CD groups. Overgrowth of Enterococcus and Enterobacteriaceae occurred in most AT infants. Antibiotic administration significantly influences the initial development of the neonatal gut microbiota, with a high impact on Bifidobacterium colonization. |
| Corvaglia et al (2016) [61] | Intrapartum | Effect of IPA on gut microbiota in healthy, full-term infants. Secondary objective: influence of type of feeding on the gut microbiota. | 84 | 84 healthy, full-term infants, born by vaginal delivery. IPA group n = 35; No-IPA group n = 49 Feeding types: exclusive breastfeeding, exclusive formula feeding, or mixed feeding. Fecal samples: d7, d30 | IPA group: significantly lower levels of Bifidobacterium at d7 vs. no-IPA group: log CFU/g (5.51–6.98) vs. 7.80 (6.61–8.26) (p = 0.000). No significant differences of Lactobacilli and Bacteroides fragilis at d7 and d30 between groups. No differences in Bifidobacteria at d30. Higher counts of Bifidobacteria at d7 in no-IPA groups exclusively breastfed. Higher Lactobacillus counts both at d7 and d30 in infants exclusively fed human milk, regardless of IPA treatment. | IPA modifies gut microbiota by reducing Bifidobacteria, which is further affected in infants receiving formula feeding. Long-term consequences require further investigation. |
| Aloisio et al. (2014) [76] | Intrapartum | To assess the influence of IPA on the main microbial groups present in the newborn gut microbiota. | 52 | 52 full-term infants, vaginal delivery, exclusively breastfed. IPA group n = 26; No-IPA group n = 26. Fecal samples: d6–d7. | No-IPA group: E. coli, Bacteroides fragilis group, and Bifidobacteria were the most abundant (9.03, 8.53, and 7.29 log CFU/g, respectively). Lactobacilli and C. difficile showed lower counts (6.73 and 3.70 log CFU/g, respectively). IPA group: significant reduction in Bifidobacterium (from an average of 7.29 to 5.85 log CFU/g). Strong decrement in the frequency of Bifidobacterium breve, B. bifidum and B. dentium in IPA group. B. pseudocatenulatum, B. pseudolongum, and B. longum less influenced by IPA. Lactobacillus, C. difficile, and Bacteroides fragilis were not significantly affected by IPA. | Significant influence of IPA on the early bifidobacterial pattern of newborns. Further studies are necessary to evaluate the long-term effects of IPA. |
| Fouhy et al. (2012) [67] | Postnatal | Assessment of consequences after four and eight weeks of postnatal antibiotic treatment within the first 48 h after birth. | 18 | Treatment group (n = 9): combination of ampicillin and gentamicin within 48 h of birth; Control group (n = 9): no antibiotics. Fecal samples: w4 and w8 after the end of antibiotic treatment. | week4: Shannon–Wiener index > 3.6 in all samples (high level of biodiversity). Increased Proteobacteria colonization (54%) in the treatment group compared to 37% in the control group (p = 0.0049). Bacteroidetes were detected in less than half of infants treated with antibiotics, notably low levels if present. Actinobacteria levels significantly lower in the antibiotic-treated group (3% vs. 24%; p = 0.00001). Enterobacteriaceae predominant (55% vs. 37%; p = 0.0073) and lower levels of Bifidobacteriaceae (3% vs. 24%; p = 0.0132) in the treatment group. Significantly higher levels of Bifidobacterium (25% vs. 5%; p = 0.0132) and Lactobacillus (4% vs. 1%; p = 0.0088) present in the untreated group. week8: significantly higher proportions of Proteobacteria (44%) vs. control (23%). Actinobacteria increased significantly after four weeks in the treatment group, until there were no significant differences vs. control. Enterobacteriaceae decreased after four weeks (p = 0.0136) but remained dominant in the antibiotic group (45%). | Postnatal antibiotic therapy induces alterations in the gut microbiota, over eight weeks. The combined use of ampicillin and gentamicin in early life may have significant effects on gut microbiota, but the long-term health implications remain unknown. |
| Stearns et al. (2017) [64] | Intrapartum | Effects of IPA on the development of gut microbiome among a low-risk population. | 74 | 74 mother–infant pairs IPA group n = 21; No IPA group n = 53. Fecal samples: d3, d10, w6, w12 postpartum. | Bacterial species richness and Shannon–Wiener diversity index were significantly lower (p < 0.01) in infants born vaginally and exposed to IPA at early time points, but reached levels similar to communities in unexposed infants by w12. IPA group: delayed Actinobacteria colonization without differences between delivery modes (vaginal/C-section). Firmicutes showed delayed colonization in vaginally born infants. Prolonged persistence of Proteobacteria. Longer duration of IPA exposure increased the magnitude of the effect on Bifidobacterium populations. Infants born by C-section lacked Bacteroidetes up to w12 and showed a greater abundance of Firmicutes. | IPA affected all aspects of gut microbial ecology including species richness, diversity, community structure, and the abundance of colonizing bacterial genera. |
| Source | Intervention Period | Test/Control | Population/Country | Strain(s)/Dose/Administration | Food Allergy-Related Variable | Results |
|---|---|---|---|---|---|---|
| Boyle et al. (2011) [133] | Prenatal only (from the 36th week of pregnancy to delivery) | 125/125 | Infants at high risk of allergy; Australia | Lactobacillus rhamnosus GG (LGG) 1.8 × 1010 CFU/day in drops. | Incidence of positive SPTs to food allergens (cow’s milk, eggs, and peanuts) at 12 months. | No significant differences. |
| Kim et al. (2010) [130] | Prenatal and postnatal (from the 32nd week of pregnancy to six months after delivery) | 57/55 | Infants at high risk of atopic disease; Korea | Mothers: mixture of Bifidobacterium bifidum BGN4; Bifidobacterium lactis AD011 and Lactobacillus acidophilus AD031; 1.6 × 109 CFU/day for each strain for five months. Infants: same mixture from four to six months of age; dissolved in breast milk, infant formula, or sterile water. | Specific IgE against common food allergens (egg white, cow’s milk, wheat, peanuts, soybeans, and buckwheat). | Lower sensitization to any one of the common food allergens in the probiotic group (38.7% vs. 51.7%), but not significant. |
| Kuitunen et al. (2009) [131] | Prenatal and postnatal (from the 35th week of pregnancy to six months after delivery) | 445/446 | Infants at high risk of allergy; Finland | Mothers: Lactobacillus rhamnosus GG (ATCC 53103); 1 × 1010 CFU; L. rhamnosus LC705 1 × 1010 CFU/day; Bifidobacterium breve Bb99 4 × 108 CFU/day, and Propionibacterium freudenreichii ssp. Shermanii JS 4 × 109 CFU/day) for four weeks. Infants: same mixture for six months; mixed with syrup and 0.8 g of GOS. | Cumulative incidence of any allergic disease and any IgE-mediated allergic disease until age five. | Lower SPT+ and/or food-specific IgE in children born by cesarean vs placebo. No differences in vaginally delivered children. |
| Allen et al. (2014) [129] | Prenatal and postnatal (from the 36th week of pregnancy to six months after delivery) | 220/234 | Infants with and without high risk of atopy; UK | Mothers: Lactobacillus salivarius CUL61, 6.25 × 109 CFU/day; L. paracasei CUL08; Bifidobacterium animalis ssp. lactis CUL34, and Bifidobacterium bifidum CUL20; 1.25 × 109 CFU/day/each strain for four weeks. Infants: same mixture for six months; mixed with breast milk or formula. | Positive SPTs to food allergens (cow’s milk and egg proteins) at either age six months or two years. | Significant decrease in the proportion of SPT+ to CM and eggs in probiotic group after six months; no differences after two years. |
| Abrahamsson et al. (2007) [132] | Prenatal and postnatal (from the 36th week of pregnancy to 12 months after delivery) | 117/115 | Infants at high risk of allergy; Sweden | Mothers: Lactobacillus reuteri ATCC 55730, 1 × 108 CFU/day in drops; for four weeks. Infants: same mixture for 12 months; mixed with breast milk or hypoallergenic formula. | Incidence of positive SPTs to food allergens (cow’s milk and egg proteins) and specific IgE >0.35 kU/L against common food allergens (egg white, cow’s milk, cod, wheat, peanuts, and soybeans) until two years of age. | Lower incidence of SPT+ to egg in the L reuteri group and greater for milk but not significant. Significant lower levels of IgE to egg white in the L reuteri group at two years of age. |
| Source | Intervention Period | Test/ Control | Population/ Country | Strain(s)/Dose/Administration | Food Allergy-Related Variable | Results |
|---|---|---|---|---|---|---|
| West et al. (2013) [124] | Postnatal (4 to 13 months of age) | 84/87 | Healthy, full-term infants with no prior allergic manifestations; Sweden | Lactobacillus paracasei F19 1 × 108 CFU/day for nine months; mixed with infant cereals | Specific IgE to cow’s milk, egg white, wheat, codfish, and peanuts after a follow-up of 8–9 years | No significant differences in food allergies compared to placebo. |
| Taylor et al. (2007) [123] | Postnatal (from birth to 6 months of age) | 115/111 | Newborns of women with allergy; Australia | Lactobacillus acidophilus (LAVRI-A1) 3 × 109 CFU/day for six months; dissolved in 1–2 mL sterile water | Incidence of food allergy and evidence of allergen sensitization (SPT+) after a follow-up of 12 months | No significant differences in the rate of symptomatic food allergy. No significant differences in SPT+. |
| Viljanen et al. (2005) [125] | Postnatal | 88/76/74 | Infants with CMA (aged 1.4–11.9 months); Finland | Lactobacillus rhamnosus GG (ATCC 53103); 1 × 1010 CFU/day or a probiotic mixture (LGG; 1 × 1010 CFU/day; L. rhamnosus LC705 1 × 1010 CFU/day; Bifidobacterium breve Bb99 4 × 108 CFU/day, and Propionibacterium freudenreichii ssp. Shermanii JS 4 × 109 CFU/day) for four weeks; mixed with food | Fecal inflammatory markers as IgA, TNF- α, AT, and ECP | No significant differences. |
| Hol et al. (2008) | Postnatal | 60/59 | Infants younger than six months with a diagnosis of CMA; Netherlands | Lactobacillus casei CRL431 and Bifidobacterium lactis Bb-12; 109 CFU/day for each strain for 12 months; extensively hydrolyzed formula | Clinical tolerance to CM at 6 and 12 months after initial CMA diagnosis. | No significant differences. |
| Morisset et al. (2011) [127] | Postnatal (from birth until one year old) | 66/63 | Infants at high risk of allergy; France | Heat-killed Bifidobacterium breve C50 and Streptococcus thermophilus 065 (4.2 × 109 and 3.84 × 107 bacteria per 100 g of powder formula, respectively). EBF—administered to mothers; No EBF—administered to children (nonhypoallergenic formula) | Incidence of sensitization and allergy to CM and other foods (hen’s eggs, codfish, wheat flour, soy flour, and roasted peanuts) during the first 24 months of life. | Significant decrease in the proportion of SPT+ to CM in probiotic group after 12 months. Significant decrease in positive IgE tests against other foods than CM after 12 months. No significant decrease in incidence of CMA was observed. |
| Source | Intervention Period | Test/Control | Population/Country | Strain(s)/Dose/Administration | Allergic Outcome | Conclusions | Risk of Bias |
|---|---|---|---|---|---|---|---|
| Wickens et al. (2018) [143] | Prenatal and postnatal (from the 35th week of pregnancy to two years of age) | 157/158/159 | Infants at high risk of allergy; New Zealand | Mothers: Lactobacillus rhamnosus HN001; 6 × 109 CFU/day or Bifidobacterium lactis HN019; 6 × 109 CFU/each/day for seven months Infants: same mixture for two years | Lifetime prevalence of atopic sensitization, eczema, asthma, wheezing, hay fever, and rhinitis, and relative risks for point or 12-month prevalence at 11 years. | Significant reductions in the 12-month prevalence of eczema and hay fever at age 11 after HN001 supplementation. Significantly lower prevalence of atopic sensitization, eczema, and wheezing in HN001 group. No significant results for HN019 | Reduction in participation Rate. |
| Davies, et al. (2018) [145] | Prenatal and postnatal (from the 36th week of pregnancy to six months of age) | 220/234 | Healthy infants; UK | Mothers: Lactobacillus salivarius CUL61; L. paracasei CUL08; Bifidobacterium animalis ssp. lactis CUL34, and Bifidobacterium bifidum CUL20; 1 × 1010 CFU/day for four weeks Infants: same mixture for six months | Reports of eczema and asthma at five years using electronic follow-up data. | Higher prevalence of asthma in children in the probiotic arm at five years. | Potential intake of probiotics in both groups during follow-up. |
| Simpson et al. (2015) [147] | Prenatal and postnatal (from the 36th week of pregnancy to three months after delivery) | 211/204 | Healthy infants; Norway | Mothers: 250 mL of low-fat fermented milk containing Lactobacillus rhamnosus GG (LGG); 5 × 1010 CFU and Bifidobacterium animalis ssp. lactis Bb-12 (Bb-12) 5 × 109 CFU and Lactobacillus acidophilus La-5; 5 × 109 CFU for four months Infants: no probiotic supplementation | Cumulative incidence of AD and ARC, and the 12-month prevalence of asthma after six years of follow-up. | No significant differences in cumulative incidence of ARC and wheezing at six years of age. Significant differences in cumulative incidence of AD (39.1% in control vs. 29.3% in probiotic group). No statistically significant difference in 12-month prevalence of asthma. | High proportion of missing data. |
| Abrahamsson et al. (2013) [148] | Prenatal and postnatal (from the 36th week of pregnancy to one year of age) | 94/90 | Infants at high risk of allergy; Sweden | Mothers: Lactobacillus reuteri ATCC 57730; 1 × 108 CFU/day for four weeks Infants: same product for one year | Prevalence of asthma, ARC, allergic urticaria, and eczema after seven years of follow-up. | No significant differences between groups | Significantly greater intake of antibiotic during the first year of life in probiotic group. |
| Wickens et al. (2012) [144] | Prenatal and postnatal (from the 35th week of pregnancy to six months of age) | 157/158/159 | Infants at high risk of allergy; New Zealand | Mothers: Lactobacillus rhamnosus HN001; 6 × 109 CFU/day or Bifidobacterium lactis HN019; 6 × 109 CFU/each/day for seven months (if breastfeeding) Infants: same mixture for two years from birth | Cumulative prevalence of eczema and wheezing occurring between 2–3 months and at age four. Cumulative prevalence of atopic sensitization at two and four years old. RR for the effect of each probiotic on eczema, SCORAD (≥10), wheezing, ARC, and atopic sensitization after four years. | Cumulative prevalence of eczema significantly lower in HN001 group by four years. Some protection against developing SCORAD ≥10, wheezing, and atopic sensitization by age four years but not significant. Significantly reduced risks of having eczema and ARC in the last 12 months at age four. No significant effect of HN019 on any outcome. | Use of antibiotics between two and four years of age significantly higher in the B. lactis HN019 group compared to the placebo. |
| Abrahamsson et al. (2011) [149] | Prenatal and postnatal (from the 36th week of pregnancy to one year of age) | 81/80 | Infants at high risk of allergy; Sweden | Mothers: Lactobacillus reuteri ATCC 57730; 1 × 108 CFU/day for four weeks Infants: same product for one year | Circulating levels of Th1-associated CXC-chemokine ligand CXCL9, CXCL10, and CXCL11 and Th2-associated CC-chemokine ligand CCL17, CCL18, and CCL22 in venous blood at birth, six, 12, and 24 months of age. | Presence of L. reuteri in infant stool the first week of life related with low CCL17 and CCL22 and high CXCL11 levels at six months, but no differences in chemokine levels compared to the placebo group. | |
| Kukkonen et al. (2011) [150] | Prenatal and postnatal (from the 36th week of pregnancy to six months of age) | 64/67 | Infants at high risk of allergy; Finland | Mothers: Lactobacillus rhamnosus GG (ATCC 53103); 1 × 1010 CFU/day, L. rhamnosus LC705 (DSM 7061) 1 × 1010 CFU/day, Bifidobacterium breve Bb99 (DSM 13692) 4 × 108 CFU/day and Propionibacterium freudenreichii ssp. shermanii JS(DSM 7076) 4 × 109 CFU/day for seven months Infants: same mixture plus 0.8 g GOS for six months | Airway inflammation measured as levels of exhaled nitric oxide (FeNO) at age five. | No preventive effect on respiratory allergies. Probiotics had no significant effects on FeNO levels compared to placebo | |
| Dotterud et al. (2010) [146] | Prenatal and postnatal (from the 36th week of pregnancy to three months of age) | 211/204 | Infants both with and without a family history of atopy; Finland | Mothers: 250 mL of low-fat fermented milk containing Lactobacillus rhamnosus GG (LGG) 5 × 1010 CFU and Bifidobacterium animalis ssp. lactis Bb-12 (Bb-12) 5 × 1010 CFU and Lactobacillus acidophilus La-5; 5 × 109 CFU for four months Infants: no probiotic supplementation | Diagnosed AD, ARC, or asthma, during the first two years of life. | Significant reduction in the cumulative incidence of AD at two years of age. No reduction in the incidence of asthma or ARC | The nonsignificant results in asthma and ARC may be a result of insufficient statistical power. |
| Source | Intervention Period | Test/ Control | Population/ Country | Strain(s)/Dose/Administration | Allergic Outcome | Conclusions | Risk of Bias |
|---|---|---|---|---|---|---|---|
| Schmidt et al. (2019) [151] | Postnatal | 144/146 | Healthy infants aged 8–14 months; Denmark | Lactobacillus rhamnosus (LGG) and Bifidobacterium animalis ssp. lactis (BB-12) 1 × 109 CFU/each/day for six months; sachet | Incidence of allergic diseases, sensitization, and food reactions. | Significantly lower incidence of eczema in the probiotic group (4.2% vs 11.5%).No differences in the incidence of rhinitis, conjunctivitis asthma, sensitization, or food reactions. | Asthma, rhinitis, and conjunctivitis usually develop later in childhood.Detection of Bifidobacterium animalis ssp. lactis in the placebo group. |
| Cabana et al. (2017) [139] | Postnatal (from birth to six months of age) | 92/92 | Infants at high risk of allergy; USA | Lactobacillus rhamnosus GG (LGG); 1 × 1010 CFU/day and 225 mg of inulin; dissolve in 2 mL of pumped breast milk, partially hydrolyzed wheyinfant formula, or water | Incidence of eczema within two years of birth and incidences of asthma and allergic rhinitis within five years of birth. | No significant differences in cumulative incidence of eczema (probiotic: 30.9%; control: 28.7%) or asthma (probiotic: 9.7%; control: 17.4%). | Much larger sample size needed to detect a difference in the cumulative incidence of asthma. |
| Loo et al. (2014) [140] | Postnatal (from birth until six months of age) | 124/121 | Infants at high risk of allergy; Singapore | Bifidobacterium longum BL999; 1 × 107 CFU/g and Lactobacillus rhamnosus LPR; 2 × 107 CFU/g; cow’s milk-based infant formula; infants received at least 2.8 × 108 CFU/day | Prevalence of asthma, allergic rhinitis, eczema, and food allergy after five years of follow-up. | No significant differences between groups. | Most of the subjects continued to consume probiotics during follow-up. |
| West et al. (2013) [124] | Postnatal (from four to 13 months of age) | 84/87 | Healthy infants born vaginally; Sweden | Lactobacillus paracasei ssp paracasei F19; 1 × 108 CFU/g; mixed with infant cereals | Prevalence of eczema, allergic rhinitis, asthma, food allergy and lung function after a follow-up of 8–9 years. | No statistically significant differences between the groups. | Loss to follow-up of 60–70% of the original study population. |
| Author, (Year)/Country | Objective | Type of Study, Group (n) | Intervention | Probiotic Strain (Dose) | Primary Outcomes |
|---|---|---|---|---|---|
| Luoto (2010) Finland [163] | Evaluation of the impact of the prophylactic use in VLBW preterm infants of Lactobacillus rhamnosus GG (LGG) on NEC stage II or III in all five university hospital NICUs in Finland during the VON years. | RCS Prophylactic LGG group (418) Probiotic “on demand” group (1024) CC: 1900 | The incidence of NEC was analyzed in <30 weeks or <1500 g babies, from the national database and from the VON databases separately in all five level III NICUs and additionally in three groups according to the probiotic practice. | Lactobacillus rhamnosus GG (LGG); 6 × 109 CFU/day | The incidence of NEC was 4.6% vs. 3.3% vs. 1.8% in the prophylactic LGG group, the probiotics “on demand” group, and the no-probiotics group, respectively (p = 0.0090). LGG had no influence on the clinical course of NEC. |
| Braga (2011) Brazil [168] | Evaluation of the combined use of Lactobacillus casei and Bifidobacterium breve in the prevention of NEC stage ≥2 in VLBW preterm infants. | RDBPC PG: 119 CG: 112 | 28 days of treatment after second day of life in neonates with a birth weight of 750 to 1499 g. | Multi-strain probiotic: Lactobacillus casei; Bifidobacterium breve, 3.5 × 107 to 3.5 × 109 CFU/day | Confirmed cases of NEC occurred only in the control group (4/112). Infants in PG achieved full enteral feeding faster than CG (p = 0.02). |
| Hunter (2012) USA [158] | Evaluation of the use of Lactobacillus reuteri DSM 17938 on the rate of NEC in neonates at highest risk of developing NEC (BW ≤ 1000 g). | RCS PG: 79 CG: 232 | Groups separated based on the introduction of probiotic as routine prophylaxis. Treatment from first week of life until hospital discharge. | Lactobacillus reuteri DSM 17938; ~5.5 × 107 CFU/day | Significantly lower rates of NEC in the neonates who received L. reuteri (2/79 neonates (2.5%) vs. 35/232 untreated neonates (15.1%)). Rates of late-onset Gram-negative or fungal infections were not statistically different between treated and untreated groups (22.8 vs. 31%). No adverse events related to use of L. reuteri. |
| Li (2013) USA [162] | Evaluation of the efficacy of probiotic therapy in preventing NEC in VLBW infants. | RCS PG: 291 CG: 289 | Screening of patients admitted to the NICU over eight years. Probiotic administration was implemented as part of the standard care for NEC prevention. | Multi-strain probiotic: S. thermophilus; B. infantis; B. bifidum VLBW: 1.05 × 109 CFU/day ELBW: 0.5 × 109 CFU/day | The incidence of NEC was similar between the control group (2.8%) and probiotics group (2.4%) (hazard ratio, 1.15; 95% [CI], 0.42, 3.12). Mortality of NEC similar between groups (1 vs. 2, p = 1.000). Incidence of NEC scare was decreased, from 2.8% in the control group to 1.4% in the probiotics group, not significant. |
| Demirel (2013) Turkey [170] | Evaluation of the efficacy of Saccharomyces boulardii for reducing the incidence and severity of NEC in VLBW infants. | Prospective, blinded, randomized controlled trial PG: 136 CG: 135 | Treatment from the first feed (within 48 h of birth) until neonates were discharged. The primary outcomes were death or NEC (Bell’s stage ≥2), and secondary outcomes were feeding intolerance and clinical or culture-proven sepsis. | Saccharomyces boulardii; 5 × 1010 CFU/day | No significant difference in the incidence of death (3.7% vs. 3.6%, 95% CI of the difference = −5.20–5.25; p = 1.0) or incidence of stage ≥2 NEC (4.4% vs. 5.1%, 95% CI, −0.65–5.12; p = 1.0) between the PG and CG. Feeding intolerance and clinical sepsis were significantly lower in the probiotic group compared with control (22.9% vs. 29.23% and 34.8% vs. 47.8%). |
| Fernández-Carrocera (2013) Mexico [165] | Evaluation of the effectiveness of a multispecies probiotic in the prevention of NEC in newborns with birthweight <1500 g. | RDBPC PG: 75 CG: 75 | Patients randomized into two groups to receive either a daily feeding supplementation with a multispecies probiotic, 1 g/day, or the placebo. Unspecified treatment period. | Multi-strain probiotic: Lactobacillus acidophilus, 1.0 × 109 CFU/g; Lactobacillus rhamnosus, 4.4 × 108 CFU/g; Lactobacillus casei, 1.0 × 109 CFU/g; Lactobacillus plantarum, 1.76 × 108 CFU/g; Bifidobacterium infantis, 2.76 × 107 CFU/g Streptococcus thermophilus, 6.6 × 105 CFU/g | No differences detected in NEC risk reduction (RR: 0.54, 95% CI 0.21 to 1.39), trend in the reduction in NEC frequency in the studied cases: six (8%) vs. 12 (16%) in the CG. Fewer infants in the PG died or developed NEC vs. CG; RR 0.39 (95% CI 0.17 to 0.87). Lactobacillus or Bifidobacteria not present in blood cultures in cases of sepsis. |
| Serce (2013) Turkey [182] | To investigate the efficacy of S. boulardii in preventing NEC or sepsis in very-low-birth-weight infants. | RDBPC PG: 104 PG: 104 | VLBW neonates (BW ≤ 1500 g) treated from the first feed until discharge. The median duration of probiotic supplementation and follow-up was 44 days. The study was conducted in preterm infants (≤ 32 GWs, ≤ 1500 g birth weight). They were randomized either to receive feeding supplementation with S. boulardii 50 mg/kg every 12 h or a placebo, starting with the first feed and continuing until discharge. | Saccharomyces boulardii, 5 × 1010 CFU/day | Same incidence of stage ≥2 NEC in both groups (7/104; 6.7%). No differences between PG vs. CG in late-onset, culture-proven sepsis (18.3% vs. 24.3%, p = 0.29); 28.8% vs. 23%, p = 0.34), deaths (4.8% vs. 3.8%) or time to reach 100 mL/kg/day of oral feeding (day) (11 ± 7 vs. 12 ± 7, p = 0.37). |
| Bonsante (2013) France [157] | To report outcomes in infants receiving the probiotic cohort (PC) compared with the historical cohort. | RCS PG: 347 CG: 783 | Treatment with Lactobacillus rhamnosus Lcr35 in neonates born at 24 to 31 weeks’ gestation. Supplementation at the beginning of enteral feeding until a gestational age of 36 weeks or discharge. | Lactobacillus rhamnosus Lcr35, 4 × 108 CFU/day | Infants in PG presented a reduced rate of NEC (OR 0.20; 95% CI 0.07 to 0.58), mortality (OR 0.46; 95% CI 0.21 to 1.00), and LOS (OR 0.60; 95% CI 0.40 to 0.89) and achieved FEF significantly earlier (11.7 ± 10 vs. 16.5 ± 13.3; p = 0.01). IRB was significantly lower in PG (4.6% vs. 7.5%; p = 0.07) |
| Oncel (2014) Turkey [166] | To evaluate the effect of oral Lactobacillus reuteri in the frequency of NEC and/or death after seven days, frequency of proven sepsis, rates of feeding intolerance, and duration of hospital stay. | RDBPC PG: 200 CG: 200 | Treatment with Lactobacillus reuteri DSM 17938 in preterm infants (≤32 weeks). Supplementation started with the first feed and lasted until death or discharge. | Lactobacillus reuteri DSM 17938, 10 × 108 CFU/day (5 drops) | No statistically significant difference between PG and CG in terms of frequency of NEC stage ≥2 (4% vs. 5%; p = 0.63), overall NEC, or mortality rates (10% vs. 13.5%; p = 0.27). Significantly lower frequency of proven sepsis in PG vs. CG (6.5% vs. 12.5%; p = 0.041). Significant difference in rates of feeding intolerance (28% vs. 39.5%; p = 0.015) and duration of hospital stay (38 (10–131) vs. 46 (10–180) days; p = 0.022). |
| Janvier (2014) Canada [159] | To determine whether routine probiotic administration to very preterm infants would reduce the incidence of NEC without adverse consequences. | Prospective cohort study, with a historical comparison cohort PG: 264 CG: 317 | Treatment with a probiotic mixture as routine administration in preterm infants (≤32 weeks). Supplementation started with the first feed and went until death or 34 weeks postmenstrual age. Comparation with those admitted during the previous 17 months (no probiotic intake). | Multispecies probiotic: B. breve; B. bifidum; B. infantis; B. longum; L. rhamnosus GG (2 × 109 CFU/day) | Significant differences in NEC between PG and CG (5% vs. 10%; p < 0.05) and in the combined outcome of death or NEC (11% vs. 17%). No significant differences in death rate between groups. Improvements remained significant after adjustment for gestational age, intrauterine growth restriction, and sex, (OR for NEC, 0.51; 95% CI, 0.26–0.98; OR for death or NEC, 0.56; 95% CI, 0.33–0.93). No probiotic effect on healthcare-associated infection. |
| Hartel (2014) Germany [161] | To evaluate outcome data in an observational cohort of very-low-birth-weight infants of the German Neonatal Network stratified to prophylactic use of Lactobacillus acidophilus/Bifidobacterium infantis probiotics. | Observational, prospective, multicentric PG: 2310 CG: 518 | Treatment with a probiotic mixture as prophylactic in VLBW infants. Variability regarding dosage and time of probiotic administration: 1 × 1 capsule/day or 2 × 1/2 capsule/day) from day 2 or 3 of life for 14 days or until full enteral feeds. Primary outcome data of all eligible infants were determined according to the center-specific strategy. | Multispecies probiotic: L. acidophilus/B. infantis* | PG associated with a reduced risk for NE surgery (OR 0.58, 95% CI 0.37–0.91; p = 0.017), any abdominal surgery (OR 0.7, 95% CI 0.51–0.95; p = 0.02), and the combined outcome abdominal surgery and/or death (OR 0.43; 95% CI 0.33–0.56; p < 0.001). Probiotics had no effect on the risk of blood-culture confirmed sepsis. |
| Dang (2015) USA [169] | To investigate the role of probiotics supplementation in improving nutritional outcomes. | RCS PG: 108 CG: 113 | Treatment with probiotic mixture as routine administration in preterm infants (≤28 weeks and/or ≤1250g). Supplementation started with the first enteral feeding (48 h of life) and until 34 weeks postmenstrual age. Comparison with those admitted when probiotic intake was not instituted. | Multispecies probiotic: L. rhamnosus GG (LGG), 5 × 108 CFU/day; B. infantis, 5 × 108 CFU/day | OR of EUGR significantly lower in PG (–70%): (OR: 0.3, 95% CI: 0.138–0.611). Time to reach full feeds significantly reduced and weight gain significantly better in PG. Significant reduction in number of total parental nutrition days, central line days, nil per os days, and number of feeding intolerance episodes in PG. No significant difference in the incidence of NEC. |
| Dilli (2015) Turkey [164] | To test the efficacy of probiotic and prebiotic, alone or combined (symbiotic), on the prevention of NEC in VLBW infants. | RDBPC PG: 108 CG: 113 | VLBW infant randomized in four groups: G1: probiotic G2: Prebiotic (insulin: 900 mg) G3: Symbiotic: probiotic + prebiotic CG: placebo 1 sachet per day with breast milk or formula until discharge or death, for a maximum of eight weeks. | B. lactis, 5 × 109 CFU/day | Significantly lower NEC rate in G1 (2.0%) and G2 (4.0%) groups compared with G3 (12.0%) and placebo (18.0%) groups (p < 0.001). Significantly faster times to reach full enteral feeding (p < 0.001), lower rates of clinical nosocomial sepsis (p = 0.004), and lower mortality rates (p = 0.003) in G1, G2, and G3 groups vs. CG. Significantly shorter stays in the neonatal intensive care unit (p = 0.002) in G1, G2, and G3 groups vs. CG. |
| Lambaek (2016) Denmark [167] | To evaluate the benefit of implementing prophylactic use of probiotics as standard care for preterm infants. | Prospective cohort study, with a historical comparison cohort PG: 333 CG: 381 | Treatment with a probiotic mixture as routine administration in preterm infants (≤30 weeks). Supplementation started on the third day of life and continued until discharge from hospital. Comparison with a prior period without probiotic use. | Multispecies probiotic: B. lactis Bb12, 1 × 108 CFU/day; L. rhamnosus GG, 2 × 109 CFU/day | Incidence of NEC not significant between groups: (OR) 0.75, (p = 0.34, 95% CI: 0.43–1.30). Difference in mortality between groups not statistically significant: OR 0.92 (p = 0.55, 95% CI: 0.62–1.40). No side effects; no presence of probiotic strains in blood. |
| Robertson (2019) UK [160] | To compare the rates of NEC, LOS, and mortality for five-year periods before and after the implementation of routine daily multistrain probiotics administration in high-risk neonates. | RCS PG: 513 CG: 469 | Treatment with probiotic mixture as routine administration in preterm neonates at high risk of NEC: Supplementation started on postnatal day 1 and continued until 34 weeks postmenstrual age. Comparison with those admitted when probiotic intake was not instituted. | Multispecies probiotic: Mix 1: Lactobacillus acidophilus and Bifidobacterium bifidum, 1 × 109 CFU/day, each strain Mix 2: L. acidophilus; B. bifidum; and B. longum subsp infantis, ~0.5 × 109 CFU/day, each strain The mix used depended on the time of the study. | Rates of NEC significantly decreased from 7.5% (35/469) in CG to 3.1% (16/513) in PG (p = 0.014). Cases of LOS significantly decreased from 106/469 (22.6%) in CG to 59/513 (11.5%) in PG (p < 0.0001). All-cause mortality decreased from 67/469 (14.3%) to 47/513 (9.2%), although not significant. No episode of sepsis due to Lactobacillus or Bifidobacterium. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro-Tapia, E.; Sebastiani, G.; Sailer, S.; Almeida Toledano, L.; Serra-Delgado, M.; García-Algar, Ó.; Andreu-Fernández, V. Probiotic Supplementation during the Perinatal and Infant Period: Effects on gut Dysbiosis and Disease. Nutrients 2020, 12, 2243. https://doi.org/10.3390/nu12082243
Navarro-Tapia E, Sebastiani G, Sailer S, Almeida Toledano L, Serra-Delgado M, García-Algar Ó, Andreu-Fernández V. Probiotic Supplementation during the Perinatal and Infant Period: Effects on gut Dysbiosis and Disease. Nutrients. 2020; 12(8):2243. https://doi.org/10.3390/nu12082243
Chicago/Turabian StyleNavarro-Tapia, Elisabet, Giorgia Sebastiani, Sebastian Sailer, Laura Almeida Toledano, Mariona Serra-Delgado, Óscar García-Algar, and Vicente Andreu-Fernández. 2020. "Probiotic Supplementation during the Perinatal and Infant Period: Effects on gut Dysbiosis and Disease" Nutrients 12, no. 8: 2243. https://doi.org/10.3390/nu12082243
APA StyleNavarro-Tapia, E., Sebastiani, G., Sailer, S., Almeida Toledano, L., Serra-Delgado, M., García-Algar, Ó., & Andreu-Fernández, V. (2020). Probiotic Supplementation during the Perinatal and Infant Period: Effects on gut Dysbiosis and Disease. Nutrients, 12(8), 2243. https://doi.org/10.3390/nu12082243