Myostatin as a Biomarker of Muscle Wasting and other Pathologies-State of the Art and Knowledge Gaps


{kind=link}
Abstract
:1. Introduction
2. Myostatin and Muscle Wasting
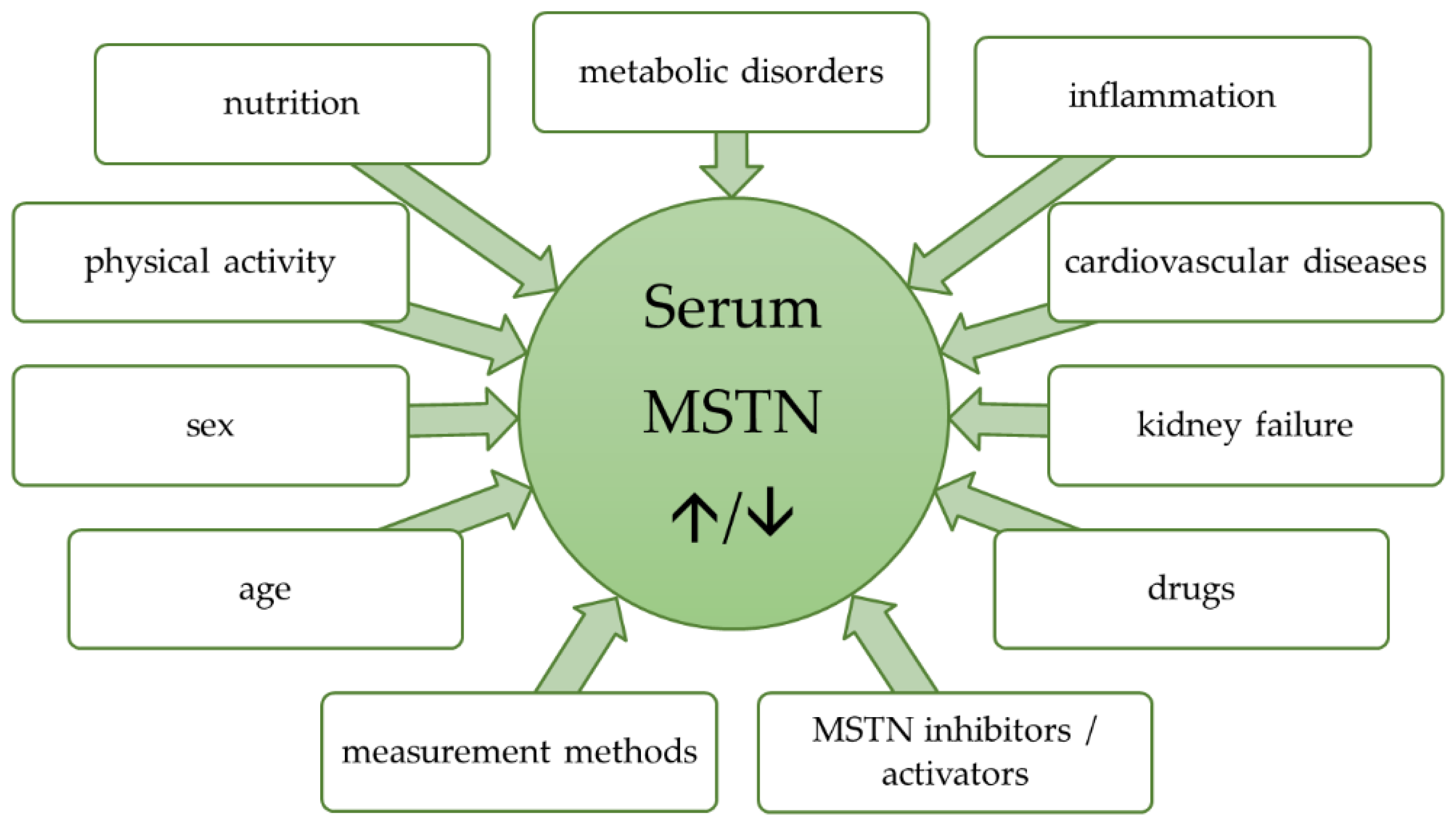
3. Factors Affecting Myostatin Concentration
3.1. Age and Sex Differences
3.2. Physical Activity
3.3. Nutritional and Metabolic Status
3.4. Inflammation and Injuries
3.5. Heart
3.6. Kidneys
3.7. Gynecological Disorders
3.8. Pharmaceutics
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ActRIIA | type IIA activin receptorv |
| ActRIIB | activin type IIB receptor |
| ALK4/5 | activin receptor-like kinase 4 or 5 |
| AMPK | 5’AMP-activated protein kinase |
| AngII | angiotensin II |
| AT1R | angiotensin II type I receptor |
| BHD | bicarbonate hemodialysis |
| BMP1 | bone morphogenic protein 1 |
| BMP-11 | bone morphogenetic protein 11 |
| COPD | chronic obstructive pulmonary disease |
| CRP | C-reactive protein |
| DF-3 | follistatin-derived myostatin inhibitory peptide |
| EHC | euglycemic-hyperinsulinemic clamp |
| ELISA | enzyme-linked immunosorbent assay |
| EWGSOP | European Working Group in Sarcopenia in Older People |
| FLRG | follistatin–related gene |
| FNDC5 | fibronectin type III domain-containing protein 5 |
| FoxO | forkhead box protein O |
| GASP-1 | growth and serum protein-1 |
| GDF-8 | growth differentiation factor -8 |
| GDF-11 | growth differentiation factor 11 |
| HD | hemodialysis |
| HDF | hemodiafiltration |
| HF | heart failure |
| HOMA-IR | Homeostatic Model Assessment for Insulin Resistance |
| IL-6 | interleukin 6 |
| IL-15 | interleukin 15 |
| IGF-1 | insulin-like growth factor |
| MEF2 | myocyte-specific enhancer factor |
| miRNA | micro RNA |
| MRF4 | myogenic regulatory factor 4 |
| mTOR | mammalian target of rapamycin |
| mTORC1 | mammalian target of rapamycin complex 1 |
| MYF5 | myogenic factor 5 |
| MYF6 | myogenic factor 6 |
| MyoD | myoblast determination protein 1 |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NT-proBNP | N-terminal pro B-type natriuretic peptide |
| PPARγ | peroxisome proliferator-activated receptor gamma |
| p38MAPK | p 38 mitogen-activated kinases |
| RVMPI | right ventricular myocardial performance index |
| TAK, | 1Transforming growth factor beta-activated kinase 1 |
| TAPSE | tricuspid annular plane systolic excursion |
| TGF-β | transforming growth factor β |
| TNF-α | tumor necrosis factor -α |
References
- Cruz-Jentoft, A.J.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyere, O.; Cederholm, T.; Cooper, C.; Landi, F.; Rolland, Y.; Sayer, A.A.; et al. Sarcopenia: Revised European consensus on definition and diagnosis. Age Ageing 2019, 48, 601. [Google Scholar] [CrossRef] [Green Version]
- Mayhew, A.J.; Amog, K.; Phillips, S.; Parise, G.; McNicholas, P.D.; de Souza, R.J.; Thabane, L.; Raina, P. The prevalence of sarcopenia in community-dwelling older adults, an exploration of differences between studies and within definitions: A systematic review and meta-analyses. Age Ageing 2019, 48, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Uemura, K.; Doi, T.; Lee, S.; Shimada, H. Sarcopenia and Low Serum Albumin Level Synergistically Increase the Risk of Incident Disability in Older Adults. J. Am. Med. Dir. Assoc. 2019, 20, 90–93. [Google Scholar] [CrossRef]
- Beaudart, C.; Reginster, J.Y.; Petermans, J.; Gillain, S.; Quabron, A.; Locquet, M.; Slomian, J.; Buckinx, F.; Bruyere, O. Quality of life and physical components linked to sarcopenia: The SarcoPhAge study. Exp. Gerontol. 2015, 69, 103–110. [Google Scholar] [CrossRef]
- Arango-Lopera, V.E.; Arroyo, P.; Gutierrez-Robledo, L.M.; Perez-Zepeda, M.U.; Cesari, M. Mortality as an adverse outcome of sarcopenia. J. Nutr. Health Aging 2013, 17, 259–262. [Google Scholar] [CrossRef]
- Beaudart, C.; Zaaria, M.; Pasleau, F.; Reginster, J.Y.; Bruyere, O. Health Outcomes of Sarcopenia: A Systematic Review and Meta-Analysis. PLoS ONE 2017, 12, e0169548. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Jentoft, A.J.; Baeyens, J.P.; Bauer, J.M.; Boirie, Y.; Cederholm, T.; Landi, F.; Martin, F.C.; Michel, J.P.; Rolland, Y.; Schneider, S.M.; et al. Sarcopenia: European consensus on definition and diagnosis: Report of the European Working Group on Sarcopenia in Older People. Age Ageing 2010, 39, 412–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fielding, R.A.; Vellas, B.; Evans, W.J.; Bhasin, S.; Morley, J.E.; Newman, A.B.; Abellan van Kan, G.; Andrieu, S.; Bauer, J.; Breuille, D.; et al. Sarcopenia: An undiagnosed condition in older adults. Current consensus definition: Prevalence, etiology, and consequences. International working group on sarcopenia. J. Am. Med. Dir. Assoc. 2011, 12, 249–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welle, S.; Brooks, A.I.; Delehanty, J.M.; Needler, N.; Thornton, C.A. Gene expression profile of aging in human muscle. Physiol. Genom. 2003, 14, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Schrager, M.A.; Roth, S.M.; Ferrell, R.E.; Metter, E.J.; Russek-Cohen, E.; Lynch, N.A.; Lindle, R.S.; Hurley, B.F. Insulin-like growth factor-2 genotype, fat-free mass, and muscle performance across the adult life span. J. Appl. Physiol. (1985) 2004, 97, 2176–2183. [Google Scholar] [CrossRef]
- Phillips, T.; Leeuwenburgh, C. Muscle fiber specific apoptosis and TNF-alpha signaling in sarcopenia are attenuated by life-long calorie restriction. FASEB J. 2005, 19, 668–670. [Google Scholar] [CrossRef] [PubMed]
- Rong, Y.D.; Bian, A.L.; Hu, H.Y.; Ma, Y.; Zhou, X.Z. Study on relationship between elderly sarcopenia and inflammatory cytokine IL-6, anti-inflammatory cytokine IL-10. BMC Geriatr. 2018, 18, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrucci, L.; Penninx, B.W.; Volpato, S.; Harris, T.B.; Bandeen-Roche, K.; Balfour, J.; Leveille, S.G.; Fried, L.P.; Md, J.M. Change in muscle strength explains accelerated decline of physical function in older women with high interleukin-6 serum levels. J. Am. Geriatr. Soc. 2002, 50, 1947–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visser, M.; Deeg, D.J.; Lips, P.; Longitudinal Aging Study Amsterdam. Low vitamin D and high parathyroid hormone levels as determinants of loss of muscle strength and muscle mass (sarcopenia): The Longitudinal Aging Study Amsterdam. J. Clin. Endocrinol. Metab. 2003, 88, 5766–5772. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, R.N.; Waters, D.L.; Gallagher, D.; Morley, J.E.; Garry, P.J. Predictors of skeletal muscle mass in elderly men and women. Mech. Ageing Dev. 1999, 107, 123–136. [Google Scholar] [CrossRef]
- Schaap, L.A.; Pluijm, S.M.; Smit, J.H.; van Schoor, N.M.; Visser, M.; Gooren, L.J.; Lips, P. The association of sex hormone levels with poor mobility, low muscle strength and incidence of falls among older men and women. Clin. Endocrinol. (Oxf.) 2005, 63, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Bian, A.; Ma, Y.; Zhou, X.; Guo, Y.; Wang, W.; Zhang, Y.; Wang, X. Association between sarcopenia and levels of growth hormone and insulin-like growth factor-1 in the elderly. BMC Musculoskelet. Disord. 2020, 21, 214. [Google Scholar] [CrossRef] [Green Version]
- Hiona, A.; Sanz, A.; Kujoth, G.C.; Pamplona, R.; Seo, A.Y.; Hofer, T.; Someya, S.; Miyakawa, T.; Nakayama, C.; Samhan-Arias, A.K.; et al. Mitochondrial DNA mutations induce mitochondrial dysfunction, apoptosis and sarcopenia in skeletal muscle of mitochondrial DNA mutator mice. PLoS ONE 2010, 5, e11468. [Google Scholar] [CrossRef]
- Chabi, B.; Ljubicic, V.; Menzies, K.J.; Huang, J.H.; Saleem, A.; Hood, D.A. Mitochondrial function and apoptotic susceptibility in aging skeletal muscle. Aging Cell 2008, 7, 2–12. [Google Scholar] [CrossRef]
- Whitman, S.A.; Wacker, M.J.; Richmond, S.R.; Godard, M.P. Contributions of the ubiquitin-proteasome pathway and apoptosis to human skeletal muscle wasting with age. Pflugers Arch. 2005, 450, 437–446. [Google Scholar] [CrossRef]
- Kuh, D.; Bassey, E.J.; Butterworth, S.; Hardy, R.; Wadsworth, M.E.; Musculoskeletal Study Team. Grip strength, postural control, and functional leg power in a representative cohort of British men and women: Associations with physical activity, health status, and socioeconomic conditions. J. Gerontol. A Biol. Sci. Med. Sci. 2005, 60, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Kortebein, P.; Symons, T.B.; Ferrando, A.; Paddon-Jones, D.; Ronsen, O.; Protas, E.; Conger, S.; Lombeida, J.; Wolfe, R.; Evans, W.J. Functional impact of 10 days of bed rest in healthy older adults. J. Gerontol. A Biol. Sci. Med. Sci. 2008, 63, 1076–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hai, S.; Cao, L.; Wang, H.; Zhou, J.; Liu, P.; Yang, Y.; Hao, Q.; Dong, B. Association between sarcopenia and nutritional status and physical activity among community-dwelling Chinese adults aged 60 years and older. Geriatr. Gerontol. Int. 2017, 17, 1959–1966. [Google Scholar] [CrossRef] [PubMed]
- Bahat, G.; Saka, B.; Tufan, F.; Akin, S.; Sivrikaya, S.; Yucel, N.; Erten, N.; Karan, M.A. Prevalence of sarcopenia and its association with functional and nutritional status among male residents in a nursing home in Turkey. Aging Male 2010, 13, 211–214. [Google Scholar] [CrossRef] [PubMed]
- McDermott, M.M.; Greenland, P.; Liu, K.; Guralnik, J.M.; Criqui, M.H.; Dolan, N.C.; Chan, C.; Celic, L.; Pearce, W.H.; Schneider, J.R.; et al. Leg symptoms in peripheral arterial disease: Associated clinical characteristics and functional impairment. JAMA 2001, 286, 1599–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDermott, M.M.; Guralnik, J.M.; Albay, M.; Bandinelli, S.; Miniati, B.; Ferrucci, L. Impairments of muscles and nerves associated with peripheral arterial disease and their relationship with lower extremity functioning: The InCHIANTI Study. J. Am. Geriatr. Soc. 2004, 52, 405–410. [Google Scholar] [CrossRef]
- Park, S.W.; Goodpaster, B.H.; Strotmeyer, E.S.; de Rekeneire, N.; Harris, T.B.; Schwartz, A.V.; Tylavsky, F.A.; Newman, A.B. Decreased muscle strength and quality in older adults with type 2 diabetes: The health, aging, and body composition study. Diabetes 2006, 55, 1813–1818. [Google Scholar] [CrossRef] [Green Version]
- Park, S.W.; Goodpaster, B.H.; Lee, J.S.; Kuller, L.H.; Boudreau, R.; de Rekeneire, N.; Harris, T.B.; Kritchevsky, S.; Tylavsky, F.A.; Nevitt, M.; et al. Excessive loss of skeletal muscle mass in older adults with type 2 diabetes. Diabetes Care 2009, 32, 1993–1997. [Google Scholar] [CrossRef] [Green Version]
- Riuzzi, F.; Sorci, G.; Arcuri, C.; Giambanco, I.; Bellezza, I.; Minelli, A.; Donato, R. Cellular and molecular mechanisms of sarcopenia: The S100B perspective. J. Cachexia Sarcopenia Muscle 2018, 9, 1255–1268. [Google Scholar] [CrossRef] [Green Version]
- Morley, J.E. Treatment of sarcopenia: The road to the future. J. Cachexia Sarcopenia Muscle 2018, 9, 1196–1199. [Google Scholar] [CrossRef]
- Sousa-Santos, A.R.; Afonso, C.; Borges, N.; Santos, A.; Padrao, P.; Moreira, P.; Amaral, T.F. Sarcopenia, physical frailty, undernutrition and obesity cooccurrence among Portuguese community-dwelling older adults: Results from Nutrition UP 65 cross-sectional study. BMJ Open 2020, 10, e033661. [Google Scholar] [CrossRef] [PubMed]
- Landi, F.; Calvani, R.; Cesari, M.; Tosato, M.; Martone, A.M.; Bernabei, R.; Onder, G.; Marzetti, E. Sarcopenia as the Biological Substrate of Physical Frailty. Clin. Geriatr. Med. 2015, 31, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Yaxley, A.; Miller, M.D.; Fraser, R.J.; Cobiac, L.; Crotty, M. The complexity of treating wasting in ambulatory rehabilitation: Is it starvation, sarcopenia, cachexia or a combination of these conditions? Asia Pac. J. Clin. Nutr. 2012, 21, 386–393. [Google Scholar]
- Gingrich, A.; Volkert, D.; Kiesswetter, E.; Thomanek, M.; Bach, S.; Sieber, C.C.; Zopf, Y. Prevalence and overlap of sarcopenia, frailty, cachexia and malnutrition in older medical inpatients. BMC Geriatr. 2019, 19, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narumi, T.; Watanabe, T.; Kadowaki, S.; Takahashi, T.; Yokoyama, M.; Kinoshita, D.; Honda, Y.; Funayama, A.; Nishiyama, S.; Takahashi, H.; et al. Sarcopenia evaluated by fat-free mass index is an important prognostic factor in patients with chronic heart failure. Eur. J. Intern. Med. 2015, 26, 118–122. [Google Scholar] [CrossRef]
- Hu, G.F.; Tian, J.H.; Yang, J.; Fu, H.X.; Liu, J. Sarcopenia and long-term prognosis of patients with chronic heart failure in patients aged 70 years and over. Zhonghua Nei Ke Za Zhi 2018, 57, 212–215. [Google Scholar] [CrossRef]
- Pereira, R.A.; Cordeiro, A.C.; Avesani, C.M.; Carrero, J.J.; Lindholm, B.; Amparo, F.C.; Amodeo, C.; Cuppari, L.; Kamimura, M.A. Sarcopenia in chronic kidney disease on conservative therapy: Prevalence and association with mortality. Nephrol. Dial. Transplant. 2015, 30, 1718–1725. [Google Scholar] [CrossRef] [Green Version]
- Mauro, E.; Crespo, G.; Martinez-Garmendia, A.; Gutierrez-Acevedo, M.N.; Diaz, J.M.; Saidman, J.; Bermudez, C.; Ortiz-Patron, J.; Garcia-Olveira, L.; Zalazar, F.; et al. Cystatin C and Sarcopenia Predict Acute on Chronic Liver Failure Development and Mortality in Patients on the Liver Transplant Waiting List. Transplantation 2020, 104, e188–e198. [Google Scholar] [CrossRef]
- Scherbakov, N.; Doehner, W. Cachexia as a common characteristic in multiple chronic disease. J. Cachexia Sarcopenia Muscle 2018, 9, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Bowen, T.S.; Schuler, G.; Adams, V. Skeletal muscle wasting in cachexia and sarcopenia: Molecular pathophysiology and impact of exercise training. J. Cachexia Sarcopenia Muscle 2015, 6, 197–207. [Google Scholar] [CrossRef]
- Argiles, J.M.; Busquets, S.; Felipe, A.; Lopez-Soriano, F.J. Muscle wasting in cancer and ageing: Cachexia versus sarcopenia. Adv. Gerontol. 2006, 18, 39–54. [Google Scholar] [PubMed]
- Douglas, E.; McMillan, D.C. Towards a simple objective framework for the investigation and treatment of cancer cachexia: The Glasgow Prognostic Score. Cancer Treat. Rev. 2014, 40, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Silva, G.A.D.; Wiegert, E.V.M.; Calixto-Lima, L.; Oliveira, L.C. Clinical utility of the modified Glasgow Prognostic Score to classify cachexia in patients with advanced cancer in palliative care. Clin. Nutr. 2020, 39, 1587–1592. [Google Scholar] [CrossRef] [PubMed]
- Califf, R.M. Biomarker definitions and their applications. Exp. Biol. Med. (Maywood) 2018, 243, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Calvani, R.; Marini, F.; Cesari, M.; Tosato, M.; Anker, S.D.; von Haehling, S.; Miller, R.R.; Bernabei, R.; Landi, F.; Marzetti, E.; et al. Biomarkers for physical frailty and sarcopenia: State of the science and future developments. J. Cachexia Sarcopenia Muscle 2015, 6, 278–286. [Google Scholar] [CrossRef] [PubMed]
- McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member. Nature 1997, 387, 83–90. [Google Scholar] [CrossRef]
- Kambadur, R.; Sharma, M.; Smith, T.P.; Bass, J.J. Mutations in myostatin (GDF8) in double-muscled Belgian Blue and Piedmontese cattle. Genome Res. 1997, 7, 910–916. [Google Scholar] [CrossRef] [Green Version]
- Clop, A.; Marcq, F.; Takeda, H.; Pirottin, D.; Tordoir, X.; Bibe, B.; Bouix, J.; Caiment, F.; Elsen, J.M.; Eychenne, F.; et al. A mutation creating a potential illegitimate microRNA target site in the myostatin gene affects muscularity in sheep. Nat. Genet. 2006, 38, 813–818. [Google Scholar] [CrossRef]
- Mosher, D.S.; Quignon, P.; Bustamante, C.D.; Sutter, N.B.; Mellersh, C.S.; Parker, H.G.; Ostrander, E.A. A mutation in the myostatin gene increases muscle mass and enhances racing performance in heterozygote dogs. PLoS Genet. 2007, 3, e79. [Google Scholar] [CrossRef]
- Schuelke, M.; Wagner, K.R.; Stolz, L.E.; Hubner, C.; Riebel, T.; Komen, W.; Braun, T.; Tobin, J.F.; Lee, S.J. Myostatin mutation associated with gross muscle hypertrophy in a child. N. Engl. J. Med. 2004, 350, 2682–2688. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Kambadur, R.; Matthews, K.G.; Somers, W.G.; Devlin, G.P.; Conaglen, J.V.; Fowke, P.J.; Bass, J.J. Myostatin, a transforming growth factor-beta superfamily member, is expressed in heart muscle and is upregulated in cardiomyocytes after infarct. J. Cell Physiol. 1999, 180, 1–9. [Google Scholar] [CrossRef]
- McPherron, A.C.; Lee, S.J. Suppression of body fat accumulation in myostatin-deficient mice. J. Clin. Investig. 2002, 109, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; McPherron, A.C. Regulation of myostatin activity and muscle growth. Proc. Natl. Acad. Sci. USA 2001, 98, 9306–9311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thies, R.S.; Chen, T.; Davies, M.V.; Tomkinson, K.N.; Pearson, A.A.; Shakey, Q.A.; Wolfman, N.M. GDF-8 propeptide binds to GDF-8 and antagonizes biological activity by inhibiting GDF-8 receptor binding. Growth Factors 2001, 18, 251–259. [Google Scholar] [CrossRef]
- Wolfman, N.M.; McPherron, A.C.; Pappano, W.N.; Davies, M.V.; Song, K.; Tomkinson, K.N.; Wright, J.F.; Zhao, L.; Sebald, S.M.; Greenspan, D.S.; et al. Activation of latent myostatin by the BMP-1/tolloid family of metalloproteinases. Proc. Natl. Acad. Sci. USA 2003, 100, 15842–15846. [Google Scholar] [CrossRef] [Green Version]
- Langley, B.; Thomas, M.; Bishop, A.; Sharma, M.; Gilmour, S.; Kambadur, R. Myostatin inhibits myoblast differentiation by down-regulating MyoD expression. J. Biol. Chem. 2002, 277, 49831–49840. [Google Scholar] [CrossRef] [Green Version]
- Rebbapragada, A.; Benchabane, H.; Wrana, J.L.; Celeste, A.J.; Attisano, L. Myostatin signals through a transforming growth factor beta-like signaling pathway to block adipogenesis. Mol. Cell Biol. 2003, 23, 7230–7242. [Google Scholar] [CrossRef] [Green Version]
- Hennebry, A.; Berry, C.; Siriett, V.; O’Callaghan, P.; Chau, L.; Watson, T.; Sharma, M.; Kambadur, R. Myostatin regulates fiber-type composition of skeletal muscle by regulating MEF2 and MyoD gene expression. Am. J. Physiol. Cell Physiol. 2009, 296, C525–C534. [Google Scholar] [CrossRef]
- Amirouche, A.; Durieux, A.C.; Banzet, S.; Koulmann, N.; Bonnefoy, R.; Mouret, C.; Bigard, X.; Peinnequin, A.; Freyssenet, D. Down-regulation of Akt/mammalian target of rapamycin signaling pathway in response to myostatin overexpression in skeletal muscle. Endocrinology 2009, 150, 286–294. [Google Scholar] [CrossRef] [Green Version]
- Bonala, S.; Lokireddy, S.; McFarlane, C.; Patnam, S.; Sharma, M.; Kambadur, R. Myostatin induces insulin resistance via Casitas B-lineage lymphoma b (Cblb)-mediated degradation of insulin receptor substrate 1 (IRS1) protein in response to high calorie diet intake. J. Biol. Chem. 2014, 289, 7654–7670. [Google Scholar] [CrossRef] [Green Version]
- Bonala, S.; McFarlane, C.; Ang, J.; Lim, R.; Lee, M.; Chua, H.; Lokireddy, S.; Sreekanth, P.; Leow, M.K.; Meng, K.C.; et al. Pid1 induces insulin resistance in both human and mouse skeletal muscle during obesity. Mol. Endocrinol. 2013, 27, 1518–1535. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Mallidis, C.; Artaza, J.; Taylor, W.; Gonzalez-Cadavid, N.; Bhasin, S. Characterization of 5’-regulatory region of human myostatin gene: Regulation by dexamethasone in vitro. Am. J. Physiol. Endocrinol. Metab. 2001, 281, E1128–E1136. [Google Scholar] [CrossRef] [PubMed]
- Valdes, J.A.; Flores, S.; Fuentes, E.N.; Osorio-Fuentealba, C.; Jaimovich, E.; Molina, A. IGF-1 induces IP3 -dependent calcium signal involved in the regulation of myostatin gene expression mediated by NFAT during myoblast differentiation. J. Cell Physiol. 2013, 228, 1452–1463. [Google Scholar] [CrossRef] [PubMed]
- Lenk, K.; Schur, R.; Linke, A.; Erbs, S.; Matsumoto, Y.; Adams, V.; Schuler, G. Impact of exercise training on myostatin expression in the myocardium and skeletal muscle in a chronic heart failure model. Eur. J. Heart Fail. 2009, 11, 342–348. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.W.; Chang, H.; Kuan, P.; Shyu, K.G. Angiotensin II activates myostatin expression in cultured rat neonatal cardiomyocytes via p38 MAP kinase and myocyte enhance factor 2 pathway. J. Endocrinol. 2008, 197, 85–93. [Google Scholar] [CrossRef]
- Carneiro, I.; Castro-Piedras, I.; Munoz, A.; Labandeira-Garcia, J.L.; Devesa, J.; Arce, V.M. Hypothyroidism is associated with increased myostatin expression in rats. J. Endocrinol. Investig. 2008, 31, 773–778. [Google Scholar] [CrossRef]
- Qiu, J.; Thapaliya, S.; Runkana, A.; Yang, Y.; Tsien, C.; Mohan, M.L.; Narayanan, A.; Eghtesad, B.; Mozdziak, P.E.; McDonald, C.; et al. Hyperammonemia in cirrhosis induces transcriptional regulation of myostatin by an NF-kappaB-mediated mechanism. Proc. Natl. Acad. Sci. USA 2013, 110, 18162–18167. [Google Scholar] [CrossRef] [Green Version]
- Drummond, M.J.; Glynn, E.L.; Fry, C.S.; Dhanani, S.; Volpi, E.; Rasmussen, B.B. Essential amino acids increase microRNA-499, -208b, and -23a and downregulate myostatin and myocyte enhancer factor 2C mRNA expression in human skeletal muscle. J. Nutr. 2009, 139, 2279–2284. [Google Scholar] [CrossRef]
- Kim, J.S.; Petrella, J.K.; Cross, J.M.; Bamman, M.M. Load-mediated downregulation of myostatin mRNA is not sufficient to promote myofiber hypertrophy in humans: A cluster analysis. J. Appl. Physiol. (1985) 2007, 103, 1488–1495. [Google Scholar] [CrossRef]
- Li, L.; Wang, J.; Bai, Y.; Li, J.; Yu, X.; Luo, X.; Zhu, Z.; He, X.; Dong, Y.; Li, H.; et al. Effect of Hypoxia on the Muscle Fiber Switching Signal Pathways CnA/NFATc1 and Myostatin in Mouse Myocytes. Acta Histochem. 2019, 121, 539–545. [Google Scholar] [CrossRef]
- Forbes, D.; Jackman, M.; Bishop, A.; Thomas, M.; Kambadur, R.; Sharma, M. Myostatin auto-regulates its expression by feedback loop through Smad7 dependent mechanism. J. Cell. Physiol. 2006, 206, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Callis, T.E.; Pandya, K.; Seok, H.Y.; Tang, R.H.; Tatsuguchi, M.; Huang, Z.P.; Chen, J.F.; Deng, Z.; Gunn, B.; Shumate, J.; et al. MicroRNA-208a is a regulator of cardiac hypertrophy and conduction in mice. J. Clin. Investig. 2009, 119, 2772–2786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Williams, A.H.; Kim, Y.; McAnally, J.; Bezprozvannaya, S.; Sutherland, L.B.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. An intragenic MEF2-dependent enhancer directs muscle-specific expression of microRNAs 1 and 133. Proc. Natl. Acad. Sci. USA 2007, 104, 20844–20849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, L.; Caldow, M.K.; Watts, R.; Levinger, P.; Cameron-Smith, D.; Levinger, I. Age and sex differences in human skeletal muscle fibrosis markers and transforming growth factor-beta signaling. Eur. J. Appl. Physiol. 2017, 117, 1463–1472. [Google Scholar] [CrossRef]
- Welle, S.; Bhatt, K.; Shah, B.; Thornton, C. Insulin-like growth factor-1 and myostatin mRNA expression in muscle: Comparison between 62–77 and 21–31 yr old men. Exp. Gerontol. 2002, 37, 833–839. [Google Scholar] [CrossRef]
- Elliott, B.; Renshaw, D.; Getting, S.; Mackenzie, R. The central role of myostatin in skeletal muscle and whole body homeostasis. Acta Physiol. (Oxf.) 2012, 205, 324–340. [Google Scholar] [CrossRef]
- Huang, Z.; Chen, X.; Chen, D. Myostatin: A novel insight into its role in metabolism, signal pathways, and expression regulation. Cell. Signal. 2011, 23, 1441–1446. [Google Scholar] [CrossRef]
- Dschietzig, T.B. Myostatin—From the Mighty Mouse to cardiovascular disease and cachexia. Clin. Chim. Acta 2014, 433, 216–224. [Google Scholar] [CrossRef]
- Yamada, S.; Tsuruya, K.; Yoshida, H.; Tokumoto, M.; Ueki, K.; Ooboshi, H.; Kitazono, T. Factors Associated with the Serum Myostatin Level in Patients Undergoing Peritoneal Dialysis: Potential Effects of Skeletal Muscle Mass and Vitamin D Receptor Activator Use. Calcif. Tissue Int. 2016, 99, 13–22. [Google Scholar] [CrossRef]
- Zhou, Y.; Hellberg, M.; Hellmark, T.; Hoglund, P.; Clyne, N. Muscle mass and plasma myostatin after exercise training: A substudy of Renal Exercise (RENEXC)—A randomized controlled trial. Nephrol. Dial. Transplant. 2019. [Google Scholar] [CrossRef] [Green Version]
- Loumaye, A.; de Barsy, M.; Nachit, M.; Lause, P.; Frateur, L.; van Maanen, A.; Trefois, P.; Gruson, D.; Thissen, J.P. Role of Activin A and myostatin in human cancer cachexia. J. Clin. Endocrinol. Metab. 2015, 100, 2030–2038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Planella-Farrugia, C.; Comas, F.; Sabater-Masdeu, M.; Moreno, M.; Moreno-Navarrete, J.M.; Rovira, O.; Ricart, W.; Fernandez-Real, J.M. Circulating Irisin and Myostatin as Markers of Muscle Strength and Physical Condition in Elderly Subjects. Front. Physiol. 2019, 10, 871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furihata, T.; Kinugawa, S.; Fukushima, A.; Takada, S.; Homma, T.; Masaki, Y.; Abe, T.; Yokota, T.; Oba, K.; Okita, K.; et al. Serum myostatin levels are independently associated with skeletal muscle wasting in patients with heart failure. Int. J. Cardiol. 2016, 220, 483–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delanaye, P.; Bataille, S.; Quinonez, K.; Buckinx, F.; Warling, X.; Krzesinski, J.M.; Pottel, H.; Burtey, S.; Bruyere, O.; Cavalier, E. Myostatin and Insulin-Like Growth Factor 1 Are Biomarkers of Muscle Strength, Muscle Mass, and Mortality in Patients on Hemodialysis. J. Ren. Nutr. 2019, 29, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Bergen, H.R., 3rd; Farr, J.N.; Vanderboom, P.M.; Atkinson, E.J.; White, T.A.; Singh, R.J.; Khosla, S.; LeBrasseur, N.K. Myostatin as a mediator of sarcopenia versus homeostatic regulator of muscle mass: Insights using a new mass spectrometry-based assay. Skelet Muscle 2015, 5, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, L.N.; Lee, W.J.; Liu, L.K.; Lin, M.H.; Chen, L.K. Healthy community-living older men differ from women in associations between myostatin levels and skeletal muscle mass. J. Cachexia Sarcopenia Muscle 2018, 9, 635–642. [Google Scholar] [CrossRef]
- Skladany, L.; Koller, T.; Molcan, P.; Vnencakova, J.; Zilincan, M.; Jancekova, D.; Kukla, M. Prognostic usefulness of serum myostatin in advanced chronic liver disease: Its relation to gender and correlation with inflammatory status. J. Physiol. Pharmacol. 2019, 70. [Google Scholar] [CrossRef]
- Moriwaki, K.; Matsumoto, H.; Tanishima, S.; Tanimura, C.; Osaki, M.; Nagashima, H.; Hagino, H. Association of serum bone- and muscle-derived factors with age, sex, body composition, and physical function in community-dwelling middle-aged and elderly adults: A cross-sectional study. BMC Musculoskelet. Disord. 2019, 20, 276. [Google Scholar] [CrossRef]
- Hofmann, M.; Halper, B.; Oesen, S.; Franzke, B.; Stuparits, P.; Tschan, H.; Bachl, N.; Strasser, E.M.; Quittan, M.; Ploder, M.; et al. Serum concentrations of insulin-like growth factor-1, members of the TGF-beta superfamily and follistatin do not reflect different stages of dynapenia and sarcopenia in elderly women. Exp. Gerontol. 2015, 64, 35–45. [Google Scholar] [CrossRef]
- Negaresh, R.; Ranjbar, R.; Baker, J.S.; Habibi, A.; Mokhtarzade, M.; Gharibvand, M.M.; Fokin, A. Skeletal Muscle Hypertrophy, Insulin-like Growth Factor 1, Myostatin and Follistatin in Healthy and Sarcopenic Elderly Men: The Effect of Whole-body Resistance Training. Int. J. Prev. Med. 2019, 10, 29. [Google Scholar] [CrossRef]
- Koyun, D.; Nergizoglu, G.; Kir, K.M. Evaluation of the relationship between muscle mass and serum myostatin levels in chronic hemodialysis patients. Saudi J. Kidney Dis. Transpl. 2018, 29, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, L.P.; Basso-Vanelli, R.P.; Di Thommazo-Luporini, L.; Mendes, R.G.; Oliveira-Junior, M.C.; Vieira, R.P.; Bonjorno-Junior, J.C.; Oliveira, C.R.; Luporini, R.; Borghi-Silva, A. Myostatin and adipokines: The role of the metabolically unhealthy obese phenotype in muscle function and aerobic capacity in young adults. Cytokine 2018, 107, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Arrieta, H.; Hervas, G.; Rezola-Pardo, C.; Ruiz-Litago, F.; Iturburu, M.; Yanguas, J.J.; Gil, S.M.; Rodriguez-Larrad, A.; Irazusta, J. Serum Myostatin Levels Are Higher in Fitter, More Active, and Non-Frail Long-Term Nursing Home Residents and Increase after a Physical Exercise Intervention. Gerontology 2019, 65, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Barrios-Silva, L.V.; Parnell, M.; Shinwari, Z.B.; Chaudhary, G.A.; Xenofontos, T.; van Bekhoven, A.; McArthur, S.; Elliott, B.T. Activin subfamily peptides predict chronological age in humans. Physiol. Rep. 2018, 6, e13823. [Google Scholar] [CrossRef] [PubMed]
- Han, D.S.; Chang, K.V.; Li, C.M.; Lin, Y.H.; Kao, T.W.; Tsai, K.S.; Wang, T.G.; Yang, W.S. Skeletal muscle mass adjusted by height correlated better with muscular functions than that adjusted by body weight in defining sarcopenia. Sci. Rep. 2016, 6, 19457. [Google Scholar] [CrossRef]
- Tay, L.; Ding, Y.Y.; Leung, B.P.; Ismail, N.H.; Yeo, A.; Yew, S.; Tay, K.S.; Tan, C.H.; Chong, M.S. Sex-specific differences in risk factors for sarcopenia amongst community-dwelling older adults. Age (Dordr) 2015, 37, 121. [Google Scholar] [CrossRef] [Green Version]
- Roh, Y.H.; Hong, S.W.; Chung, S.W.; Lee, Y.S. Altered gene and protein expressions of vitamin D receptor in skeletal muscle in sarcopenic patients who sustained distal radius fractures. J. Bone Miner Metab. 2019, 37, 920–927. [Google Scholar] [CrossRef]
- Hedayati, M.; Nozhat, Z.; Hannani, M. Can the Serum Level of Myostatin be Considered as an Informative Factor for Cachexia Prevention in Patients with Medullary Thyroid Cancer? Asian Pac. J. Cancer Prev. 2016, 17, 119–123. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Kim, J.W.; Hwang, I.G.; Jang, J.S.; Hong, S.; Kim, T.Y.; Baek, J.Y.; Shin, S.H.; Sun, S.; Hong, D.S.; et al. Serum biomarkers for predicting overall survival and early mortality in older patients with metastatic solid tumors. J. Geriatr. Oncol. 2019, 10, 749–756. [Google Scholar] [CrossRef]
- Burch, P.M.; Pogoryelova, O.; Palandra, J.; Goldstein, R.; Bennett, D.; Fitz, L.; Guglieri, M.; Bettolo, C.M.; Straub, V.; Evangelista, T.; et al. Reduced serum myostatin concentrations associated with genetic muscle disease progression. J. Neurol. 2017, 264, 541–553. [Google Scholar] [CrossRef]
- Mariot, V.; Joubert, R.; Hourde, C.; Feasson, L.; Hanna, M.; Muntoni, F.; Maisonobe, T.; Servais, L.; Bogni, C.; Le Panse, R.; et al. Downregulation of myostatin pathway in neuromuscular diseases may explain challenges of anti-myostatin therapeutic approaches. Nat. Commun. 2017, 8, 1859. [Google Scholar] [CrossRef] [PubMed]
- Koch, C.; Buono, S.; Menuet, A.; Robe, A.; Djeddi, S.; Kretz, C.; Gomez-Oca, R.; Depla, M.; Monseur, A.; Thielemans, L.; et al. Myostatin: A Circulating Biomarker Correlating with Disease in Myotubular Myopathy Mice and Patients. Mol. Ther. Methods Clin. Dev. 2020, 17, 1178–1189. [Google Scholar] [CrossRef] [PubMed]
- Mariot, V.; Le Guiner, C.; Barthelemy, I.; Montus, M.; Blot, S.; Torelli, S.; Morgan, J.; Muntoni, F.; Voit, T.; Dumonceaux, J. Myostatin Is a Quantifiable Biomarker for Monitoring Pharmaco-gene Therapy in Duchenne Muscular Dystrophy. Mol. Ther. Methods Clin. Dev. 2020, 18, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Vernerova, L.; Horvathova, V.; Kropackova, T.; Vokurkova, M.; Klein, M.; Tomcik, M.; Oreska, S.; Spiritovic, M.; Storkanova, H.; Hermankova, B.; et al. Alterations in activin A-myostatin-follistatin system associate with disease activity in inflammatory myopathies. Rheumatology (Oxford) 2020. [Google Scholar] [CrossRef] [PubMed]
- Loffredo, F.S.; Steinhauser, M.L.; Jay, S.M.; Gannon, J.; Pancoast, J.R.; Yalamanchi, P.; Sinha, M.; Dall’Osso, C.; Khong, D.; Shadrach, J.L.; et al. Growth differentiation factor 11 is a circulating factor that reverses age-related cardiac hypertrophy. Cell 2013, 153, 828–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, M.; Jang, Y.C.; Oh, J.; Khong, D.; Wu, E.Y.; Manohar, R.; Miller, C.; Regalado, S.G.; Loffredo, F.S.; Pancoast, J.R.; et al. Restoring systemic GDF11 levels reverses age-related dysfunction in mouse skeletal muscle. Science 2014, 344, 649–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egerman, M.A.; Cadena, S.M.; Gilbert, J.A.; Meyer, A.; Nelson, H.N.; Swalley, S.E.; Mallozzi, C.; Jacobi, C.; Jennings, L.L.; Clay, I.; et al. GDF11 Increases with Age and Inhibits Skeletal Muscle Regeneration. Cell Metab. 2015, 22, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Lakshman, K.M.; Bhasin, S.; Corcoran, C.; Collins-Racie, L.A.; Tchistiakova, L.; Forlow, S.B.; St Ledger, K.; Burczynski, M.E.; Dorner, A.J.; Lavallie, E.R. Measurement of myostatin concentrations in human serum: Circulating concentrations in young and older men and effects of testosterone administration. Mol. Cell Endocrinol. 2009, 302, 26–32. [Google Scholar] [CrossRef]
- Wintgens, K.F.; Dschietzig, T.; Stoeva, S.; Paulsson, M.; Armbruster, F.P. Plasma myostatin measured by a competitive ELISA using a highly specific antiserum. Clin. Chim. Acta 2012, 413, 1288–1294. [Google Scholar] [CrossRef]
- Lehallier, B.; Gate, D.; Schaum, N.; Nanasi, T.; Lee, S.E.; Yousef, H.; Moran Losada, P.; Berdnik, D.; Keller, A.; Verghese, J.; et al. Undulating changes in human plasma proteome profiles across the lifespan. Nat. Med. 2019, 25, 1843–1850. [Google Scholar] [CrossRef]
- Breitbart, A.; Scharf, G.M.; Duncker, D.; Widera, C.; Gottlieb, J.; Vogel, A.; Schmidt, S.; Brandes, G.; Heuft, H.G.; Lichtinghagen, R.; et al. Highly specific detection of myostatin prodomain by an immunoradiometric sandwich assay in serum of healthy individuals and patients. PLoS ONE 2013, 8, e80454. [Google Scholar] [CrossRef] [PubMed]
- Cote, S.M.; Jackson, J.; Pirruccello-Straub, M.; Carven, G.J.; Wawersik, S. A Sensitive and Selective Immunoassay for the Quantitation of Serum Latent Myostatin after In Vivo Administration of SRK-015, a Selective Inhibitor of Myostatin Activation. SLAS Discov. 2020, 25, 95–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Chen, S.; Song, K.; Pan, P.; Qiu, Y.; Amaravadi, L.; Wu, J. A sensitive antibody-free 2D-LC-MS/MS assay for the quantitation of myostatin in the serum of different species. Bioanalysis 2019, 11, 957–970. [Google Scholar] [CrossRef] [PubMed]
- Camparini, L.; Kollipara, L.; Sinagra, G.; Loffredo, F.S.; Sickmann, A.; Shevchuk, O. Targeted Approach to Distinguish and Determine Absolute Levels of GDF8 and GDF11 in Mouse Serum. Proteomics 2020, 20, e1900104. [Google Scholar] [CrossRef] [PubMed]
- Gilson, H.; Schakman, O.; Kalista, S.; Lause, P.; Tsuchida, K.; Thissen, J.P. Follistatin induces muscle hypertrophy through satellite cell proliferation and inhibition of both myostatin and activin. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E157–E164. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.J.; Lee, Y.S.; Zimmers, T.A.; Soleimani, A.; Matzuk, M.M.; Tsuchida, K.; Cohn, R.D.; Barton, E.R. Regulation of muscle mass by follistatin and activins. Mol. Endocrinol. 2010, 24, 1998–2008. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Li, Y.; Lu, A.; Gharaibeh, B.; Ma, J.; Kobayashi, T.; Quintero, A.J.; Huard, J. Follistatin improves skeletal muscle healing after injury and disease through an interaction with muscle regeneration, angiogenesis, and fibrosis. Am. J. Pathol. 2011, 179, 915–930. [Google Scholar] [CrossRef]
- Saitoh, M.; Takayama, K.; Hitachi, K.; Taguchi, A.; Taniguchi, A.; Tsuchida, K.; Hayashi, Y. Discovery of a follistatin-derived myostatin inhibitory peptide. Bioorg. Med. Chem. Lett. 2020, 30, 126892. [Google Scholar] [CrossRef]
- Schafer, M.J.; Atkinson, E.J.; Vanderboom, P.M.; Kotajarvi, B.; White, T.A.; Moore, M.M.; Bruce, C.J.; Greason, K.L.; Suri, R.M.; Khosla, S.; et al. Quantification of GDF11 and Myostatin in Human Aging and Cardiovascular Disease. Cell Metab. 2016, 23, 1207–1215. [Google Scholar] [CrossRef] [Green Version]
- Chew, J.; Tay, L.; Lim, J.P.; Leung, B.P.; Yeo, A.; Yew, S.; Ding, Y.Y.; Lim, W.S. Serum Myostatin and IGF-1 as Gender-Specific Biomarkers of Frailty and Low Muscle Mass in Community-Dwelling Older Adults. J. Nutr. Health Aging 2019, 23, 979–986. [Google Scholar] [CrossRef]
- Fife, E.; Kostka, J.; Kroc, L.; Guligowska, A.; Piglowska, M.; Soltysik, B.; Kaufman-Szymczyk, A.; Fabianowska-Majewska, K.; Kostka, T. Relationship of muscle function to circulating myostatin, follistatin and GDF11 in older women and men. BMC Geriatr. 2018, 18, 200. [Google Scholar] [CrossRef]
- Kalampouka, I.; van Bekhoven, A.; Elliott, B.T. Differing Effects of Younger and Older Human Plasma on C2C12 Myocytes in Vitro. Front. Physiol. 2018, 9, 152. [Google Scholar] [CrossRef] [PubMed]
- Yarasheski, K.E.; Bhasin, S.; Sinha-Hikim, I.; Pak-Loduca, J.; Gonzalez-Cadavid, N.F. Serum myostatin-immunoreactive protein is increased in 60-92 year old women and men with muscle wasting. J. Nutr. Health Aging 2002, 6, 343–348. [Google Scholar] [PubMed]
- Agergaard, J.; Trostrup, J.; Uth, J.; Iversen, J.V.; Boesen, A.; Andersen, J.L.; Schjerling, P.; Langberg, H. Does vitamin-D intake during resistance training improve the skeletal muscle hypertrophic and strength response in young and elderly men?—A randomized controlled trial. Nutr. Metab. (Lond.) 2015, 12, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covington, J.D.; Tam, C.S.; Bajpeyi, S.; Galgani, J.E.; Noland, R.C.; Smith, S.R.; Redman, L.M.; Ravussin, E. Myokine Expression in Muscle and Myotubes in Response to Exercise Stimulation. Med. Sci. Sports Exerc. 2016, 48, 384–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansson, B.; Olsen, L.A.; Nicoll, J.X.; von Walden, F.; Melin, M.; Stromberg, A.; Rullman, E.; Gustafsson, T.; Fry, A.C.; Fernandez-Gonzalo, R.; et al. Skeletal muscle signaling responses to resistance exercise of the elbow extensors are not compromised by a preceding bout of aerobic exercise. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 317, R83–R92. [Google Scholar] [CrossRef]
- Hinkley, J.M.; Konopka, A.R.; Suer, M.K.; Harber, M.P. Short-term intense exercise training reduces stress markers and alters the transcriptional response to exercise in skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 312, R426–R433. [Google Scholar] [CrossRef] [Green Version]
- Popov, D.V.; Lysenko, E.A.; Bachinin, A.V.; Miller, T.F.; Kurochkina, N.S.; Kravchenko, I.V.; Furalyov, V.A.; Vinogradova, O.L. Influence of resistance exercise intensity and metabolic stress on anabolic signaling and expression of myogenic genes in skeletal muscle. Muscle Nerve 2015, 51, 434–442. [Google Scholar] [CrossRef]
- Pugh, J.K.; Faulkner, S.H.; Jackson, A.P.; King, J.A.; Nimmo, M.A. Acute molecular responses to concurrent resistance and high-intensity interval exercise in untrained skeletal muscle. Physiol. Rep. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Ross, C.I.; Shute, R.J.; Ruby, B.C.; Slivka, D.R. Skeletal Muscle mRNA Response to Hypobaric and Normobaric Hypoxia After Normoxic Endurance Exercise. High Alt. Med. Biol. 2019, 20, 141–149. [Google Scholar] [CrossRef]
- Schwarz, N.A.; McKinley-Barnard, S.K.; Spillane, M.B.; Andre, T.L.; Gann, J.J.; Willoughby, D.S. Effect of resistance exercise intensity on the expression of PGC-1alpha isoforms and the anabolic and catabolic signaling mediators, IGF-1 and myostatin, in human skeletal muscle. Appl. Physiol. Nutr. Metab. 2016, 41, 856–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silvennoinen, M.; Ahtiainen, J.P.; Hulmi, J.J.; Pekkala, S.; Taipale, R.S.; Nindl, B.C.; Laine, T.; Hakkinen, K.; Selanne, H.; Kyrolainen, H.; et al. PGC-1 isoforms and their target genes are expressed differently in human skeletal muscle following resistance and endurance exercise. Physiol. Rep. 2015, 3, e12563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, E.L.; Viana, J.L.; Wimbury, D.; Martin, N.; Greening, N.J.; Barratt, J.; Smith, A.C. The Effect of Resistance Exercise on Inflammatory and Myogenic Markers in Patients with Chronic Kidney Disease. Front. Physiol. 2017, 8, 541. [Google Scholar] [CrossRef] [Green Version]
- Hjorth, M.; Pourteymour, S.; Gorgens, S.W.; Langleite, T.M.; Lee, S.; Holen, T.; Gulseth, H.L.; Birkeland, K.I.; Jensen, J.; Drevon, C.A.; et al. Myostatin in relation to physical activity and dysglycaemia and its effect on energy metabolism in human skeletal muscle cells. Acta Physiol. (Oxf.) 2016, 217, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Shad, B.J.; Thompson, J.L.; Holwerda, A.M.; Stocks, B.; Elhassan, Y.S.; Philp, A.; LJC, V.A.N.L.; Wallis, G.A. One Week of Step Reduction Lowers Myofibrillar Protein Synthesis Rates in Young Men. Med. Sci. Sports Exerc. 2019, 51, 2125–2134. [Google Scholar] [CrossRef] [PubMed]
- Han, D.S.; Hsiao, M.Y.; Wang, T.G.; Chen, S.Y.; Yang, W.S. Association of serum myokines and aerobic exercise training in patients with spinal cord injury: An observational study. BMC Neurol. 2016, 16, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Z.; Tian, Y.; Valenzuela, P.L.; Huang, C.; Zhao, J.; Hong, P.; He, Z.; Yin, S.; Lucia, A. Myokine Response to High-Intensity Interval vs. Resistance Exercise: An Individual Approach. Front. Physiol. 2018, 9, 1735. [Google Scholar] [CrossRef]
- He, Z.; Tian, Y.; Valenzuela, P.L.; Huang, C.; Zhao, J.; Hong, P.; He, Z.; Yin, S.; Lucia, A. Myokine/Adipokine Response to “Aerobic” Exercise: Is It Just a Matter of Exercise Load? Front. Physiol. 2019, 10, 691. [Google Scholar] [CrossRef]
- Kabak, B.; Belviranli, M.; Okudan, N. Irisin and myostatin responses to acute high-intensity interval exercise in humans. Horm. Mol. Biol. Clin. Investig. 2018, 35. [Google Scholar] [CrossRef]
- Kerschan-Schindl, K.; Thalmann, M.M.; Weiss, E.; Tsironi, M.; Foger-Samwald, U.; Meinhart, J.; Skenderi, K.; Pietschmann, P. Changes in Serum Levels of Myokines and Wnt-Antagonists after an Ultramarathon Race. PLoS ONE 2015, 10, e0132478. [Google Scholar] [CrossRef] [Green Version]
- Paoli, A.; Pacelli, Q.F.; Neri, M.; Toniolo, L.; Cancellara, P.; Canato, M.; Moro, T.; Quadrelli, M.; Morra, A.; Faggian, D.; et al. Protein supplementation increases postexercise plasma myostatin concentration after 8 weeks of resistance training in young physically active subjects. J. Med. Food 2015, 18, 137–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amani-Shalamzari, S.; Farhani, F.; Rajabi, H.; Abbasi, A.; Sarikhani, A.; Paton, C.; Bayati, M.; Berdejo-Del-Fresno, D.; Rosemann, T.; Nikolaidis, P.T.; et al. Blood Flow Restriction During Futsal Training Increases Muscle Activation and Strength. Front. Physiol. 2019, 10, 614. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, R.; Moghadam, B.H.; Church, D.D.; Tinsley, G.M.; Eskandari, M.; Moghadam, B.H.; Motevalli, M.S.; Baker, J.S.; Robergs, R.A.; Wong, A. The effects of concurrent training order on body composition and serum concentrations of follistatin, myostatin and GDF11 in sarcopenic elderly men. Exp. Gerontol. 2020, 133, 110869. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, R.; Rashidlamir, A.; Motevalli, M.S.; Elliott, B.T.; Mehrabani, J.; Wong, A. Effects of upper-body, lower-body, or combined resistance training on the ratio of follistatin and myostatin in middle-aged men. Eur. J. Appl. Physiol. 2019, 119, 1921–1931. [Google Scholar] [CrossRef]
- Mafi, F.; Biglari, S.; Ghardashi Afousi, A.; Gaeini, A.A. Improvement in Skeletal Muscle Strength and Plasma Levels of Follistatin and Myostatin Induced by an 8-Week Resistance Training and Epicatechin Supplementation in Sarcopenic Older Adults. J. Aging Phys. Act. 2019, 27, 384–391. [Google Scholar] [CrossRef]
- Micielska, K.; Gmiat, A.; Zychowska, M.; Kozlowska, M.; Walentukiewicz, A.; Lysak-Radomska, A.; Jaworska, J.; Rodziewicz, E.; Duda-Biernacka, B.; Ziemann, E. The beneficial effects of 15 units of high-intensity circuit training in women is modified by age, baseline insulin resistance and physical capacity. Diabetes Res. Clin. Pract. 2019, 152, 156–165. [Google Scholar] [CrossRef]
- Wu, Y.; Qu, J.; Li, H.; Yuan, H.; Guo, Q.; Ouyang, Z.; Lu, Q. Relationship between serum level of growth differentiation factors 8, 11 and bone mineral density in girls with anorexia nervosa. Clin. Endocrinol. (Oxf.) 2019, 90, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Amor, M.; Itariu, B.K.; Moreno-Viedma, V.; Keindl, M.; Jurets, A.; Prager, G.; Langer, F.; Grablowitz, V.; Zeyda, M.; Stulnig, T.M. Serum Myostatin is Upregulated in Obesity and Correlates with Insulin Resistance in Humans. Exp. Clin. Endocrinol. Diabetes 2019, 127, 550–556. [Google Scholar] [CrossRef]
- Tsioga, M.N.; Oikonomou, D.; Vittas, S.; Kalscheuer, H.; Roeder, E.; Wintgens, K.F.; Nawroth, P.P.; Wolfrum, C.; Rudofsky, G. Plasma myostatin is only a weak predictor for weight maintenance in obese adults. Exp. Clin. Endocrinol. Diabetes 2015, 123, 466–472. [Google Scholar] [CrossRef]
- Assyov, Y.S.; Velikova, T.V.; Kamenov, Z.A. Myostatin and carbohydrate disturbances. Endocr. Res. 2017, 42, 102–109. [Google Scholar] [CrossRef]
- Tanaka, M.; Masuda, S.; Yamakage, H.; Inoue, T.; Ohue-Kitano, R.; Yokota, S.; Kusakabe, T.; Wada, H.; Sanada, K.; Ishii, K.; et al. Role of serum myostatin in the association between hyperinsulinemia and muscle atrophy in Japanese obese patients. Diabetes Res. Clin. Pract. 2018, 142, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Fontana, B.; Reyes-Garcia, R.; Morales-Santana, S.; Avila-Rubio, V.; Munoz-Garach, A.; Rozas-Moreno, P.; Munoz-Torres, M. Relationship between myostatin and irisin in type 2 diabetes mellitus: A compensatory mechanism to an unfavourable metabolic state? Endocrine 2016, 52, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Han, X.; Meng, Q.; Luo, Q. Early second trimester maternal serum markers in the prediction of gestational diabetes mellitus. J. Diabetes Investig. 2018, 9, 967–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Yang, Q.; Walker, R.G.; Thompson, T.B.; Du, M.; Rodgers, B.D. Myostatin Attenuation In Vivo Reduces Adiposity, but Activates Adipogenesis. Endocrinology 2016, 157, 282–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.; Yao, T.; Zhou, P.; Kazak, L.; Tenen, D.; Lyubetskaya, A.; Dawes, B.A.; Tsai, L.; Kahn, B.B.; Spiegelman, B.M.; et al. Brown Adipose Tissue Controls Skeletal Muscle Function via the Secretion of Myostatin. Cell Metab. 2018, 28, 631–643. [Google Scholar] [CrossRef] [Green Version]
- Steculorum, S.M.; Ruud, J.; Karakasilioti, I.; Backes, H.; Engstrom Ruud, L.; Timper, K.; Hess, M.E.; Tsaousidou, E.; Mauer, J.; Vogt, M.C.; et al. AgRP Neurons Control Systemic Insulin Sensitivity via Myostatin Expression in Brown Adipose Tissue. Cell 2016, 165, 125–138. [Google Scholar] [CrossRef] [Green Version]
- Consitt, L.A.; Clark, B.C. The Vicious Cycle of Myostatin Signaling in Sarcopenic Obesity: Myostatin Role in Skeletal Muscle Growth, Insulin Signaling and Implications for Clinical Trials. J. Frailty Aging 2018, 7, 21–27. [Google Scholar] [CrossRef]
- Akerfeldt, T.; Helmersson-Karlqvist, J.; Gunningberg, L.; Swenne, C.L.; Larsson, A. Postsurgical Acute Phase Reaction is Associated with Decreased Levels of Circulating Myostatin. Inflammation 2015, 38, 1727–1730. [Google Scholar] [CrossRef]
- Wallner, C.; Wagner, J.M.; Dittfeld, S.; Drysch, M.; Lehnhardt, M.; Behr, B. Myostatin serum concentration as an indicator for deviated muscle metabolism in severe burn injuries. Scand. J. Surg. 2019, 108, 297–304. [Google Scholar] [CrossRef]
- Zhao, C.; Shao, Y.; Lin, C.; Zeng, C.; Fang, H.; Pan, J.; Cai, D. Myostatin serum concentrations are correlated with the severity of knee osteoarthritis. J. Clin. Lab. Anal. 2017, 31. [Google Scholar] [CrossRef]
- Su, C.M.; Hu, S.L.; Sun, Y.; Zhao, J.; Dai, C.; Wang, L.; Xu, G.; Tang, C.H. Myostatin induces tumor necrosis factor-alpha expression in rheumatoid arthritis synovial fibroblasts through the PI3K-Akt signaling pathway. J. Cell. Physiol. 2019, 234, 9793–9801. [Google Scholar] [CrossRef] [PubMed]
- Kerschan-Schindl, K.; Ebenbichler, G.; Foeger-Samwald, U.; Leiss, H.; Gesslbauer, C.; Herceg, M.; Stummvoll, G.; Marculescu, R.; Crevenna, R.; Pietschmann, P. Rheumatoid arthritis in remission: Decreased myostatin and increased serum levels of periostin. Wien. Klin. Wochenschr. 2019, 131, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anker, S.D.; Ponikowski, P.; Varney, S.; Chua, T.P.; Clark, A.L.; Webb-Peploe, K.M.; Harrington, D.; Kox, W.J.; Poole-Wilson, P.A.; Coats, A.J. Wasting as independent risk factor for mortality in chronic heart failure. Lancet 1997, 349, 1050–1053. [Google Scholar] [CrossRef]
- Heineke, J.; Auger-Messier, M.; Xu, J.; Sargent, M.; York, A.; Welle, S.; Molkentin, J.D. Genetic deletion of myostatin from the heart prevents skeletal muscle atrophy in heart failure. Circulation 2010, 121, 419–425. [Google Scholar] [CrossRef]
- Gruson, D.; Ahn, S.A.; Ketelslegers, J.M.; Rousseau, M.F. Increased plasma myostatin in heart failure. Eur. J. Heart Fail. 2011, 13, 734–736. [Google Scholar] [CrossRef]
- Ishida, J.; Konishi, M.; Saitoh, M.; Anker, M.; Anker, S.D.; Springer, J. Myostatin signaling is up-regulated in female patients with advanced heart failure. Int. J. Cardiol. 2017, 238, 37–42. [Google Scholar] [CrossRef]
- Chen, P.; Liu, Z.; Luo, Y.; Chen, L.; Li, S.; Pan, Y.; Lei, X.; Wu, D.; Xu, D. Predictive value of serum myostatin for the severity and clinical outcome of heart failure. Eur. J. Intern. Med. 2019, 64, 33–40. [Google Scholar] [CrossRef]
- Meloux, A.; Rochette, L.; Maza, M.; Bichat, F.; Tribouillard, L.; Cottin, Y.; Zeller, M.; Vergely, C. Growth Differentiation Factor-8 (GDF8)/Myostatin is a Predictor of Troponin I Peak and a Marker of Clinical Severity after Acute Myocardial Infarction. J. Clin. Med. 2019, 9, 116. [Google Scholar] [CrossRef] [Green Version]
- Han, D.S.; Hsiao, M.Y.; Wang, T.G.; Chen, S.Y.; Yang, W.S. Relation between serum myokines and phase II cardiac rehabilitation. Medicine (Baltimore) 2017, 96, e6579. [Google Scholar] [CrossRef]
- Baan, J.A.; Varga, Z.V.; Leszek, P.; Kusmierczyk, M.; Baranyai, T.; Dux, L.; Ferdinandy, P.; Braun, T.; Mendler, L. Myostatin and IGF-I signaling in end-stage human heart failure: A qRT-PCR study. J. Transl. Med. 2015, 13, 1. [Google Scholar] [CrossRef] [Green Version]
- Castillero, E.; Ali, Z.A.; Akashi, H.; Giangreco, N.; Wang, C.; Stohr, E.J.; Ji, R.; Zhang, X.; Kheysin, N.; Park, J.S.; et al. Structural and functional cardiac profile after prolonged duration of mechanical unloading: Potential implications for myocardial recovery. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H1463–H1476. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Sola, J.; Borrisser-Pairo, F.; Antunez, E.; Tobias, E. Myostatin and insulin-like growth factor-1 in hypertensive heart disease: A prospective study in human heart donors. J. Hypertens. 2015, 33, 851–858; discussion 859. [Google Scholar] [CrossRef] [PubMed]
- Fernlund, E.; Gyllenhammar, T.; Jablonowski, R.; Carlsson, M.; Larsson, A.; Arnlov, J.; Liuba, P. Serum Biomarkers of Myocardial Remodeling and Coronary Dysfunction in Early Stages of Hypertrophic Cardiomyopathy in the Young. Pediatr. Cardiol. 2017, 38, 853–863. [Google Scholar] [CrossRef] [PubMed]
- Ju, C.R.; Chen, M.; Zhang, J.H.; Lin, Z.Y.; Chen, R.C. Higher Plasma Myostatin Levels in Cor Pulmonale Secondary to Chronic Obstructive Pulmonary Disease. PLoS ONE 2016, 11, e0150838. [Google Scholar] [CrossRef] [PubMed]
- Ju, C.R.; Zhang, J.H.; Chen, M.; Chen, R.C. Plasma myostatin levels are related to the extent of right ventricular dysfunction in exacerbation of chronic obstructive pulmonary disease. Biomarkers 2017, 22, 246–252. [Google Scholar] [CrossRef]
- Zhang, Y.B.; Zuo, H.Y.; Tian, D.H.; Ouyang, X.H.; Wang, X.A. Correlation between peripheral skeletal muscle functions and the stable phase of COPD in older patients. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5317–5326. [Google Scholar] [CrossRef]
- Qi, H.; Ren, J.; Ba, L.; Song, C.; Zhang, Q.; Cao, Y.; Shi, P.; Fu, B.; Liu, Y.; Sun, H. MSTN Attenuates Cardiac Hypertrophy through Inhibition of Excessive Cardiac Autophagy by Blocking AMPK /mTOR and miR-128/PPARgamma/NF-kappaB. Mol. Ther. Nucleic Acids 2019, 19, 507–522. [Google Scholar] [CrossRef]
- Biesemann, N.; Mendler, L.; Kostin, S.; Wietelmann, A.; Borchardt, T.; Braun, T. Myostatin induces interstitial fibrosis in the heart via TAK1 and p38. Cell Tissue Res. 2015, 361, 779–787. [Google Scholar] [CrossRef]
- Yano, S.; Nagai, A.; Isomura, M.; Yamasaki, M.; Kijima, T.; Takeda, M.; Hamano, T.; Nabika, T. Relationship between Blood Myostatin Levels and Kidney Function:Shimane CoHRE Study. PLoS ONE 2015, 10, e0141035. [Google Scholar] [CrossRef]
- Raptis, V.; Bakogiannis, C.; Loutradis, C.; Boutou, A.K.; Sioulis, A.; Balaskas, E.; Zebekakis, P.; Sarafidis, P.A. Serum Fas Ligand, Serum Myostatin and Urine TGF-beta1 Are Elevated in Autosomal Dominant Polycystic Kidney Disease Patients with Impaired and Preserved Renal Function. Kidney Blood Press. Res. 2018, 43, 744–754. [Google Scholar] [CrossRef]
- Esposito, P.; La Porta, E.; Calatroni, M.; Grignano, M.A.; Milanesi, S.; Verzola, D.; Battaglia, Y.; Gregorini, M.; Libetta, C.; Garibotto, G.; et al. Modulation of Myostatin/Hepatocyte Growth Factor Balance by Different Hemodialysis Modalities. BioMed Res. Int. 2017, 2017, 7635459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, P.; Battaglia, Y.; La Porta, E.; Grignano, M.A.; Caramella, E.; Avella, A.; Peressini, S.; Sessa, N.; Albertini, R.; Di Natali, G.; et al. Significance of serum Myostatin in hemodialysis patients. BMC Nephrol. 2019, 20, 462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.T.; Yang, Y.J.; Huang, R.H.; Zhang, Z.H.; Lin, X. Myostatin Activates the Ubiquitin-Proteasome and Autophagy-Lysosome Systems Contributing to Muscle Wasting in Chronic Kidney Disease. Oxid. Med. Cell. Longev. 2015, 2015, 684965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrarelli, P.; Yen, C.F.; Arcuri, F.; Funghi, L.; Tosti, C.; Wang, T.H.; Huang, J.S.; Petraglia, F. Myostatin, follistatin and activin type II receptors are highly expressed in adenomyosis. Fertil. Steril. 2015, 104, 744–752 e741. [Google Scholar] [CrossRef] [PubMed]
- Carrarelli, P.; Funghi, L.; Ciarmela, P.; Centini, G.; Reis, F.M.; Dela Cruz, C.; Mattei, A.; Vannuccini, S.; Petraglia, F. Deep Infiltrating Endometriosis and Endometrial Adenocarcinoma Express High Levels of Myostatin and Its Receptors Messenger RNAs. Reprod. Sci. 2017, 24, 1577–1582. [Google Scholar] [CrossRef]
- Peiris, H.N.; Georgiou, H.; Lappas, M.; Kaitu’u-Lino, T.; Salomon, C.; Vaswani, K.; Rice, G.E.; Mitchell, M.D. Expression of Myostatin in Intrauterine Growth Restriction and Preeclampsia Complicated Pregnancies and Alterations to Cytokine Production by First-Trimester Placental Explants Following Myostatin Treatment. Reprod. Sci. 2015, 22, 1202–1211. [Google Scholar] [CrossRef]
- Tsigkou, A.; Reis, F.M.; Ciarmela, P.; Lee, M.H.; Jiang, B.; Tosti, C.; Shen, F.R.; Shi, Z.; Chen, Y.G.; Petraglia, F. Expression Levels of Myostatin and Matrix Metalloproteinase 14 mRNAs in Uterine Leiomyoma are Correlated With Dysmenorrhea. Reprod. Sci. 2015, 22, 1597–1602. [Google Scholar] [CrossRef]
- Trappe, T.A.; Standley, R.A.; Jemiolo, B.; Carroll, C.C.; Trappe, S.W. Prostaglandin and myokine involvement in the cyclooxygenase-inhibiting drug enhancement of skeletal muscle adaptations to resistance exercise in older adults. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R198–R205. [Google Scholar] [CrossRef] [Green Version]
- D’Lugos, A.C.; Patel, S.H.; Ormsby, J.C.; Curtis, D.P.; Fry, C.S.; Carroll, C.C.; Dickinson, J.M. Prior acetaminophen consumption impacts the early adaptive cellular response of human skeletal muscle to resistance exercise. J. Appl. Physiol. (1985) 2018, 124, 1012–1024. [Google Scholar] [CrossRef]
- Heisterberg, M.F.; Andersen, J.L.; Schjerling, P.; Bulow, J.; Lauersen, J.B.; Roeber, H.L.; Kjaer, M.; Mackey, A.L. Effect of Losartan on the Acute Response of Human Elderly Skeletal Muscle to Exercise. Med. Sci. Sports Exerc. 2018, 50, 225–235. [Google Scholar] [CrossRef]
- Schakman, O.; Gilson, H.; Kalista, S.; Thissen, J.P. Mechanisms of muscle atrophy induced by glucocorticoids. Horm. Res. 2009, 72 (Suppl. 1), 36–41. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Jiao, H.; Zhao, J.; Wang, X.; Lin, H. Glucocorticoids Enhance Muscle Proteolysis through a Myostatin-Dependent Pathway at the Early Stage. PLoS ONE 2016, 11, e0156225. [Google Scholar] [CrossRef] [PubMed]
- Macedo, A.G.; Krug, A.L.; Souza, L.M.; Martuscelli, A.M.; Constantino, P.B.; Zago, A.S.; Rush, J.W.; Santos, C.F.; Amaral, S.L. Time-course changes of catabolic proteins following muscle atrophy induced by dexamethasone. Steroids 2016, 107, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Dhindsa, S.; Ghanim, H.; Green, K.; Abuaysheh, S.; Batra, M.; Makdissi, A.; Chaudhuri, A.; Sandhu, S.; Dandona, P. Acute effects of insulin on skeletal muscle growth and differentiation genes in men with type 2 diabetes. Eur. J. Endocrinol. 2019, 181, K55–K59. [Google Scholar] [CrossRef] [PubMed]
- Hong, O.K.; Choi, Y.H.; Kwon, H.S.; Jeong, H.K.; Son, J.W.; Lee, S.S.; Kim, S.R.; Yoon, K.H.; Yoo, S.J. Long-term insulin treatment leads to a change in myosin heavy chain fiber distribution in OLETF rat skeletal muscle. J. Cell. Biochem. 2019, 120, 2404–2412. [Google Scholar] [CrossRef] [PubMed]
- Yamakage, H.; Tanaka, M.; Inoue, T.; Odori, S.; Kusakabe, T.; Satoh-Asahara, N. Effects of dapagliflozin on the serum levels of fibroblast growth factor 21 and myokines and muscle mass in Japanese patients with type 2 diabetes: A randomized, controlled trial. J. Diabetes Investig. 2020, 11, 653–661. [Google Scholar] [CrossRef] [Green Version]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baczek, J.; Silkiewicz, M.; Wojszel, Z.B. Myostatin as a Biomarker of Muscle Wasting and other Pathologies-State of the Art and Knowledge Gaps. Nutrients 2020, 12, 2401. https://doi.org/10.3390/nu12082401
Baczek J, Silkiewicz M, Wojszel ZB. Myostatin as a Biomarker of Muscle Wasting and other Pathologies-State of the Art and Knowledge Gaps. Nutrients. 2020; 12(8):2401. https://doi.org/10.3390/nu12082401
Chicago/Turabian StyleBaczek, Jan, Marta Silkiewicz, and Zyta Beata Wojszel. 2020. "Myostatin as a Biomarker of Muscle Wasting and other Pathologies-State of the Art and Knowledge Gaps" Nutrients 12, no. 8: 2401. https://doi.org/10.3390/nu12082401
APA StyleBaczek, J., Silkiewicz, M., & Wojszel, Z. B. (2020). Myostatin as a Biomarker of Muscle Wasting and other Pathologies-State of the Art and Knowledge Gaps. Nutrients, 12(8), 2401. https://doi.org/10.3390/nu12082401