Casein Protein Processing Strongly Modulates Post-Prandial Plasma Amino Acid Responses In Vivo in Humans

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
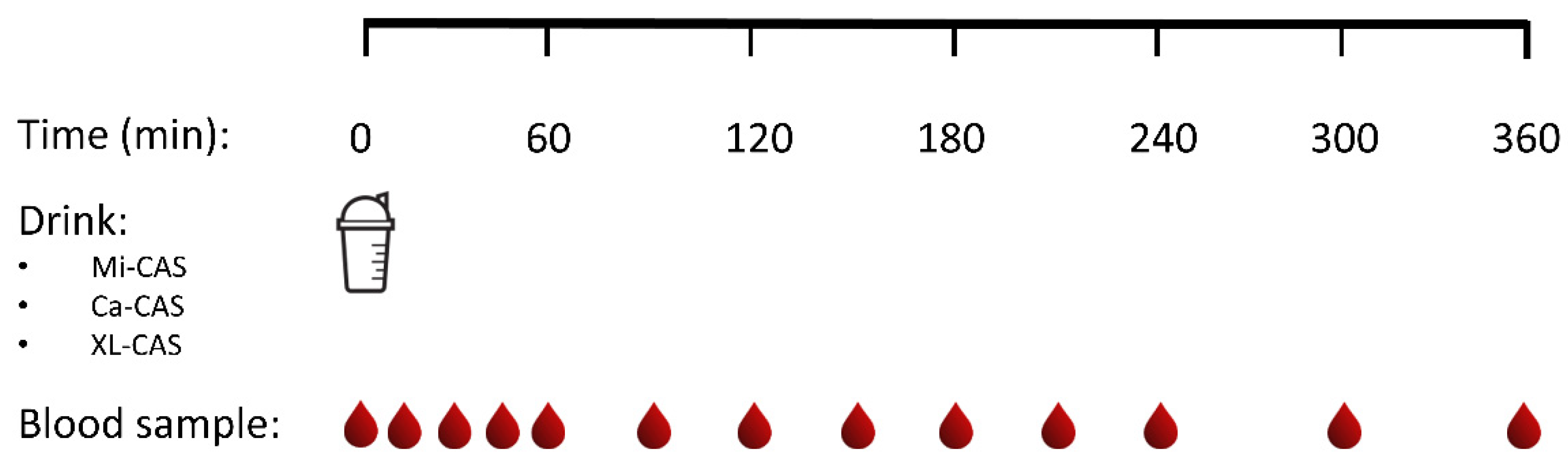
2.2. Design
2.3. Pretesting
2.4. Standardization of Physical Activity and Diet
2.5. Experimental Procedures
2.6. Protein
2.7. Plasma Analysis
2.8. Solubility of Protein Ingredients
2.9. Statistical Analysis
3. Results
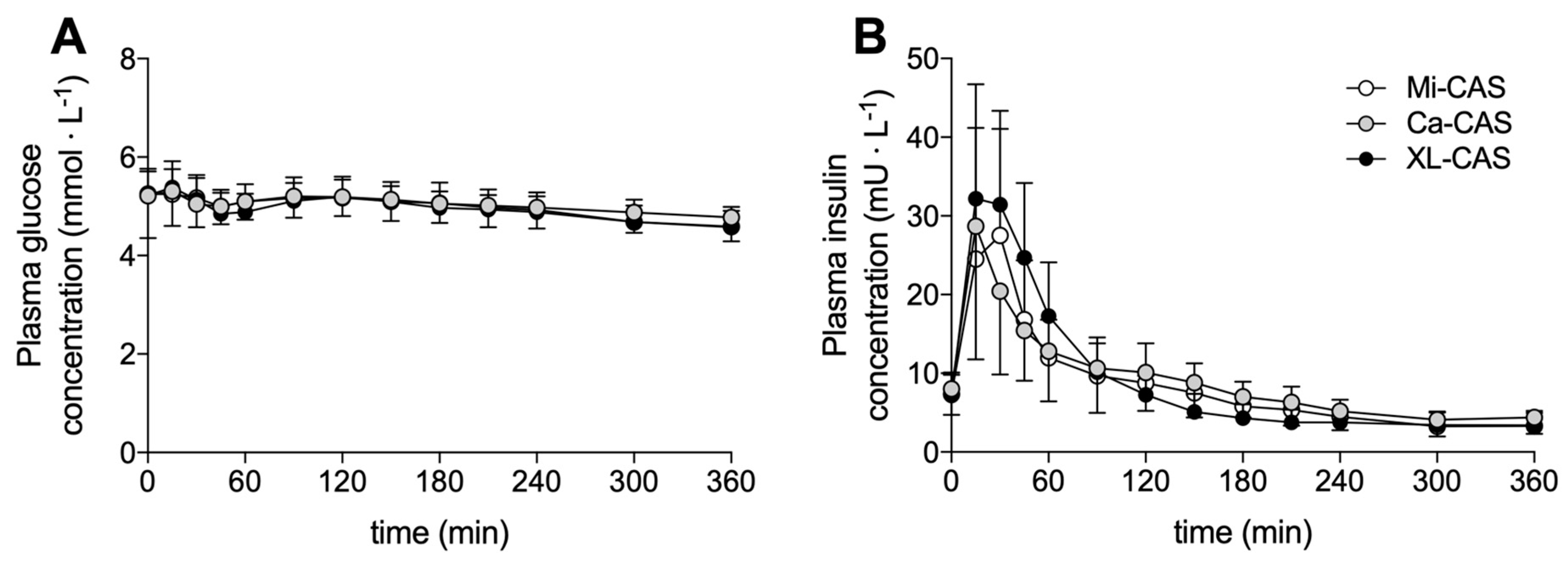
3.1. Plasma Glucose and Insulin Concentrations
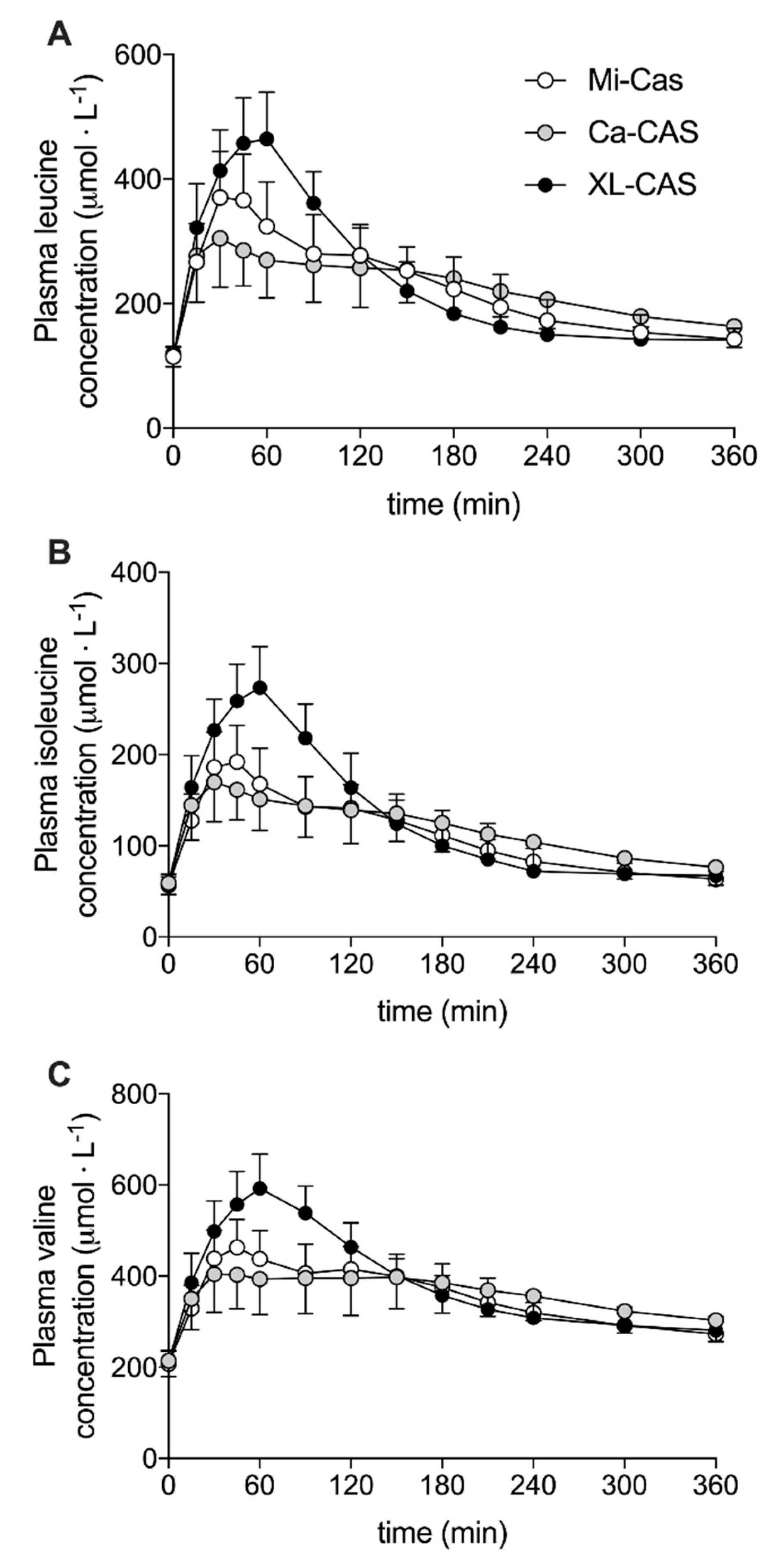
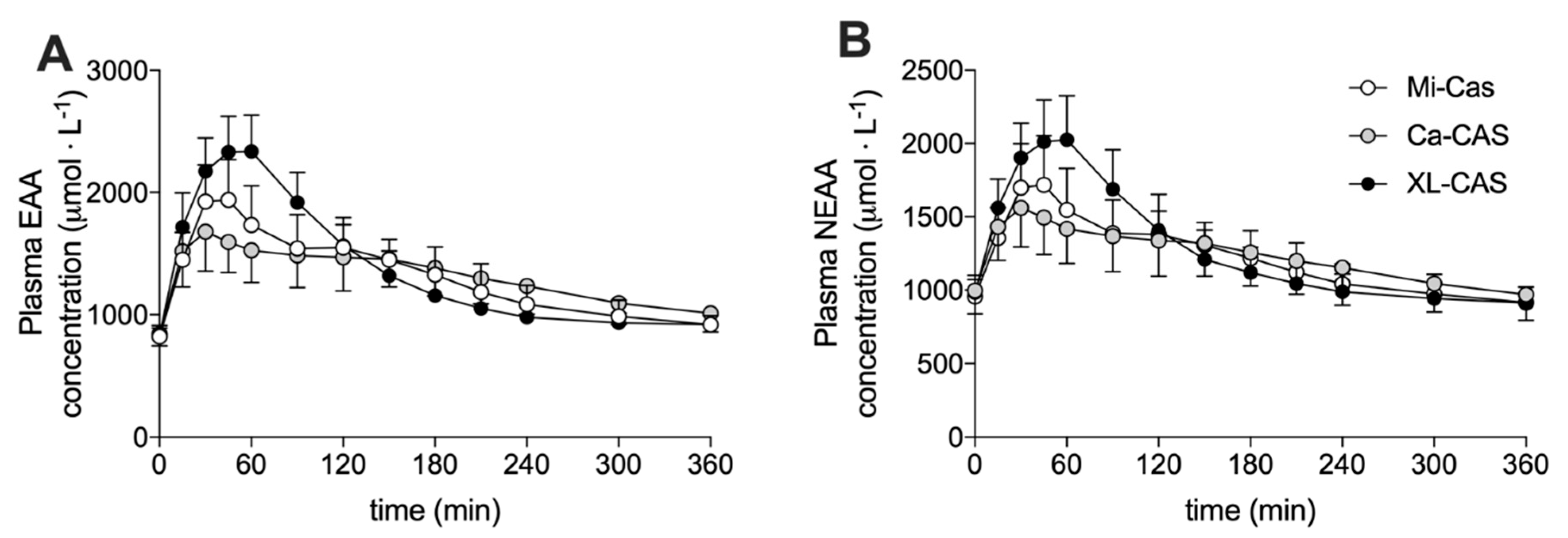
3.2. Plasma Amino Acid Concentrations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Biolo, G.; Tipton, K.D.; Klein, S.; Wolfe, R.R. An abundant supply of amino acids enhances the metabolic effect of exercise on muscle protein. Am. J. Physiol. Content 1997, 273, E122–E129. [Google Scholar] [CrossRef] [PubMed]
- Groen, B.B.L.; Horstman, A.M.; Hamer, H.M.; De Haan, M.; Van Kranenburg, J.; Bierau, J.; Poeze, M.; Wodzig, W.K.W.H.; Rasmussen, B.B.; Van Loon, L.J.C. Post-Prandial Protein Handling: You Are What You Just Ate. PLoS ONE 2015, 10, e0141582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trommelen, J.; Betz, M.; Van Loon, L.J.C. The Muscle Protein Synthetic Response to Meal Ingestion Following Resistance-Type Exercise. Sports Med. 2019, 49, 185–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koopman, R.; Crombach, N.; Gijsen, A.P.; Walrand, S.; Fauquant, J.; Kies, A.K.; Lemosquet, S.; Saris, W.H.; Boirie, Y.; Van Loon, L.J.C. Ingestion of a protein hydrolysate is accompanied by an accelerated in vivo digestion and absorption rate when compared with its intact protein. Am. J. Clin. Nutr. 2009, 90, 106–115. [Google Scholar] [CrossRef] [Green Version]
- West, D.W.; Burd, N.A.; Coffey, V.G.; Baker, S.K.; Burke, L.M.; Hawley, J.A.; Moore, D.R.; Stellingwerff, T.; Phillips, S.M. Rapid aminoacidemia enhances myofibrillar protein synthesis and anabolic intramuscular signaling responses after resistance exercise. Am. J. Clin. Nutr. 2011, 94, 795–803. [Google Scholar] [CrossRef] [Green Version]
- Fujita, S.; Rasmussen, B.B.; Cadenas, J.G.; Grady, J.J.; Volpi, E. Effect of insulin on human skeletal muscle protein synthesis is modulated by insulin-induced changes in muscle blood flow and amino acid availability. Am. J. Physiol. Metab. 2006, 291, E745–E754. [Google Scholar] [CrossRef] [Green Version]
- Trommelen, J.; Groen, B.B.L.; Hamer, H.M.; De Groot, L.C.P.G.M.; Van Loon, L.J.C. Mechanisms in Endocrinology: Exogenous insulin does not increase muscle protein synthesis rate when administered systemically: A systematic review. Eur. J. Endocrinol. 2015, 173, R25–R34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickinson, J.M.; Rasmussen, B.B. Amino acid transporters in the regulation of human skeletal muscle protein metabolism. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 638–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuthbertson, D.J.; Smith, K.; Babraj, J.; Leese, G.; Waddell, T.; Atherton, P.; Wackerhage, H.; Taylor, P.M.; Rennie, M.J. Anabolic signaling deficits underlie amino acid resistance of wasting, aging muscle. FASEB J. 2004, 19, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Trommelen, J.; Kouw, I.W.K.; Holwerda, A.M.; Snijders, T.; Halson, S.L.; Rollo, I.; Verdijk, L.B.; Van Loon, L.J.C. Presleep dietary protein-derived amino acids are incorporated in myofibrillar protein during postexercise overnight recovery. Am. J. Physiol. Metab. 2018, 314, E457–E467. [Google Scholar] [CrossRef] [PubMed]
- Holwerda, A.M.; Paulussen, K.J.M.; Overkamp, M.; Goessens, J.P.B.; Kramer, I.F.; Wodzig, W.K.W.H.; Verdijk, L.B.; Van Loon, L.J.C. Dose-Dependent Increases in Whole-Body Net Protein Balance and Dietary Protein-Derived Amino Acid Incorporation into Myofibrillar Protein during Recovery from Resistance Exercise in Older Men. J. Nutr. 2019, 149, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.R.; Robinson, M.J.; Fry, J.L.; Tang, J.E.; Glover, E.I.; Wilkinson, S.B.; Prior, T.; Tarnopolsky, M.A.; Phillips, S.M. Ingested protein dose response of muscle and albumin protein synthesis after resistance exercise in young men. Am. J. Clin. Nutr. 2008, 89, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.E.; Moore, D.R.; Kujbida, G.W.; Tarnopolsky, M.A.; Phillips, S.M. Ingestion of whey hydrolysate, casein, or soy protein isolate: Effects on mixed muscle protein synthesis at rest and following resistance exercise in young men. J. Appl. Physiol. 2009, 107, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Boirie, Y.; Dangin, M.; Gachon, P.; Vasson, M.-P.; Maubois, J.-L.; Beaufrère, B. Slow and fast dietary proteins differently modulate postprandial protein accretion. Proc. Natl. Acad. Sci. USA 1997, 94, 14930–14935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pennings, B.; Boirie, Y.; Senden, J.M.; Gijsen, A.P.; Kuipers, H.; Van Loon, L.J.C. Whey protein stimulates postprandial muscle protein accretion more effectively than do casein and casein hydrolysate in older men. Am. J. Clin. Nutr. 2011, 93, 997–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorissen, S.H.M.; Trommelen, J.; Kouw, I.W.K.; Holwerda, A.M.; Pennings, B.; Groen, B.B.L.; Wall, B.T.; Churchward-Venne, T.A.; Horstman, A.M.H.; Koopman, R.; et al. Protein Type, Protein Dose, and Age Modulate Dietary Protein Digestion and Phenylalanine Absorption Kinetics and Plasma Phenylalanine Availability in Humans. J. Nutr. 2020. [Google Scholar] [CrossRef] [PubMed]
- A Burd, N.; Yang, Y.; Moore, D.R.; Tang, J.E.; Tarnopolsky, M.A.; Phillips, S.M. Greater stimulation of myofibrillar protein synthesis with ingestion of whey protein isolate v. micellar casein at rest and after resistance exercise in elderly men. Br. J. Nutr. 2012, 108, 958–962. [Google Scholar] [CrossRef] [Green Version]
- Ye, A.; Cui, J.; Dalgleish, D.; Singh, H. The formation and breakdown of structured clots from whole milk during gastric digestion. Food Funct. 2016, 7, 4259–4266. [Google Scholar] [CrossRef]
- Dalgleish, D.G.; Corredig, M. The Structure of the Casein Micelle of Milk and Its Changes during Processing. Annu. Rev. Food Sci. Technol. 2012, 3, 449–467. [Google Scholar] [CrossRef]
- Badem, A.; Uçar, G. Production of caseins and their usages. Int. J. Food Sci. Nutr. 2017, 2, 4–9. [Google Scholar]
- Miranda, G.; Pélissier, J.-P. In vivo studies on the digestion of bovine caseins in the rat stomach. J. Dairy Res. 1981, 48, 319–326. [Google Scholar] [CrossRef]
- Phillips, S.M. A comparison of whey to caseinate. Am. J. Physiol. Metab. 2011, 300, E610. [Google Scholar] [CrossRef] [PubMed]
- Buchert, J.; Ercili-Cura, D.; Ma, H.; Gasparetti, C.; Monogioudi, E.; Faccio, G.; Mattinen, M.; Boer, H.; Partanen, R.; Selinheimo, E.; et al. Crosslinking Food Proteins for Improved Functionality. Annu. Rev. Food Sci. Technol. 2010, 1, 113–138. [Google Scholar] [CrossRef] [PubMed]
- Juvonen, K.; Lille, M.E.; E Laaksonen, D.; Mykkänen, H.; Niskanen, L.K.; Herzig, K.-H.; Poutanen, K.; Karhunen, L.J. Crosslinking with transglutaminase does not change metabolic effects of sodium caseinate in model beverage in healthy young individuals. Nutr. J. 2012, 11, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorissen, S.; A Burd, N.; Hamer, H.M.; Gijsen, A.P.; Groen, B.B.; Van Loon, L.J.C. Carbohydrate Coingestion Delays Dietary Protein Digestion and Absorption but Does Not Modulate Postprandial Muscle Protein Accretion. J. Clin. Endocrinol. Metab. 2014, 99, 2250–2258. [Google Scholar] [CrossRef]
- Holwerda, A.M.; Lenaerts, K.; Bierau, J.; Van Loon, L.J.C. Body Position Modulates Gastric Emptying and Affects the Post-Prandial Rise in Plasma Amino Acid Concentrations Following Protein Ingestion in Humans. Nutrients 2016, 8, 221. [Google Scholar] [CrossRef] [Green Version]
- Holwerda, A.M.; Lenaerts, K.; Bierau, J.; Wodzig, W.K.; Van Loon, L.J.C. Food ingestion in an upright sitting position increases postprandial amino acid availability when compared with food ingestion in a lying down position. Appl. Physiol. Nutr. Metab. 2017, 42, 738–743. [Google Scholar] [CrossRef] [Green Version]
- Pennings, B.; Koopman, R.; Beelen, M.; Senden, J.M.; Saris, W.H.; Van Loon, L.J.C. Exercising before protein intake allows for greater use of dietary protein–derived amino acids for de novo muscle protein synthesis in both young and elderly men. Am. J. Clin. Nutr. 2010, 93, 322–331. [Google Scholar] [CrossRef]
- Res, P.T.; Groen, B.; Pennings, B.; Beelen, M.; Wallis, G.A.; Gijsen, A.P.; Senden, J.M.G.; Van Loon, L. Protein Ingestion before Sleep Improves Postexercise Overnight Recovery. Med. Sci. Sport. Exerc. 2012, 44, 1560–1569. [Google Scholar] [CrossRef] [Green Version]
- Gorissen, S.H.; Burd, N.A.; Kramer, I.F.; Van Kranenburg, J.; Gijsen, A.P.; Rooyackers, O.; Van Loon, L. Co-ingesting milk fat with micellar casein does not affect postprandial protein handling in healthy older men. Clin. Nutr. 2017, 36, 429–437. [Google Scholar] [CrossRef] [Green Version]
- Traylor, D.A.; Gorissen, S.H.; Hopper, H.; Prior, T.; McGlory, C.; Phillips, S.M. Aminoacidemia following ingestion of native whey protein, micellar casein, and a whey-casein blend in young men. Appl. Physiol. Nutr. Metab. 2019, 44, 103–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Klebach, M.; Visser, M.; Hofman, Z. Amino Acid Availability of a Dairy and Vegetable Protein Blend Compared to Single Casein, Whey, Soy, and Pea Proteins: A Double-Blind, Cross-Over Trial. Nutrients 2019, 11, 2613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassis, A.; Godin, J.-P.; Moille, S.E.; Nielsen-Moennoz, C.; Groulx, K.; Oguey-Araymon, S.; Praplan, F.; Beaumont, M.; Sauser, J.; Monnard, I.; et al. Effects of protein quantity and type on diet induced thermogenesis in overweight adults: A randomized controlled trial. Clin. Nutr. 2019, 38, 1570–1580. [Google Scholar] [CrossRef] [PubMed]
- Trommelen, J.; Holwerda, A.M.; Kouw, I.W.K.; Langer, H.; Halson, S.L.; Rollo, I.; Verdijk, L.B.; Van Loon, L.J.C. Resistance Exercise Augments Postprandial Overnight Muscle Protein Synthesis Rates. Med. Sci. Sport. Exerc. 2016, 48, 2517–2525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wall, B.T.; Hamer, H.M.; De Lange, A.; Kiskini, A.; Groen, B.B.; Senden, J.M.; Gijsen, A.P.; Verdijk, L.B.; Van Loon, L. Leucine co-ingestion improves post-prandial muscle protein accretion in elderly men. Clin. Nutr. 2013, 32, 412–419. [Google Scholar] [CrossRef]
- Kouw, I.W.; Holwerda, A.M.; Trommelen, J.; Kramer, I.F.; Bastiaanse, J.; Halson, S.L.; Wodzig, W.K.; Verdijk, L.B.; Van Loon, L.J. Protein Ingestion before Sleep Increases Overnight Muscle Protein Synthesis Rates in Healthy Older Men: A Randomized Controlled Trial. J. Nutr. 2017, 147, 2252–2261. [Google Scholar] [CrossRef] [Green Version]
- Farup, J.; Rahbek, S.K.; Storm, A.C.; Klitgaard, S.; Jørgensen, H.; Bibby, B.M.; Serena, A.; Vissing, K. Effect of degree of hydrolysis of whey protein on in vivo plasma amino acid appearance in humans. SpringerPlus 2016, 5, 382. [Google Scholar] [CrossRef] [Green Version]
- Dangin, M.; Boirie, Y.; Garcia-Rodenas, C.; Gachon, P.; Fauquant, J.; Callier, P.; Ballèvre, O.; Beaufrere, B. The digestion rate of protein is an independent regulating factor of postprandial protein retention. Am. J. Physiol. Metab. 2001, 280, E340–E348. [Google Scholar] [CrossRef] [Green Version]
- Sikand, V.; Tong, P.; Roy, S.; Rodriguez-Saona, L.; Murray, B. Solubility of commercial milk protein concentrates and milk protein isolates. J. Dairy Sci. 2011, 94, 6194–6202. [Google Scholar] [CrossRef]
- Schokker, E.P.; Church, J.S.; Mata, J.P.; Gilbert, E.; Puvanenthiran, A.; Udabage, P. Reconstitution properties of micellar casein powder: Effects of composition and storage. Int. Dairy J. 2011, 21, 877–886. [Google Scholar] [CrossRef]
- Anema, S.; Pinder, D.; Hunter, R.; Hemar, Y. Effects of storage temperature on the solubility of milk protein concentrate (MPC85). Food Hydrocoll. 2006, 20, 386–393. [Google Scholar] [CrossRef]
- Guo, C.; Campbell, B.; Chen, K.; Lenhoff, A.M.; Velev, O.D. Casein precipitation equilibria in the presence of calcium ions and phosphates. Colloids Surf. B Biointerfaces 2003, 29, 297–307. [Google Scholar] [CrossRef]
- Nyakayiru, J.; Van Lieshout, G.A.A.; Trommelen, J.; Van Kranenburg, J.; Verdijk, L.B.; Bragt, M.C.E.; Van Loon, L.J.C. The glycation level of milk protein strongly modulates post-prandial lysine availability in humans. Br. J. Nutr. 2019, 123, 545–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Lieshout, G.A.A.; Lambers, T.T.; Bragt, M.C.E.; Hettinga, K. How processing may affect milk protein digestion and overall physiological outcomes: A systematic review. Crit. Rev. Food Sci. Nutr. 2019, 1–24. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasma Metabolites | Micellar Casein | Calcium Caseinate | Cross-Linked Sodium Caseinate | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Glucose | |||||||||
| Cmax (mmol·L−1) | 5.4 | ± | 0.5 a | 5.5 | ± | 0.8 a | 5.5 | ± | 0.5 a |
| Tmax (min) | 23 | ± | 31 a | 61 | ± | 64 a | 28 | ± | 38 a |
| iAUC (mmol·L−1·6 h−1) | −92 | ± | 73 a | −67 | ± | 196 a | −89 | ± | 80 a |
| Insulin | |||||||||
| Cmax (mU·L−1) | 31.3 | ± | 15.7 a | 30.2 | ± | 16.2 a | 34.6 | ± | 13.5 a |
| Tmax (min) | 28 | ± | 8 a | 28 | ± | 34 a | 24 | ± | 8 a |
| iAUC (mU·L−1·6 h−1) | 321 | ± | 660 a | 419 | ± | 653 a | 619 | ± | 658 a |
| Branched-chain amino acids | |||||||||
| Cmax (µmol·L−1) | 1074 | ± | 148 a | 949 | ± | 156 a | 1351 | ± | 178 b |
| Tmax (min) | 72 | ± | 58 a | 76 | ± | 74 a | 57 | ± | 6 a |
| iAUC (mmol·L−1·6 h−1) | 113 | ± | 23 a | 114 | ± | 26 a | 132 | ± | 21 b |
| Essential amino acids | |||||||||
| Cmax (µmol·L−1) | 2028 | ± | 301 c | 1775 | ± | 226 a | 2394 | ± | 275 b |
| Tmax (min) | 47 | ± | 38 a | 71 | ± | 76 a | 56 | ± | 7 a |
| iAUC (mmol·L−1·6 h−1) | 176 | ± | 38 a | 174 | ± | 43 a | 190 | ± | 36 b |
| Non-essential | |||||||||
| Cmax (µmol·L−1) | 1779 | ± | 316 a | 1623 | ± | 203 a | 2063 | ± | 287 b |
| Tmax (min) | 48 | ± | 38 a | 74 | ± | 76 a | 57 | ± | 6 a |
| iAUC (mmol·L−1·6 h−1) | 94 | ± | 28 a,b | 86 | ± | 36 a | 104 | ± | 31 b |
| Total amino acids | |||||||||
| Cmax (µmol·L−1) | 3804 | ± | 604 a | 3394 | ± | 398 a | 4454 | ± | 518 b |
| Tmax (min) | 48 | ± | 38 a | 69 | ± | 75 a | 56 | ± | 7 a |
| iAUC (mmol·L−1·6 h−1) | 270 | ± | 63 a,b | 260 | ± | 75 a | 294 | ± | 63 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trommelen, J.; Weijzen, M.E.G.; van Kranenburg, J.; Ganzevles, R.A.; Beelen, M.; Verdijk, L.B.; van Loon, L.J.C. Casein Protein Processing Strongly Modulates Post-Prandial Plasma Amino Acid Responses In Vivo in Humans. Nutrients 2020, 12, 2299. https://doi.org/10.3390/nu12082299
Trommelen J, Weijzen MEG, van Kranenburg J, Ganzevles RA, Beelen M, Verdijk LB, van Loon LJC. Casein Protein Processing Strongly Modulates Post-Prandial Plasma Amino Acid Responses In Vivo in Humans. Nutrients. 2020; 12(8):2299. https://doi.org/10.3390/nu12082299
Chicago/Turabian StyleTrommelen, Jorn, Michelle E. G. Weijzen, Janneau van Kranenburg, Renate A. Ganzevles, Milou Beelen, Lex B. Verdijk, and Luc J. C. van Loon. 2020. "Casein Protein Processing Strongly Modulates Post-Prandial Plasma Amino Acid Responses In Vivo in Humans" Nutrients 12, no. 8: 2299. https://doi.org/10.3390/nu12082299
APA StyleTrommelen, J., Weijzen, M. E. G., van Kranenburg, J., Ganzevles, R. A., Beelen, M., Verdijk, L. B., & van Loon, L. J. C. (2020). Casein Protein Processing Strongly Modulates Post-Prandial Plasma Amino Acid Responses In Vivo in Humans. Nutrients, 12(8), 2299. https://doi.org/10.3390/nu12082299