1. Introduction
Cancer remains the major cause of human death in the world. Many factors and molecules are involved in the initiation, promotion and progression of cancer. One of the causal factors of tumor progression is the suppression of immune functions, namely the innate and acquired immunity. When fighting against tumor cells, this suppression means that immune functions cannot operate properly and their activities are reduced [
1]. Restoring or activating immune competence can thus be one of the primary strategies for active anticancer therapy. Immunotherapy, as an important anticancer modality, has received considerable attention and many immunotherapeutic drugs have been developed and used in clinics. Additionally, the use of alternative agents from natural products as immunomodulators against cancers, have also attracted great interest in the past [
2,
3].
Polysaccharides from mushrooms and microbial cell-wall components have been shown to be efficient immunomodulators. It has also been reported that extracts of Kumaizasa bamboo (
Sasa senanensis) leaves demonstrated anti-inflammatory properties [
4] and our previous studies clearly showed the strong effect of these bamboo leaf extracts to activate macrophages and neutral killer (NK) cells, inducing the production of proinflammatory cytokines, e.g., interleukin-12 (IL-12) and nitric oxide (NO), consequently preventing carcinogenesis and inhibiting tumor growth [
5].
Phellinus linteus is a medicinal mushroom that has been used in traditional medicine in China, Japan and Korea for centuries as an ailment drug for many diseases, such as gastroenteric dysfunction, diarrhea, hemorrhage and cancers. The activity of
Phellinus linteus against cancer has been reported in many studies using different cancers and it is thus considered as a promising anti-cancer agent [
6,
7,
8]. Though the mechanisms behind its anti-cancer activity are not fully clear, a previous study using extracts from fruit-bodies and the mycelium of
Phellinus linteus suggested that it could stimulate the hormonal and cell-mediated immune function, in addition to modulating the inflammatory reactions caused by a variety of stimuli, consequently suppressing tumor growth and metastasis [
9].
The Chaga mushroom (
Inonotus obliquus) is a type of fungus that grows mainly on the bark of birch trees in cold climates, such as Northern Europe, Siberia, Russia, Korea, Northern Canada and Alaska. It has been used in traditional medicine for centuries to treat various health problems, especially by Khanty people. In China, Korea, Japan, Russia, and the Baltics, extracts of Chaga mushroom have been used because of their favorable effects, for example, promoting lipid metabolism and modulating cardiac function, as well as antibacterial, anti-inflammatory, antioxidant, and antitumor activities [
10,
11,
12]. To date, several compounds have been identified from the Chaga mushroom, such as polysaccharides, triterpenes, polyphenols, betulin and betulinic acid, which are considered to be responsible for most of its therapeutic activities [
10,
11,
12].
Hot water extraction method is commonly used to obtain extracts from plants, however the efficacy and yield are relatively low. Previously we modified the extraction method using vigorous steam conditions at high temperature and high pressure (vigorous extract) for Kumaizasa (
Sasa senanesis) leaves, of which the extracts contained a larger quantity of active compounds than conventional hot-water extracts [
5]. Along this line, an extract mixture of
Phellinus linteus, Kumaizasa bamboo and Chaga mushroom using vigorous steam conditions was prepared. In this study, we investigated the activity of this extract mixture (MeshimaMax) for cancer prevention and treatment by using different solid tumor models, including chemical carcinogen induced cancer. The possible mechanisms involved in the activity of the extracts was examined especially focusing on their potential to stimulate innate immunity.
2. Materials and Methods
2.1. Materials
Dextran Sulfate Sodium Salt (DSS) was purchased from MP Biomedicals, LLC (Irvine, CA, USA). Latex beads-Rabbit IgG-FITC Complex was from Cayman chemical (Ann Arbor, MI, USA). Thiazolyl Blue Tetrazolium Bromide (MTT) was purchased from Sigma-Aldrich Chemical (St. Louis, MO, USA). Azoxymethane (AOM), 7,12-dimethylbenz anthracene (DMBA), cell culture medium (RPMI-1640, DMEM, DMEM:F12) were purchased from Fujifilm Wako Pure Chemical Corporation (Osaka, Japan). MeshimaMax was provided by ACSYS UN (Tokyo, Japan). Anticancer nanomedicine HPMA copolymer conjugated pirarubicin (P-THP) was synthesized in our laboratory as reported previously [
13].
2.2. Animals
ddY mice (male, 6-weeks old), ICR mice (male, 5-weeks old) and SD rats (female, 6-weeks old) were bought from SLC (Shizuoka, Japan). They were housed in groups of four or five per cage and were maintained at 22 ±10 °C and 55% ±5% relative humidity. Lighting was automatic on a 12-h light/dark cycle. All experiments were carried out according to the Laboratory Protocol of Animal Handling, Sojo University. Ethical approval was given by Sojo University, Animal Ethical Committee (decision no. P/009/2019) in 2019.
2.3. Cell Lines
RAW264.7 cells of mice macrophages were purchased from ATCC (Manassas, VA). Human ovarian cancer A2780 cells were distributed by Professor Kobayashi, Faculty of Medicine, Kagoshima University. Human cervical cancer HeLa cells were from Riken cell bank (Tsukuba, Japan). Mouse hepatocytes AML12 were kindly provided by Dr. Zhang of Department of toxicology, Anhui Medical University, China. Monkey kidney epithelial cells (CCL-81) and yeast (Saccharomyces cerevisiae) cells were kindly provided by Professor Yokomizo, Microbiology Laboratory, Faculty of Pharmaceutical Sciences, Sojo University. RAW264.7 cells were cultured in DMEM (high glucose) containing 10% fetal bovine serum (FBS), CCL-81 cells and A2780 cells were maintained in RPMI-1640 containing 10% FBS. HeLa cells were cultured in DMEM containing 10% FBS and AML12 cells were cultured in DMEM/F12 medium containing 10% FBS with 10 µg/mL insulin, 5.5 µg/mL transferrin, 5 ng/mL selenium, and 40 ng/mL dexamethasone. All of the cells were cultured at 37 °C with 5% CO2.
2.4. Antitumor Effect of MeshimaMax
The potential effect of MeshimaMax on tumor growth and carcinogenesis was investigated in various murine solid tumor models, including an AOM/DSS induced mouse colon cancer model, a DMBA induced rat breast cancer model and a mouse sarcoma S180 subcutaneously transplanted model.
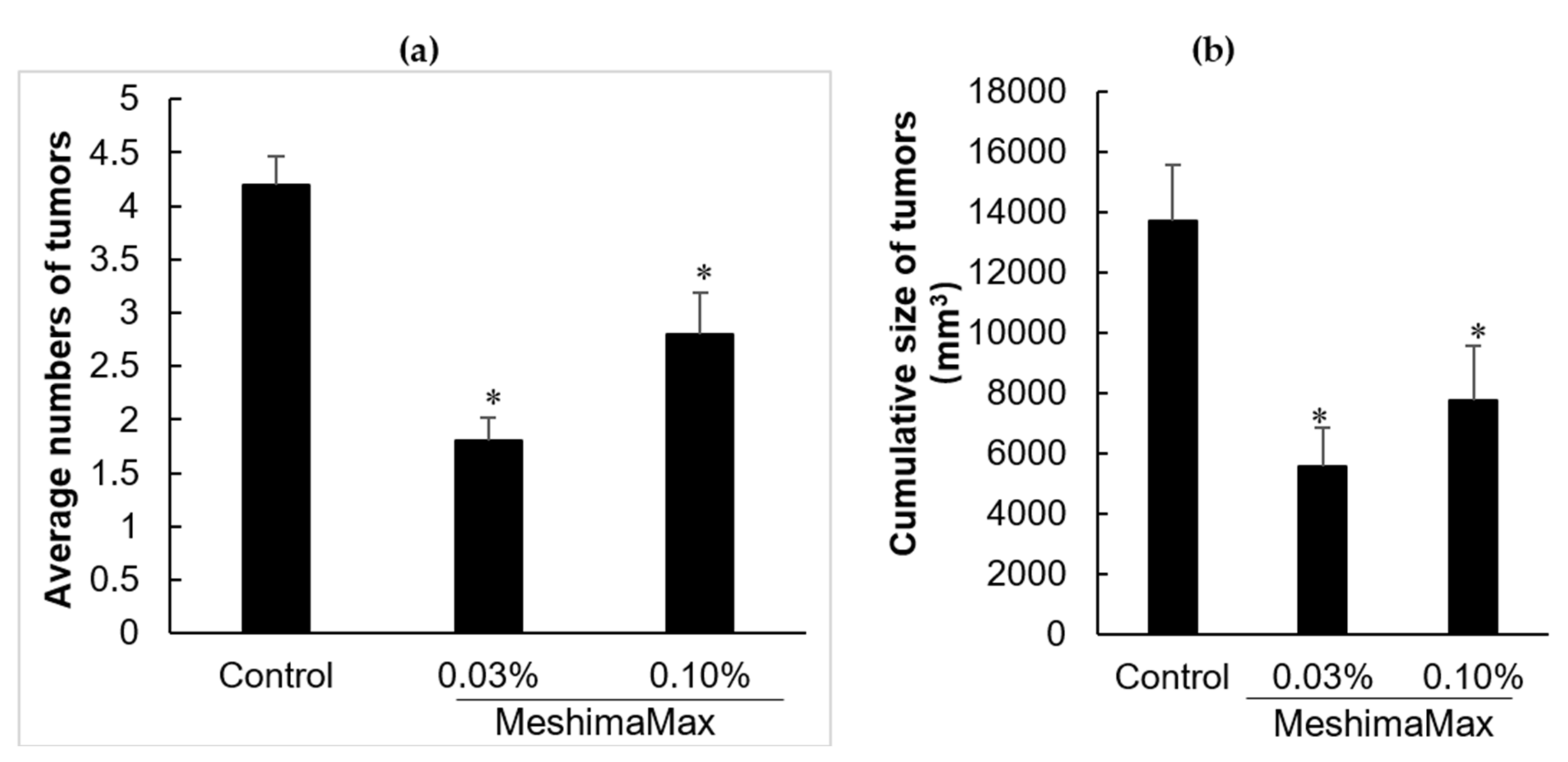
AOM/DSS induced mouse colon cancer model was prepared by administering AOM (10 mg/kg) intraperitoneally into ICR mice, after 1 week, 2% DSS was added into drinking water and applied to the mice for 1 week. In the MeshimaMax treatment group, different concentrations of MeshimaMax (0.03%, 0.1%, 0.3%) were applied to the mice as drinking during the entire experiment, from the time of AOM administration, during which it was mixed in the food when DSS was applied through drinking water. Twelve weeks after DSS administration, the mice were sacrificed and the number of colon cancers and the diameter of tumor nodules were measured.
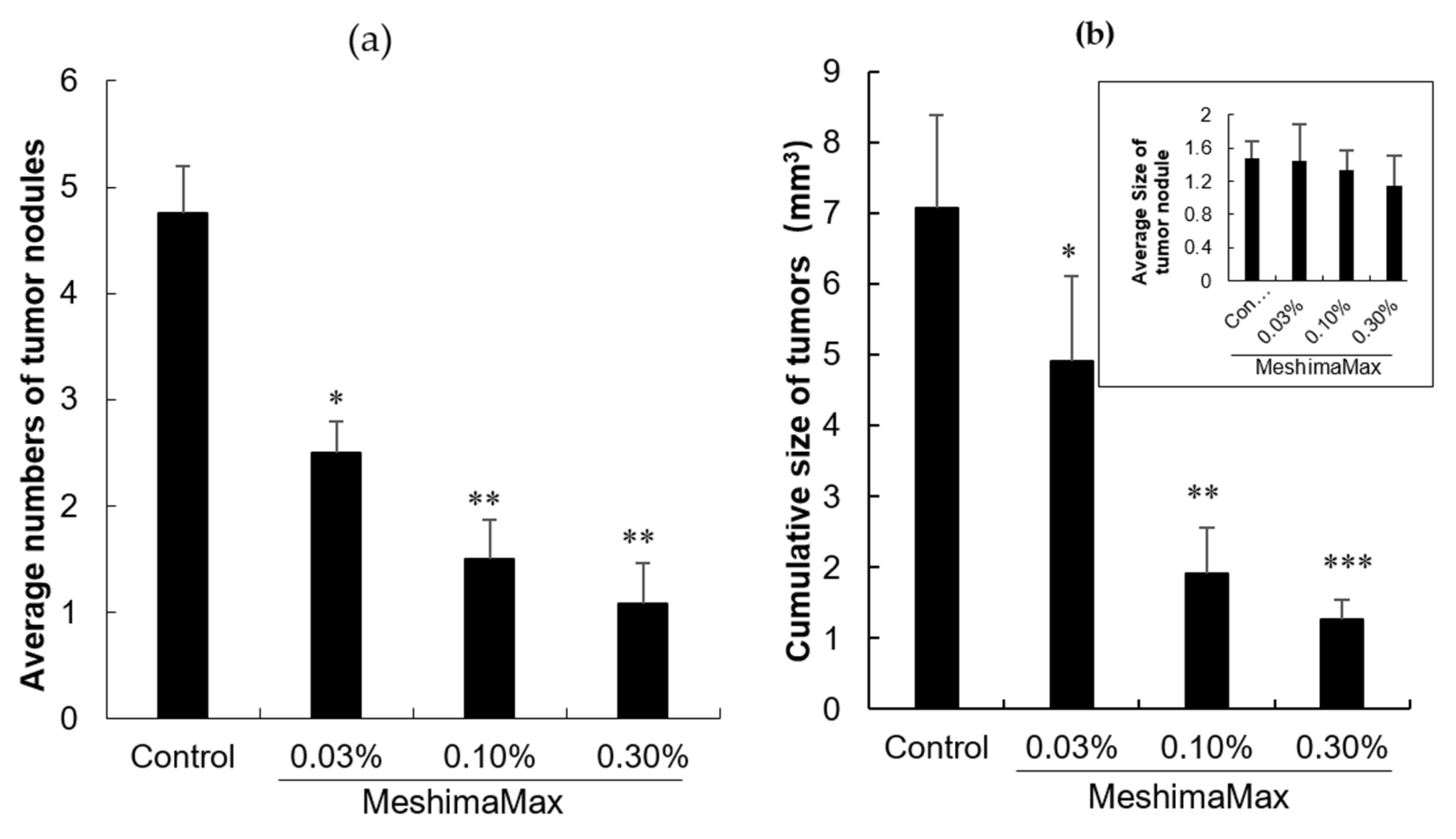
The DMBA induced rat breast cancer model was established by oral administration of DMBA (10 mg/mL/rat) which was dissolved in corn oil. For the treatment, MeshimaMax (0.03%, 0.1%, 0.3%) was added to the drinking water of rats during the entire experiment. During the experiment, the tumor development status was observed regularly and the tumor size and number were measured.
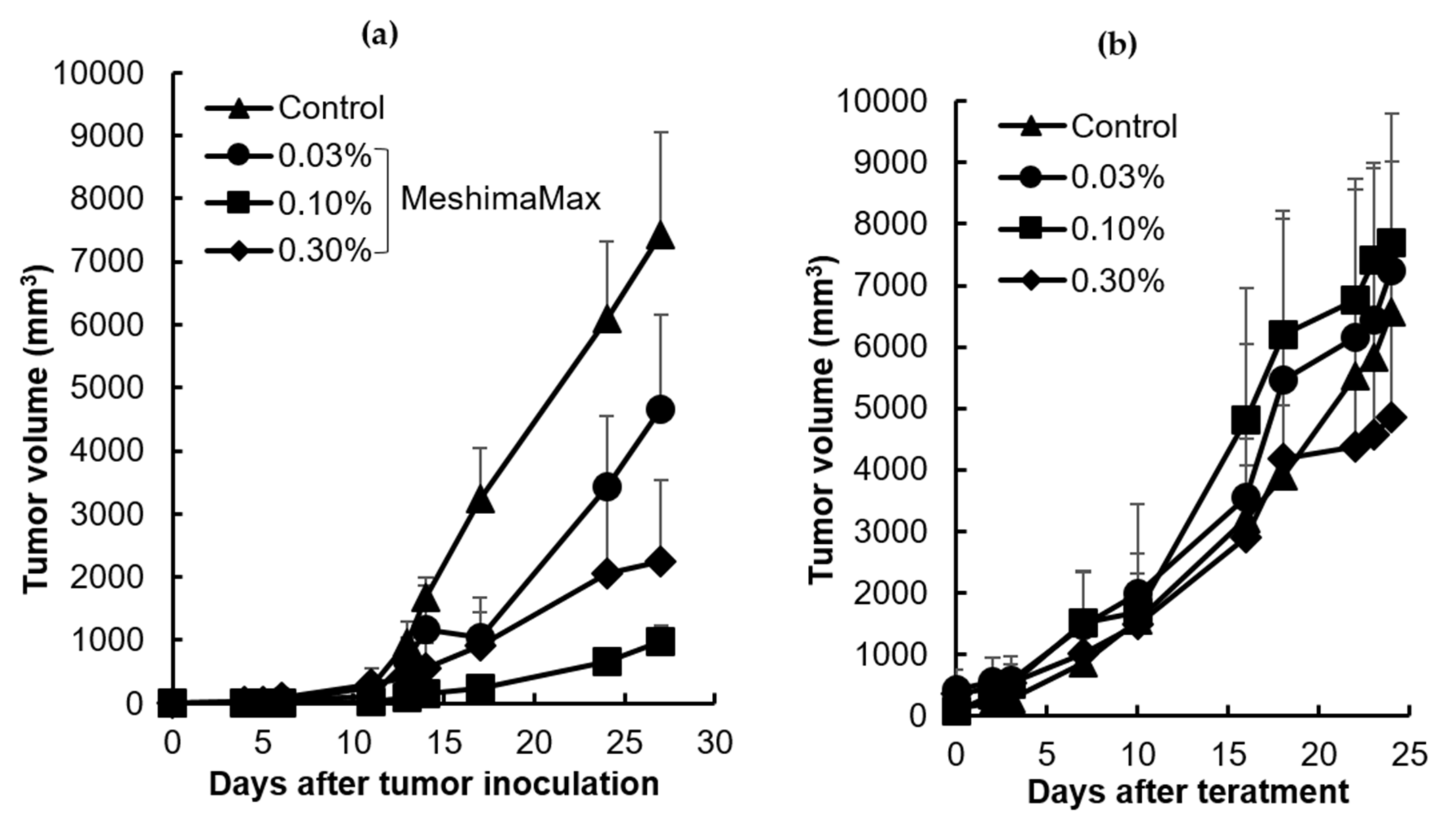
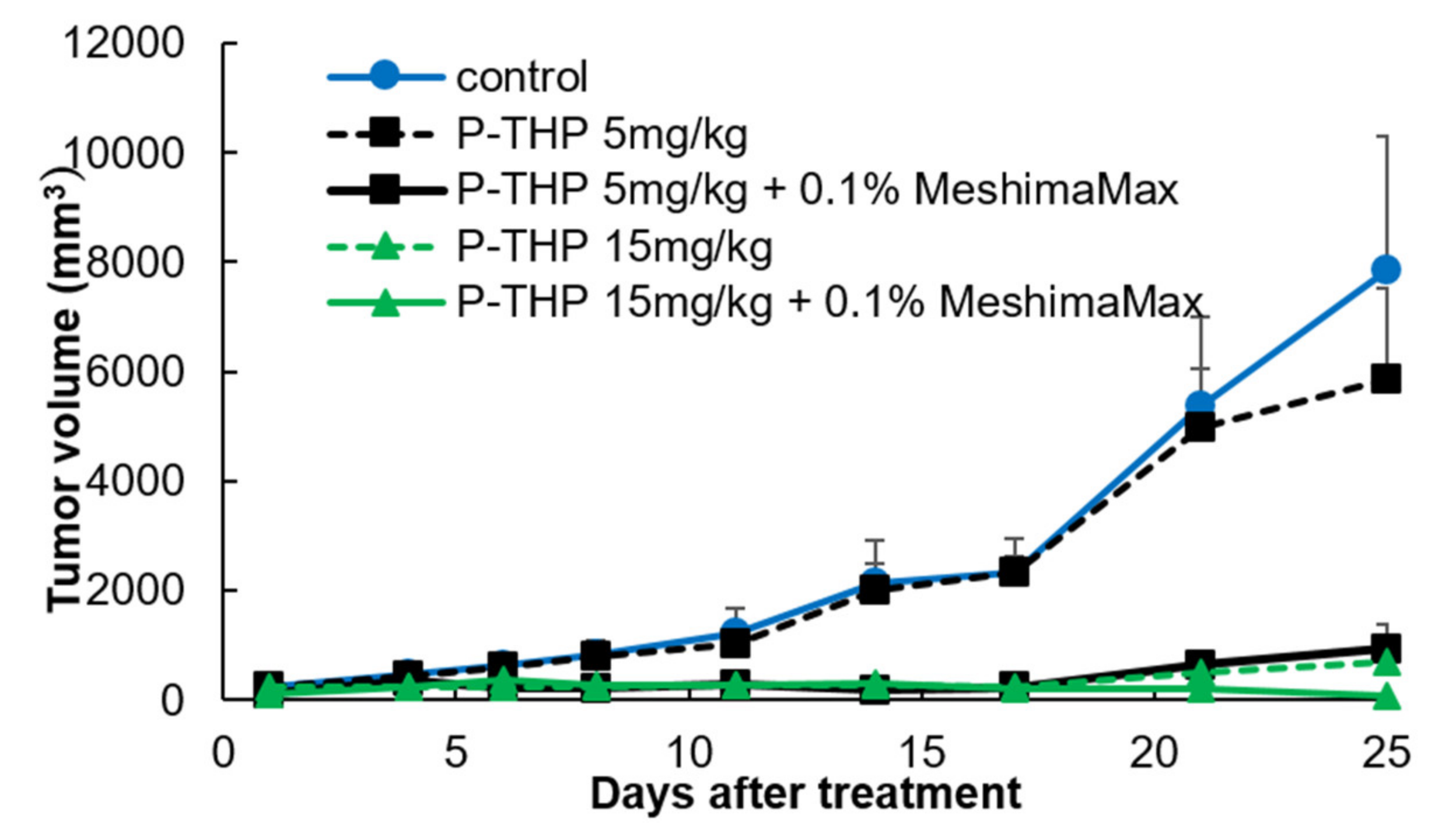
The mouse S180 tumor model was prepared by injecting the cancer cells (2 × 106/100 uL) subcutaneously into the dorsal skin of a ddY mouse. MeshimaMax (0.03%, 0.1%, 0.3%) was added to the drinking water of mice for treatment. In some experiments, MeshimaMax was administered from the day that the S180 cells were transplanted (preventive administration). In another set of experiments, MeshimaMax was applied one week after the transplantation of S180 cells when the diameter of the tumor had reached 6–8 mm (therapeutic administration). In a separate experiment, MeshimaMax was combined with anticancer nanomedicine P-THP, in which P-THP was injected i.v. 7 days after the inoculation of S180 cells, 0.1% MeshimaMax was then administered as drinking water till the end of the experiment. During the experiment, the tumor regression or growth was observed periodically and tumor size was measured.
2.5. Cytotoxicity Assay
The cytotoxicity of MeshimaMax was investigated in cancer cells (A2780, HeLa) as well as normal cells (CCL-81, AML12) by MTT assay. Briefly, Cells were seeded in 96-well plates (3000 cells/well), after overnight preincubation, MeshimaMax of difference concentrations was added and the cells were further cultured for 48 hours. Then, the medium was removed, and MTT was added to the final concentration of 0.56 mg/mL. After incubation for 4 hours at 37 °C, 5% CO2, the medium was removed, followed by adding 150 ul DMSO to each well. The cell viability was calculated by measuring the optical density (OD) at 570 nm.
2.6. Effect of MeshimaMax on Phagocytosis of Macrophages
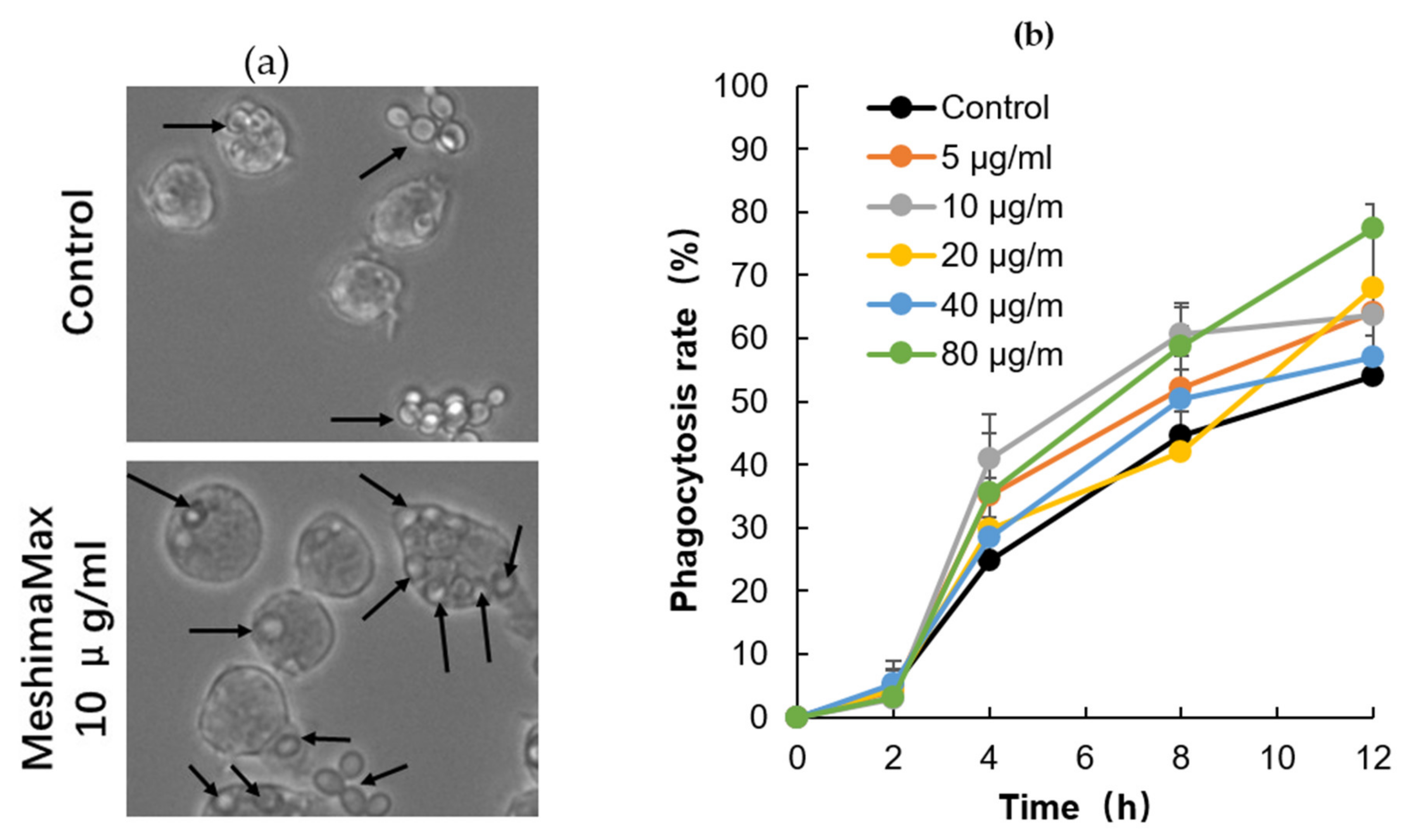
The effect of MeshimaMax on phagocytosis of macrophages was examined by using RAW264.7 cells. Cells (1 × 105/1.5 mL) were seeded on a 6-well plate, after overnight preincubation, MeshimaMax was added at different concentrations and the cells were cultured for 24 hours. Then, yeast cells (3 × 105 cells) were administered to the macrophage culture system. The macrophages incorporating yeast were observed and counted with a microscope (BZ-X700, Keyence Co. Ltd., Osaka, Japan). Two-hundred RAW264.7 cells were counted and the phagocytic ability of macrophages was calculated by the following equation, phagocytosis = A / 200 × 100%, in which A was the number of macrophages which took up yeast cells.
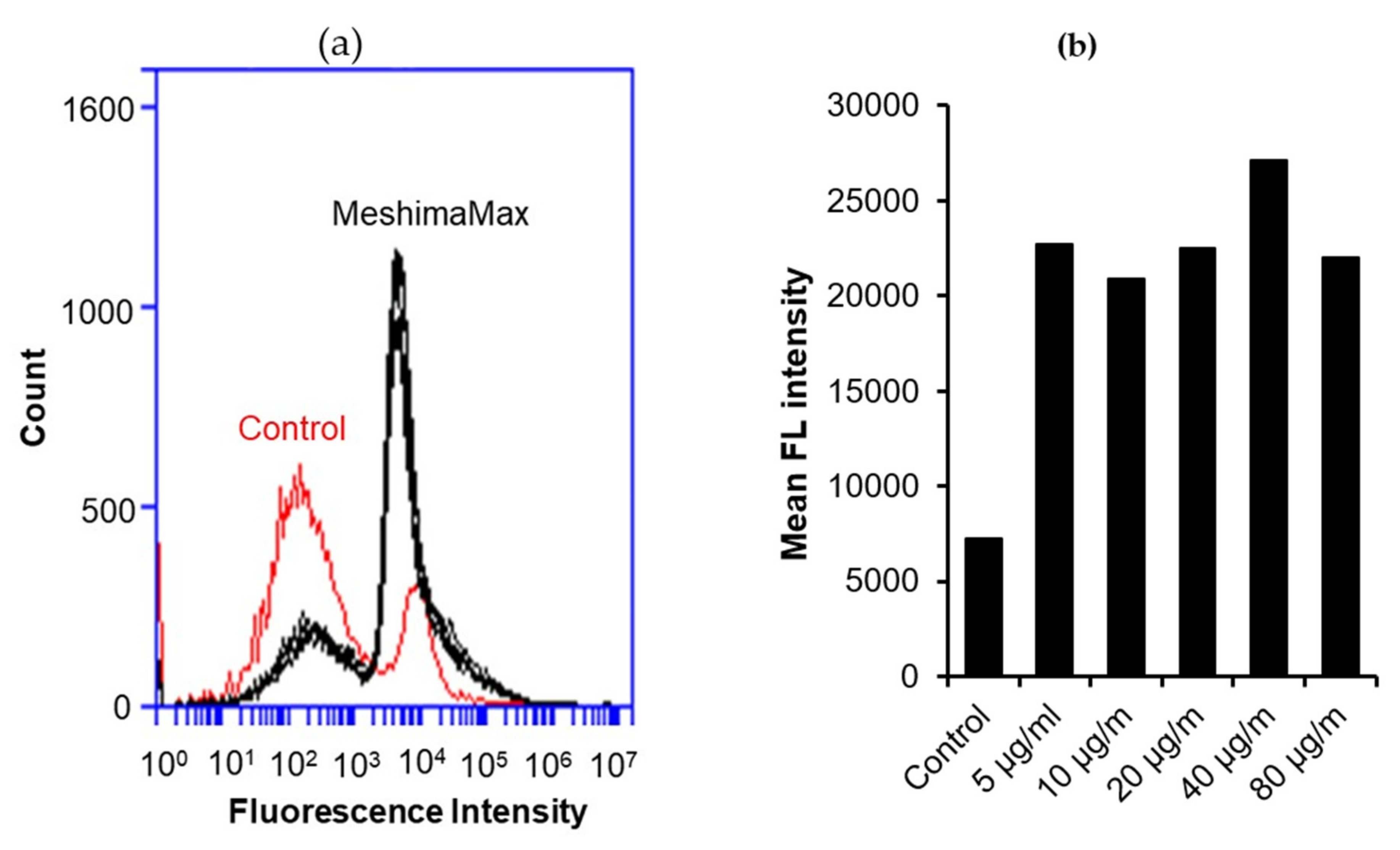
In a separate experiment, fluorescence nanoparticles (Latex beads-Rabbit IgG-FITC complex) were used instead of yeast, in which the phagocytosed nanoparticles were quantified by flow cytometry (Accuri™ C6 Plus, BD, Franklin Lakes, New Jersey, USA).
2.7. Statistical Analyses
All data were expressed as means ±SD. Data were analyzed using ANOVA followed by the Bonferroni multiple comparison test. A difference was considered statistically significant when P < 0.05.
4. Discussion
In a previous study, we reported the antitumor activity of bamboo (
Sasa senanensis) leaf extracts through immunopotentiation, i.e., inducing the production of inflammatory cytokines and activating macrophages and NK cells [
5]. Such natural products from plants with immunostimulating activity provide an alternative or supplemental anticancer regimen. They may particularly benefit those who are in an immunosuppressed state, such as cancer patients during and after chemotherapy and older populations. Investigation and development of such supplemental nutrients is thus of great importance. Along this line, in this study, we first demonstrated that MeshimaMax, the extract mixture of
Phellinus linteus, bamboo (
Sasa senanensis) leaf and Chaga mushroom
(Inonotus obliquus), exhibited a cancer preventive effect, which was probably at least partly through the activation of macrophages—the innate immunity.
The antitumor effect of MeshimaMax was considered best for tumor prevention, but not direct tumor killing. When a tumor grew to a palpable size, administration of MeshimaMax did not show a significant effect on tumor growth suppression (
Figure 3b). This was not consistent with our previous study using bamboo leaf extracts [
5], which may partly due to the different sizes of tumors at the starting point, the tumors were 6–8 mm in the present study which was larger than those used in the previous study (5–6 mm in diameter). However, more importantly when MeshimaMax was applied to the mice at the same time of tumor inoculation, remarkable suppression of tumor growth was achieved (
Figure 3a). To confirm the cancer preventive effect of MeshimaMax, we carried out experiments using carcinogen induced tumor models, i.e., AOM/DSS induced mouse colon cancer model and DMBA induced rat breast cancer model, which are optimal models to mimic the initiation and progression of human cancer. In both models, we found the formation and growth of tumors were significantly inhibited by MeshimaMax (
Figure 1 and
Figure 2). It should also be noted that breast cancer is the second most common cancer, after lung cancer, in the world and the leading cause of cancer deaths in women [
14], while colon cancer has recently become the second most common cause of cancer in women and the third most common in men [
15]. Our current results may thus offer options to prevent the incidence of those cancers, especially for those of high risk. The cancer preventive effect of MeshimaMax was similarly observed in chemical carcinogenesis in both breast cancer and colon cancer, as well as transplanted mouse sarcoma, suggesting the effect is not tumor type-specific, but is applicable to many types of cancers which warrants further investigations. More importantly, though MeshimaMax alone did not show a direct antitumor effect, the combination therapy of MeshimaMax with anticancer drug P-THP significantly improved the therapeutic effect of P-THP (
Figure 4), suggesting its potential application as a supplement regimen during anticancer chemotherapy.
Regarding the mechanisms of MeshimaMax on cancer prevention, along with our previous study using bamboo (
Sasa senanensis) leaf extracts [
5], we focused on its potential for activation of innate immunity and investigated its effect on macrophage activation by examining the phagocytotic activity of macrophages. As expected, we found significantly increased phagocytosis of cultured macrophages under the treatment of MeshimaMax, which was demonstrated by using both living yeast and the FITC-labeled latex nanoparticles (
Figure 6 and
Figure 7). Phagocytosis is one of the most important functions of macrophages against microbial, foreign bodies and damaged cells including cancer cells in innate immunity. It is thus reasonable that MeshimaMax fulfills its cancer preventive effect through macrophage activation. It has been also known that macrophages have two major activation phenotypes upon different stimuli. Firstly, macrophage polarization, where M1 macrophages show mostly inflammatory properties excreting inflammatory cytokines such as IL-2, IL-6, inducible NO synthase and having high phagocytotic activity to clear microbial and damaged cells. Secondly, M2 macrophages exhibit mainly anti-inflammatory effects by generating anti-inflammatory cytokines such as IL-10, a transforming growth factor for supporting tissue repair [
16]. We thus consider that MeshimaMax may induce the activation of macrophages towards M1 polarization, which will be a forthcoming challenge in our future study. Many studies have reported the versatile effects of
Phellinus linteus and its bioactive components including anti-inflammatory activity, anti-oxidative activity, anti-microbial activity and anticancer activity etc., in which immunomodulation is one of the critically important mechanisms involved in its functions [
17].
Phellinus linteus could enhance the expression and activation of macrophages and NK cells, upregulating the innate immunity [
18,
19], our present study confirmed this notion.
Phellinus linteus is also known to increase the proliferation and activity of antigen presentation cells, B lymphocytes, as well as T lymphocytes [
20,
21,
22], suggesting its effect on acquired immunity. Accordingly, the immunoregulating effect of MeshimaMax may also include multiple mechanisms, the forthcoming comprehensive study will focus in depth on the immunomodulating mechanisms of MeshimaMax.
However, in this study, we found the effect of MeshimaMax was not in a linear dose-dependent manner in either in vivo (
Figure 1,
Figure 2 and
Figure 3) or in vitro studies (
Figure 6 and
Figure 7). Relatively lower concentrations (e.g., 0.03%, 0.1%) showed better effects whereas higher concentrations (e.g., 0.3%) resulted in no significant increase in effect (
Figure 1) or, on the contrary, a decreased effect (
Figure 2 and
Figure 3). Actually, similar results were reported in our previous studies using Kumaizasa bamboo leaf extract [
5]. Though the present study and our previous study [
5] strongly suggest the immunostimulating, proinflammatory activity of the tested extracts, an earlier study also reported the anti-inflammatory effect of bamboo leaf extract [
4]. Accordingly, we considered that the effect of MeshimaMax, similar to Kumaizasa bamboo leaf extract, may be an inverted “U-shaped” dose-effect, which low or moderate concentrations exhibit proinflammatory activity or M1 polarization of macrophages, whereas higher doses may inhibit inflammatory reaction pushing the macrophages to M2 polarization. Such “U-shaped” or convex “U-shaped” dose-effects are common biological events, particularly for chemicals with hormonal activities [
23]. However, the detailed mechanisms involved in the dose-effect in the present study are not clear and further investigations are warranted to elucidate this issue. More importantly, we also found the dose responses varied in different models or animal species, for example, SD rat models seemed to respond at much lower doses (i.e., 0.03%) (
Figure 1), whereas balb/c mice showed almost a linear effect up to 0.3% of MeshimaMax (
Figure 2), these findings may reflect the different immune responses of different species of animals, which should be considered in future studies. In other words, overdosing of immunopotentiator may exhibit opposite functions.
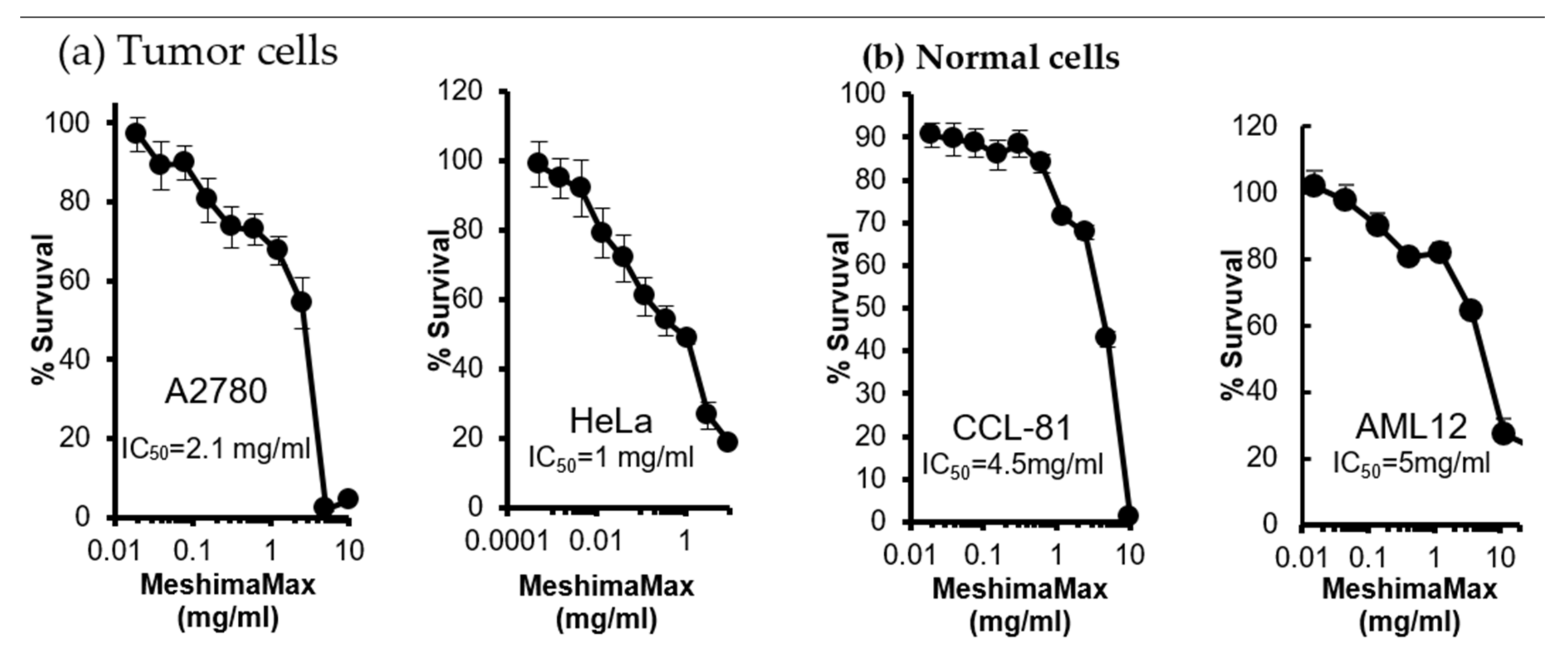
Additionally, in this study, we also examined the cytotoxicity of MeshimaMax on cancer cells, as well as normal cells. Here, the IC
50 was more than 1 mg/mL which is an extremely high concentration compared with that of macrophage activation (
Figure 5). This suggests that MeshimaMax does not have an apparent direct antitumor effect. However, interestingly, the IC
50 against normal cells was 2–3 times higher than that of cancer cells (
Figure 5). This finding indicates that normal cells are more tolerant to MeshimaMax, namely MeshimaMax is a friendly, non-toxic supplement to our bodies.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}