Select Polyphenol-Rich Berry Consumption to Defer or Deter Diabetes and Diabetes-Related Complications

 , , , and
, , , and 
Abstract
:1. Introduction
2. Methods
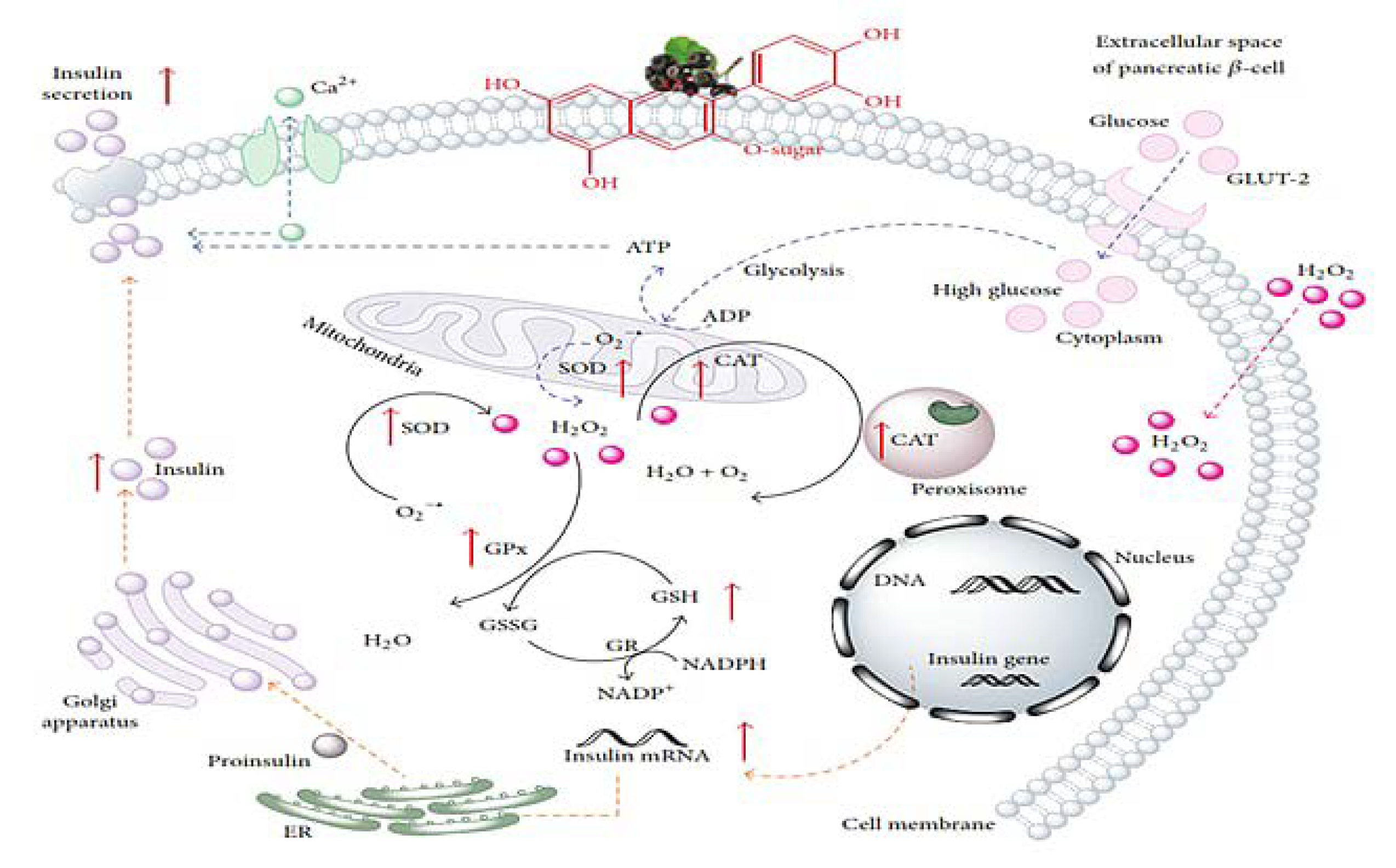
3. Blueberries

{kind=link}
{kind=link}
| No. | Study Design | Study Subject | Duration | Berry Interventions | Intervention Diet | Significant Findings | Ref. |
|---|---|---|---|---|---|---|---|
| (I) Blueberries (BlB) (Animal studies) | |||||||
| 1 | RCT | C57bl/6J mice (n = NS) | 12 wk | BlB anthocyanins 300–500 mg/kg.BW.day | LFD (20% kcal from lard fat) and HFD (70%kcal from lard fat) | Lower the blood glucose level and dyslipidemia markers | [13] |
| 2 | RCT | Male C57BL/6 mice (n = 24) | 8 wk | 4% (w/w) F/D whole BlB powder with HFD provided ad libitum | HFD (60% kcal from fat) | Offer protection against HFD-induced obesity, adipose tissue macrophages inflammatory gene expression, and oxidative stress | [11] |
| 3 | RCT | C57/Bl6 mice (n = 200) | 12 wk | 5% or 10% (w/w) of whole BlB with HFD provided ad libitum | HFD (45% kcal from fat) | Reduced HFD-induced cellular inflammatory cytokines, chemokines, interleukins, and proinflammatory interferon gamma -producing T-cells | [18] |
| 4 | RCT | Male Zucker Fatty and Zucker Lean rats (n = 48) | 8 wk | 4% (w/w) F/D whole BlB powder with HFD provided ad libitum | HFD (45% kcal from fat) | Hypolipidemic, Hyper-insulinemic, hypoglycemic and anti-inflammatory | [40] |
| 5 | RCT | C57BL/6 mice (n = 48) | 12 wk | Ad libitum 100% BlBJ with HFD provided ad libitum | LFD (20% kcal from fat) and HFD (45%kcal from fat) | Reduced expressions of inflammatory and FA synthesis genes. Reduced IR and plasma dyslipidemia markers | [12] |
| 6 | RCT | C57BL/6 mice (n = 72) | 8 wk | 65.1 ± 1.6 mg cyanidin-3-O-glucoside/L (from 30% BlB + 70% blackberry juice available ad libitum) | HFD (60% kcal from fat) | Anti-obesity, hypoglycemic, antidiabetic | [22] |
| 7 | RCT | C57BL/6 and KKAy mice (n = 20) | 4 wk | BlBJ (40–80 mL/kg per day in drinking water) | Normal chow diet | Improved glucose tolerance, reduced glycemic response suggesting increased insulin sensitivity | [23] |
| 8 | RCT | Obese Zucker rat (n = 20) | 8 wk | 8% wild BlB diet (WB) provided ad libitum | NA | Downregulated expression and plasma concentrations of NF-kB, TNFα, IL-6, CRP in liver and adipose tissues | [25] |
| Human studies | |||||||
| 9 | SB and RCT | Obese men and women (n = 66) | 8 wk | 50 g F/DBlB per day | NA | Reduction in plasma oxidized LDL and other plasma lipid oxidation products | [20] |
| 10 | DB, PC, RCT | overweight or obese individuals (n = 30) | 4 wk | 4 g of inulin/day from BlB (equivalent to two cups of whole BlB) | NA | Improvement in glycemic response, insulin sensitivity, satiety, serum lipid parameters, and fecal markers of gut microbiota | [41] |
| 11 | DB, PC, RCT | Diabetic patients (n = 58) | 24 wk | 160 mg of BlB anthocyanins twice daily | NA | Reduced serum concentration of LDL-C, TG, apolipoprotein, apolipoprotein C-III, lipid and protein oxidation markers with strengthening the inherent antioxidative system | [42] |
| 12 | DB, PC, RCT | Healthy adults (n = 44) | 6 wk | 45 g/day F/D BlBpowder | 12-oz yogurt and skim milk-based smoothie | Improvement in endothelial function in subjects with metabolic syndrome | [21] |
| 13 | DB, CO, RCT, | Healthy human beings (n = 17) | 4 wk | 310–724 mg/kg.BW.day BlBanthocyanin | Sugar-matched smoothie | Extend the postprandial glucose response beyond the period observed for a sugar-matched control | [24] |
| (I) Bilberries (BB) (Animal studies) | |||||||
| 1 | RCT | Male KK-Ay mice (n = 16) | 5 wk | 27 g of BB extract/kg diet daily | NA | Activation of AMP-activated protein kinase (AMPK) resulting in increased insulin sensitivity, upregulation of glucose transporter GLUT4, suppression of glucose production in liver | [15] |
| 2 | RCT | diabetic groups of Wistar rats (n = 32) | 4 wk | BB extracts 2 g/day by gavage | Normal chow diet | Increased serum insulin, reduced TC, VLDL-C, LDL-C, and TG levels, and prevented HDL-C decline | [17,43] |
| 3 | RCT | Brown Norway (BN) rats (n = 96) | 6 wk | BB extract 100 mg/kg.BW.day | Normal chow diet | Prevent diabetic retinopathy | [44] |
| 4 | RCT | Male KM mice (n = 60) | 5 d | BB extract (containing 42.04% anthocyanins) 200 mg/kg BW·day) | Normal chow diet | Reduced the live damage and oxidative stress markers (ALT, MDA, NO) with improvements in enzymatic antioxidative system (GSH) | [45] |
| 5 | RCT | Male Wistar rats (n = 15) | 12 wk | 40 mg/kg.day BB extracts in 5 mL drinking water | Normal chow diet | Prevent capillary albumin filtration | [46] |
| 6 | RCT | Goto-Kakizaki (GK) rat (n = NS) | 4 wk | BB decoction with rodent chow | powdered rodent chow | Improved mitochondrial respiratory and biogenesis parameters | [47] |
| (Human studies) | |||||||
| 7 | DB, CO, RCT | T2DM Male volunteer subjects (n = 8) | 24 h | A single capsule of 0 × 47 g BB extract (36% w/w) | NA | Decrease in the incremental AUC for both glucose and insulin without alterations in GLP1, glucagon, amylin, and anti-inflammatory peptides | [48] |
| 8 | CO, DB, RCT design | Obese/Overweight/diabetic men and women (n = 16) | 3 wk | 3 × 0.47 g of Mirtoselect capsules per day, a standardized BB extract (36% w/w) anthocyanins) | NA | Reduced activity of digestion enzymes without alterations in anti-inflammatory markers, vascular health markers and reducing capacity | [49] |
| 9 | RCT | Healthy men and women (n = 9) | 1 d | 10% BB in fermented drink up to 300 g/day | White bread | Lower the insulin response than glycemic response | [50] |
| 10 | RCT | Healthy men and women (n = 62) | 4 wk | BB juice 330 mL/day | NA | Anti-inflammatory | [51] |
| 11 | Parallel RCT | Healthy men (n = 40) | 8 wk | Fresh BB 100 g/day of BB | NA | Increased intestinal bioavailability of antioxidative and antidiabetic compounds | [52] |
| 12 | RCT | Healthy men and women (n = 27) | 8 wk | Fresh BB 400 g/day | NA | Reduction in the low-grade inflammation with different cytoplasmic ribosomal proteins, Toll-like receptor, and B-cell receptor signaling pathways | [53] |
| (I) Cranberries (CrB) (Animal studies) | |||||||
| 1 | RCT | 36 C57Bl/6J male mice | 8 wk | CrB extracts 200 mg/kg BW on daily basis by gavage | HFD (65% lipids, 15% proteins and 20% carbohydrates) | Attenuated HFD-induced obesity, TC and TG accumulation, oxidative stress, with improvements in glycemic response, insulin sensitivity, HOMAIR, alleviate intestinal inflammation | [54] |
| 2 | RCT | Male Fischer rats (n = 24) | 16 m | 2% whole CrB powder standard NIH-31 standard rodent chow available ad libitum | NA | Increased β-cell glucose responsiveness; age related decline in in basal plasma insulin concentrations was delayed by cranberry | [55] |
| Human studies | |||||||
| 3 | CO, RCT | Obese participants (n = 25) | 2–4 h | Cranberries (40 g) | HF breakfast (70 g fat, 974 kcal) | Improved postprandial glycemic control, reduction in lipid oxidation products and inflammatory cytokines | [56] |
| 4 | PC, DB, RCT | T2DM men and women (n = 30) | 12 wk | 3 capsules of CrB extract/day (1 capsule = 500 mg) | NA | Decrease in the TC:HDLC ratio | [57] |
| 5 | single CO RCT | Healthy men and women (n = 12) | OTCS | Dextrose sweetened normal calorie CrB juice (NCCBJ; 27% CBJ, v/v; 130 Cal/240 mL) and low-calorie CrB juice (LCCBJ; 27%, v/v CrBJ;19 Cal/240 mL) | 5 g Vanilla Crisp Power Bar (contained 230 Cal, 2.5 g total fat, 3 g dietary fiber, 20 g sugars, 22 g other carbo-hydrates, and 9 g protein | Improved metabolic response towards glucose | [58] |
| 6 | RCT | Non-diabetic men and women (n = 187) | OTCS | low-calorie 27% CrBJ (19 Cal/240 mL); normal-calorie 27%CrBJ (140 Cal/240 mL) at weight-adjusted serving size (480 mL/70 kg) | NA | Improved glycemic response | [59] |
| 7 | RCT | T2DM men and women (n = 13) | OTCS | Raw CrB (55 g, 21 cal, 1 g fiber); Sweetened dried CrB (40 g, 138 cal, 2.1 g fiber); Sweetened dried CrB containing less sugar (SDC-LS, 40 g, 113 cal, 1.8 g fiber + 10 g polydextrose) | White bread (57 g, 160 cal, 1 g fiber) | favorable glycemic and insulinemic response | [60] |
| 8 | CS Nutrition Examination Survey (n = 10 891) | Healthy men and women | 2 days | Average 2-day CrBJ intake 158 to 404 mL | Routine diet | Lowered the weight-gain, TC, and proinflammatory serum CRP levels | [61] |
| 9 | DB, CO, RCT | Healthy men and women (n = 12) | OTCS | 37·5 g of CrB in addition to 37.5 g × 3 of other berries (bilberries, strawberries, blueberries) + 35 g added sugar | NA | Hypoglycemic and hypo-insulinemic | [62] |
| 10 | Parallel RCT | Diabetic men and women (n = 48) | 8 wk | 200 × 2 mL RCCJ was enriched with omega-3 fatty acid (180 mg EPA + 120 mg DHA) on daily basis | usual diet and physical activity during the study | Anti-dyslipidemic and hypoglycemic | [63] |
| 11 | Parallel DB RCT | T2DM male patients (n = 58) | 12 wk | 1 cup (240 mL) CrB juice daily | NA | Antiglycation, antidiabetic, reducing CVD risk factors in T2DM male patients | [64] |
| 12 | Parallel DB, PC RCT | Healthy men and women (n = 56) | 8 wk | 480 mL (80 kcal) whole CrB juice daily | Complete diets in addition to Background diets consisted of typical American foods (HFD) and 3–5 servings of fruits or vegetables daily (328– 618 g/d depending on energy intake) | Anti-dyslipidemic, hypoglycemic, improved HOAM-IR | [65] |
| 13 | RCT | Patients with metabolic syndrome (n = 55) | 60 d | 0·7 L/day (J (20 kcal) of reduced-energy CrB juice containing 66 mg proanthocyanidins; total phenolics of 104 and 0·12 mg folic acid | NA | An increase in adiponectin and folic acid and a decrease in homocysteine, decreased lipoperoxidation and protein oxidation levels | [66] |
| 14 | CO, DB | Obese yet healthy men (n = 35) | 4 wk | 500 mL CrB juice/day | NA | Improved augmentation in obese men | [67] |
| 15 | DB, CO | Obese men (n = 30) | 12 wk | Increasing doses of low-calories CrBJ during three successive periods of 4 wk (wk 1–4: 125 mL/day, wk 5–8: 250 mL/day, and wk 9–12: 500 mL/day) | NA | Improved antioxidative defense system | [68] |
| 16 | DB, CO | Obese men (n = 30) | 12 wk | Increasing doses of low-calories CrBJ during three successive periods of 4 wk (wk 1–4: 125 mL/day, wk 5–8: 250 mL/day, and wk 9–12: 500 mL/day) | NA | Decrease in plasma OxLDL, intercellular adhesion molecule-1 (ICAM-1), vascular cell adhesion molecule-1 (VCAM-1) and E-selectin concentrations | [69] |
| 17 | CO, DB | Abdominally obese men (n = 30) | 12 wk | Increasing doses of low-calories CrBJ during three successive periods of 4 wk (wk 1–4: 125 mL/day, wk 5–8: 250 mL/day, and wk 9–12: 500 mL/day) | NA | Increased plasma HDL-cholesterol concentrations | [70] |
| 18 | RCT | Healthy men (n = 21) | 2 wk | CrBJ 7 mL/kg.BW.day | NA | Reduction in plasma OxLDL levels and Improved antioxidative defense system | [71] |
| (I) Raspberries (RB) (Animal studies) | |||||||
| 1 | RCT | Weanling male Syrian golden hamsters | 12 wk | RBJ 275 mL/day (1 mL = 0.6 g of berries) | semi-purified hyperlipidic diet (0.5% cholesterol and 15% lard) | Hypo-cholestrolemic and antioxidative | [72] |
| 2 | RCT | Male Wistar rats (n = 30) | 10 d | Dose of ellagitannins enriched RB extracts equivalent to daily consumption of 125 g of fresh fruit by a human healthy adult of 70 kg (i.e., 20 mg/kg BW.day orally) | NA | Protection from the ethanol induced oxidative stress and inflammatory biomarkers | [73] |
| 3 | RCT | male Lewis rats (n = 24) | 30 d | RB extracts at 30–120 mg/kg.BW | NA | Inhibition of inflammation, pannus formation, cartilage damage, and bone resorption | [74] |
| 4 | RCT | CD1 male mice (n = 36) | 4 wk | RB infusion by gavage (100 mg/kg BW.day) | NA | Improved antioxidative defense system | [75] |
| 5 | RCT | obese diabetic (db/db) mice (n = 30) | 8 wk | 5.3% RB supplementation along agar-based diet finally containing polyphenolics (963 mg extractable GAE/kg agar-based diet) | agar-based diet | Hyper-cholestrolemic and diabetes-induced oxidative stress | [76] |
| 6 | RCT | Male Zucker Fatty rats (n = NS) | 12 wk | 20 g of diet per day containing RB (2% red raspberry F/D powder) | NA | Upregulation of the expression of myocardial adiponectin receptor 1 and apolipoprotein E, improving the plasma cholesterol and triglyceride homeostasis | [77] |
| 7 | RCT | Male Wistar rats (n = 42) | 5 d | 313 g whole RB with/without Lactobacillus plantarum HEAL19 (HEAL19 at 109 cfu) per day with diet | Normal chow diet | Increased intestinal SCFA load and anti-inflammatory | [78] |
| 8 | RCT | Male F-344 rats (n = NS) | 6 wk | AIN-76A diet containing either 5% whole BRB powder, 0.2% BRB anthocyanins, or 2.25% of the residue fraction provided ad libitum | NA | Anti-dysbiosis, anti-inflammatory, anti-obesity | [79] |
| 9 | RCT | Male db/db mice with C57BL/6J Background (n = 48) | 8 wk | 150 mg/kg.BW.day per mice RB derived pelargonidin-3-O-glucoside | NA | Hypoglycemic, anti-inflammatory, anti-obesity | [80] |
| 10 | RCT | Specific-pathogen free C57BL/6 mice (n = 20) | 7 wk | AIN-76A diet with 10% black raspberry powder provided ad libitum | NA | Hypo-glycemic, anti-metabolic syndromic | [81] |
| 11 | RCT | Male db/db mice (n = 30) | 8 wk | 10% F/D RB in a isocaloric standard diet | Isocaloric standard diet | Hypo-cholestrolemic, antioxidative, improved insulin sensitivity | [82] |
| 12 | RCT | C57BL/6J mice (n = NS) | 10 wk | Energy-containing RB foods (juice and puree concentrate and whole fruit powder) containing 10% raspberry and HFD supplemented with 0.2% (w/w) RB extract provided ad libitum | HFD (45% energy from fat) + high-carbohydrate food (35% energy from starch) | Anti-obesity and antidiabetic | [83] |
| 13 | RCT | C57BL/6J, C57BL/Ks db/db, and db/+ male mice (n = NS) | 8 wk | 0.2% Cyanidin 3-glucoside in HFD | HFD (58% of calories from coconut hydrogenated fat) | Anti-obesity, anti-inflammatory, improvement in the insulin sensitivity | [84] |
| 14 | RCT | Male Sprague Dawley rats (n = 40) | 8 wk | Application of RB derived EA (0.1–10 mg/mL) on ischemic stomach (1.5 mL/100 g.BW) in an in an ex vivo chamber | NA | Gastric protective action against gastric lesions induced by NH4OH, due to anti-oxidative activity of EA | [85] |
| 15 | RCT | Male Wistar rats (n = 22) | 4 wk | Oral administration of 10–20 mg/kg.BW of RB derived elagic acid | NA | Anti-inflammatory and anti-oxidative | [86] |
| 16 | RCT | Male Wistar rats AMPKα1−/− (n = 12) | 10 wk | 5% supplementation of RB extracts (contains polyphenols at ~11 g gallic acid equivalent (GAE)/kg of DW) along HFD | HFD (60% from fat) | reduced ectopic lipid storage, alleviated inflammation responses, improved whole-body insulin sensitivity, and promoted mitochondrial biogenesis | [87] |
| 17 | RCT | Male mice (C57BL/6) (n = 40) | 12 wk | 5% F/D RB powder in HFD provided ad libitum | HFD (60% energy from fat) | Anti-dyslipidemic, hypoglycemic | [88] |
| 18 | RCT | Male KK-Ay mice (n = NS) | 5 weeks | Cyanidin 3-glucoside 2 g/kg.BW.day in the normal chow diet | NA | Anti-obesity, anti-inflammatory, improvement in the insulin sensitivity | [89] |
| 19 | RCT | Male mice (C57BL/6) (n = 40) | 12 wk | 3% RB seed floor (equivalent to 0.03% ellagic acid) in HFD and HFD + High-sucrose diet | HFD (41% energy from fat) HFD + High-sucrose diet (37% energy from sucrose) | Anti-dyslipidemic, hypoglycemic, attenuated hepatic ER and oxidative stresses, as well as adipocyte inflammation | [90] |
| Human studies | |||||||
| 20 | PC, CO, RCT | Healthy men and women (n = 20) | 4 wk | High-carbohydrate bars (120–123 g) containing freeze-dried black RB (10% (LOW-Rasp) or 20% (HIGH-Rasp)), One bar each day after overnight fasting. | macronutrient-matched high-carbohydrate cereal bars (45% total sugars) | [91] | |
| 21 | RCT | Healthy men and women (n = 12) | NS | 100 g RB along the designated diet | High-carbohydrate food in the form of pancakes (50 g available carbohydrate from 333 kcal pancake) | Alter postprandial hyperglycemia to sustainable glycemic response | [92] |
| 22 | 3 randomized, controlled, CO, | Healthy women (n = 13–20) | OTCS | 150 g whole berries puree along each meal study 1: white bread + strawberries, bilberries, or lingonberries study 2: white bread + h raspberries, cloudberries, or chokeberries study 3: white bread or rye bread + mix berries consisting of equal amounts of strawberries, bilberries, cranberries, and blackcurrants | White bread or rye bread with 50 g available starch | Reduced the postprandial insulin response, improved the glycemic profile, improved postprandial glucose metabolism. | [93] |
| 23 | CO, RCT | T2DM men and women (n = NS) | 12 wk | 250 g frozen red raspberries puree with each breakfast | NA | Anti-dyslipidemic, anti-inflammatory, anti-obesity | [94] |
| (I) Mulberries (MBs) (Animal studies) | |||||||
| 1 | Randomized block design | Male C57BL/6 mice (n = 60) | 8 wk | MB anthocyanins at 200 mg/kg HFD provided ad libitum | HFD (45% kcal from fat) | Anti-dyslipidemia, anti-inflammatory, anti-obesity | [95] |
| 2 | RCT | Male db/db mice with C57BL6/J genetic background (n = 50) | 8 wk | MB fruit extracts 50 and 125 mg/kg BW every day orally by gavage | NA | Antioxidative and hypoglycemic | [96] |
| 3 | RCT | male adult Wistar rats (n = 70) | 6 wk | MB fruit wine 400 mL/70 kg of body weight daily | NA | Antioxidative and hypoglycemic | [97] |
| 4 | RCT | male Sprague–Dawley rats (n = 50) | 8 wk | MB fruit derived cyanidin-3-O-β-D-glucopyranoside (10 mg/kg.BW. daily) orally by gavage | NA | Antidiabetic cystopathy | [98] |
| 5 | RCT | Adult diabetic male Wistar rats (n = 12) | 6 wk | MB polysaccharides (200 mg/kg.BW daily) | HFD | Improved oral glucose tolerance/insulin resistance, bioactivities of superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPx), were increased | [99] |
| 6 | RCT | Male Gold Syrian hamsters (n = NS) | 12 wk | Water extracts of MB fruit at 1–2% (w/w) in HFD provided ad libitum | HFD (1% cholesterol and 10% corn oil) | Hypolipidemic | [100] |
| 7 | RCT | Male C57BL/6 mice (n = 48) | 12 wk | Anthocyanin from MB of 40–200 mg/kg of HFD | HFD (45% kcal from fat) | Inhibit body weight gain, reduce the resistance to insulin, lower the size of adipocytes, attenuate lipid accumulation and decrease the leptin secretion. | [101] |
| 8 | RCT | Male Syrian golden hamsters (n = 32) | 12 wk | Water extracts of MB fruit at 0.5–2% (w/w) in HFD provided ad libitum | HFD (10% corn oil + 0.1% cholesterol) | Anti-obese and hypolipidemic effects | [102] |
| 9 | RCT | Male Wistar rats (n = 32) | 4 wk | 5–10% (w/w) mulberry fruit polysaccharide fractions in HFD provided ad libitum | HFD (10% lard, 1% cholesterol, 0.5% sodium cholate, and 88.5% commercial diet) | Hypolipidemic and improved the enzymatic antioxidant system | [103] |
| 10 | RCT | Male C57BL/6 mice (n = 48) | 6 wk | 0.5–2% (w/w) water extracts of MB fruit in high-fat (35% kcal from fat) ethanol rich liquid diet (36%kcal from ethanol) provided ad libitum | high-fat (35% kcal from fat) ethanol rich liquid diet (36%kcal from ethanol) | Anti-obesity, hypoglycemic, antioxidative, anti-inflammatory | [104] |
| 11 | RCT | Male Sprague-Dawley rats (n = 40) | 10 wk | MB fruit extracts 100 or 200 mg/kg.BW.day | HFD (1% cholesterol, 18% lipid (lard), 40% sucrose) | Anti-dyslipidemic, antioxidative, ameliorates nonalcoholic fatty liver disease (NAFLD) | [105] |
| 12 | RCT | Female Wistar rats (n = 48) | 20 wk | Microencapsulated 50 to 250 mg/kg.BW.day mulberry fruit extracts (microencapsulated) with HCHF | High-carbohydrate high-fat (HCHF) diet which contained total energy around 4.62 kcal/g (fat 31.54%, protein 20.25%, and carbohydrate 48.21%). | Anti-inflammatory, antioxidative, improved metabolic syndrome | [106] |
| 13 | RCT | Male, C57BL/6J mice (n = 12) | 13 wk | 20% MB powder in HFD provided ad libitum | HFD, 60% calories from fat | Anti-obesity, antidiabetic, increase of Bacteroidetes/Firmicutes ratio | [107] |
| 14 | RCT | db/m mice (n = 50) | NS | MB fruit polysaccharide fractions (200–800 mg/kg.BW) | NA | Improved antioxidant enzymatic defense system, antihyperglycemic and antihyperlipidemic effects | [108] |
| 15 | RCT | Male C57BL6/J genetic background (db/db) mice (n = 60) | 8 wk | Mulberry fruit extract 25–250 mg/kg BW daily | NA | Upregulation of gluconeogenesis pathway | [109] |
| 16 | RCT | Adult diabetic male Wistar (n = 40) | 7 wk | MB fruit polysaccharide fractions MFP50 and MFP90 (400 mg/kg.BW) | HFD | Antihyperglycemic and antihyperlipidemic effects | [110] |
| (I) Lingonberries (LB) (Animal studies) | |||||||
| 1 | RCT | Male C57BL/6 mice (n = NS) | 8 wk | LB extracts (125, 250, and 500 mg/kg) in HFD provided ad libitum | HFD (35% fat, 20% protein, and 36.5% carbohydrate) | Attenuates hepatic steatosis hyperglycemia, hyperlipidemia. Improves insulin signaling | [111] |
| 2 | RCT | SHR rats (n = NS) | 8 wk | Cold-compressed LB juice provided ad libitum | NA | Reduced hypertension and pro-inflammatory markers | [112] |
| 3 | RCT | Male C57BL/6JBomTac mice (n = 120) | 13 wk | 20% (w/w) F/D LB in HFD provided ad libitum | HFD (45 kcal% fat) | Significantly reduced body fat, lipid accumulation, and plasma levels of the inflammatory marker PAI-1, as well as mediated positive effects on glucose metabolism homeostasis. | [113] |
| 4 | RCT | Male C57BL/6JBomTac mice (n = NS) | 11 wk | 20% (w/w) F/D LB in HFD provided ad libitum | HFD (45 kcal% fat) | Reduced plasma levels of markers of endotoxemia and inflammation | [114] |
| 5 | RCT | Male Apoe-/- mice (n = 35) | 8 wk | 44% lingonberry + HFD | HFD (38 kcal% fat) | Decreased triglyceridemia and reduced atherosclerosis | [115] |
| 6 | RCT | Male C57BL/6JBomTac mice (n = NS) | 11 wk | 20% (w/w) F/D LB in HFD provided ad libitum | HFD (45 kcal% fat) | Improvement in glycaemia, reduction in inflammation and hepatic steatosis | [116] |
| 7 | RCT | C57BL/6JBomTac (n = NS) | 13 wk | 20% (w/w) freeze-dried LB + blackcurrants, bilberries or açai berry in HFD provided ad libitum | HFD (45 kcal% fat) | Downregulation of inflammatory pathways, NF-κB, STAT3 and mTOR as possible targets for antidiabetic therapy | [117] |
| 8 | RCT | Male ApoE−/− mice (n = 50) | 8 wk | Two LB polysaccharide fractions 15–60 g/kg BW with HFD daily | HFD (38 kcal% fat) | Hypoglycemic, hypolipidemic, altered caecal microbiota composition | [118] |
| Human studies | |||||||
| 9 | RCT | Scandinavian type 2 diabetes patients (n = 30) | 12 wk | Recommended daily intake of LB/berries/fruits | Okinawan-based Nordic diet of about 1,900 kcal/day | Improved metabolic and anthropometric parameters | [119] |
| 10 | CO, DB, RCT | Healthy normal-weight nonsmoking men (n = NS) | 6 d | Glycemic diet + 40 g lingonberry powder Lipemic diet + 60 g lingonberry powder | Glycemic diet: 200 g yoghurt (lactose-free and fat-free non-flavored natural yoghurt + 50 g glucose) Lipemic diet: 200 g Yoghurt (lactose-free and fat-free non-flavored natural yoghurt + 35 canola oil) | Nullified the glycemic effect of the sugars present in the meals without affecting the postprandial lipemic response | [120] |
| 11 | CO, DB, RCT | 13 Healthy, over-weight, non-smoking male and female volunteers | Single meal challenge | 100 g lingonberry | Hyperlipidic and hypercaloric meals (38 kcal% fat) | Reduced glycemic response, rarified the increase of cholesterolemia | [121] |
| 12 | RCT | Normal, healthy subjects (n = 9) | 12 wk | LB polysaccharides + fibers (2 g/Kg of oat bread) | Oat bread | In reduced glucose and C-peptide response | [122] |
| 13 | SB, CO, RCT | Healthy women volunteers (n = 20) | 2-h meal tests | Diet 1: 150 g whole LB puree containing 35 g sucrose per day Diet 2: 300 mL LB nectar (equal to 150 g fresh berries) containing 35 g sucrose | NA | Optimized postprandial metabolic responses to sucrose with delayed digestion and absorption of sucrose/glucose | [123] |
| 14 | RCT | Healthy non-smoking males (n = 14) | 2-h meal tests | 60 g of LB juice press residue corresponding to 270 g of fresh LB with standard diet | Standard diet: white wheat bread, cucumber, water, and a banana | Gut microfloral metabolism of polyphenols resulting in increased levels of hippuric acid and 4-hydroxyhippuric acid | [124] |
| (I) Blackberries (BBR) (Animal studies) | |||||||
| 1 | RCT | Male Wistar rats (n = 32) | 5 wk | Anthocyanin-enriched fraction (AF) and Ellagitannin-enriched fraction (EF) equivalent to (4 mg cyanidin eq/kg BW) and 2.68 mg EA eq/kg BW respectively | NA | Reinforce the antioxidative defense system and lipid oxidation markers | [125] |
| 2 | RCT | C57BL/6 mice (n = 60) | 12 wk | BBR extracts at 200 mg/kg food BBR extracts: cyanidin-3-glucoside (51.24%), cyanidin-3-rutinoside (42.31%), and peonidin-3glucoside (6.91%) | HFD (45% kcal from fat) | Anti-inflammatory, anti-hypertensive, anti-hypercholesterolemia, antioxidative | [126] |
| 3 | RCT | Male DIO C57BL/6J mice (n = 40) | 12 wk | 6.3%, (w/w) BBR extracts in HFD provided ad libitum | HFD (45% kcal from fat) | Anti-obesity, Anti-inflammatory, anti-hypertensive, | [127] |
| 4 | RCT | Male Wistar rats (n = 24) | 17 wk | 25 mg/kg.BW BBR extracts in HFD provided ad libitum | HFD (45% kcal from fat) | Anti-obesity, anti-inflammatory, anti-dyslipidemic | [128] |
| 5 | RCT | Male diabetic Sprague-Dawley rats (n = 40) | 40 d | Microfiltrated 12.5–25% BBR juices | Reduced glycaemia (−10.4%), TG (−4.6%) and TC (21.0%), lipid peroxidation, attenuation of oxidative stress | [129] | |
| 6 | RCT | Male Wistar strain rats (n = 40) | 4 wk | Normal standard diet with 0.98% BBR polyphenols and 6% BBR fiber | Normal chow diet | Anti-inflammatory and anti-dyslipidemic | [130] |
| 7 | RCT | Female obese (BKS(D)-Leprdb/J72) and lean (C57BL/6J) mice (n = 24) | 10 wk | Aged or fresh BBR supplemented at 10% (w/w) of diet provided ad libitum | Normal chow diet | Increased in total beneficial bacterial population | [131] |
| 8 | RCT | Male C57BL/6J mice (n = 72) | 10 wk | Alcohol-free blueberry–blackberry fermented beverage (AFFB) a) AFFB [70% blackberry and 30% blueberry, 8.4 mg cyanidin-3-O-glucoside (C3G) eq./kg.BW)/day]; (b) dose 0.1 × ostamberlite extract (PAE), 1.1 mgC3G eq./kg BW/day; (c) dose 1 × PAE, 9.0 mg C3G eq./kg BW/day; (d) dose 2 × PAE, 18.9 mg C3G eq./kg BW/day | HFD (60.3% fat, 21.3% carbohydrate and 18.4% protein) | Reduced percent fat mass, mean adipocyte diameters, epididymal fat pad weights, and plasma TG and TC. | [22] |
| Human studies | |||||||
| 9 | RCT | Diabetic and obese men and women (n = 152) | 1 wk | Consumption of daily recommended amount of low glycemic index fruit (0.7–1.4 servings/day) | NA | Anti-dyslipidemic | [132] |
| 10 | open, single-center RCT | Healthy human subjects (n = 6) | 4 h | 200 mL of BBR juice equivalent to 400 mg of cyanidin equivalent/50 kg of body weight | NA | Improved plasma and urine antioxidant system | [133] |
| 11 | RCT | Dyslipidemic patients (n = 72) | 8 wk | 300 mL of BBR juice (equivalent to 316 mg/100 g polyphenols) of BBR with pulp every day | NA | Increased apo A-1 and HDL-C along reduction in apo B and hsCRP | [134] |
| (I) Strawberries (SB) (Animal studies) | |||||||
| 1 | RCT | Diabetic male albino Wistar rats (n = 36) | 4 wk | Aqueous, alcoholic and hydro-alcoholic SB extract (2 g/kg b.w.day | NA | Reduced expression level of genes involving glucose, lipid metabolism with improvement in glucose metabolism and liver function | [135] |
| 2 | RCT | Male Wistar rats (n = 20) | 12 wk | HFD supplemented with 0.2% irradiated/non-irradiated SB extracts | HFD (47.5% kcal from fat) | Reduction in the oxidative damage in brain and peripheral tissues | [136] |
| 3 | RCT | Male C57BL/6J mice (n = 36) | 24 wk | HFD supplemented with 2.6% freeze-dried SB | HFD containing approximately 20% higher in energy density compared to the low-fat diets | Reduction in the HFD led increase of FBS, adhesion molecule-1, leptin, E-selectin, resistin, and plasminogen activator protein-1 | [137] |
| 4 | RCT | Male Wistar rats (n = 48) | 8 wk | Supplementation of the diet with a 6% w/w (equivalent to a 5 g/kg 65 BW dose) of a F/D SB-BlB (5:1) powder (FDSB) | High-fat-sucrose diet (D12451, Research Diet) | Anti-obesogenic and anti-inflammatory effects | [138] |
| 5 | RCT | Male Wistar rats (n = 24) | 16 wk | AIN93-modified diet with lyophilized SB extract at 10 g/kg of diet | AIN93-modified diet | Improvement of oxidative stress biomarkers, mitochondrial performance, antioxidant enzyme activities, reduction of DNA damage and ROS concentration | [139] |
| 6 | RCT | Male Wistar rats (n = 20) | 12 wk | Supplementation of 0.2% SB | HFD (47.5% calories from fat) | Antioxidative, anti-stress | [140] |
| 7 | RCT | German Landrace pigs (n = 48) | 4 wk | 205–745 g of SB with normal feed per day | Linseed oil (15 g/day) enriched feed | Anti-stress and antioxidative | [22] |
| 8 | RCT | db/db mice homozygous for the diabetes spontaneous mutation (Leprdb) with C57BL/6J background (n = 24) | 10 wk | 2.35% F/D SB powder in the diet pellets (w/w) (equivalent to two human servings of SB i.e., ~160 g SB) | NA | Increased Bacteriodetes to Firmicutes ratio | [141] |
| 9 | RCT | Male CD-1 mice (n = 60) | 8 wk | 5% (w/w) of diet freeze-dried whole SB powder | AIN93G diet | Increased Bacteriodetes to Firmicutes ratio | [142] |
| (Human studies) | |||||||
| 10 | DB, RCT, parallel study | Insulin resistant and obese males and females (n = 41) | 6 wk | Beverage containing 1·84 g of a mixture of dry SB and CrB providing 333 mg of polyphenols on daily basis (also equivalent to 112 g consumption of fresh berry fruit) | NA | Improved insulin sensitivity and release | [143] |
| 11 | CO, SB, PC, RCT | Hyperlipidemic men and women (n = 24) | 12 wk | SB beverage containing 10 g/serving of freeze-dry SB powder providing 338 mg of polyphenols daily (also equivalent to 110 g consumption of fresh berry fruit) | HFD consisting of typical breakfast food items (i.e., bagel, cream cheese, whole milk, egg, margarine, cantaloupe) | Reduced postprandial lipemia and oxidative stress markers | [144] |
| 12 | CO, RCT | Healthy males and females (n = 30) | 5 d | 20 g of five types SB jams each with sugar of different glycemic index | 60 g white bread slice | Non-significant reduction in the postprandial glucose level | [145] |
| 13 | CO DB RCT | Healthy males and females (n = 16) | 3 wk | 60 g of three types SB jams each with sugar of different glycemic index and polyphenolic contents | Strawberry jam with high sugar level produced less levels of FFA. | [146] | |
| 14 | DB RCT | T2DM males and female subjects (n = 36) | 6 wk | Two cups of F/D SB beverage containing 25 g × 2 = 50 g | NA | Reduction in LDL-C and LDL-C/TC and LDL-C/HDL-C ratio | [147] |
| 15 | SB, CO parallel, RCT | Obese and overweight men and women (n = 24) | 6 wk | SB beverage containing 10 g/serving of freeze-dry SB powder providing 96 mg of polyphenols on daily (also equivalent to 100 g consumption of fresh berry fruit) | High-carbohydrate-fat diet | Attenuation of diet-induced inflammatory markers | [148] |
| 16 | Single-center, CO, SB, PC, | Men and women (n = 26) | OTCS | SB Milk based beverage containing 10 g/305 mL of F/D SB powder | high-carbohydrate, moderate-fat meal (HCFM) | Reduced postprandial insulin and inflammatory response | [149] |
| 17 | Four-arm, SB, PC, CO, RCT | Males and females with insulin resistance (n = 23) | NS | SB milkshake containing 10–40 g freeze-dried SB powder where 10 g freeze dried powder = 110 g fresh strawberries | Standard western type meal | Reduced lipid oxidation and post-meal insulin demand | [150] |
| 18 | Observatory study | Healthy men and women (n = 247) | 20 years | Dietary flavonoids intake (47–560 mg/day) from fruits and berries | - | Flavonoid Compounds in Driving Patterns of Microbial Community Assembly | [151] |
| 19 | RCT | Obese men and women (n = 66) | 12 wk | SB beverage containing 25–50 g freeze-dry SB powder daily | HFD (50% calories from fat) | Increased the glutathione level, serum catalase activity, and plasma antioxidant capacity | [152] |
| 20 | DB RCT | T2DM patients (n = 40) | 6 wk | 50 g of freeze-dried SB powder (equivalent to 500 g fresh strawberries) each day | NA | Reduction in the markers of lipid peroxidation (MDA), inflammatory markers (CRP). Reducing trend in HbA1c. | [153] |
| (I) Goji berries (GB) (Animal studies) | |||||||
| 1 | RCT | Alloxan-induced hyperglycemic/hyperlipidemic adult rabbits (n = 35) and male mice (n = 24) | 10 d | Water decoction (0.25 g/kg BW day), crude GB polysaccharides (10 mg/kg BW day), and purified GB polysaccharides (10 mg/kg BW day) | NA | Hypoglycemic and hypolipidemic effect with increased plasma antioxidant capacity | [154] |
| 2 | RCT | Male Wistar rats (n = 70) | 8 wk | Ethanolic and aqueous GB extracts at 50 mg/kg b.w. or 100 mg/kg BW daily | HFD | Significantly reduced liver damage and oxidative changes | [155] |
| 3 | RCT | Diabetic male mice of original Kun-ming strain (n = NS) | 4 wk | GB polysaccharides (20–40 mg/kg BW day) orally | NA | Hypoglycemic and hypolipidemic | [156] |
| 4 | randomized block design | Obese male Sprague-Dawley rats (n = 60) | 8 wk | GB anthocyanins at 50–200 mg/kg BW.day | HFD | Reduced body-weight-gain with anti-inflammatory properties | [157] |
| 5 | RCT | STZ-diabetic Male Wistar rats (n = NS) | 8 wk | GB polysaccharides (10 mg/kg, BW.day) | NA | Increased antioxidative scavenging and antioxidant enzymes. Increased activity of protein kinase C (PKC) | [158] |
| 6 | RCT | STZ-induced diabetic Sprague-Dawley male rats (n = 60) | 8 wk | Water decoction of GB (5 g/kg.BW.day) | NA | Protective effects in diabetic retinopathy | [159] |
| 8 | RCT | Male Wistar rats (n = 16) | 4 wk | GB polysaccharides 10 mg/kg BW.day dissolved in physiological saline | High-fat-sucrose diet | hypoglycemic and improving hyperinsulinemia | [160] |
| 9 | RCT | Diabetic male C57BL/6J mice (n = 48) | 7 wk | GB polysaccharides 100–500 mg/kg BW.day by gastric perfusion | HFD | Hypoglycemic effects with increased insulin-sensitizing, glucose metabolism, insulin secretion, and promoting pancreatic cell proliferation. | [26] |
| 10 | RCT | Swiss Albino rat (n = 30) | 3 wk | Water-soluble polysaccharides (galactomannan) 250–500 mg/kg BW.day by oral gavage | NA | Hypolipidemic, reduced lipid oxidation, increased insulin-sensitizing and serum antioxidant level | [161] |
| 11 | RCT | Diabetic Wistar rats (n = 48) | 8 wk | Water-soluble GB polysaccharides 250–500 mg/kg BW.day by oral gavage | HFD and HCD (12% protein, 5% fat, 67% carbohydrate, 5% cholesterol, and 5% other additives) | Reduced serum level of IL-2, IL-6, TNF-α, IFN-α, MCP-1, and ICAM-1 with increased activities of SOD and GSH-Px activities | [162] |
| 12 | RCT | Postnatal Royal College of Surgeons (RCS) rats (n = 60) | 4 wk | Whole GB powder 1 mg/kg of per day | NA | Reduced Caspase-2 activity in experimental group at 25th post-neonatal day | [163] |
| 13 | RCT | Male IL-10-deficient mice (n = 14) | 10 wk | Diet supplemented with 1% GB | Normal diet | Increased gut population of SCFA producing bacteria | [164] |
| Human studies | |||||||
| 14 | RCT | Kunming mice of clean grade (n = 14) | 2 wk | GBPS at a dose of 0.1 mL/10 g body weigh daily via intragastric administration | Normal diet | Increased gut population of SCFA producing bacteria, Firmicutes, Akkermansia, Lactobacillus, and Prevotellaceae | [165] |
| 15 | DB, PC, RCT | Healthy males and females (n = 50) | 30 d | Intake of 120 mL of GB juice (equivalent to 1632 mg/ daily serving (120 mL) of goji berry polyphenols | Traditional Chinese diet rich in carbohydrate | Increased serum levels of glutathione peroxidase (GSH-Px) and superoxide dismutase (SOD) with reduced level of MDA | [166] |
| 16 | RCT | Metabolic syndrome patients (n = NS) | 45 d | 14 g of GB with meals | Normal diet | Reduction in transaminases, waist circumference with improvements in lipid profile, glutathione and catalase level. | [167] |
| 17 | RCT | Male and female C57BL/b6N mice (n = 56) | 8 wk | GB polysaccharides (1–10 mg/kg BW day) orally | NA | Increased hepatic antioxidant enzymes, y inhibited cytochrome P450 2E1, nitric oxide metabolism and lipid peroxidation | [168] |
| 18 | DB, CO, RCT | healthy overweight men (n = NS) | Single meal challenge | meal containing 25 g of dried GB fruit | Ready-made meal with a fixed macronutrient composition (30–40% fat, 40–50% carbohydrates, and 13–16% proteins) | No-single-dose-effect on substrate oxidation and prospandial-energy-expenditure | [169] |
| (I) Acai berries (AB) (Animal studies) | |||||||
| 1 | RCT | Male mice of the C57BL/6 strain (n = NS) | 12 wk | AB seed extracts 300 mg/kg.BW.day by intragastric gavage | HFD (60% calorie from fat) | Reduced expressions of lipogenic proteins (SREBP-1c, pACC, ACC, HMG-CoA reductase) with increased expression of pAMPK, pACC/ACC, and cholesterol transporters (ABCG5 and ABCG8) | [170] |
| 2 | RCT | Zebrafish (n = 70) | 5 wk | HC diet supplemented with10% w/w of AB puree powder | high cholesterol (HC) diet (47.5% crude protein, 6.5% crude fat, 4% cholesterol, 2.0% crude fiber, 10.5% crude ash) | Reduced oxidative markers with lipid lowering effects | [171] |
| 3 | RCT | Oxidatively damaged sod1/sod1 mutant strains Drosophila melanogaster (n = 120) | 5-6d | AB supplemented sugar-yeast (SY) medium to a final concentration of 0.25%, 0.5%, 1% or 2% (w/v) of the food | SY medium | Increased transcript level of gluconeogenesis gene phosphoenolpyruvate carboxykinase (Pepck) with reduction in oxidative stress | [172] |
| 4 | RCT | ApoE-deficient (ApoE 2/2) male mice (n = 23) | 12 wk | AIN-93M diet formulated to contain 2% F/D açai’ pulp + exercise in progressive treadmill for 30 min daily at a speed of 12 m/min, 0% incline | AIN-93M diet | Hepatic superoxide dismutase activity, mRNA expression of monocyte chemotactic protein-1, percentages of hepatic lipid droplets | [173] |
| 5 | RCT | STZ-induced diabetic Male Wistar rats (n = NS) | 45 d | AB seed extracts 200 mg/kg.BW.day in drinking water | NA | Reduced oxidative damage by reducing the expression of caspase-3, IL-6, TNF-α and MCP-1 | [174] |
| 6 | RCT | Female Fischer rats (n = 32) | 6 wk | Hypercholesterolemic diet (25% soy oil and 1% cholesterol) supplemented with 2% AB (dry wt/wt) | Hypercholesterolemic diet (25% soy oil and 1% cholesterol) | Reduced expression of cholesterol biosynthesis genes HMG CoA-R, EBP-2, ApoB100, LDL-R, ABCG8, and CYP7A1 | [175] |
| 7 | RCT | STZ-induced diabetic Male Wistar rats (n = 40) | 9 wk | AB seed extracts 200 mg/kg.BW.day by intragastric gavage | HFD (55% calorie from fat) | Hypoglycemic and hypolipidemic with reduced expression of TNF-α and activating the insulin-signaling pathway in muscle and adipose tissue | [176] |
| 8 | RCT | Diabetic female Fisher rats (n = NS) | 30 d | Standard AIN-93 diet supplemented with 2% (w/w) AB pulp | AIN-93 | Modulate ROS production by neutrophils and improve the liver oxidant/antioxidant balance | [177] |
| (Human studies) | |||||||
| 9 | CO, DB, RCT | Overweight healthy males (n = 23) | Single day meal challenge | Frozen AB pulp (150 g) was prepared in a smoothie with 50 g banana | 50 g banana and matched for fat with 1.5 g hexadecanoic acid [palmitic acid (16:0)] and 8.5 g sunflower oil [30% (9Z)-Octadec-9-enoic acid (oleic acid [18:1]), 60% (9Z,12Z)-9,12-Octadecadienoic acid (linoleic acid [18:2]), and 10% palmitic acid (16:0)], | Lower incremental area under the curve (iAUC) for total peroxide oxidative status after açai and increased the iAUC for insulin | [178] |
| 10 | RCT | Male Swiss mice (n = 32) | 12 wk | A single daily dose freeze-dried AB pulp (3 g/kg) via gavage | HFD (32% lard and 1% cholesterol) | Attenuated hepatic steatosis and reduced lipid accumulation | [179] |
| 11 | four-way CO | Healthy men and women (n = 11) | Single dose study | 100% clarified AB juice/pulp 7 mL/kg BW of each study | NA | Increased plasma antioxidant capacity without affecting generation of reactive oxygen species, and uric acid concentrations in plasma | [180] |
| 12 | open label pilot study | Overweight adults (n = 10) | 30 d | Intake of 100 g AB pulp twice daily | NA | Postprandial increase in the AUC of plasma glucose with reduced TC, LDL-C, and LDL-C/HDL-C | [181] |
| (I) Chokeberries (CB) (Animal studies) | |||||||
| 1 | RCT | C57BL/6JmsSlc and KK-Ay male mice (N = 10, EACH GROUP) | 4 wk | CB provided ad libitum | Normal chow diet | Duction of glucose-dependent insulinotropicpolypeptide (GIP) level | [182] |
| 2 | RCT | STZ-induced-diabetic-male ICR mice (n = 32) | 4 wk | CB extract (10–100 mg/kg.BW) daily administered orally | NA | Hypoglycemic, hypolipidemic, antioxidative | [183] |
| 3 | RCT | C57BL/6N mice (n = 20) | 12 wk | CB powder dissolved in water (50 mg/kg daily) | HFD (60 kcal% Lard) | Reduced the body and liver weight, lipid accumulation, PPARγ2, FAS, hepatic TG and leptin. Serum transaminases, indicators for liver antioxidant capacity were significantly increased. | [184] |
| 4 | RCT | Male C57BL/6J (n = 60) | 8 wk | CB extracts (100 mg/kg.BW) dissolved in 0.5% carboxymethyl cellulose | HFD (containing 60% kcal fat) | Attenuated weight-gain, increase in serum TG, TC, LDL-C and better glucose tolerance | [185] |
| 5 | RCT | Male Wistar rats (n = NS) | 6 wk | Aronia melanocarpa fruit juice (AMFJ) at doses 10 and 20 mL/kg | NA | Hypoglycemic, hypolipidemic | [186] |
| 6 | RCT | Polish Merino lambs (n = 24) | 12 wk | 150-300 g of chokeberry pomace per each kg of the complete feed mixture | Complete feed mixture | Hypoglycemic, hypolipidemic | [187] |
| 7 | RCT | Middle-aged non-medicated subjects with MS (n = 38) an healthy volunteers (n = 14) | 8 wk | CB extracts 100 mg/kg.BW three times daily | NA | Beneficial changes in lipid profile, coagulation parameters, inhibition of platelet aggregation | [188] |
| 8 | RCT | Male Wistar rats (n = 24) | 4 wk | Diet was supplemented by the extract from CB fruits (0.2% W/W) added at the expense of corn starch | Standard casein diet enriched with 0.5% of cholesterol. Exp group: the diets were modified by 8% of lard and 65% of fructose added at the expense of soybean oil and maize starch, | Maltase and sucrase, e improvement of antioxidant status, cholesterol-lowering, | [189] |
| 9 | RCT | Male Wistar rats (n = NS) | 4 wk | CB juice 10 mL/kg.BW.day | NA | Hypoglycemic, hypolipidemic, antioxidative | [190] |
| 10 | RCT | Male Wistar rats (n = 72) | 8 wk | CB juice 50 mL/kg.BW.day | High-carbohydrate, high-fat + purple maize flour (HPM) | Reduced Inflammatory cell infiltration, visceral adiposity index, total body fat mass, improved glucose tolerance | [191] |
| 11 | RCT | Male Wistar rats (n = 36) | 6 wk | CBE at 100 or 200 mg/kg BW.day | Fructose rich diet containing (g/kg diet): casein, 207; DL-methionine, 3·0; fructose, 600; lard, 50; cellulose, 79·8; | Elevated plasma adiponectin levels and inhibited plasma TNF-α and IL6. Increased in the expression level of glucose and lipid metabolizing genes | [192] |
| 12 | RCT | Male Wistar albino rats (n = 60) | 4 wk | Standardized Aronia extract (SAE) 0.45 mL/kg.BW day) for 4 weeks | HFD (25% fat, 15% protein, 51% starch, and 5% fiber) | Reduced serum level of TC, TG, LDL-C, with increased serum levels of SFA and PUFA. | [193] |
| (Human studies) | |||||||
| 13 | CO open-label trial | T2DM patients (n = 35) | 12 wk | Oral CB juice supplementation (150 mL/day, three times a day for 50 mL) | NA | Significantly improved the renal /hematological and lipid parameters (TG, TC, LDL-C, LDL-C/HDL-C) in diabetic patients | [194] |
| 14 | RCT | Healthy female volunteers (n = 29) | 12 wk | 100 mL of polyphenol-rich organic CB juice per day | NA | Reduced TBARS, pro-oxidantantioxidant balance, increase in paroxonase-1 activity | [195] |
| 15 | RCT | Apparently healthy women (n = 25) | 12 wk | Consume 100 mL of polyphenol-rich organic CB juice daily | NA | Increased SOD and GPX activities, C22:6n-3, PUFAs, total PUFAs and unsaturation index and decrease in n-6:n-3 ratio | [196] |
| 16 | RCT | Healthy volunteers and 25 patients with metabolic syndrome (n = 22) | 8 wk | CB extract (3 × 100 mg/day) | NA | Improvement in serum lipids, and oxidative status (GSH-Px, SOD, TBARS) | [197] |
| 17 | RCT | Healthy subjects (n = 33) | 4 wk | Consume 200 mL of polyphenol-rich organic CB juice daily (containing 386 ± 9.7 mg of total phenolics expressed as gallic acid equivalents per 100 g) | NA | Positive effects on BP and lipid status in hypertensive subjects | [198] |
| 18 | RCT | Diabetic Wistar white male rats (n = 48) | 16 wk | dose of polyphenols extracts 0.040 g/kg BW every 2 day | NA | Reduced TNF-α and IFN-γ levels | [199] |
| 19 | RCT | Healthy, non-smoking volunteers (n = 11) | 3 wk | CrB juice between meals (250 mL per day) (560 mg GAE/100 mL) | NA | Increased serum antioxidant capacity with no significant change in the blood lipid profile | [200] |
| 20 | RCT | Men with the diagnosed mild hypercholesterolemia (n = 58) | 6 wk | CB juice between meals (250 mL per day) (560 mg GAE/100 mL) | NA | Improved lipid profile with reduced lipid peroxides (LPO), C-reactive high sensitivity protein (hsCRP), homocysteine, | [201] |
| 21 | 3-arm, DB, parallel RCT | Healthy male volunteers (n = 66) | 12 wk | CB extract” capsules (containing 116 mg total (poly)phenols). CB whole fruit” capsules (containing the equivalent to 10 g of the whole CB fruit, and 12 mg of total (poly)phenols) | NA | Increased Anaerostipes, Bifidobacterium, Faecalibacterium, and Clostridium genera | [202] |
| (I) Black Currants (BCT) (Animal studies) | |||||||
| 1 | RCT | Old male Sprague-Dawley (SD) rats (n = NS) | Single meal challenge test | BCE 5 mg/kg.BW (1 mg D3R/kg.BW) | Normal diet with IP administration of glucose solution (2 g/kg) | Improved hyperglycemic and hypoinsulinemic condition | [203] |
| 2 | RCT | Male KK-Ay (n = 16) | 7 wk | BC extracts (2 g/Kg.diet) (equivalent to delphinidine-3-glucoside (D3R) 2 g/Kg.diet) | NA | Improved glucose tolerance with increased GLP-1 concentration, and upregulation of AMPK-α and prohormone convertase 1/3(GLP-1 precursor) | [204] |
| 3 | RCT | Male C57BL/6J mice (n = 48) | 8 wk | Diet supplemented with 1% BC powdered extract (32% anthocyanins) | HFD (60 kcal% fat diet) | Protective effect of BC anthocyanins against obesity and associated insulin resistance. | [205] |
| 4 | RCT | Male C57BL/6J mice (n = 24) | 12 wk | HF/HC diet supplemented with 0.1% of BCE (containing 25% anthocyanins and 40% polyphenols) by weight | AIN-93M high fat/high cholesterol (HF/HC) diet (16% fat, 0.25% cholesterol by weight; 55.7%, 125.5% and 31.8% energy from carbohydrate, protein and fat, respectively; 4529 kcal/Kg | Reduced BW and adipocyte size of the epididymal fat, energy expenditure and mitochondrial biogenesis genes | [206] |
| 5 | RCT | Male New Zealand white rabbits (n = 20) | 4 wk | Diet supplemented with 1.5% BC polyphenolic extract | HFD (10% lard) was 17% from protein, 32% from fat and 51% from carbohydrates | Reduced concentration of putrefactive metabolites, β-glucuronidase activity, ameliorated hyperlipidemia, and antioxidative capacity | [207] |
| 6 | RCT | Sprague–Dawley male rats (n = 40) | 4 wk | 2 mL of BC extract (containing 30 mg BC /kg BW) or 2 mL of CAM30 extract (containing 13.4 mg CAM30/kg body weight), respectively, three times weekly by oral gavage | NA | Reduced β-glucuronidase activity and undesirable bacteria in the caeca. Increased lactobacilli and bifidobacterial gut species | [208] |
| 7 | RCT | Male Sprague-Dawley (SD) rats (n = 40) | 8 wk | BC extract 100–300 mg/kg.BW.day administered orally | High-fructose (HF) diet (60% fructose diet) | Improvements in hypertension, dyslipidemia, insulin resistance, and obesity | [209] |
| 8 | RCT | Male Sprague-Dawley rats (n = 128) | 6 wk | Diets with dietary fiber and BC extracts (Currantex 30) (containing total anthocyanin 32% (w/w)) | NA | Increased intestinal population of SCFA and total beneficial bacterial population | [210] |
| (Human studies) | |||||||
| 9 | RCT | Healthy volunteers (n = 30) | 2 wk | BC extracts (1500 mg/day; 375 mg × 4 capsules) BC powder CAM30 (672 mg/day; 168 mg × 4 capsules) | NA | Increased intestinal population of SCFA and total beneficial bacterial population | [211] |
| 10 | DB, CO, RCT | Healthy subjects (n = 26) | Single meal challenge test | Apple and BC polyphenol-rich drinks (1200 mg apple polyphenols (AE), or 600 mg apple polyphenols + 600 mg BC anthocyanins (AE+BE)) | Standardized high-carbohydrate meal 100 g of white bread | Reduced Postprandial insulin, C-peptide and GIP, GLUT and SGLT1-mediated glucose transport | [212] |
| 11 | DB, CO, RCT | Healthy subjects (n = 22) | Single meal challenge test | Low-sugar-BC drink containing 300–600 mg anthocyanins | Standardized high-carbohydrate meal 100 g of white bread | Reduced postprandial insulinemia, glycemia, and incretin secretion | [213] |
| 12 | RCT | Healthy participants (n = 17) | 6 d | BC powder 6 g/day with water | NA | Improved postprandial AUC of glucose and insulin | [214] |
| 13 | DB, CO, RCT | Endurance-trained females (n = 16) | 7 d | BC extract 600 mg/day | NA | Increased fat oxidation | [215] |
| 14 | RCT | Healthy sedentary male and female participants (n = 40) | Single meal challenge test | BC juice 200 mL/participant | standardized meal bar to consume for breakfast at least 1 h prior to starting the trial. | Supported positive affective responses | [216] |
| 15 | parallel, four-arm, study design + DB, CO parallel trial | Healthy individuals (n = 24) (n = 32) | A single meal challenge study | Two opaque gelatin capsules containing BC anthocyanin (3.2 mg/kg total anthocyanins) | NA | Dose-dependent increase in plasma anthocyanins and recovery from exercise-induced oxidative stress | [217] |
| (I) Maqui berries (MqB) (Animal studies) | |||||||
| 1 | open exploratory study | Pre-diabetic volunteers (n = 43) | Single dose study | A single dose of Delphinol Capsules bearing either 60, 120, or 180 mg Delphinol on each day with one-week washout period | NA | Dose dependently lowered basal insulinemia and glycemia | [218] |
| 2 | RCT | Male balb/c mice (n = NS) | 7 d | MqB extracts (25, 50 and 100 mg/kg.BW) | NA | Ameliorate the oxidative stress condition | [219] |
| 3 | RCT | Male C57BL/BJ mice | 12 wk | MqB anthocyanins (ANC), Labrasol/water: 66/34 + ANC (LAB + ANC) | HFD (60% calories from fat) | Decreased glucose production, down-regulation of gluconeogenic enzyme | [220] |
| 4 | DB, CO, RCT | Fifty overweight volunteer smokers (n = 42) | 4 wk | 3 capsules of 150 mg standardized maqui berry extract containing 54 mg of anthocyanin daily (equivalent to 162 mg anthocyanins/day) | NA | Reduced levels of Ox-LDL in the anthocyanin group | [221] |
| 5 | RCT | Male C57BL/6Nhsd mice (n = 18) | 4 wk | MqB derived Delphinidine (15 mg/kg.BW) daily | High-fat diet and high-carbohydrate drinking water (45% kcal from fat) | Reduced TG accumulation with no effect on metabolic alterations related glucose metabolism | [222] |
| 6 | Prospective observational study | Middle-aged participants (n = 21) | 8 wk | Two tablets per day of an MCN (Eonlipid) (containing maqui, 300 mg in each tablet) | NA | Improvement of most atherogenesis and oxidative stress biomarkers | [223] |
| 7 | CO, RCT | Healthy male subjects (n = 11) | ONCS | Intake of 250 mL of the MqB drink containing an number of total polyphenols ~1000 µmol equivalents of gallic acid | Meals containing food-grade glucose and rice, containing 50 g of carbohydrates by each meal | Reduced glycemic indexed for high-carbohydrate diets. | [224] |
| 8 | RCT | C57BL/6J littermates’ male mice (n = 23) | 16 wk | HFD supplemented with 4–5 mg of MqB polyphenols/ 10–15 kcal per day | HFD (45% calories from fat) | Reduced body-weight-gain, improved glucose tolerance and insulin resistance. Differential expression of genes involved in fatty acid oxidation, de novo lipogenesis, thermogenesis, and multilocular lipid droplet formation | [225] |
| Compounds | Bilberry (mg/100 g fw) | Blueberry (mg/100 g fw) | Cranberry (mg/100 g fw) | Raspberry (mg/100 g dw) | Mulberry (mg/100 g fw) | Lingonberry (mg/g DE) | Blackberry (mg/g DE) | Strawberry (mg/100 g fw) | Goji Berry (mg/100 g dw) | Acai Berry (mg/100 g dw) | Black Chokeberry (mg/100 g dw) | Black Currant (mg/100 g fw) | Maqui Berry (mg/100 g fw) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| References | [38,51] | [10,35,36,37,38,39] | [226,227,228,229] | [230,231,232] | [233,234,235] | [236,237] | [131,238,239] | [240,241] | [62,123,242,243] | [244,245,246] | [247] | [248,249] | [219,250,251] |
| Cyanidin | 18–290 | - | - | - | - | - | - | 27–175 | 27.5 | - | - | - | 22.8–26.0 |
| Delphinidin | 29–280 | - | - | - | - | - | - | - | - | - | - | - | 105.0–120.3 |
| Quercetin | 1.5–8 | 0.07 * | 104 | - | 0.3–10.04 | - | - | 0.09–0.54 | - | 39.02 | 37-400 | - | - |
| Myricetin | nd-3 | - | 69 | - | - | - | - | 0.05–0.77 | - | - | - | - | - |
| p-Coumaric acid | 1–9 | - | 0.25 | 67.03–2792.6 | 0.3–4.2 | 0.13 | - | 2.64 | 0.07–0.22 | - | - | - | - |
| m-Coumaric acid | 7–30 | - | - | - | 0.3–14.2 | - | 0.93 | - | - | - | - | - | - |
| Sinapic acid | - | - | 0.211 | - | - | - | - | 0.61 | - | - | - | - | - |
| Gallic acid | - | - | - | 3–72.2 | 3.8–8.6 | - | - | 26.5–47.54 | - | 701.6 | - | - | 75 |
| Ascorbic acid | - | - | 0.011 | 2.4–5.34 | - | - | - | - | - | - | - | - | - |
| Ferulic acid | - | - | 0.087 | - | 5.3–294 | - | - | 0.95 | 753.6 | 2.46 | - | - | - |
| Chlorogenic acid | - | 3.08 * | - | 177.4 | 4.3–22.3 | - | - | 0.35–1.10 | - | 37.65 | - | - | - |
| Protocatechuic acid | - | - | - | - | 3 | - | - | - | - | - | - | - | - |
| 5-O-Caffeoylquinic Acid | - | - | - | - | 283–1735 | - | 5.57–8.88 | - | 8.4–37.9 | - | 346–413 | - | - |
| 1,3-di-O-Caffeoylquinic Acid | - | - | - | - | 0.2–0.3 | - | 0.15–0.22 | - | 0.6–4.27 | - | 13–508 | - | - |
| Caffeic acid | - | - | 0.15 | 2.41–5.31 | 1.3–9.2 | 0.26 | - | 0.52 | 0.76–1.52 | 8.12 | - | - | - |
| Protocatechuic acid | 4–8 | - | - | - | - | - | - | - | - | - | - | - | - |
| Ellagic acid | - | - | 120 | 1151.7 | 23.9 | - | 2.012 | 2.72 | - | - | - | - | - |
| Benzoic acid | - | - | 4.7 | - | - | 3.79 | - | - | - | - | - | - | - |
| p−Hydroxyphenylacetic acid | - | - | 0.007 | - | 4.3–12.9 | - | - | - | - | - | - | - | - |
| 2,3-Dihydroxybenzoic acid | - | - | 0.003 | - | 12.9 | - | - | - | - | 28.18 | - | - | - |
| 2,4-Dihydroxy benzoic acid | - | - | 0.04 | - | - | - | - | - | 0.13–0.51 | 3.37 | - | - | - |
| Vanillic acid | - | - | 0.05 | 3–4.41 | - | - | - | 2.91–3.1 | 4–6.37 | 57.7 | - | - | - |
| Trans-cinnamic acid | - | - | 0.02 | - | - | - | - | - | - | - | - | - | - |
| O-Hydroxycinnamic acid | - | - | 0.089 | - | - | - | - | - | - | - | - | - | - |
| p-Hydroxybenzoic acid | - | - | 0.021 | - | - | - | - | - | - | 172 | - | - | - |
| Resveratrol | 1–12 | - | - | - | - | 0.13 | - | - | - | - | - | - | - |
| Epigallocatechin | - | - | 1.5 | - | 25.6 | - | - | - | - | - | - | - | - |
| (+/−)-Catechins | 6–7 | - | 4.5 | 129.3 | - | - | - | 19.56–135.19 | 106.6 | 49.1 | 593 | - | - |
| (+/−)Epicatechin | 6–7 | - | 4.5 | 791.7 | 0.2–24 | - | - | 1.07 | - | 44.6 | 6767 | - | - |
| Gallocatechin gallate | - | - | 0.4 | - | 10.2–63.7 | - | - | - | - | - | - | - | - |
| Epigallocatechin gallate | - | - | 1.9 | - | 4.5–8.4 | - | - | 5.65 | - | - | - | - | - |
| Delphinidin 3-galactoside | 167.1 | 23.4 | - | - | - | Up to 1.25 | - | - | - | - | - | 52 *** | - |
| Delphinidin 3-glucoside | 169.1 | 15.4 | - | - | - | 26.8–29.40 | - | - | - | - | 839 *** | 389.9 | |
| Cyanidin 3-galactoside | 122.6 | 4.2 | 2 | - | - | - | - | - | - | 105–2407 | - | - | |
| Delphinidin 3-arabinoside | 152.3 | - | - | - | - | - | - | - | - | - | - | - | |
| Cyanidin 3-glucoside | 130.4 | 2.6 | 0.1 | 15.02–53.94 | 4.72 | - | - | - | 998.74 | 5–113 | 327 ** | 679 | |
| Petunidin 3-galactoside | 50 | 11.7 | - | - | - | - | - | - | - | - | 103 *** | - | |
| Cyanidin 3-arabinoside | 110.6 | 3.5 | 1.4 | - | - | - | - | - | - | 215–1148 | - | - | |
| Petunidin 3-glucoside | 101.9 | 12.4 | - | - | - | 10.02–15.25 | - | - | 21 | - | - | - | |
| Peonidin 3-galactoside | 13.3 | 1.8 | 2.8 | - | - | - | - | - | - | - | - | - | |
| Petunidin 3-arabinoside | 23.9 | 9.3 | - | - | - | - | - | - | - | - | - | - | |
| Peonidin 3-glucoside | 56.7 | 2.1 | 0.3 | - | - | 2.04–3.62 | - | - | 193 | - | 71 *** | - | |
| Malvidin 3-galactoside | 27.5 | 34.9 | - | - | - | - | - | - | - | - | - | - | |
| Peonidin 3-arabinoside | 4.5 | 1 | 1.1 | - | - | - | - | - | - | - | - | - | |
| Malvidin 3-glucoside | 67.7 | 31.2 | - | - | - | 9.49–10.57 | - | - | - | - | - | - | |
| Malvidin 3-arabinoside | 12.8 | 34.7 | - | - | - | - | - | - | - | - | - | - | |
| Quercetin-3-galactoside | - | - | 70.4 | - | - | - | - | - | - | - | - | - | |
| Quercetin-3-α-arabinopyranoside | - | - | 34.4 | - | - | - | - | - | - | - | - | - | |
| Quercetin-3-rhamnoside | - | - | 41.6 | - | - | - | - | - | - | - | - | - | |
| Kaempferol-3-glucoside | - | - | 5.6 | - | - | - | 5.12–17.67 | - | - | - | - | - | |
| Myricetin 3-α-arabinofuranoside | - | - | 37.5 | - | - | - | - | - | - | - | - | - | |
| Quercetin 3-O-glucuronide | - | - | - | 717.57 | - | - | 9.4–39 | - | - | - | - | - | |
| Quercetin pentoside | - | - | - | 252 | - | - | - | - | - | - | - | - | |
| Cyanidin-3-O-sophoroside | - | - | - | 43.27–800.3 | - | - | - | - | - | - | - | - | |
| Cyanidin-3-O-rutinoside | - | - | - | 5.49–104.58 | 2.73 | - | - | - | 433.98 | - | 1693 *** | - | |
| Pelargonidin-3-glucoside | - | - | - | - | 0.14 | - | - | - | - | - | - | - | |
| Quercetin 3-O-rutinoside | - | - | - | - | 192–398 | - | - | 0.9–23.2 | - | - | 1.8–2.37 | - | |
| Quercetin 3-O-galactoside | - | - | - | - | 0.2–345 | - | - | - | - | - | - | - | |
| Quercetin 3-O-glucoside | - | - | - | - | 72.4–345.7 | 0.23–0.88 | 9.8–25.1 | 16.9–90.9 | 44–3756 | - | 1.5–2.0 | - | |
| Kaempferol 3-O-glucoside | - | - | - | - | 35.5–478 | - | 5.96–14.39 | 0.5–1.94 | - | - | - | - | |
| Pelargonidin 3-O-rutinoside | - | - | - | - | 17.8–290 | - | - | - | - | - | - | - | |
| Delphinidin-O-(pentosyl)hexoside | - | - | - | - | - | - | 0.82–1.88 | - | - | - | - | - | - |
| Delphinidin-O-rhamnoside | - | - | - | - | - | - | 2.14 | - | - | - | - | - | - |
| Malvidin-O-pentoside | - | - | - | - | - | - | 1.08–2.13 | - | - | - | - | - | - |
| Malvidin-O-rhamnoside | - | - | - | - | - | - | 0.13–0.63 | - | - | - | - | - | - |
| Caffeoylisocitrate | - | - | - | - | - | - | 0.35 | - | - | - | - | - | - |
| Caffeic acid-O-hexoside | - | - | - | - | - | - | 0.4–0.56 | - | - | - | - | - | - |
| Myricetin-O-hexoside | - | - | - | - | - | - | 0.19–0.29 | - | - | - | - | 29 | - |
| Pelargonidin-3-glucoside | - | - | - | - | - | - | - | 17.82–20.85 | - | - | - | - | - |
| Pelargonidin-3-malonylglucoside | - | - | - | - | - | - | - | 5.51–8.16 | - | - | - | - | - |
| Pelargonidin-3-glucoside | - | - | - | - | - | - | - | 114–348 | - | 17.58 | - | - | - |
| Pelargonidin-3-rutinoside | - | - | - | - | - | - | - | 18–62 | - | - | - | - |
4. Bilberries
5. Cranberries
6. Raspberries
7. Mulberries
8. Lingonberries
9. Blackberries
10. Strawberries
11. Goji Berries
12. Acai Berries
13. Chokeberries
14. Black Currants
15. Maqui Berries
16. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- World Health Organization (WHO). Organisation for Economic Co-Operation and Development (OECD); World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Nathan, D.M.; Buse, J.B.; Davidson, M.B.; Heine, R.J.; Holman, R.R.; Sherwin, R.; Zinman, B. Management of Hyperglycemia in Type 2 Diabetes: A consensus algorithm for the initiation and adjustment of therapy: A consensus statement from the American Diabetes Association and the European Association for the Study of Diabetes. Diabetes Care 2006, 29, 1963–1972. [Google Scholar] [CrossRef] [Green Version]
- Enwere, O.; Salako, B.; Falade, C. Prescription and cost consideration at a diabetic clinic in Ibadan, Nigeria: A report. Ann. Ib. Postgrad. Med. 2010, 4. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.B.; Etienne, C. Health systems financing and the path to universal coverage. Bull. World Health Organ. 2010, 88, 402. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.D.; Hu, F.B. Precision nutrition for prevention and management of type 2 diabetes. Lancet Diabetes Endocrinol. 2018, 6, 416–426. [Google Scholar] [CrossRef]
- Ley, S.H.; Hamdy, O.; Mohan, V.; Hu, F.B. Prevention and management of type 2 diabetes: Dietary components and nutritional strategies. Lancet 2014, 383, 1999–2007. [Google Scholar] [CrossRef] [Green Version]
- Mursu, J.; Virtanen, J.; Tuomainen, T.P.; Nurmi, T.; Voutilainen, S. Intake of fruit, berries, and vegetables and risk of type 2 diabetes in Finnish men: The Kuopio Ischaemic Heart Disease Risk Factor Study. Am. J. Clin. Nutr. 2013, 99, 328–333. [Google Scholar] [CrossRef]
- Moher, D. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS ONE Med. 2009, 6, e1000097. [Google Scholar]
- Barberis, A.; Garbetta, A.; Cardinali, A.; Bazzu, G.; D’Antuono, I.; Rocchitta, G.; Fadda, A.; Linsalata, V.; D’Hallewin, G.; Serra, P.A.; et al. Real-time monitoring of glucose and phenols intestinal absorption through an integrated Caco-2TC7cells/biosensors telemetric device: Hypoglycemic effect of fruit phytochemicals. Biosens. Bioelectron. 2017, 88, 159–166. [Google Scholar] [CrossRef]
- Martineau, L.C.; Couture, A.; Spoor, D.; Benhaddou-Andaloussi, A.; Harris, C.; Meddah, B.; LeDuc, C.; Burt, A.; Vuong, T.; Le, P.M.; et al. Anti-diabetic properties of the Canadian lowbush blueberry Vaccinium angustifolium Ait. Phytomedicine 2006, 13, 612–623. [Google Scholar] [CrossRef]
- DeFuria, J.; Bennett, G.; Strissel, K.J.; Perfield, J.W.; Milbury, P.E.; Greenberg, A.S.; Obin, M.S. Dietary blueberry attenuates whole-body insulin resistance in high fat-fed mice by reducing adipocyte death and its inflammatory sequelae. J. Nutr. 2009, 139, 1510–1516. [Google Scholar] [CrossRef]
- Wu, T.; Tang, Q.; Gao, Z.; Yu, Z.; Song, H.; Zheng, X.; Chen, W. Blueberry and mulberry juice prevent obesity development in C57BL/6 Mice. PLoS ONE 2013, 8, e77585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grace, M.H.; Ribnicky, D.M.; Kuhn, P.; Poulev, A.; Logendra, S.; Yousef, G.G.; Raskin, I.; Lila, M.A. Hypoglycemic activity of a novel anthocyanin-rich formulation from lowbush blueberry, Vaccinium angustifolium Aiton. Phytomedicine 2009, 16, 406–415. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Yao, L.; He, X.; Wang, L.; Li, M.; Yang, Y.; Wan, C. Hypoglycemic activity and constituents analysis of blueberry (Vaccinium corymbosum) fruit extracts. Diabetes Metab. Syndr. Obes. Targets Ther. 2018, 11, 357–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takikawa, M.; Inoue, S.; Horio, F.; Tsuda, T. Dietary anthocyanin-rich bilberry extract ameliorates hyperglycemia and insulin sensitivity via Activation of AMP-activated protein kinase in diabetic mice. J. Nutr. 2010, 140, 527–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, A.R.; Elks, C.M.; Vila, J.; Del Piero, F.; Paulsen, D.B.; Francis, J. A Blueberry-enriched diet improves renal function and reduces oxidative stress in metabolic syndrome animals: Potential mechanism of TLR4-MAPK signaling pathway. PLoS ONE 2014, 9, e111976. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, J.I.; Tanaka, I.; Seo, S.; Yamazaki, M.; Saito, K. LC/PDA/ESI-MS profiling and radical scavenging activity of anthocyanins in various berries. J. Biomed. Biotechnol. 2004, 2004, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Mykkänen, O.T.; Huotari, A.; Herzig, K.-H.; Dunlop, T.W.; Mykkänen, H.; Kirjavainen, P.V. Wild blueberries (Vaccinium myrtillus) alleviate inflammation and hypertension associated with developing obesity in mice fed with a high-fat Diet. PLoS ONE 2014, 9, e114790. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Gu, L.; Hager, T.J.; Hager, A.; Howard, L.R. Whole berries versus berry anthocyanins: Interactions with dietary fat levels in the C57BL/6J mouse model of obesity. J. Agric. Food Chem. 2008, 56, 647–653. [Google Scholar] [CrossRef]
- Basu, A.; Du, M.; Leyva, M.J.; Sanchez, K.; Betts, N.M.; Wu, M.; Aston, C.E.; Lyons, T.J. Blueberries decrease cardiovascular risk factors in obese men and women with metabolic syndrome. J. Nutr. 2010, 140, 1582–1587. [Google Scholar] [CrossRef] [Green Version]
- Stull, A.J.; Cash, K.C.; Champagne, C.M.; Gupta, A.K.; Boston, R.C.; Beyl, R.A.; Johnson, W.D.; Cefalu, W.T. Blueberries improve endothelial function, but not blood pressure, in adults with metabolic syndrome: A randomized, double-blind, placebo-controlled clinical trial. Nutrients 2015, 7, 4107–4123. [Google Scholar] [CrossRef]
- Johnson, M.H.; Wallig, M.; Luna-Vital, D.A.; De Mejia, E.G. Alcohol-free fermented blueberry–blackberry beverage phenolic extract attenuates diet-induced obesity and blood glucose in C57BL/6J mice. J. Nutr. Biochem. 2016, 31, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Vuong, T.; Benhaddou-Andaloussi, A.; Brault, A.; Harbilas, D.; Martineau, L.C.; Vallerand, D.; Ramassamy, C.; Matar, C.; Haddad, P.S. Antiobesity and antidiabetic effects of biotransformed blueberry juice in KKAy mice. Int. J. Obes. 2009, 33, 1166–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, L.; Lamport, D.J.; Butler, L.T.; Williams, C. A study of glycaemic effects following acute anthocyanin-rich blueberry supplementation in healthy young adults. Food Funct. 2017, 8, 3104–3110. [Google Scholar] [CrossRef] [Green Version]
- Vendrame, S.; Daugherty, A.; Kristo, A.S.; Riso, P.; Klimis-Zacas, D. Wild blueberry (Vaccinium angustifolium) consumption improves inflammatory status in the obese Zucker rat model of the metabolic syndrome. J. Nutr. Biochem. 2013, 24, 1508–1512. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Liu, W.; Yu, J.; Zou, S.; Wang, J.; Yao, W.; Gao, X. Characterization and hypoglycemic effect of a polysaccharide extracted from the fruit of Lycium barbarum L. Carbohydr. Polym. 2013, 98, 8–16. [Google Scholar] [CrossRef]
- Bharat, D.; Cavalcanti, R.R.M.; Petersen, C.; Begaye, N.; Cutler, B.R.; Costa, M.M.A.; Ramos, R.K.L.G.; Ferreira, M.R.; Li, Y.; Bharath, L.P.; et al. Blueberry metabolites attenuate lipotoxicity-induced endothelial dysfunction. Mol. Nutr. Food Res. 2017, 62, 1700601. [Google Scholar] [CrossRef]
- Bloedon, T.; Vendrame, S.; Bolton, J.; Lehnhard, R.; Riso, P.; Klimis-Zacas, D. The effect of wild blueberry (Vaccinium angustifolium) consumption on oxidative stress, inflammation, and DNA damage associated with exercise. Comp. Exerc. Physiol. 2015, 11, 173–181. [Google Scholar] [CrossRef]
- Li, F.; Zhang, B.; Chen, G.; Fu, X. The novel contributors of anti-diabetic potential in mulberry polyphenols revealed by UHPLC-HR-ESI-TOF-MS/MS. Food Res. Int. 2017, 100, 873–884. [Google Scholar] [CrossRef]
- Ren, G.; Rimando, A.M.; Mathews, S.T. AMPK activation by pterostilbene contributes to suppression of hepatic gluconeogenic gene expression and glucose production in H4IIE cells. Biochem. Biophys. Res. Commun. 2018, 498, 640–645. [Google Scholar] [CrossRef]
- Lee, S.I.; Keirsey, K.; Kirkland, R.I.; Grunewald, Z.; Fischer, J.G.; De La Serre, C.B. Blueberry supplementation influences the gut microbiota, inflammation, and insulin resistance in high-fat-diet–fed rats. J. Nutr. 2018, 148, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Vendrame, S.; Guglielmetti, S.; Riso, P.; Arioli, S.; Klimis-Zacas, D.; Porrini, M. Six-week consumption of a wild blueberry powder drink increases bifidobacteria in the human gut. J. Agric. Food Chem. 2011, 59, 12815–12820. [Google Scholar] [CrossRef] [PubMed]
- Polewski, M.A.; Esquivel-Alvarado, D.; Wedde, N.S.; Kruger, C.G.; Reed, J.D. Isolation and characterization of blueberry polyphenolic components and their effects on gut barrier dysfunction. J. Agric. Food Chem. 2019, 68, 2940–2947. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Daza, M.-C.; Daoust, L.; Boutkrabt, L.; Pilon, G.; Varin, T.; Dudonné, S.; Levy, É.; Marette, A.; Roy, D.; Desjardins, Y. Wild blueberry proanthocyanidins shape distinct gut microbiota profile and influence glucose homeostasis and intestinal phenotypes in high-fat high-sucrose fed mice. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Pang, W.; Lu, H.; Cheng, D.; Yan, X.; Cheng, Y.; Jiang, Y. Comparison of metabolic profiling of cyanidin-3-o-galactoside and extracts from blueberry in aged mice. J. Agric. Food Chem. 2011, 59, 2069–2076. [Google Scholar] [CrossRef]
- Harris, C.; Burt, A.J.; Saleem, A.; Le, P.M.; Martineau, L.C.; Haddad, P.S.; Bennett, S.A.L.; Arnason, J.T. A single HPLC-PAD-APCI/MS method for the quantitative comparison of phenolic compounds found in leaf, stem, root and fruit extracts of Vaccinium angustifolium. Phytochem. Anal. 2007, 18, 161–169. [Google Scholar] [CrossRef]
- Lätti, A.K.; Kainulainen, P.S.; Hayirlioglu-Ayaz, S.; Ayaz, F.A.; Riihinen, K.R. Characterization of anthocyanins in caucasian blueberries (Vaccinium arctostaphylos L.) native to Turkey. J. Agric. Food Chem. 2009, 57, 5244–5249. [Google Scholar] [CrossRef]
- Može, S.; Polak, T.; Gašperlin, L.; Koron, D.; Vanzo, A.; Ulrih, N.P.; Abram, V. Phenolics in slovenian bilberries (Vaccinium myrtillus L.) and blueberries (Vaccinium corymbosum L.). J. Agric. Food Chem. 2011, 59, 6998–7004. [Google Scholar] [CrossRef]
- Shi, M.; Loftus, H.; McAinch, A.J.; Su, X.Q. Blueberry as a source of bioactive compounds for the treatment of obesity, type 2 diabetes and chronic inflammation. J. Funct. Foods 2017, 30, 16–29. [Google Scholar] [CrossRef] [Green Version]
- Seymour, E.M.; Tanone, I.I.; Urcuyo-Llanes, D.E.; Lewis, S.K.; Kirakosyan, A.; Kondoleon, M.G.; Kaufman, P.B.; Bolling, S.F. Blueberry intake alters skeletal muscle and adipose tissue peroxisome proliferator-activated receptor activity and reduces insulin resistance in obese rats. J. Med. Food 2011, 14, 1511–1518. [Google Scholar] [CrossRef] [Green Version]
- Rebello, C.J.; Burton, J.; Heiman, M.; Greenway, F.L. Gastrointestinal microbiome modulator improves glucose tolerance in overweight and obese subjects: A randomized controlled pilot trial. J. Diabetes Complicat. 2015, 29, 1272–1276. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Zhang, Y.; Liu, Y.; Sun, R.; Xia, M. Purified anthocyanin supplementation reduces dyslipidemia, enhances antioxidant capacity, and prevents insulin resistance in diabetic patients. J. Nutr. 2015, 145, 742–748. [Google Scholar] [CrossRef] [PubMed]
- Asgary, S.; Rafieian-Kopaei, M.; Sahebkar, A.; Shamsi, F.; Goli-Malekabadi, N. Anti-hyperglycemic and anti-hyperlipidemic effects of Vaccinium myrtillusfruit in experimentally induced diabetes (antidiabetic effect of Vaccinium myrtillusfruit). J. Sci. Food Agric. 2015, 96, 764–768. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, C.-S.; Lee, Y.M.; Sohn, E.; Jo, K.; Kim, J.S. Vaccinium myrtillus extract prevents or delays the onset of diabetes—Induced blood–retinal barrier breakdown. Int. J. Food Sci. Nutr. 2015, 66, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Yao, X.S.; Yau, C.C.; Tsi, D.; Chia, C.-S.; Nagai, H.; Kurihara, H. Protective effects of bilberry (Vaccinium myrtillus L.) extract on restraint stress-induced liver damage in mice. J. Agric. Food Chem. 2008, 56, 7803–7807. [Google Scholar] [CrossRef]
- Cohen-Boulakia, F.; Valensi, P.E.; Boulahdour, H.; Lestrade, R.; Dufour-Lamartinie, J.-F.; Hort-Legrand, C.; Behar, A. In vivo sequential study of skeletal muscle capillary permeability in diabetic rats: Effect of anthocyanosides. Metabolism 2000, 49, 880–885. [Google Scholar] [CrossRef]
- Ferreira, F.M.; Peixoto, F.; Nunes, E.; Sena, C.; Seiça, R.; Santos, M.S. MitoTeas: Vaccinium myrtillus and Geranium robertianum decoctions improve diabetic Goto–Kakizaki rats hepatic mitochondrial oxidative phosphorylation. Biochim. Biophys. Acta 2010, 1797, 79–80. [Google Scholar] [CrossRef] [Green Version]
- Hoggard, N.; Cruickshank, M.; Moar, K.M.; Bestwick, C.; Holst, J.J.; Russell, W.R.; Horgan, G. A single supplement of a standardised bilberry (Vaccinium myrtillus L.) extract (36 % wet weight anthocyanins) modifies glycaemic response in individuals with type 2 diabetes controlled by diet and lifestyle. J. Nutr. Sci. 2013, 2, e22. [Google Scholar] [CrossRef] [Green Version]
- Alnajjar, M.; Barik, S.K.; Bestwick, C.; Campbell, F.; Cruickshank, M.; Farquharson, F.; Holtrop, G.; Horgan, G.; Louis, P.; Moar, K.-M.; et al. Anthocyanin-enriched bilberry extract attenuates glycaemic response in overweight volunteers without changes in insulin. J. Funct. Foods 2020, 64, 103597. [Google Scholar] [CrossRef]
- Granfeldt, Y.; Björck, I. A bilberry drink with fermented oatmeal decreases postprandial insulin demand in young healthy adults. Nutr. J. 2011, 10, 57. [Google Scholar] [CrossRef] [Green Version]
- Karlsen, A.; Paur, I.; Bøhn, S.K.; Sakhi, A.K.; Borge, G.I.; Serafini, M.; Erlund, I.; Laake, P.; Tonstad, S.; Blomhoff, R. Bilberry juice modulates plasma concentration of NF-κB related inflammatory markers in subjects at increased risk of CVD. Eur. J. Nutr. 2010, 49, 345–355. [Google Scholar] [CrossRef]
- Erlund, I.; Marniemi, J.; Hakala, P.; Alfthan, G.; Meririnne, E.; Aro, A. Consumption of black currants, lingonberries and bilberries increases serum quercetin concentrations. Eur. J. Clin. Nutr. 2003, 57, 37–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolehmainen, M.; Mykkänen, O.; Kirjavainen, P.V.; Leppänen, T.; Moilanen, E.; Adriaens, M.E.; Laaksonen, D.E.; Hallikainen, M.; Puupponen-Pimia, R.; Pulkkinen, L.; et al. Bilberries reduce low-grade inflammation in individuals with features of metabolic syndrome. Mol. Nutr. Food Res. 2012, 56, 1501–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anhê, F.F.; Roy, D.; Pilon, G.; Dudonné, S.; Matamoros, S.; Varin, T.V.; Garofalo, C.; Moine, Q.; Desjardins, Y.; Levy, E.; et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increasedAkkermansiaspp. population in the gut microbiota of mice. Gut 2014, 64, 872–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, M.; Hu, J.; Perez, E.; Phillips, D.; Kim, W.; Ghaedian, R.; Napora, J.K.; Zou, S. Effects of long-term cranberry supplementation on endocrine pancreas in aging Rats. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2011, 66, 1139–1151. [Google Scholar] [CrossRef] [Green Version]
- Schell, J.; Betts, N.M.; Foster, M.; Scofield, R.H.; Basu, A. Cranberries improve postprandial glucose excursions in type 2 diabetes. Food Funct. 2017, 8, 3083–3090. [Google Scholar] [CrossRef]
- Lee, I.T.; Chan, Y.C.; Lin, C.W.; Lee, W.J.; Sheu, W.H.-H. Effect of cranberry extracts on lipid profiles in subjects with Type 2 diabetes. Diabet. Med. 2008, 25, 1473–1477. [Google Scholar] [CrossRef]
- Wilson, T.; Meyers, S.; Singh, A.; Limburg, P.; Vorsa, N. Favorable glycemic response of Type 2 diabetics to low-calorie cranberry juice. J. Food Sci. 2008, 73, H241–H245. [Google Scholar] [CrossRef]
- Wilson, T.; Singh, A.P.; Vorsa, N.; Goettl, C.D.; Kittleson, K.M.; Roe, C.M.; Kastello, G.M.; Ragsdale, F.R. Human glycemic response and phenolic content of unsweetened cranberry juice. J. Med. Food 2008, 11, 46–54. [Google Scholar] [CrossRef]
- Wilson, T.; Luebke, J.L.; Morcomb, E.F.; Carrell, E.J.; Leveranz, M.C.; Kobs, L.; Schmidt, T.P.; Limburg, P.J.; Vorsa, N.; Singh, A.P. Glycemic responses to sweetened dried and raw cranberries in humans with Type 2 diabetes. J. Food Sci. 2010, 75, H218–H223. [Google Scholar] [CrossRef]
- Duffey, K.J.; Sutherland, L.A. Adult consumers of cranberry juice cocktail have lower C-reactive protein levels compared with nonconsumers. Nutr. Res. 2015, 35, 118–126. [Google Scholar] [CrossRef]
- Törrönen, R.; Sarkkinen, E.; Niskanen, T.; Tapola, N.; Kilpi, K.; Niskanen, L. Postprandial glucose, insulin and glucagon-like peptide 1 responses to sucrose ingested with berries in healthy subjects. Br. J. Nutr. 2011, 107, 1445–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javid, A.Z.; Maghsoumi-Norouzabad, L.; Ashrafzadeh, E.; Yousefimanesh, H.A.; Zakerkish, M.; Angali, K.A.; Ravanbakhsh, M.; Babaei, H. Impact of cranberry juice enriched with Omega-3 Fatty acids adjunct with nonsurgical periodontal treatment on metabolic control and periodontal status in Type 2 patients with diabetes with periodontal Disease. J. Am. Coll. Nutr. 2017, 37, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Shidfar, F.; Heydari, I.; Hajimiresmaiel, S.J.; Hosseini, S.; Shidfar, S.; Amiri, F. The effects of cranberry juice on serum glucose, apoB, apoA-I, Lp (a), and Paraoxonase-1 activity in type 2 diabetic male patients. J. Res. Med. Sci. 2012, 17, 355–360. [Google Scholar] [PubMed]
- Novotny, A.; Baer, J.; Khoo, D.J.; Gebauer, C.; Charron, S.K. Cranberry juice consumption lowers markers of cardiometabolic risk, including blood pressure and circulating c-reactive protein, triglyceride, and glucose concentrations in adults. J. Nutr. 2015, 145, 1185–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simão, T.N.C.; Lozovoy, M.A.B.; Simão, A.N.C.; Oliveira, S.R.; Venturini, D.; Morimoto, H.K.; Miglioranza, L.H.S.; Dichi, I. Reduced-energy cranberry juice increases folic acid and adiponectin and reduces homocysteine and oxidative stress in patients with the metabolic syndrome. Br. J. Nutr. 2013, 110, 1885–1894. [Google Scholar] [CrossRef]
- Ruel, G.; Lapointe, A.; Pomerleau, S.; Couture, P.; Lemieux, S.; Lamarche, B.; Couillard, C. Evidence that cranberry juice may improve augmentation index in overweight men. Nutr. Res. 2013, 33, 41–49. [Google Scholar] [CrossRef]
- Ruel, G.; Pomerleau, S.; Couture, P.; Lemieux, S.; Lamarche, B.; Couillard, C. Plasma matrix metalloproteinase (MMP)-9 levels are reduced following low-calorie cranberry juice supplementation in men. J. Am. Coll. Nutr. 2009, 28, 694–701. [Google Scholar] [CrossRef]
- Ruel, G.; Pomerleau, S.; Couture, P.; Lemieux, S.; Lamarche, B.; Couillard, C. Low-calorie cranberry juice supplementation reduces plasma oxidized LDL and cell adhesion molecule concentrations in men. Br. J. Nutr. 2008, 99, 352–359. [Google Scholar] [CrossRef] [Green Version]
- Ruel, G.; Pomerleau, S.; Couture, P.; Lemieux, S.; Lamarche, B.; Couillard, C. Favourable impact of low-calorie cranberry juice consumption on plasma HDL-cholesterol concentrations in men. Br. J. Nutr. 2006, 96, 357–364. [Google Scholar] [CrossRef]
- Ruel, G.; Pomerleau, S.; Couture, P.; Lamarche, B.; Couillard, C. Changes in plasma antioxidant capacity and oxidized low-density lipoprotein levels in men after short-term cranberry juice consumption. Metabolism 2005, 54, 856–861. [Google Scholar] [CrossRef]
- Suh, J.H.; Romain, C.; González-Barrio, R.; Cristol, J.P.; Teissedre, P.L.; Crozier, A.; Rouanet, J.M. Raspberry juice consumption, oxidative stress and reduction of atherosclerosis risk factors in hypercholesterolemic golden Syrian hamsters. Food Funct. 2011, 2, 400–405. [Google Scholar] [CrossRef] [PubMed]
- SanGiovanni, E.; Vrhovšek, U.; Rossoni, G.; Colombo, E.; Brunelli, C.; Brembati, L.; Trivulzio, S.; Gasperotti, M.; Mattivi, F.; Bosisio, E.; et al. Ellagitannins from rubus berries for the control of gastric inflammation: In vitro and in vivo studies. PLoS ONE 2013, 8, e71762. [Google Scholar] [CrossRef] [Green Version]
- Jean-Gilles, D.; Li, L.; Ma, H.; Yuan, T.; Chichester, I.C.O.; Seeram, N.P. Anti-inflammatory effects of polyphenolic-enriched red raspberry extract in an antigen-induced arthritis rat model. J. Agric. Food Chem. 2011, 60, 5755–5762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gião, M.; Pestana, D.; Faria, A.; Guimarães, J.T.; Pintado, M.; Calhau, C.; Azevedo, I.; Malcata, F.X. Effects of extracts of selected medicinal plants upon hepatic oxidative stress. J. Med. Food 2010, 13, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Noratto, G.; Chew, B.P.; Atienza, L.M. Red raspberry (Rubus idaeus L.) intake decreases oxidative stress in obese diabetic (db/db) mice. Food Chem. 2017, 227, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Kirakosyan, A.; Seymour, E.M.; Kondoleon, N.; Gutiérrez, E.; Wolforth, J.; Bolling, S. The intake of red raspberry fruit is inversely related to cardiac risk factors associated with metabolic syndrome. J. Funct. Foods 2018, 41, 83–89. [Google Scholar] [CrossRef]
- Jakobsdottir, G.; Blanco, N.; Xu, J.; Ahrné, S.; Molin, G.; Sterner, O.; Nyman, M. Formation of short-chain fatty acids, excretion of anthocyanins, and microbial diversity in rats fed blackcurrants, blackberries, and raspberries. J. Nutr. Metab. 2013, 2013, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pan, P.; Lam, V.; Salzman, N.H.; Huang, Y.W.; Yu, J.; Zhang, J.; Wang, L. Black raspberries and their anthocyanin and fiber fractions alter the composition and diversity of gut microbiota in F-344 rats. Nutr. Cancer 2017, 69, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Xie, L.; Xu, Y.; Ke, H.; Bao, T.; Li, Y.; Chen, W. Pelargonidin-3-O-glucoside derived from wild raspberry exerts antihyperglycemic effect by inducing autophagy and modulating gut microbiota. J. Agric. Food Chem. 2019. [Google Scholar] [CrossRef]
- Tu, P.; Bian, X.; Chi, L.; Gao, B.; Ru, H.; Knobloch, T.J.; Weghorst, C.M.; Lu, K. Characterization of the functional changes in mouse gut microbiome associated with increased akkermansia muciniphila population modulated by dietary black raspberries. ACS Omega 2018, 3, 10927–10937. [Google Scholar] [CrossRef]
- Atienza, L.E.; Garcia, P.; Noratto, G. Effects of raspberry on biomarkers of diabetes, cardiovascular disease (cvd) and oxidative stress in obese diabetic (db/db) mice. FASEB J. 2015, 29, LB283. [Google Scholar]
- Luo, T.; Miranda-Garcia, O.; Adamson, A.; Sasaki, G.; Shay, N. Development of obesity is reduced in high-fat fed mice fed whole raspberries, raspberry juice concentrate, and a combination of the raspberry phytochemicals ellagic acid and raspberry ketone. J. Berry Res. 2016, 6, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Xia, M.; Zou, T.; Ling, W.; Zhong, R.; Zhang, W. Cyanidin 3-glucoside attenuates obesity-associated insulin resistance and hepatic steatosis in high-fat diet-fed and db/db mice via the transcription factor FoxO1. J. Nutr. Biochem. 2012, 23, 349–360. [Google Scholar] [CrossRef]
- Iino, T.; Tashima, K.; Umeda, M.; Ogawa, Y.; Takeeda, M.; Takata, K.; Takeuchi, K. Effect of ellagic acid on gastric damage induced in ischemic rat stomachs following ammonia or reperfusion. Life Sci. 2002, 70, 1139–1150. [Google Scholar] [CrossRef]
- Rosillo, M.A. Protective effect of ellagic acid, a natural polyphenolic compound, in a murine model of Crohn’s disease. Biochem. Pharmacol. 2011, 82, 737–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Zou, T.; Gomez, N.A.; Wang, B.; Zhu, M.-J.; Du, M. Raspberry alleviates obesity-induced inflammation and insulin resistance in skeletal muscle through activation of AMP-activated protein kinase (AMPK) α1. Nutr. Diabetes 2018, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Kang, Y.; Xue, Y.; Liang, X.; García, M.P.G.; Rodgers, D.; Kagel, D.R.; Du, M. Red raspberries suppress NLRP3 inflammasome and attenuate metabolic abnormalities in diet-induced obese mice. J. Nutr. Biochem. 2017, 53, 96–103. [Google Scholar] [CrossRef]
- Sasaki, R.; Nishimura, N.; Hoshino, H.; Isa, Y.; Kadowaki, M.; Ichi, T.; Tanaka, A.; Nishiumi, S.; Fukuda, I.; Ashida, H.; et al. Cyanidin 3-glucoside ameliorates hyperglycemia and insulin sensitivity due to downregulation of retinol binding protein 4 expression in diabetic mice. Biochem. Pharmacol. 2007, 74, 1619–1627. [Google Scholar] [CrossRef]
- Kang, I.; Espín, J.C.; Carr, T.P.; Tomás-Barberán, F.A.; Chung, S. Raspberry seed flour attenuates high-sucrose diet-mediated hepatic stress and adipose tissue inflammation. J. Nutr. Biochem. 2016, 32, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.J.; Karl, J.P.; Wilson, M.A.; Whitney, C.C.; Barrett, A.; Farhadi, N.F.; Chen, C.Y.O.; Montain, S.J. Glycaemic regulation, appetite and ex vivo oxidative stress in young adults following consumption of high-carbohydrate cereal bars fortified with polyphenol-rich berries. Br. J. Nutr. 2019, 121, 1026–1038. [Google Scholar] [CrossRef]
- Clegg, M.E.; Pratt, M.; Meade, C.M.; Henry, C.J. The addition of raspberries and blueberries to a starch-based food does not alter the glycaemic response. Br. J. Nutr. 2011, 106, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Törrönen, R.; Kolehmainen, M.; Sarkkinen, E.; Poutanen, K.; Mykkänen, H.; Niskanen, L. Berries reduce postprandial insulin responses to wheat and rye breads in healthy women. J. Nutr. 2013, 143, 430–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schell, J.; Betts, N.M.; Lyons, T.J.; Basu, A. Raspberries improve postprandial glucose and acute and chronic inflammation in adults with Type 2 Diabetes. Ann. Nutr. Metab. 2019, 74, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Yin, J.; Zhang, G.; Long, H.; Zheng, X. Mulberry and cherry anthocyanin consumption prevents oxidative stress and inflammation in diet-induced obese mice. Mol. Nutr. Food Res. 2016, 60, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Zheng, X. Anthocyanin-rich mulberry fruit improves insulin resistance and protects hepatocytes against oxidative stress during hyperglycemia by regulating AMPK/ACC/mTOR pathway. J. Funct. Foods 2017, 30, 270–281. [Google Scholar] [CrossRef]
- Srikanta, A.H.; Kumar, A.; Sukhdeo, S.V.; Peddha, M.S.; Govindaswamy, V. The antioxidant effect of mulberry and jamun fruit wines by ameliorating oxidative stress in streptozotocin-induced diabetic Wistar rats. Food Funct. 2016, 7, 4422–4431. [Google Scholar] [CrossRef]
- Ha, U.S.; Bae, W.J.; Kim, S.J.; Yoon, B.I.; Jang, H.; Hong, S.H.; Lee, J.Y.; Hwang, S.Y.; Kim, S.W. Protective effect of cyanidin-3-O-β-D-glucopyranoside fraction from mulberry fruit pigment against oxidative damage in streptozotocin-induced diabetic rat bladder. Neurourol. Urodyn. 2012, 32, 493–499. [Google Scholar] [CrossRef]
- Ren, C.; Zhang, Y.; Cui, W.; Lu, G.; Wang, Y.; Gao, H.; Huang, L.; Mu, Z. A polysaccharide extract of mulberry leaf ameliorates hepatic glucose metabolism and insulin signaling in rats with type 2 diabetes induced by high fat-diet and streptozotocin. Int. J. Biol. Macromol. 2015, 72, 951–959. [Google Scholar] [CrossRef]
- Liu, L.K.; Chou, F.-P.; Chen, Y.C.; Chyau, C.C.; Ho, H.H.; Wang, C.J. Effects of mulberry (Morus albal.) extracts on lipid homeostasis in vitro and in vivo. J. Agric. Food Chem. 2009, 57, 7605–7611. [Google Scholar] [CrossRef]
- Wu, T.; Qi, X.; Liu, Y.; Guo, J.; Zhu, R.; Chen, W.; Zheng, X.; Yu, T. Dietary supplementation with purified mulberry (Morus australis Poir) anthocyanins suppresses body weight gain in high-fat diet fed C57BL/6 mice. Food Chem. 2013, 141, 482–487. [Google Scholar] [CrossRef]
- Du, Q.; Zheng, J.; Xu, Y. Composition of anthocyanins in mulberry and their antioxidant activity. J. Food Compos. Anal. 2008, 21, 390–395. [Google Scholar] [CrossRef]
- Peng, C.H.; Liu, L.K.; Chyau, C.C.; Wang, C.J.; Chuang, C.M.; Huang, C.N. Mulberry water extracts possess an anti-obesity effect and ability to inhibit hepatic lipogenesis and promote lipolysis. J. Agric. Food Chem. 2011, 59, 2663–2671. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.C.; Huang, H.P.; Lee, Y.J.; Tang, Y.H.; Wang, C.J. Hepatoprotective effect of mulberry water extracts on ethanol-induced liver injury via anti-inflammation and inhibition of lipogenesis in C57BL/6J mice. Food Chem. Toxicol. 2013, 62, 786–796. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.K.; Joo-Yong, L. Mulberry fruit extract ameliorates nonalcoholic fatty liver disease (NAFLD) through inhibition of mitochondrial oxidative stress in rats. Evid. Based Complement. Altern. Med. 2018, 2018, 8165716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wattanathorn, J.; Kawvised, S.; Thukham-Mee, W. Encapsulated Mulberry Fruit Extract Alleviates Changes in an Animal Model of Menopause with Metabolic Syndrome. Oxidative Med. Cell. Longev. 2019, 2019, 5360560. [Google Scholar] [CrossRef]
- Sheng, Y.; Liu, J.; Zheng, S.; Liang, F.; Luo, Y.; Huang, K.; Xu, W.; He, X. Mulberry leaves ameliorate obesity through enhancing brown adipose tissue activity and modulating gut microbiota. Food Funct. 2019, 10, 4771–4781. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; You, L.; Huang, Q.; Fu, X.; Zhang, B.; Liu, R.-H.; Li, C. Modulation of gut microbiota by mulberry fruit polysaccharide treatment of obese diabetic db/db mice. Food Funct. 2018, 9, 3732–3742. [Google Scholar] [CrossRef]
- Pierre, W. Hypoglycemic and hypolipidemic effects of Bersama engleriana leaves in nicotinamide/streptozotocin-induced type 2 diabetic rats. BMC Complement. Altern. Med. 2012, 12, 264. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Wang, X.; Jiang, X.; Kong, F.; Wang, S.; Yan, C. Antidiabetic effects of Morus alba fruit polysaccharides on high-fat diet- and streptozotocin-induced type 2 diabetes in rats. J. Ethnopharmacol. 2017, 199, 119–127. [Google Scholar] [CrossRef]
- Eid, H.M.; Ouchfoun, M.; Brault, A.; Vallerand, D.; Musallam, L.; Arnason, J.T.; Haddad, P.S. Lingonberry (Vaccinium vitis-idaea L.) exhibits antidiabetic activities in a mouse model of diet-induced obesity. Evid. Based Complement. Altern. Med. 2014, 2014, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kivimäki, A.S.; Ehlers, P.I.; Siltari, A.; Turpeinen, A.M.; Vapaatalo, H.; Korpela, R. Lingonberry, cranberry and blackcurrant juices affect mRNA expressions of inflammatory and atherothrombotic markers of SHR in a long-term treatment. J. Funct. Foods 2012, 4, 496–503. [Google Scholar] [CrossRef]
- Heyman-Lindén, L.; Axling, U.; Blanco, N.; Sterner, O.; Holm, C.; Berger, K. Evaluation of beneficial metabolic effects of berries in high-fat fed C57BL/6J Mice. J. Nutr. Metab. 2014, 2014, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Heyman-Lindén, L.; Kotowska, D.; Sand, E.; Bjursell, M.; Plaza, M.; Turner, C.; Holm, C.; Fåk, F.; Berger, K. Lingonberries alter the gut microbiota and prevent low-grade inflammation in high-fat diet fed mice. Food Nutr. Res. 2016, 60, 29993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matziouridou, C.; Marungruang, N.; Nguyen, T.D.; Nyman, M.; Fak, F. Lingonberries reduce atherosclerosis inApoe-/-mice in association with altered gut microbiota composition and improved lipid profile. Mol. Nutr. Food Res. 2016, 60, 1150–1160. [Google Scholar] [CrossRef]
- Al-Hamimi, S.; Heyman-Lindén, L.; Plaza, M.; Turner, C.; Berger, K.; Spégel, P. Alterations in the plasma metabolite profile associated with improved hepatic function and glycemia in mice fed lingonberry supplemented high-fat diets. Mol. Nutr. Food Res. 2016, 61, 1600442. [Google Scholar] [CrossRef]
- Heyman-Lindén, L.; Seki, Y.; Storm, P.; Jones, H.A.; Charron, M.J.; Berger, K.; Holm, C. Berry intake changes hepatic gene expression and DNA methylation patterns associated with high-fat diet. J. Nutr. Biochem. 2016, 27, 79–95. [Google Scholar] [CrossRef] [Green Version]
- Marungruang, N.; Kovalenko, T.; Osadchenko, I.; Voss, U.; Huang, F.; Burleigh, S.; Ushakova, A.A.; Skibo, G.; Nyman, M.; Prykhod’Ko, O.; et al. Lingonberries and their two separated fractions differently alter the gut microbiota, improve metabolic functions, reduce gut inflammatory properties, and improve brain function in ApoE−/− mice fed high-fat diet. Nutr. Neurosci. 2018, 23, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Nilholm, C.; Roth, B.; Linninge, C.; Hoglund, P.; Nyman, M.; Ohlsson, B. Anthropometric and metabolic improvements in human type 2 diabetes after introduction of an Okinawan-based Nordic diet are not associated with changes in microbial diversity or SCFA concentrations. Int. J. Food Sci. Nutr. 2017, 69, 729–740. [Google Scholar] [CrossRef]
- Linderborg, K.M.; Järvinen, R.; Lehtonen, H.M.; Viitanen, M.; Kallio, H. The fiber and/or polyphenols present in lingonberries null the glycemic effect of the sugars present in the berries when consumed together with added glucose in healthy human volunteers. Nutr. Res. 2012, 32, 471–478. [Google Scholar] [CrossRef]
- Furlan, C.; Valle, S.C.; Maróstica, M.R.; Östman, E.; Björck, I.; Tovar, J. Effect of bilberries, lingonberries and cinnamon on cardiometabolic risk-associated markers following a hypercaloric-hyperlipidic breakfast. J. Funct. Foods 2019, 60, 103443. [Google Scholar] [CrossRef]
- Rokka, S.; Ketoja, E.; Järvenpää, E.; Tahvonen, R. The glycaemic and C-peptide responses of foods rich in dietary fibre from oat, buckwheat and lingonberry. Int. J. Food Sci. Nutr. 2013, 64, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Törrönen, R.; Kolehmainen, M.; Sarkkinen, E.; Mykkänen, H.; Niskanen, L. Postprandial glucose, insulin, and free fatty acid responses to sucrose consumed with blackcurrants and lingonberries in healthy women. Am. J. Clin. Nutr. 2012, 96, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, H.-M.; Lindstedt, A.; Järvinen, R.; Sinkkonen, J.; Graça, G.; Viitanen, M.; Kallio, H.; Gil, A.M. 1H NMR-based metabolic fingerprinting of urine metabolites after consumption of lingonberries (Vaccinium vitis-idaea) with a high-fat meal. Food Chem. 2013, 138, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Hassimotto, N.M.A.; Lajolo, F.M. Antioxidant status in rats after long-term intake of anthocyanins and ellagitannins from blackberries. J. Sci. Food Agric. 2010, 91, 523–531. [Google Scholar] [CrossRef]
- Wu, T.; Gao, Y.; Guo, X.; Zhang, M.; Gong, L. Blackberry and blueberry anthocyanin supplementation counteract high-fat-diet-induced obesity by alleviating oxidative stress and inflammation and accelerating energy expenditure. Oxidative Med. Cell. Longev. 2018, 2018, 1–9. [Google Scholar] [CrossRef]
- Kim, B.; Gil Lee, S.; Park, Y.-K.; Ku, C.S.; Pham, T.X.; Wegner, C.J.; Yang, Y.; Koo, S.I.; Chun, O.K.; Lee, J.Y. Blueberry, blackberry, and blackcurrant differentially affect plasma lipids and pro-inflammatory markers in diet-induced obesity mice. Nutr. Res. Pract. 2016, 10, 494–500. [Google Scholar] [CrossRef]
- Meireles, M.; Rodríguez-Alcalá, L.M.; Marques, C.; Norberto, S.; Freitas, J.; Fernandes, I.; Mateus, N.; Gomes, A.; Faria, A.; Calhau, C. Effect of chronic consumption of blackberry extract on high-fat induced obesity in rats and its correlation with metabolic and brain outcomes. Food Funct. 2016, 7, 127–139. [Google Scholar] [CrossRef]
- Azofeifa, G.; Quesada, S.; Navarro, L.; Hidalgo, O.; Portet, K.; Pérez, A.M.; Vaillant, F.; Poucheret, P.; Michel, A. Hypoglycaemic, hypolipidaemic and antioxidant effects of blackberry beverage consumption in streptozotocin-induced diabetic rats. J. Funct. Foods 2016, 26, 330–337. [Google Scholar] [CrossRef]
- Kosmala, M.; Jurgoński, A.; Juskiewicz, J.; Karlińska, E.; Macierzynski, J.; Rój, E.; Zduńczyk, Z. Chemical composition of blackberry press cake, polyphenolic extract, and defatted seeds, and their effects on cecal fermentation, bacterial metabolites, and blood lipid profile in rats. J. Agric. Food Chem. 2017, 65, 5470–5479. [Google Scholar] [CrossRef]
- Salyer, J.; Park, S.H.; Ricke, S.C.; Lee, S.O. Analysis of microbial populations and metabolism of anthocyanins by mice gut microflora fed with blackberry powder. J. Nutr. Food Sci. 2013, 3. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, D.J.A.; Srichaikul, K.; Kendall, C.W.C.; Sievenpiper, J.L.; Abdulnour, S.; Mirrahimi, A.; Meneses, C.; Nishi, S.; He, X.; Lee, S.; et al. The relation of low glycaemic index fruit consumption to glycaemic control and risk factors for coronary heart disease in type 2 diabetes. Diabetologia 2010, 54, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Hassimotto, N.M.A.; Pinto, M.D.S.; Lajolo, F.M. Antioxidant status in humans after consumption of blackberry (Rubus fruticosusL.) juices with and without defatted milk. J. Agric. Food Chem. 2008, 56, 11727–11733. [Google Scholar] [CrossRef] [PubMed]
- Vafa, M.; Aghababaee, S.K.; Shidfar, F.; Tahavorgar, A.; Gohari, M.; Katebi, D.; Mohammadi, V. Effects of blackberry (Morus nigra L.) consumption on serum concentration of lipoproteins, apo A-I, apo B, and high-sensitivity-C-reactive protein and blood pressure in dyslipidemic patients. J. Res. Med. Sci. 2015, 20, 684–691. [Google Scholar] [CrossRef] [PubMed]
- Mandave, P.C.; Khadke, S.; Karandikar, M.; Pandit, V.; Ranjekar, P.; Kuvalekar, A.; Mantri, N. Antidiabetic, lipid normalizing, and nephroprotective actions of the strawberry: A potent supplementary fruit. Int. J. Mol. Sci. 2017, 18, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oviedo-Solís, C.I.; Sandoval-Salazar, C.; Lozoya, E.; Maldonado-Aguilera, G.A.; Aguilar-Zavala, H.; Beltrán-Campos, V.; Pérez-Vázquez, V.; Ramírezemiliano, J. Ultraviolet light-C increases antioxidant capacity of the strawberry (Fragaria x ananassa) in vitro and in high-fat diet-induced obese rats. Food Sci. Nutr. 2017, 5, 1004–1014. [Google Scholar] [CrossRef]
- Parelman, M.A.; Storms, D.H.; Kirschke, C.P.; Huang, L.; Zunino, S.J. Dietary strawberry powder reduces blood glucose concentrations in obese and lean C57BL/6 mice, and selectively lowers plasma C-reactive protein in lean mice. Br. J. Nutr. 2012, 108, 1789–1799. [Google Scholar] [CrossRef] [Green Version]
- Aranaz, P.; Romo-Hualde, A.; Zabala, M.; Navarro-Herrera, D.; De Galarreta, M.R.; Gil, A.G.; Martínez, J.A.; Aranaz, P.; Gonzalez, C.J. Freeze-dried strawberry and blueberry attenuates diet-induced obesity and insulin resistance in rats by inhibiting adipogenesis and lipogenesis. Food Funct. 2017, 8, 3999–4013. [Google Scholar] [CrossRef]
- Diamanti, J.; Mezzetti, B.; Giampieri, F.; Alvarez-Suarez, J.M.; Quiles, J.L.; Gonzalez-Alonso, A.; Ramirez-Tortosa, M.D.C.; Granados-Principal, S.; Gonzáles-Paramás, A.M.; Santos-Buelga, C.; et al. Doxorubicin-induced oxidative stress in rats is efficiently counteracted by dietary anthocyanin differently enriched strawberry (Fragaria×ananassaDuch.). J. Agric. Food Chem. 2014, 62, 3935–3943. [Google Scholar] [CrossRef]
- Sandoval-Salazar, C.; Oviedo-Solís, C.I.; Lozoya, E.; Aguilar-Zavala, H.; Solís-Ortiz, S.; Pérez-Vázquez, V.; Balcón-Pacheco, C.D.; Ramírezemiliano, J. Strawberry intake ameliorates oxidative stress and decreases GABA levels induced by high-fat diet in frontal cortex of rats. Antioxidants 2019, 8, 70. [Google Scholar] [CrossRef] [Green Version]
- Petersen, C.; Wankhade, U.D.; Bharat, D.; Wong, K.; Mueller, J.E.; Chintapalli, S.V.; Piccolo, B.D.; Jalili, T.; Jia, Z.; Symons, J.D.; et al. Dietary supplementation with strawberry induces marked changes in the composition and functional potential of the gut microbiome in diabetic mice. J. Nutr. Biochem. 2019, 66, 63–69. [Google Scholar] [CrossRef]
- Han, Y.; Song, M.; Gu, M.; Ren, D.; Zhu, X.; Cao, X.; Li, F.; Wang, W.; Cai, X.; Yuan, B.; et al. Dietary intake of whole strawberry inhibited colonic inflammation in dextran-sulfate-sodium-treated mice via restoring immune homeostasis and alleviating gut microbiota dysbiosis. J. Agric. Food Chem. 2019, 67, 9168–9177. [Google Scholar] [CrossRef]
- Paquette, M.; Larqué, A.S.M.; Weisnagel, S.J.; Desjardins, Y.; Marois, J.; Pilon, G.; Dudonné, S.; Marette, A.; Jacques, H. Strawberry and cranberry polyphenols improve insulin sensitivity in insulin-resistant, non-diabetic adults: A parallel, double-blind, controlled and randomised clinical trial. Br. J. Nutr. 2017, 117, 519–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton-Freeman, B.; Linares, A.; Hyson, D.; Kappagoda, T. Strawberry modulates LDL oxidation and postprandial lipemia in response to high-fat meal in overweight hyperlipidemic men and women. J. Am. Coll. Nutr. 2010, 29, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Kurotobi, T.; Fukuhara, K.; Inage, H.; Kimura, S. Glycemic index and postprandial blood glucose response to Japanese strawberry jam in normal adults. J. Nutr. Sci. Vitaminol. 2010, 56, 198–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibero-Baraibar, I.; Cuervo, M.; Navas-Carretero, S.; Abete, I.; Zulet, M.A.; Martínez, J.A. Different postprandial acute response in healthy subjects to three strawberry jams varying in carbohydrate and antioxidant content: A randomized, crossover trial. Eur. J. Nutr. 2013, 53, 201–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amani, R.; Moazen, S.; Shahbazian, H.; Ahmadi, K.; Jalali, M.T. Flavonoid-rich beverage effects on lipid profile and blood pressure in diabetic patients. World J. Diabetes 2014, 5, 962–968. [Google Scholar] [CrossRef]
- Ellis, C.L.; Edirisinghe, I.; Kappagoda, T.; Burton-Freeman, B. Attenuation of meal-induced inflammatory and thrombotic responses in overweight men and women after 6-week daily strawberry (Fragaria) intake. J. Atheroscler. Thromb. 2011, 18, 318–327. [Google Scholar] [CrossRef] [Green Version]
- Edirisinghe, I.; Banaszewski, K.; Cappozzo, J.; Sandhya, K.; Ellis, C.L.; Tadapaneni, R.K.; Kappagoda, C.T.; Burton-Freeman, B.M. Strawberry anthocyanin and its association with postprandial inflammation and insulin. Br. J. Nutr. 2011, 106, 913–922. [Google Scholar] [CrossRef]
- Park, E.; Edirisinghe, I.; Wei, H.; Vijayakumar, L.P.; Banaszewski, K.; Cappozzo, J.C.; Burton-Freeman, B. A dose-response evaluation of freeze-dried strawberries independent of fiber content on metabolic indices in abdominally obese individuals with insulin resistance in a randomized, single-blinded, diet-controlled crossover trial. Mol. Nutr. Food Res. 2016, 60, 1099–1109. [Google Scholar] [CrossRef]
- Ivey, K.L.; Chan, A.T.; Izard, J.; Cassidy, A.; Rogers, G.B.; Rimm, E.B. Role of dietary flavonoid compounds in driving patterns of microbial community assembly. mBio 2019, 10, e01205-19. [Google Scholar] [CrossRef] [Green Version]
- Basu, A.; Morris, S.; Nguyen, A.; Betts, N.M.; Fu, D.; Lyons, T.J. Effects of dietary strawberry supplementation on antioxidant biomarkers in obese adults with above optimal serum lipids. J. Nutr. Metab. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moazen, S.; Amani, R.; Rad, A.H.; Ahmadi, K.; Jalali, M.-T.; Shahbazian, H. Effects of freeze-dried strawberry supplementation on metabolic biomarkers of atherosclerosis in subjects with Type 2 Diabetes: A randomized double-blind controlled trial. Ann. Nutr. Metab. 2013, 63, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Cai, Y.; Yan, J.; Sun, M.; Corke, H. Hypoglycemic and hypolipidemic effects and antioxidant activity of fruit extracts from Lycium barbarum. Life Sci. 2004, 76, 137–149. [Google Scholar] [CrossRef]
- Cui, B.K.; Liu, S.; Lin, X.; Wang, J.; Li, S.H.; Wang, Q.-B.; Li, S. Effects of Lycium barbarum aqueous and ethanol extracts on high-fat-diet induced oxidative stress in rat liver tissue. Molecules 2011, 16, 9116–9128. [Google Scholar] [CrossRef] [PubMed]
- Jing, L.; Cui, G.; Feng, Q.; Xiao, Y. Evaluation of hypoglycemic activity of the polysaccharides extracted from Lycium Barbarum. Afr. J. Tradit. Complement. Altern. Med. 2009, 6, 579–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Wu, T. Anthocyanins from black wolfberry (Lycium ruthenicum Murr.) prevent inflammation and increase fecal fatty acid in diet-induced obese rats. RSC Adv. 2017, 7, 47848–47853. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Li, Q.W.; Li, J.; Zhang, T. Protective effect of Lycium barbarum polysaccharide 4 on kidneys in streptozotocin-induced diabetic rats. Can. J. Physiol. Pharmacol. 2009, 87, 711–719. [Google Scholar] [CrossRef]
- Hu, C.; Lee, Y.J.; Colitz, C.M.; Chang, C.J.; Lin, C.T. The protective effects of Lycium barbarum and Chrysanthemum morifolum on diabetic retinopathies in rats. Veter. Ophthalmol. 2012, 15, 65–71. [Google Scholar] [CrossRef]
- Zhao, R.; Gao, X.; Zhang, T.; Li, X. Effects of Lycium barbarum. polysaccharide on type 2 diabetes mellitus rats by regulating biological rhythms. Iran. J. Basic Med. Sci 2016, 19, 1024–1030. [Google Scholar]
- Al-Fartosy, A.J.M. Protective effect of galactomannan extracted from Iraqi Lycium Barbarum L. fruits against alloxan-induced diabetes in rats. Am. J. Biochem. Biotechnol. 2015, 11, 73–83. [Google Scholar] [CrossRef]
- Du, M.; Hu, X.; Kou, L.; Zhang, B.; Zhang, C. Lycium barbarum Polysaccharide mediated the antidiabetic and antinephritic effects in diet-streptozotocin-induced diabetic sprague dawley rats via regulation of NF-κB. BioMed Res. Int. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ni, T.; Wei, G.; Yin, X.; Liu, X.; Liu, D. Original article Neuroprotective effect of Lycium barbarum on retina of Royal College of Surgeons (RCS) rats: A preliminary study. Folia Neuropathol. 2013, 2, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Yang, G.; Zhang, S.; Ross, C.F.; Zhu, M. Goji berry modulates gut microbiota and alleviates colitis in IL-10-deficient mice. Mol. Nutr. Food Res. 2018, 62, 1800535. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Zhou, S.; Liu, J.; McLean, R.J.; Chu, W. Prebiotic, immuno-stimulating and gut microbiota-modulating effects of Lycium barbarum polysaccharide. Biomed. Pharmacother. 2020, 121, 109591. [Google Scholar] [CrossRef]
- Amagase, H.; Sun, B.; Borek, C. Lycium barbarum (goji) juice improves in vivo antioxidant biomarkers in serum of healthy adults. Nutr. Res. 2009, 29, 19–25. [Google Scholar] [CrossRef]
- Zanchet, M.Z.D.S.; Nardi, G.M.; Bratti, L.D.O.S.; Filippin, F.B.; Locatelli, C. Lycium Barbarum reduces abdominal fat and improves lipid profile and antioxidant status in patients with metabolic syndrome. Oxidative Med. Cell. Longev. 2017, 2017, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Liong, E.C.; Ching, Y.P.; Chang, R.C.C.; So, K.F.; Fung, M.L.; Tipoe, G.L. Lycium barbarum polysaccharides protect mice liver from carbon tetrachloride-induced oxidative stress and necroinflammation. J. Ethnopharmacol. 2012, 139, 462–470. [Google Scholar] [CrossRef]
- Driessche, J.J.V.D.; Plat, J.; Plasqui, G.; Mensink, R.P. A single dose of goji berries does not affect postprandial energy expenditure and substrate oxidation in healthy, overweight men. J. Nutr. Metab. 2019, 2019, 1–6. [Google Scholar] [CrossRef]
- De Oliveira, P.R.B.; Da Costa, C.A.; De Bem, G.F.; Cordeiro, V.S.C.; Santos, I.B.; De Carvalho, L.C.R.M.; Conceição, E.P.; Lisboa, P.C.; Ognibene, D.T.; Sousa, P.J.C.; et al. Euterpe oleracea mart.-derived polyphenols protect mice from diet-induced obesity and fatty liver by regulating hepatic lipogenesis and cholesterol Excretion. PLoS ONE 2015, 10, e0143721. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Hong, J.-H.; Jung, H.K.; Jeong, Y.S.; Cho, K.H. Grape skin and loquat leaf extracts and acai puree have potent anti-atherosclerotic and anti-diabetic activity in vitro and in vivo in hypercholesterolemic zebrafish. Int. J. Mol. Med. 2012, 30, 606–614. [Google Scholar] [CrossRef]
- Sun, X.; Seeberger, J.; Alberico, T.; Wang, C.; Wheeler, C.T.; Schauss, A.G.; Zou, S. Açai palm fruit (Euterpe oleracea Mart.) pulp improves survival of flies on a high fat diet. Exp. Gerontol. 2010, 45, 243–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Castro, C.A.; Natali, A.J.; Cardoso, L.M.; Ferreira-Machado, A.B.; Novello, A.A.; Da Silva, K.A.; Tafuri, N.F.; Da Matta, S.L.P.; Pedrosa, M.L.; Peluzio, M.D.C.G. Aerobic exercise and not a diet supplemented with jussara açaí (Euterpe edulis Martius) alters hepatic oxidative and inflammatory biomarkers in ApoE-deficient mice. Br. J. Nutr. 2014, 112, 285–294. [Google Scholar] [CrossRef] [Green Version]
- Cordeiro, V.D.S.C.; De Bem, G.F.; Da Costa, C.A.; Santos, I.B.; De Carvalho, L.C.R.M.; Ognibene, D.T.; Da Rocha, A.P.M.; De Carvalho, J.J.; De Moura, R.S.; Resende, A.C. Euterpe oleracea Mart. seed extract protects against renal injury in diabetic and spontaneously hypertensive rats: Role of inflammation and oxidative stress. Eur. J. Nutr. 2017, 57, 817–832. [Google Scholar] [CrossRef]
- De Souza, M.O.; e Silva, L.S.; Magalhães, C.L.D.B.; De Figueiredo, B.B.; Costa, D.C.; Silva, M.E.; Pedrosa, M.L. The hypocholesterolemic activity of açaí (Euterpe oleracea Mart.) is mediated by the enhanced expression of the ATP-binding cassette, subfamily G transporters 5 and 8 and low-density lipoprotein receptor genes in the rat. Nutr. Res. 2012, 32, 976–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bem, G.F.; Costa, C.A.; Santos, I.B.; Cordeiro, V.D.S.C.; De Carvalho, L.C.R.M.; De Souza, M.A.V.; Soares, R.D.A.; Sousa, P.J.D.C.; Ognibene, D.T.; Resende, A.C.; et al. Antidiabetic effect of Euterpe oleracea Mart. (açaí) extract and exercise training on high-fat diet and streptozotocin-induced diabetic rats: A positive interaction. PLoS ONE 2018, 13, e0199207. [Google Scholar] [CrossRef] [PubMed]
- Guergoletto, K.B.; Costabile, A.; Flores, G.; Garcia, S.; Gibson, G.R. In vitro fermentation of juçara pulp (Euterpe edulis) by human colonic microbiota. Food Chem. 2016, 196, 251–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AlQurashi, R.M.; Alarifi, S.N.; Walton, G.E.; Costabile, A.; Rowland, I.R.; Commane, D.M. In vitro approaches to assess the effects of açai (Euterpe oleracea) digestion on polyphenol availability and the subsequent impact on the faecal microbiota. Food Chem. 2017, 234, 190–198. [Google Scholar] [CrossRef]
- Guerra, J.F.D.C.; Maciel, P.S.; De Abreu, I.C.M.E.; Pereira, R.R.; Silva, M.; Cardoso, L.D.M.; Pinheiro-Sant’Ana, H.M.; Lima, W.G.; Silva, M.E.; Pedrosa, M.L. Dietary açai attenuates hepatic steatosis via adiponectin-mediated effects on lipid metabolism in high-fat diet mice. J. Funct. Foods 2015, 14, 192–202. [Google Scholar] [CrossRef]
- Mertens-Talcott, S.U.; Rios, J.; Jilma-Stohlawetz, P.; Pacheco-Palencia, L.A.; Meibohm, B.; Talcott, S.T.; Derendorf, H. Pharmacokinetics of anthocyanins and antioxidant effects after the consumption of anthocyanin-rich açai juice and Pulp (Euterpe oleracea Mart.) in human healthy volunteers. J. Agric. Food Chem. 2008, 56, 7796–7802. [Google Scholar] [CrossRef]
- Udani, J.; Singh, B.B.; Singh, V.J.; Barrett, M.L. Effects of Açai (Euterpe oleracea Mart.) berry preparation on metabolic parameters in a healthy overweight population: A pilot study. Nutr. J. 2011, 10, 45. [Google Scholar] [CrossRef] [Green Version]
- Yamane, T.; Kozuka, M.; Konda, D.; Nakano, Y.; Nakagaki, T.; Ohkubo, I.; Ariga, H. Improvement of blood glucose levels and obesity in mice given aronia juice by inhibition of dipeptidyl peptidase IV and α-glucosidase. J. Nutr. Biochem. 2016, 31, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.D.; Kang, S.H.; Moon, K.H.; Lee, J.H.; Kim, D.G.; Kim, W.; Kim, J.S.; Ahn, B.Y.; Jin, J.S. the effect of aronia berry on Type 1 Diabetes in vivo and in vitro. J. Med. Food 2018, 21, 244–253. [Google Scholar] [CrossRef]
- Park, C.H.; Kim, J.H.; Lee, E.B.; Hur, W.; Kwon, O.-J.; Park, H.-J.; Yoon, S.K. Aronia melanocarpa extract ameliorates hepatic lipid metabolism through PPARγ2 downregulation. PLoS ONE 2017, 12, e0169685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Baek, S.-K.; Bin Song, K. Physical and antioxidant properties of alginate films prepared from Sargassum fulvellum with black chokeberry extract. Food Packag. Shelf Life 2018, 18, 157–163. [Google Scholar] [CrossRef]
- Valcheva-Kuzmanova, S.; Kuzmanov, K.; Tancheva, S.; Belcheva, A. Hypoglycemic effects of Aronia melanocarpa fruit juice in streptozotocin-induced diabetic rats. Methods Find. Exp. Clin. Pharmacol. 2007, 29, 101–105. [Google Scholar] [CrossRef]
- Lipinska, P.; Atanasov, A.G.; Palka, M.; Jóźwik, A. Chokeberry pomace as a determinant of antioxidant parameters assayed in blood and liver tissue of polish merino and Wrzosówka lambs. Molecules 2017, 22, 1461. [Google Scholar] [CrossRef] [Green Version]
- Sikora, J.; Broncel, M.; Markowicz, M.; Chałubiński, M.; Wojdan, K.; Mikiciuk-Olasik, E. Short-term supplementation with Aronia melanocarpa extract improves platelet aggregation, clotting, and fibrinolysis in patients with metabolic syndrome. Eur. J. Nutr. 2011, 51, 549–556. [Google Scholar] [CrossRef] [Green Version]
- Jurgoński, A.; Juskiewicz, J.; Zduńczyk, Z. Ingestion of black chokeberry fruit extract leads to intestinal and systemic changes in a rat model of prediabetes and hyperlipidemia. Plant. Foods Hum. Nutr. 2008, 63, 176–182. [Google Scholar] [CrossRef]
- Kujawska, M.; Ignatowicz, E.; Ewertowska, M.; Oszmiański, J.; Jodynis-Liebert, J. Protective effect of chokeberry on chemical-induced oxidative stress in rat. Hum. Exp. Toxicol. 2010, 30, 199–208. [Google Scholar] [CrossRef]
- Bhaswant, M.; Shafie, S.R.; Mathai, M.L.; Mouatt, P.; Brown, L. Anthocyanins in chokeberry and purple maize attenuate diet-induced metabolic syndrome in rats. Nutrients 2017, 41, 24–31. [Google Scholar] [CrossRef]
- Qin, B.; Anderson, R.A. An extract of chokeberry attenuates weight gain and modulates insulin, adipogenic and inflammatory signalling pathways in epididymal adipose tissue of rats fed a fructose-rich diet. Br. J. Nutr. 2011, 108, 581–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakovljevic, V.; Milic, P.; Bradic, J.; Jeremic, J.; Zivkovic, V.; Srejovic, I.M.; Turnic, T.R.N.; Milosavljevic, I.; Jeremic, N.; Bolevich, S.; et al. Standardized aronia melanocarpa extract as novel supplement against metabolic syndrome: A rat model. Int. J. Mol. Sci. 2018, 20, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milutinović, M.; Branković, S.; Ćujić, N.; Šavikin, K.; Kostić, M.; Kitić, N.; Miladinović, B.; Kitić, D. Antispasmodic effects of black chokeberry (Aronia melanocarpa (Michx.) Elliott) extracts and juice and their potential use in gastrointestinal disorders. J. Berry Res. 2020, 10, 175–192. [Google Scholar] [CrossRef]
- Kardum, N.; Konić-Ristić, A.; Šavikin, K.; Spasic, S.; Stefanovic, A.; Ivanisevic, J.; Miljkovic, M. Effects of polyphenol-rich chokeberry juice on antioxidant/pro-oxidant status in healthy subjects. J. Med. Food 2014, 17, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Kardum, N.; Takić, M.; Šavikin, K.; Zec, M.; Zdunić, G.; Spasic, S.; Konić-Ristić, A. Effects of polyphenol-rich chokeberry juice on cellular antioxidant enzymes and membrane lipid status in healthy women. J. Funct. Foods 2014, 9, 89–97. [Google Scholar] [CrossRef]
- Broncel, M.; Kozirog, M.; Duchnowicz, P.; Koter-Michalak, M.; Sikora, J.; Chojnowska-Jezierska, J. Aronia melanocarpa extract reduces blood pressure, serum endothelin, lipid, and oxidative stress marker levels in patients with metabolic syndrome. Med. Sci. Monit. 2010, 16. [Google Scholar]
- Kardum, N.; Milovanović, B.; Šavikin, K.; Zdunić, G.; Mutavdžin, S.; Gligorijević, T.; Spasic, S. Beneficial effects of polyphenol-rich chokeberry juice consumption on blood pressure level and lipid status in hypertensive subjects. J. Med. Food 2015, 18, 1231–1238. [Google Scholar] [CrossRef]
- Badescu, M.; Badulescu, O.; Badescu, L.; Ciocoiu, M. Effects ofSambucus nigraandAronia melanocarpaextracts on immune system disorders within diabetes mellitus. Pharm. Biol. 2014, 53, 533–539. [Google Scholar] [CrossRef] [Green Version]
- Nowak, D.; Gośliński, M.; Przygoński, K.; Wojtowicz, E. The antioxidant properties of exotic fruit juices from acai, maqui berry and noni berries. Eur. Food Res. Technol. 2018, 244, 1897–1905. [Google Scholar] [CrossRef]
- Skoczynska, A.; Poręba, R.; Steinmentz-Beck, A.; Martynowicz, H.; Affelska-Jercha, A.; Turczyn, B.; Wojakowska, A.; Jedrychowska, I. The dependence between urinary mercury concentration and carotid arterial intima-media thickness in workers occupationally exposed to mercury vapour. Int. J. Occup. Med. Environ. Health 2009, 22. [Google Scholar] [CrossRef] [Green Version]
- Istas, G.; Wood, E.; Le Sayec, M.; Rawlings, C.; Yoon, J.; Dandavate, V.; Cera, E.D.; Rampelli, S.; Costabile, A.; Fromentin, E.; et al. Effects of aronia berry (poly)phenols on vascular function and gut microbiota: A double-blind randomized controlled trial in adult men. Am. J. Clin. Nutr. 2019, 110, 316–329. [Google Scholar] [CrossRef]
- Tani, T.; Nishikawa, S.; Kato, M.; Tsuda, T. Delphinidin 3-rutinoside-rich blackcurrant extract ameliorates glucose tolerance by increasing the release of glucagon-like peptide-1 secretion. Food Sci. Nutr. 2017, 5, 929–933. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, Y.; Ozeki, A.; Tani, T.; Tsuda, T. Blackcurrant extract ameliorates hyperglycemia in Type 2 Diabetic mice in association with increased basal secretion of glucagon-like peptide-1 and activation of AMP-activated protein kinase. J. Nutr. Sci. Vitaminol. 2018, 64, 258–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, D.; Damsud, T.; Wilson, M.; Grace, M.H.; Strauch, R.; Li, X.; Lila, M.A.; Komarnytsky, S. Black currant anthocyanins attenuate weight gain and improve glucose metabolism in diet-induced obese mice with intact, but not disrupted, gut microbiome. J. Agric. Food Chem. 2015, 63, 6172–6180. [Google Scholar] [CrossRef]
- Benn, T.; Kim, B.; Park, Y.K.; Wegner, C.J.; Harness, E.; Nam, T.G.; Kim, D.O.; Lee, J.S.; Lee, J.Y. Polyphenol-rich blackcurrant extract prevents inflammation in diet-induced obese mice. J. Nutr. Biochem. 2014, 25, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Jurgoński, A.; Juskiewicz, J.; Zduńczyk, Z.; Matusevicius, P.; Kołodziejczyk, K. Polyphenol-rich extract from blackcurrant pomace attenuates the intestinal tract and serum lipid changes induced by a high-fat diet in rabbits. Eur. J. Nutr. 2014, 53, 1603–1613. [Google Scholar] [CrossRef] [Green Version]
- Molan, A.; Liu, Z.; Kruger, M. The ability of blackcurrant extracts to positively modulate key markers of gastrointestinal function in rats. World, J. Microbiol. Biotechnol. 2010, 26, 1735–1743. [Google Scholar] [CrossRef]
- Park, J.H.; Kho, M.C.; Kim, H.Y.; Ahn, Y.M.; Lee, Y.J.; Kang, D.G.; Lee, H.S. Blackcurrant suppresses metabolic syndrome induced by high-fructose diet in rats. Evid. Based Complement. Altern. Med. 2015, 2015, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Paturi, G.; Butts, C.A.; Monro, J.A.; Hedderley, D.I. Effects of blackcurrant and dietary fibers on large intestinal health biomarkers in rats. Plant. Foods Hum. Nutr. 2018, 73, 54–60. [Google Scholar] [CrossRef]
- Molan, A.; Liu, Z.; Plimmer, G. Evaluation of the effect of blackcurrant products on gut microbiota and on markers of risk for colon cancer in humans. Phytother. Res. 2013, 28, 416–422. [Google Scholar] [CrossRef]
- Castro-Acosta, M.L.; Stone, S.G.; Mok, J.E.; Mhajan, R.K.; Fu, C.I.; Lenihan-Geels, G.N.; Corpe, C.P.; Hall, W. Apple and blackcurrant polyphenol-rich drinks decrease postprandial glucose, insulin and incretin response to a high-carbohydrate meal in healthy men and women. J. Nutr. Biochem. 2017, 49, 53–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Acosta, M.L.; Smith, L.; Miller, R.J.; McCarthy, D.I.; Farrimond, J.A.; Hall, W. Drinks containing anthocyanin-rich blackcurrant extract decrease postprandial blood glucose, insulin and incretin concentrations. J. Nutr. Biochem. 2016, 38, 154–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willems, M.; Parktin, N.; Widjaja, W.; Ajjimaporn, A. Effect of New Zealand blackcurrant extract on physiological responses at rest and during brisk walking in southeast asian men: A randomized, double-blind, placebo-controlled, crossover study. Nutrients 2018, 10, 1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strauss, J.A.; Willems, M.E.; Shepherd, S.O. New Zealand blackcurrant extract enhances fat oxidation during prolonged cycling in endurance-trained females. Eur. J. Appl. Physiol. 2018, 118, 1265–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lomiwes, D.; Ha, B.; Ngametua, N.; Burr, N.; Cooney, J.M.; Trower, T.; Sawyer, G.M.; Hedderley, D.I.; Hurst, R.D.; Hurst, S.M. Timed consumption of a New Zealand blackcurrant juice support positive affective responses during a self-motivated moderate walking exercise in healthy sedentary adults. J. Int. Soc. Sports Nutr. 2019, 16, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurst, R.D.; Lyall, K.A.; Roberts, J.M.; Perthaner, A.; Wells, R.W.; Cooney, J.M.; Jensen, D.J.; Burr, N.; Hurst, S.M. Consumption of an anthocyanin-rich extract made from new zealand blackcurrants prior to exercise may assist recovery from oxidative stress and maintains circulating neutrophil function: A pilot study. Front. Nutr. 2019, 6, 73. [Google Scholar] [CrossRef]
- Alvarado, J.L.; Leschot, A.; Olivera-Nappa, Á.; Salgado, A.-M.; Rioseco, H.; Lyon, C.; Vigil, P. Delphinidin-rich maqui berry extract (Delphinol®) lowers fasting and postprandial glycemia and insulinemia in prediabetic individuals during oral glucose tolerance tests. BioMed Res. Int. 2016, 2016, 9070537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Lorenzo, A.; Sobolev, A.P.; Nabavi, S.; Sureda, A.; Moghaddam, A.; Khanjani, S.; Di Giovanni, C.; Xiao, J.; Shirooie, S.; Sokeng, A.J.T.; et al. Corrigendum to Antidepressive effects of a chemically characterized maqui berry extract (Aristotelia chilensis (molina) stuntz) in a mouse model of Post-stroke depression. Food Chem. Toxicol. 2019, 131, 110641. [Google Scholar] [CrossRef]
- Rojo, L.E.; Ribnicky, D.; Logendra, S.; Poulev, A.; Rojas-Silva, P.; Kuhn, P.; Dorn, R.; Grace, M.H.; Lila, M.A.; Raskin, I. In vitro and in vivo anti-diabetic effects of anthocyanins from maqui berry (Aristotelia chilensis). Food Chem. 2011, 131, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Davinelli, S.; Bertoglio, J.C.; Zarrelli, A.; Pina, R.; Scapagnini, G. A Randomized clinical trial evaluating the efficacy of an anthocyanin–maqui berry extract (Delphinol®) on oxidative stress biomarkers. J. Am. Coll. Nutr. 2015, 34, 28–33. [Google Scholar] [CrossRef] [Green Version]
- Parra-Vargas, M.; Sandoval-Rodríguez, A.S.; Rodriguez-Echevarria, R.; Dominguez-Rosales, J.A.; Santos-Garcia, A.; Armendariz-Borunda, J. Delphinidin ameliorates hepatic triglyceride accumulation in human hepg2 cells, but not in diet-induced obese mice. Nutrients 2018, 10, 1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corsi, R.; Mosti, G.; Cavezzi, A.; Urso, S.U.; Dimitrova, G.; Fioroni, E.; Colucci, R.; Quinzi, V. A polyphenol-based multicomponent nutraceutical in dysmetabolism and oxidative stress: Results from a pilot study. J. Diet. Suppl. 2017, 15, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Ávila, F.; Jiménez-Aspee, F.; Cruz, N.; Gómez, C.; González, M.A.; Ravello, N. Additive effect of maqui (Aristotelia chilensis) and lemon (Citrus x limon) juice in the postprandial glycemic responses after the intake of high glycemic index meals in healthy men. NFS J. 2019, 17, 8–16. [Google Scholar] [CrossRef]
- Sandoval, V.; Femenias, A.; Martínez-Garza, Ú.; Sanz-Lamora, H.; Castagnini, J.M.; Quifer-Rada, P.; Lamuela-Raventós, R.-M.; Marrero, P.F.; Haro, D.; Relat, J. Lyophilized maqui (Aristotelia chilensis) berry induces browning in the subcutaneous white adipose tissue and ameliorates the insulin resistance in high fat diet-induced obese mice. Antioxidants 2019, 8, 360. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Yagiz, Y.; Buran, T.J.; Nunes, C.D.N.; Gu, L. Phytochemicals from berries and grapes inhibited the formation of advanced glycation end-products by scavenging reactive carbonyls. Food Res. Int. 2011, 44, 2666–2673. [Google Scholar] [CrossRef]
- Barrett, A.; Ndou, T.; Hughey, C.A.; Straut, C.; Howell, A.; Dai, Z.; Kaletunc, G. Inhibition of α-amylase and glucoamylase by tannins extracted from cocoa, pomegranates, cranberries, and grapes. J. Agric. Food Chem. 2013, 61, 1477–1486. [Google Scholar] [CrossRef]
- Pinto, M.D.S.; Ghaedian, R.; Shinde, R.; Shetty, K. Potential of cranberry powder for management of hyperglycemia using in vitro models. J. Med. Food 2010, 13, 1036–1044. [Google Scholar] [CrossRef]
- Pappas, E.; Schaich, K.M. Phytochemicals of cranberries and cranberry products: Characterization, potential health effects, and processing stability. Crit. Rev. Food Sci. Nutr. 2009, 49, 741–781. [Google Scholar] [CrossRef]
- Gulcin, I.; Topal, F.; Çakmakçı, R.; Bílsel, M.; Goren, A.C.; Erdogan, U.; Cakmakci, R. Pomological features, nutritional quality, polyphenol content analysis, and antioxidant properties of domesticated and 3 wild ecotype forms of raspberries (Rubus idaeus L.). J. Food Sci. 2011, 76, C585–C593. [Google Scholar] [CrossRef]
- Sparzak, B. HPLC analysis of polyphenols in the fruits of Rubus idaeus L. (Rosaceae). Nat. Prod. Res. 2010, 24, 1811–1822. [Google Scholar] [CrossRef]
- Krauze-Baranowska, M.; Głód, D.; Kula, M.; Majdan, M.; Hałasa, R.; Matkowski, A.; Kozłowska, W.; Kawiak, A. chemical composition and biological activity of Rubus idaeus shoots—A traditional herbal remedy of Eastern Europe. BMC Complement. Altern. Med. 2014, 14, 480. [Google Scholar] [CrossRef] [PubMed]
- Mena, P.; Tassotti, M.; Martínez, J.J.; Hernández, F.; Del Rio, D.; Sánchez-Salcedo, E.M. Phytochemical evaluation of eight white (Morus alba L.) and black (Morus nigra L.) mulberry clones grown in Spain based on UHPLC-ESI-MSn metabolomic profiles. Food Res. Int. 2016, 89, 1116–1122. [Google Scholar] [CrossRef]
- Natić, M.; Dabić, D.Č.; Papetti, A.; Akšić, M.M.F.; Ognjanov, V.; Ljubojević, M.; Tešić, Ž.L. Analysis and characterisation of phytochemicals in mulberry (Morus alba L.) fruits grown in Vojvodina, North Serbia. Food Chem. 2015, 171, 128–136. [Google Scholar] [CrossRef]
- Jin, Q.; Yang, J.; Ma, L.; Cai, J.; Li, J. Comparison of polyphenol profile and inhibitory activities against oxidation and α-glucosidase in mulberry (Genus Morus) cultivars from China. J. Food Sci. 2015, 80, C2440–C2451. [Google Scholar] [CrossRef]
- Bujor, O.-C.; Giniès, C.; Popa, V.I.; Dufour, C. Phenolic compounds and antioxidant activity of lingonberry (Vaccinium vitis-idaea L.) leaf, stem and fruit at different harvest periods. Food Chem. 2018, 252, 356–365. [Google Scholar] [CrossRef]
- Ieri, F.; Martini, S.; Innocenti, M.; Mulinacci, N. Phenolic distribution in liquid preparations of Vaccinium myrtillus L. and Vaccinium vitis idaea L. Phytochem. Anal. 2013, 24, 467–475. [Google Scholar] [CrossRef]
- Spínola, V.; Pinto, J.; Castilho, P. Hypoglycemic, anti-glycation and antioxidant in vitro properties of two Vaccinium species from Macaronesia: A relation to their phenolic composition. J. Funct. Foods 2018, 40, 595–605. [Google Scholar] [CrossRef]
- Spínola, V.; Pinto, J.; Llorent-Martínez, E.J.; Tomás, H.; Castilho, P. Evaluation of Rubus grandifolius L. (wild blackberries) activities targeting management of type-2 diabetes and obesity using in vitro models. Food Chem. Toxicol. 2019, 123, 443–452. [Google Scholar] [CrossRef]
- Aaby, K.; Ekeberg, D.; Skrede, G. Characterization of phenolic compounds in strawberry (Fragaria × ananassa) fruits by different HPLC detectors and contribution of individual compounds to total antioxidant Capacity. J. Agric. Food Chem. 2007, 55, 4395–4406. [Google Scholar] [CrossRef]
- Šaponjac, V.T.; Gironés-Vilaplana, A.; Djilas, S.; Mena, P.; Ćetković, G.; Moreno, D.A.; Čanadanović-Brunet, J.; Vulić, J.; Stajčić, S.; Vinčić, M. Chemical composition and potential bioactivity of strawberry pomace. RSC Adv. 2015, 5, 5397–5405. [Google Scholar] [CrossRef]
- Wojdyło, A.; Nowicka, P.; Bąbelewski, P. Phenolic and carotenoid profile of new goji cultivars and their anti-hyperglycemic, anti-aging and antioxidant properties. J. Funct. Foods 2018, 48, 632–642. [Google Scholar] [CrossRef]
- Montesano, D.; Rocchetti, G.; Cossignani, L.; Lucini, L.; Simonetti, M.S.; Blasia, F. Italian Lycium barbarum L. berry: Chemical characterization and nutraceutical value. Nat. Prod. Commun. 2018, 13. [Google Scholar] [CrossRef] [Green Version]
- Schauss, A.G.; Wu, X.; Prior, R.L.; Ou, B.; Huang, D.-J.; Owens, J.; Agarwal, A.; Jensen, G.S.; Hart, A.N.; Shanbrom, E. Antioxidant capacity and other bioactivities of the freeze-dried amazonian palm berry, euterpe oleraceaemart. (Acai). J. Agric. Food Chem. 2006, 54, 8604–8610. [Google Scholar] [CrossRef] [PubMed]
- Schauss, A.G.; Wu, X.; Prior, R.L.; Ou, B.; Patel, D.; Huang, D.-J.; Kababick, J.P. Phytochemical and nutrient composition of the freeze-dried amazonian palm berry, euterpe oleraceaemart. (Acai). J. Agric. Food Chem. 2006, 54, 8598–8603. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Palencia, L.A. Absorption and biological activity of phytochemical-rich extracts from açai (Euterpe oleracea Mart.) pulp and oil in vitro. J. Agric. Food Chem. 2008, 56, 3593–3600. [Google Scholar] [CrossRef]
- Sosnowska, D. Inhibitory effect of black chokeberry fruit polyphenols on pancreatic lipase—Searching for most active inhibitors. J. Funct. Foods 2018, 49, 196–204. [Google Scholar] [CrossRef]
- Anttonen, M.J.; Karjalainen, R.O. High-performance liquid chromatography analysis of black currant (Ribes nigrum L.) fruit phenolics grown either conventionally or organically. J. Agric. Food Chem. 2006, 54, 7530–7538. [Google Scholar] [CrossRef]
- Borges, G.; Degeneve, A.; Mullen, W.; Crozier, A. Identification of flavonoid and phenolic antioxidants in black currants, blueberries, raspberries, red currants, and cranberries. J. Agric. Food Chem. 2010, 58, 3901–3909. [Google Scholar] [CrossRef]
- Cespedes, C.L.; Pavon, N.; Dominguez, M.; Alarcon, J.; Balbontin, C.; Kubo, I.; El-Hafidi, M.; Avila, J.G.; Dominguez-Lopez, M. The chilean superfruit black-berry Aristotelia chilensis (Elaeocarpaceae), Maqui as mediator in inflammation-associated disorders. Food Chem. Toxicol. 2017, 108, 438–450. [Google Scholar] [CrossRef]
- Escribano-Bailón, M.T.; Alcalde-Eon, C.; Muñoz, O.; Rivas-Gonzalo, J.C.; Santos-Buelga, C. Anthocyanins in berries of Maqui [Aristotelia chilensis (Mol.) Stuntz]. Phytochem. Anal. 2005, 17, 8–14. [Google Scholar] [CrossRef]
- Häkkinen, S.H.; Kärenlampi, S.O.; Mykkänen, H.M.; Törrönen, A.R. Influence of domestic processing and storage on flavonol contents in berries. J. Agric. Food Chem. 2000, 48, 2960–2965. [Google Scholar] [CrossRef] [PubMed]
- Ferrier, J.; Djeffal, S.; Morgan, H.P.; Kloet, S.P.V.; Redžić, S.; Cuerrier, A.; Balick, M.J.; Arnason, J.T. Antiglycation activity of Vaccinium spp. (Ericaceae) from the Sam Vander Kloet collection for the treatment of type II diabetes1This article is part of a Special Issue entitled ‘A tribute to Sam Vander Kloet FLS: Pure and applied research from blueberries to heathland ecology’. Botany 2012, 90, 401–406. [Google Scholar] [CrossRef]
- Buchert, J.; Koponen, J.M.; Suutarinen, M.; Mustranta, A.; Lille, M.E.; Törrönen, R.; Poutanen, K. Effect of enzyme-aided pressing on anthocyanin yield and profiles in bilberry and blackcurrant juices. J. Sci. Food Agric. 2005, 85, 2548–2556. [Google Scholar] [CrossRef]
- Cermak, R.; Landgraf, S.; Wolffram, S. Quercetin glucosides inhibit glucose uptake into brush-border-membrane vesicles of porcine jejunum. Br. J. Nutr. 2004, 91, 849–855. [Google Scholar] [CrossRef] [Green Version]
- Morazzoni, P.; Livio, S.; Scilingo, A.; Malandrino, S. Vaccinium myrtillus anthocyanosides pharmacokinetics in rats. Arzneimittelforschung 1991, 41, 128–131. [Google Scholar]
- Liu, H.; Liu, H.; Wang, W.; Khoo, C.; Taylor, J.; Gu, L. Cranberry phytochemicals inhibit glycation of human hemoglobin and serum albumin by scavenging reactive carbonyls. Food Funct. 2011, 2, 475. [Google Scholar] [CrossRef] [PubMed]
- Podsędek, A.; Majewska, I.; Redzynia, M.; Sosnowska, D.; Koziolkiewicz, M. In vitro inhibitory effect on digestive enzymes and antioxidant potential of commonly consumed fruits. J. Agric. Food Chem. 2014, 62, 4610–4617. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liu, W.; Ma, H.; Marais, J.P.J.; Khoo, C.; Dain, J.A.; Rowley, D.C.; Seeram, N.P. Effect of cranberry (Vaccinium macrocarpon) oligosaccharides on the formation of advanced glycation end-products. J. Berry Res. 2016, 6, 149–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, T.; Bauer, B.A. Advising consumers about dietary supplements: Lessons from cranberry products. J. Diet. Suppl. 2009, 6, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Rao, L.V.M.; Agrawal, A.; Pendurthi, U.R. Effect of wine phenolics on cytokine-induced C-reactive protein expression. J. Thromb. Haemost. 2007, 5, 1309–1317. [Google Scholar] [CrossRef] [Green Version]
- Marathe, P.H.; Gao, H.X.; Close, K.L. American diabetes association standards of medical care in diabetes 2017. J. Diabetes 2017, 9, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Grussu, D.; Stewart, D.; McDougall, G.J. Berry polyphenols inhibit α-amylasein Vitro: Identifying active components in rowanberry and raspberry. J. Agric. Food Chem. 2011, 59, 2324–2331. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, T.; Ueno, Y.; Yoshikawa, T.; Kojo, H.; Osawa, T. Microarray profiling of gene expression in human adipocytes in response to anthocyanins. Biochem. Pharmacol. 2006, 71, 1184–1197. [Google Scholar] [CrossRef] [PubMed]
- Al-Salmi, F.A. Ameliorative and synergistic effect of red raspberry and lycopene against hepatotoxicity induced by acrylamide in male mice. Int. J. Pharmacol. 2019, 15, 166–176. [Google Scholar] [CrossRef] [Green Version]
- Zou, T.; Kang, Y.; Wang, B.; De Avila, J.M.; You, J.; Zhu, M.-J.; Du, M. Raspberry supplementation reduces lipid accumulation and improves insulin sensitivity in skeletal muscle of mice fed a high-fat diet. J. Funct. Foods 2019, 63, 103572. [Google Scholar] [CrossRef]
- Park, K.S. Raspberry ketone, a naturally occurring phenolic compound, inhibits adipogenic and lipogenic gene expression in 3T3-L1 adipocytes. Pharm. Biol. 2014, 53, 870–875. [Google Scholar] [CrossRef]
- Lee, H.; Kim, J.; An, J.; Lee, S.; Kong, H.; Song, Y.; Choi, H.R.; Lee, S.J.; Chae, K.S.; Kwon, J.W.; et al. Amelioration of hyperglycemia by Rubus occidentalis (black raspberry) and increase in short-chain fatty acids producing bacteria. J. Funct. Foods 2019, 54, 433–439. [Google Scholar] [CrossRef]
- Yan, F.; Chen, Y.; Azat, R.; Zheng, X. Mulberry anthocyanin extract ameliorates oxidative damage in HepG2 cells and prolongs the lifespan of caenorhabditis elegans through MAPK and Nrf2 pathways. Oxidative Med. Cell. Longev. 2017, 2017, 1–12. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Fang, J.; Ruan, Y.; Wang, X.; Sun, Y.; Wu, N.; Zhao, Z.; Chang, Y.; Ning, N.; Guo, H.; et al. Structures, bioactivities and future prospective of polysaccharides from Morus alba (white mulberry): A review. Food Chem. 2018, 245, 899–910. [Google Scholar] [CrossRef]
- He, H.; Yu, W.G.; Yang, J.P.; Ge, S.; Lu, Y.H. Multiple Comparisons of glucokinase activation mechanisms of five mulberry bioactive ingredients in hepatocyte. J. Agric. Food Chem. 2015, 64, 2475–2484. [Google Scholar] [CrossRef]
- Yan, F.; Dai, G.; Zheng, X. Mulberry anthocyanin extract ameliorates insulin resistance by regulating PI3K/AKT pathway in HepG2 cells and db/db mice. J. Nutr. Biochem. 2016, 36, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiang, L.; Wang, C.; Tang, C.; He, X. Antidiabetic and antioxidant effects and phytochemicals of mulberry fruit (Morus alba L.) polyphenol enhanced extract. PLoS ONE 2013, 8, e71144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ma, Z.F.; Luo, X.; Li, X. Effects of mulberry fruit (Morus alba L.) Consumption on health outcomes: A mini-review. Antioxidants 2018, 7, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.; Bao, T.; Lu, Y.; Su, H.; Ke, H.; Chen, W. Polysaccharide from Mulberry Fruit (Morus alba L.) Protects against palmitic-acid-induced hepatocyte lipotoxicity by activating the Nrf2/ARE signaling pathway. J. Agric. Food Chem. 2019. [Google Scholar] [CrossRef]
- Harbilas, D.; Martineau, L.C.; Harris, C.S.; Adeyiwola-Spoor, D.C.; Saleem, A.; Lambert, J.; Caves, D.; Johns, T.; Prentki, M.; Cuerrier, A.; et al. Evaluation of the antidiabetic potential of selected medicinal plant extracts from the Canadian boreal forest used to treat symptoms of diabetes: Part II. Can. J. Physiol. Pharmacol. 2009, 87, 479–492. [Google Scholar] [CrossRef]
- Ross, K.A.; Godfrey, D.; Fukumoto, L. The chemical composition, antioxidant activity and -glucosidase inhibitory activity of water-extractable polysaccharide conjugates from northern Manitoba lingonberry. Cogent Food Agric. 2015, 1. [Google Scholar] [CrossRef]
- Quek, R.; Henry, C.J. Influence of polyphenols from lingonberry, cranberry, and red grape onin vitrodigestibility of rice. Int. J. Food Sci. Nutr. 2015, 66, 378–382. [Google Scholar] [CrossRef]
- Beaulieu, L.-P.; Harris, C.S.; Saleem, A.; Cuerrier, A.; Haddad, P.S.; Martineau, L.C.; Bennett, S.A.; Arnason, J.T. Inhibitory effect of the cree traditional medicine wiishichimanaanh (Vaccinium vitis-idaea) on advanced glycation endproduct formation: Identification of active principles. Phytotherapy Res. 2009, 24, 741–747. [Google Scholar] [CrossRef]
- Kylli, P.; Nohynek, L.; Puupponen-Pimia, R.; Westerlund-Wikström, B.; Leppanen, T.; Welling, J.; Moilanen, E.; Heinonen, M. Lingonberry (Vaccinium vitis-idaea) and European cranberry (Vaccinium microcarpon) proanthocyanidins: Isolation, identification, and bioactivities. J. Agric. Food Chem. 2011, 59, 3373–3384. [Google Scholar] [CrossRef]
- Kowalska, K.; Olejnik, A.; Zielińska-Wasielica, J.; Olkowicz, M. Inhibitory effects of lingonberry (Vaccinium vitis-idaea L.) fruit extract on obesity-induced inflammation in 3T3-L1 adipocytes and RAW 264.7 macrophages. J. Funct. Foods 2019, 54, 371–380. [Google Scholar] [CrossRef]
- Riihinen, K.; Kamal-Eldin, A.; Mattila, P.; González-Paramás, A.M.; Törrönen, A.R. Distribution and contents of phenolic compounds in eighteen Scandinavian berry species. J. Agric. Food Chem. 2004, 52, 4477–4486. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, M.A.; Hoffmann, C.; Sherrill–Mix, S.A.; Keilbaugh, S.A.; Hamady, M.; Chen, Y.; Knight, R.; Ahima, R.S.; Bushman, F.D.; Wu, G.D.; et al. High-fat diet determines the composition of the murine gut microbiome independently of obesity. Gastroenterology 2009, 137, 1716–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krych, L.; Nielsen, D.; Hansen, A.; Hansen, C. Gut microbial markers are associated with diabetes onset, regulatory imbalance, and IFN-γ level in NOD Mice. Gut Microbes 2015, 6, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Kameyama, K.; Itoh, K. Intestinal Colonization by a Lachnospiraceae Bacterium Contributes to the Development of Diabetes in Obese Mice. Microbes Environ. 2014, 29, 427–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, D.; Orwat, J.; Hurburt, T.; Woods, F.; Pitts, J.A.; Shetty, K. Evaluation of phenolic bioactive-linked functionality of blackberry cultivars targeting dietary management of early stages type-2 diabetes using in vitro models. Sci. Hortic. 2016, 212, 193–202. [Google Scholar] [CrossRef]
- Sancho, R.A.S.; Pastore, G.M. Evaluation of the effects of anthocyanins in type 2 diabetes. Food Res. Int. 2012, 46, 378–386. [Google Scholar] [CrossRef]
- Boath, A.S.; Grussu, D.; Stewart, D.; McDougall, G.J. Berry polyphenols inhibit digestive enzymes: A source of potential health benefits? Food Dig. 2012, 3, 1–7. [Google Scholar] [CrossRef]
- Gowd, V.; Bao, T.; Chen, W. Antioxidant potential and phenolic profile of blackberry anthocyanin extract followed by human gut microbiota fermentation. Food Res. Int. 2018, 120, 523–533. [Google Scholar] [CrossRef]
- Solverson, P.; Rumpler, W.V.; Leger, J.L.; Redan, B.W.; Ferruzzi, M.; Baer, D.J.; Castonguay, T.W.A.; Novotny, J. Blackberry feeding increases fat oxidation and improves insulin sensitivity in overweight and obese males. Nutrients 2018, 10, 1048. [Google Scholar] [CrossRef] [Green Version]
- Williamson, G. Possible effects of dietary polyphenols on sugar absorption and digestion. Mol. Nutr. Food Res. 2012, 57, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Giampieri, F.; Forbes-Hernández, T.Y.; Gasparrini, M.; Alvarez-Suarez, J.M.; Afrin, S.; Bompadre, S.; Quiles, J.L.; Mezzetti, B.; Battino, M. Strawberry as a health promoter: An evidence based review. Food Funct. 2015, 6, 1386–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q. In vitro bioactivities and phytochemical profile of various parts of the strawberry (Fragaria × ananassa var. Amaou). J. Funct. Foods 2015, 13, 38–49. [Google Scholar] [CrossRef]
- Manzano, S.; Williamson, G. Polyphenols and phenolic acids from strawberry and apple decrease glucose uptake and transport by human intestinal Caco-2 cells. Mol. Nutr. Food Res. 2010, 54, 1773–1780. [Google Scholar] [CrossRef]
- da Silva Pinto, M. Evaluation of antiproliferative, anti-Type 2 Diabetes, and antihypertension potentials of ellagitannins from strawberries (Fragaria × ananassa Duch.) using in vitro models. J. Med. Food 2010, 13, 1027–1035. [Google Scholar] [CrossRef]
- Forbes-Hernández, T.Y.; Giampieri, F.; Gasparrini, M.; Afrin, S.; Mazzoni, L.; Cordero, M.D.; Mezzetti, B.; Quiles, J.L.; Battino, M. Lipid accumulation in HepG2 cells is attenuated by strawberry extract through AMPK activation. Nutrients 2017, 9, 621. [Google Scholar] [CrossRef] [Green Version]
- Basu, A.; Fu, D.X.; Wilkinson, M.; Simmons, B.; Wu, M.; Betts, N.M.; Du, M.; Lyons, T.J. Strawberries decrease atherosclerotic markers in subjects with metabolic syndrome. Nutr. Res. 2010, 30, 462–469. [Google Scholar] [CrossRef] [Green Version]
- Pajk, T.; Rezar, V.; Levart, A.; Salobir, J. Efficiency of apples, strawberries, and tomatoes for reduction of oxidative stress in pigs as a model for humans. Nutrients 2006, 22, 376–384. [Google Scholar] [CrossRef]
- Kulczyński, B.; Gramza-Michałowska, A. Goji berry (Lycium barbarum): Composition and health effects—A review. Pol. J. Food Nutr. Sci. 2016, 66, 67–75. [Google Scholar] [CrossRef]
- Jin, M.; Huang, Q.; Zhao, K.; Shang, P. Biological activities and potential health benefit effects of polysaccharides isolated from Lycium barbarum L. Int. J. Biol. Macromol. 2013, 54, 16–23. [Google Scholar] [CrossRef]
- Zou, P.; Yang, X.; Huang, W.W.; Zhao, H.; Wang, J.; Xu, R.B.; Hu, X.L.; Shen, S.Y.; Qin, D. Characterization and bioactivity of polysaccharides obtained from pine cones of pinus koraiensis by graded ethanol precipitation. Molecules 2013, 18, 9933–9948. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.L.; Chen, C.; Wang, S.K.; Sun, G.J. Biochemical analysis and hypoglycemic activity of a polysaccharide isolated from the fruit of Lycium barbarum L. Int. J. Biol. Macromol. 2015, 77, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Shen, T.; Jiang, X.; Xia, M.; Sun, X.; Guo, H.; Ling, W. Purified anthocyanins from bilberry and black currant attenuate hepatic mitochondrial dysfunction and steatohepatitis in mice with methionine and choline deficiency. J. Agric. Food Chem. 2015, 63, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Yan, Y.; Peng, Y.; Chen, D.; Mi, J.; Lu, L.; Luo, Q.; Li, X.; Zeng, X.; Cao, Y. In vitro digestion under simulated saliva, gastric and small intestinal conditions and fermentation by human gut microbiota of polysaccharides from the fruits of Lycium barbarum. Int. J. Biol. Macromol. 2019, 125, 751–760. [Google Scholar] [CrossRef]
- Girones-Vilaplana, A.; Villaño, D.; Moreno, D.A.; García-Viguera, C. New isotonic drinks with antioxidant and biological capacities from berries (maqui, açaí and blackthorn) and lemon juice. Int. J. Food Sci. Nutr. 2013, 64, 897–906. [Google Scholar] [CrossRef]
- Cheguru, P.; Chapalamadugu, K.C.; Doumit, M.E.; Murdoch, G.K.; Hill, R.A. Adipocyte differentiation-specific gene transcriptional response to C18 unsaturated fatty acids plus insulin. Pflugers Arch. 2012, 463, 429–447. [Google Scholar] [CrossRef]
- Costa, A.G.V.; Garcia-Diaz, D.F.; Jimenez, P.; Silva, P.I. Bioactive compounds and health benefits of exotic tropical red–black berries. J. Funct. Foods 2013, 5, 539–549. [Google Scholar] [CrossRef]
- Karpac, J.; Jasper, H. Insulin and JNK: Optimizing metabolic homeostasis and lifespan. Trends Endocrinol. Metab. 2009, 20, 100–106. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.C.; Mullen, K.L.; Junkin, K.A.; Nickerson, J.; Chabowski, A.; Bonen, A.; Dyck, D.J. Metformin and exercise reduce muscle FAT/CD36 and lipid accumulation and blunt the progression of high-fat diet-induced hyperglycemia. Am. J. Physiol. Metab. 2007, 293, E172–E181. [Google Scholar] [CrossRef] [Green Version]
- Umpierrez, G.E.; Bailey, T.S.; Carcia, D.; Shaefer, C.; Shubrook, J.H.; Skolnik, N. Improving postprandial hyperglycemia in patients with type 2 diabetes already on basal insulin therapy: Review of current strategies. J. Diabetes 2017, 10, 94–111. [Google Scholar] [CrossRef]
- Oyama, L.M.; Silva, F.; Carnier, J.; De Miranda, D.A.; Santamarina, A.B.; Ribeiro, E.B.; Nascimento, C.M.O.D.; De Rosso, V.V. Juçara pulp supplementation improves glucose tolerance in mice. Diabetol. Metab. Syndr. 2016, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, A.K.; Cadoná, F.C.; Assmann, C.E.; Andreazza, A.C.; Duarte, M.M.M.F.; Branco, C.D.S.; Zhou, X.; De Souza, D.V.; Ribeiro, E.E.; Da Cruz, I.B.M. Açaí (Euterpe oleracea Mart.) has anti-inflammatory potential through NLRP3-inflammasome modulation. J. Funct. Foods 2019, 56, 364–371. [Google Scholar] [CrossRef]
- Martino, H.S.D.; Dias, M.M.D.S.; Noratto, G.; Talcott, S.; Mertens-Talcott, S.U. Anti-lipidaemic and anti-inflammatory effect of açai (Euterpe oleracea Martius) polyphenols on 3T3-L1 adipocytes. J. Funct. Foods 2016, 23, 432–443. [Google Scholar] [CrossRef]
- Du, X.; Myracle, A.D. Fermentation alters the bioaccessible phenolic compounds and increases the alpha-glucosidase inhibitory effects of aronia juice in a dairy matrix followingin vitrodigestion. Food Funct. 2018, 9, 2998–3007. [Google Scholar] [CrossRef]
- Rugina, D.; Diaconeasa, Z.M.; Isvoranu, C.; Bunea, A.; Socaciu, C.; Pintea, A. Chokeberry Anthocyanin Extract as Pancreaticβ-Cell Protectors in Two Models of Induced Oxidative Stress. Oxidative Med. Cell. Longev. 2015, 2015, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Iwashima, T.; Kudome, Y.; Kishimoto, Y.; Saita, E.; Tanaka, M.; Taguchi, C.; Hirakawa, S.; Mitani, N.; Kondo, K.; Iida, K. Aronia berry extract inhibits TNF-α-induced vascular endothelial inflammation through the regulation of STAT3. Food Nutr. Res. 2019, 63, 3361. [Google Scholar] [CrossRef]
- Zapolska-Downar, D.; Bryk, D.; Małecki, M.; Hajdukiewicz, K.; Sitkiewicz, D. Aronia melanocarpa fruit extract exhibits anti-inflammatory activity in human aortic endothelial cells. Eur. J. Nutr. 2011, 51, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Törrönen, R.; Sarkkinen, E.; Tapola, N.; Hautaniemi, E.; Kilpi, K.; Niskanen, L. Berries modify the postprandial plasma glucose response to sucrose in healthy subjects. Br. J. Nutr. 2009, 103, 1094–1097. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Niu, X.; Liu, N.; Gao, Y.; Wang, L.; Xu, G.; Li, X.; Yang, Y. Characterization, antioxidant and hypoglycemic activities of degraded polysaccharides from blackcurrant (Ribes nigrum L.) fruits. Food Chem. 2017, 243, 26–35. [Google Scholar] [CrossRef]
- Hossain, A.K.M.M.; Mason, S.L.; Guo, X.-B.; Brennan, C.R. The combined effect of blackcurrant powder and wholemeal flours to improve health promoting properties of cookies. Plant. Foods Hum. Nutr. 2017, 72, 280–287. [Google Scholar] [CrossRef]
- Cespedes, C.L.; Valdez-Morales, M.; Avila, J.G.; El-Hafidi, M.; Alarcon, J.; Paredes-López, O. Phytochemical profile and the antioxidant activity of Chilean wild black-berry fruits, Aristotelia chilensis (Mol) Stuntz (Elaeocarpaceae). Food Chem. 2010, 119, 886–895. [Google Scholar] [CrossRef]
- Brauch, J.E.; Reuter, L.; Conrad, J.; Vogel, H.; Schweiggert, R.; Carle, R. Characterization of anthocyanins in novel Chilean maqui berry clones by HPLC–DAD–ESI/MSn and NMR-spectroscopy. J. Food Compos. Anal. 2017, 58, 16–22. [Google Scholar] [CrossRef]
- Rubilar, M.; Jara, C.; Poo, Y.; Acevedo, F.; Gutiérrez, C.; Sineiro, J.; Shene, C. Extracts of Maqui (Aristotelia chilensis) and Murta (Ugni molinae Turcz.): Sources of antioxidant compounds and α-glucosidase/α-amylase inhibitors. J. Agric. Food Chem. 2011, 59, 1630–1637. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Farias, M.; Vasquez, K.; Fuentes, F.; Ovalle-Marin, A.; Parra-Ruiz, C.; Zamora, O.; Pino, M.; Quitral, V.; Jimenez, P.; Garcia, L.; et al. Extracts of Chilean native fruits inhibit oxidative stress, inflammation and insulin-resistance linked to the pathogenic interaction between adipocytes and macrophages. J. Funct. Foods 2016, 27, 69–83. [Google Scholar] [CrossRef]
- Schreckinger, M.E.; Wang, J.; Yousef, G.; Lila, M.A.; De Mejia, E.G. Antioxidant capacity and in vitro inhibition of adipogenesis and inflammation by phenolic extracts of vaccinium floribundum and Aristotelia chilensis. J. Agric. Food Chem. 2010, 58, 8966–8976. [Google Scholar] [CrossRef]
- Hidalgo, J. Delphinidin reduces glucose uptake in mice jejunal tissue and human intestinal cells lines through FFA1/GPR40. Int. J. Mol. Sci. 2017, 18, 750. [Google Scholar] [CrossRef] [Green Version]
- Christensen, M.; Calanna, S.; Sparre-Ulrich, A.H.; Kristensen, P.L.; Rosenkilde, M.M.; Faber, J.; Purrello, F.; Van Hall, G.; Holst, J.J.; Vilsbøll, T.; et al. Glucose-dependent insulinotropic polypeptide augments glucagon responses to hypoglycemia in Type 1 Diabetes. Diabetes 2014, 64, 72–78. [Google Scholar] [CrossRef] [Green Version]
- Miranda-Rottmann, S.; Aspillaga, A.A.; Pérez, D.D.; Vásquez, L.; Martinez, A.L.F.; Leighton, F. Juice and Phenolic Fractions of the BerryAristotelia chilensisInhibit LDL Oxidationin vitroand protect human endothelial cells against oxidative stress. J. Agric. Food Chem. 2002, 50, 7542–7547. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hameed, A.; Galli, M.; Adamska-Patruno, E.; Krętowski, A.; Ciborowski, M. Select Polyphenol-Rich Berry Consumption to Defer or Deter Diabetes and Diabetes-Related Complications. Nutrients 2020, 12, 2538. https://doi.org/10.3390/nu12092538
Hameed A, Galli M, Adamska-Patruno E, Krętowski A, Ciborowski M. Select Polyphenol-Rich Berry Consumption to Defer or Deter Diabetes and Diabetes-Related Complications. Nutrients. 2020; 12(9):2538. https://doi.org/10.3390/nu12092538
Chicago/Turabian StyleHameed, Ahsan, Mauro Galli, Edyta Adamska-Patruno, Adam Krętowski, and Michal Ciborowski. 2020. "Select Polyphenol-Rich Berry Consumption to Defer or Deter Diabetes and Diabetes-Related Complications" Nutrients 12, no. 9: 2538. https://doi.org/10.3390/nu12092538
APA StyleHameed, A., Galli, M., Adamska-Patruno, E., Krętowski, A., & Ciborowski, M. (2020). Select Polyphenol-Rich Berry Consumption to Defer or Deter Diabetes and Diabetes-Related Complications. Nutrients, 12(9), 2538. https://doi.org/10.3390/nu12092538