A High Phosphorus Diet Impairs Testicular Function and Spermatogenesis in Male Mice with Chronic Kidney Disease

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Model and Experimental Design
2.2. Biochemical Markers in the Serum
2.3. Semen Quality Analysis
2.4. Testicular Histological Analysis
2.5. Testicular Hormone Levels Analysis
2.6. Western Blot Analysis
2.7. Statistical Analysis
3. Results
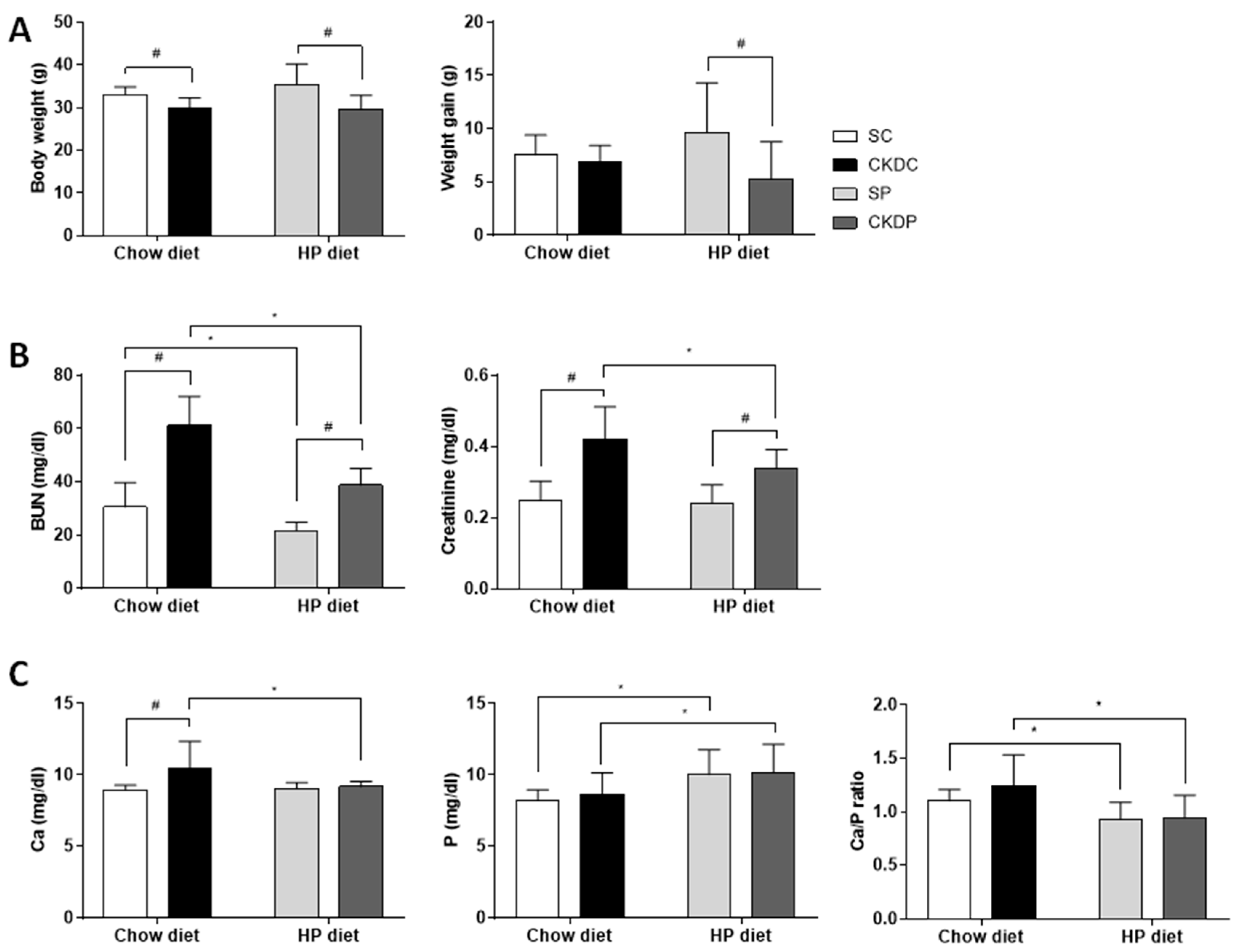
3.1. Physiological and Serum Biochemical Parameters
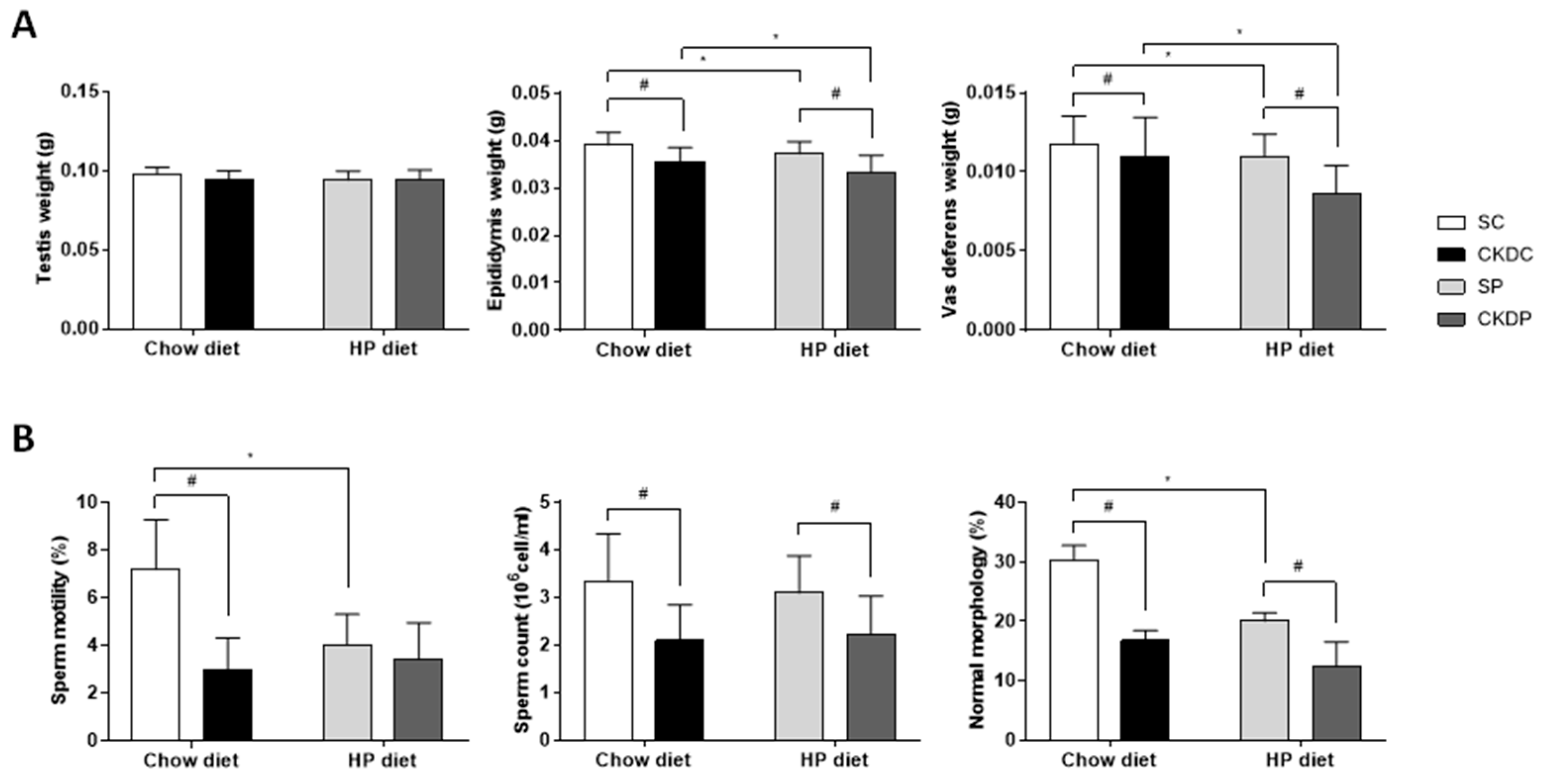
3.2. Reproductive Organ Weights and Semen Quality
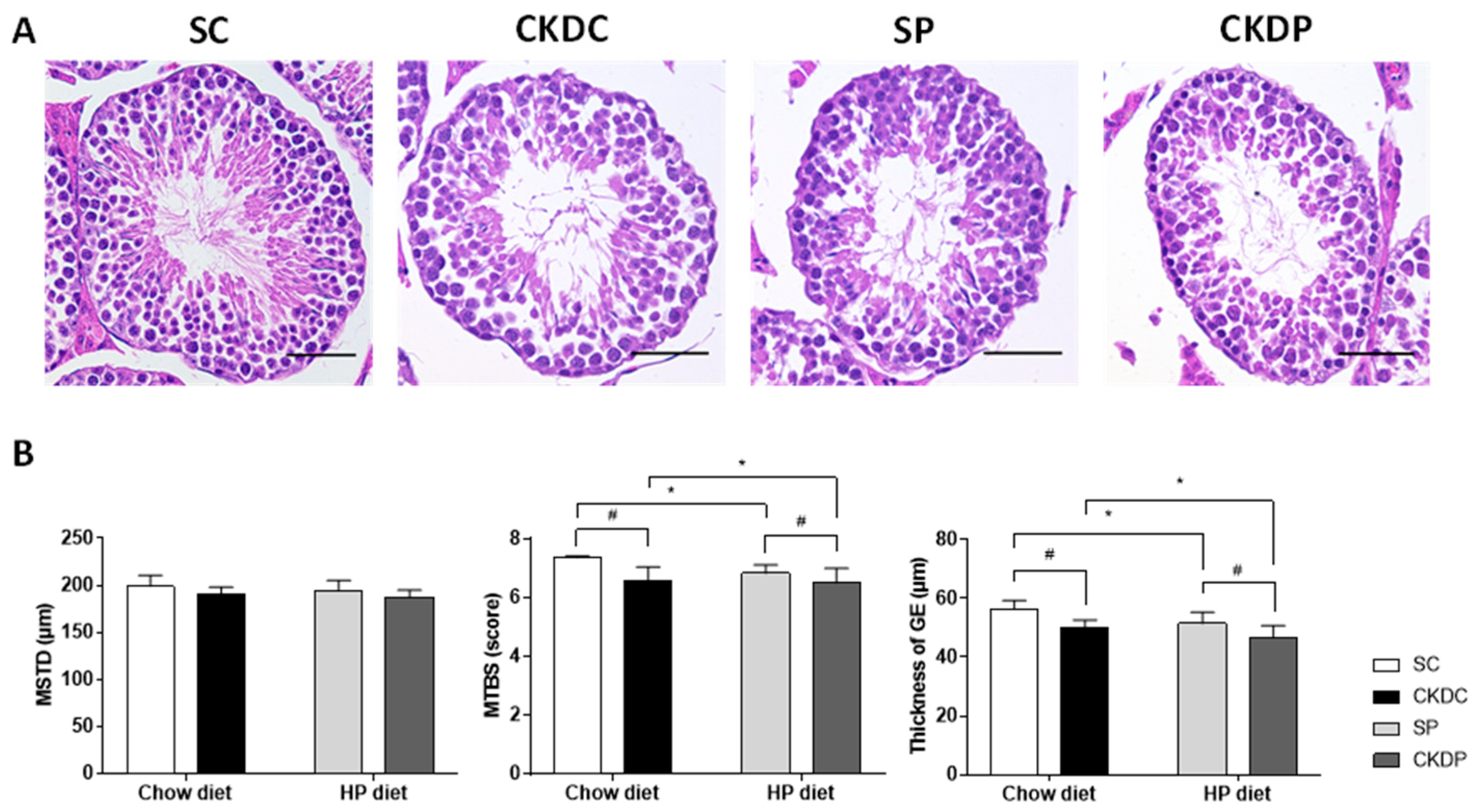
3.3. Testicular Morphology and Related Parameters
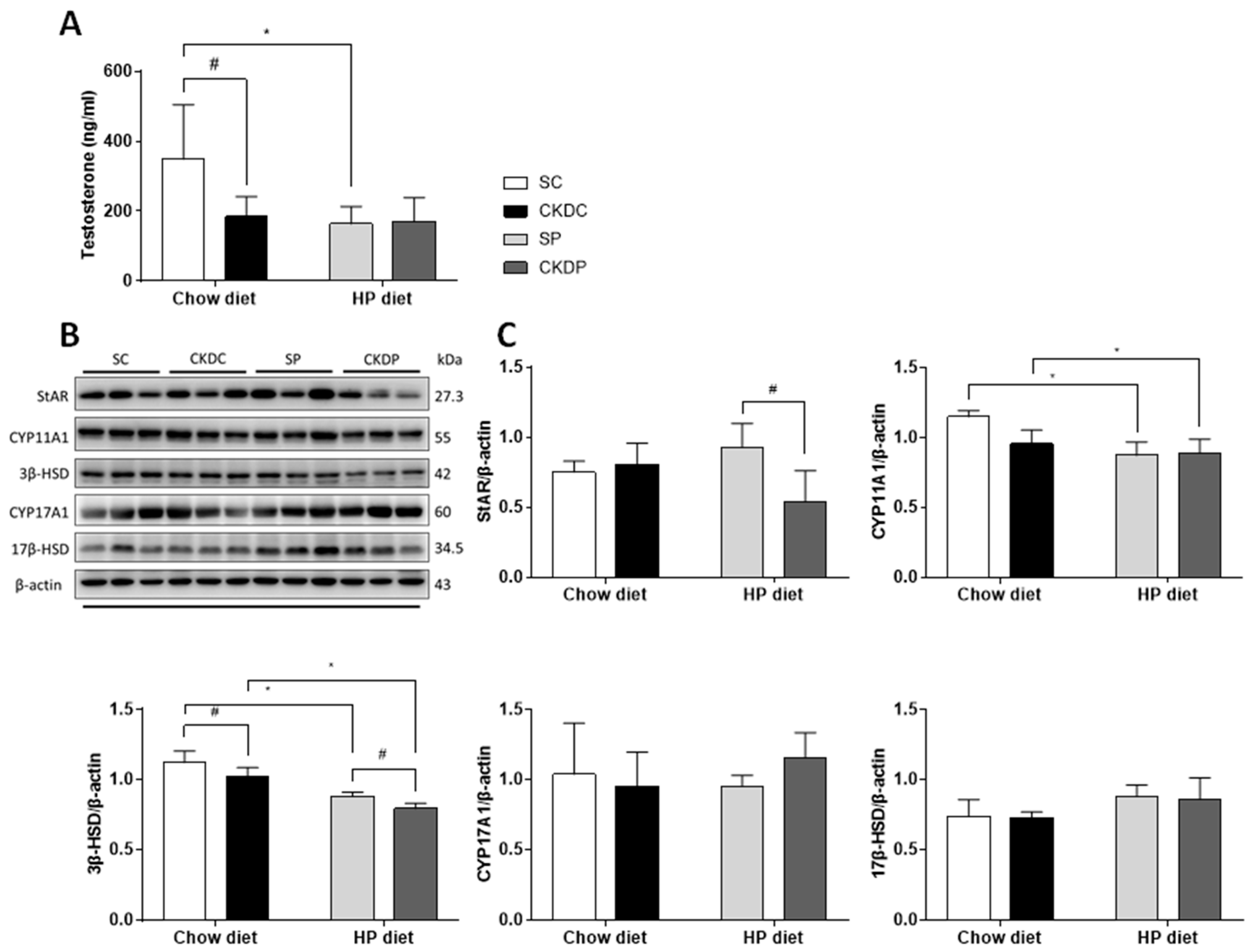
3.4. Testicular Testosterone Level and Protein Expressions of Testosterone Biosynthesis Enzymes in the Testis
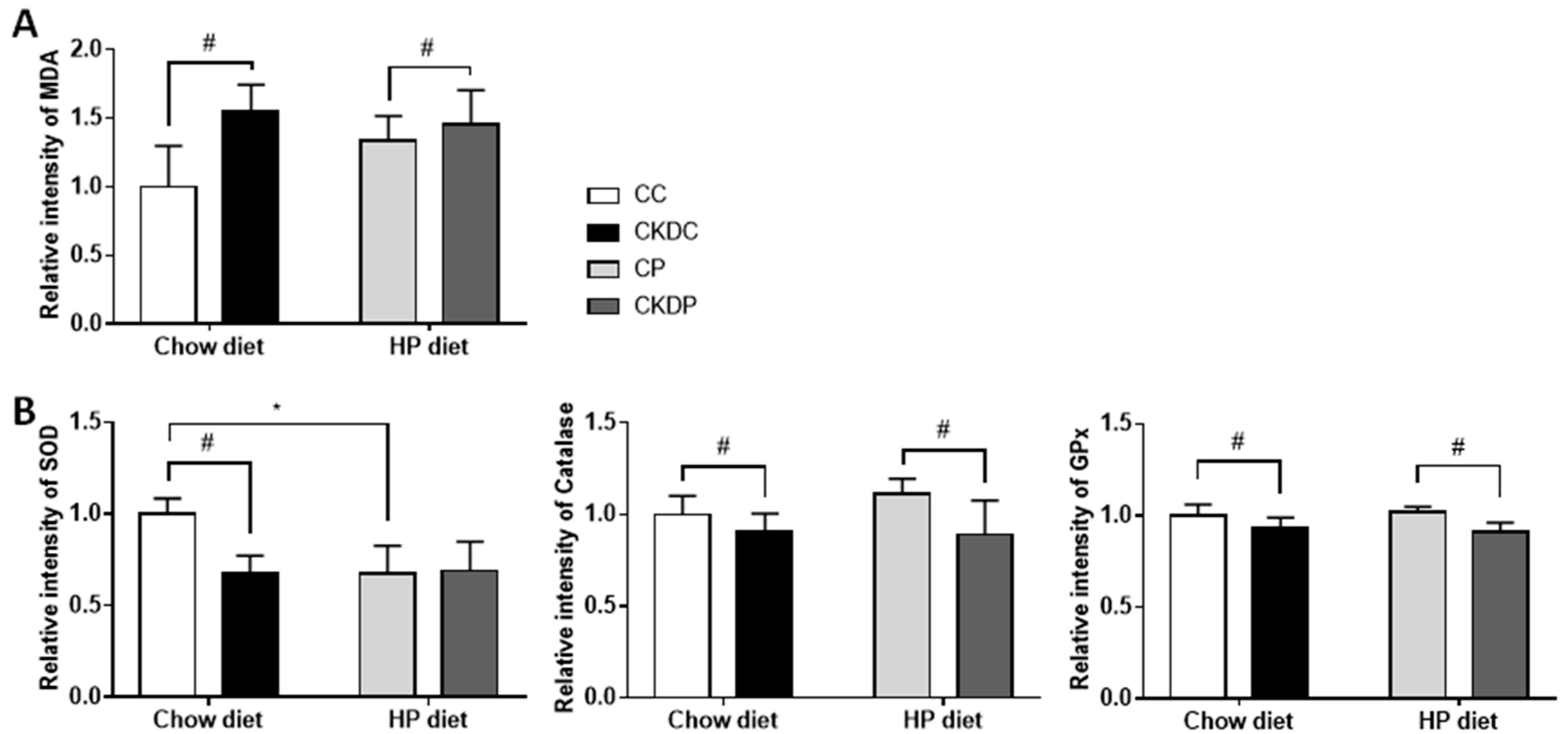
3.5. Oxidative Stress Biomarkers in Testis
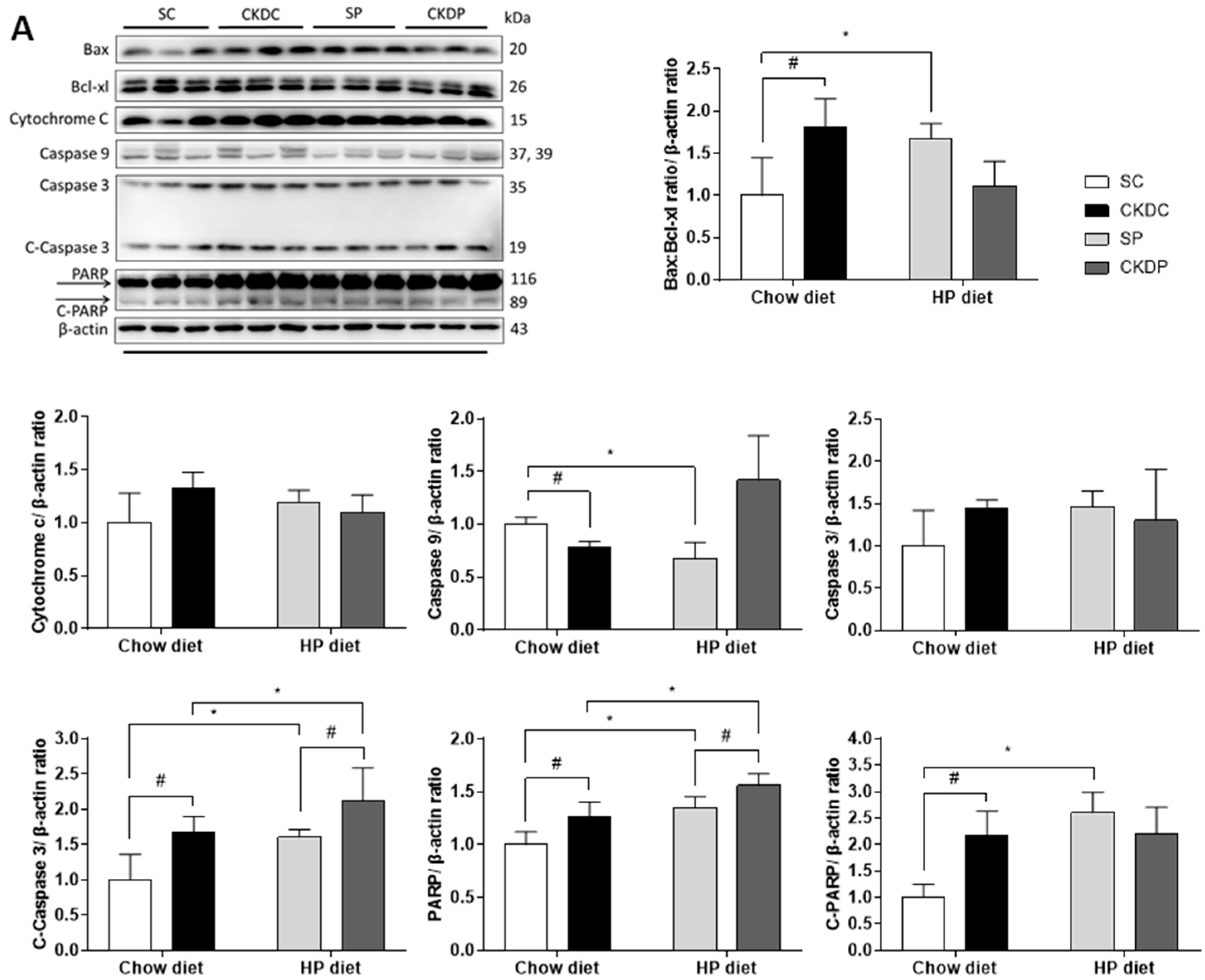
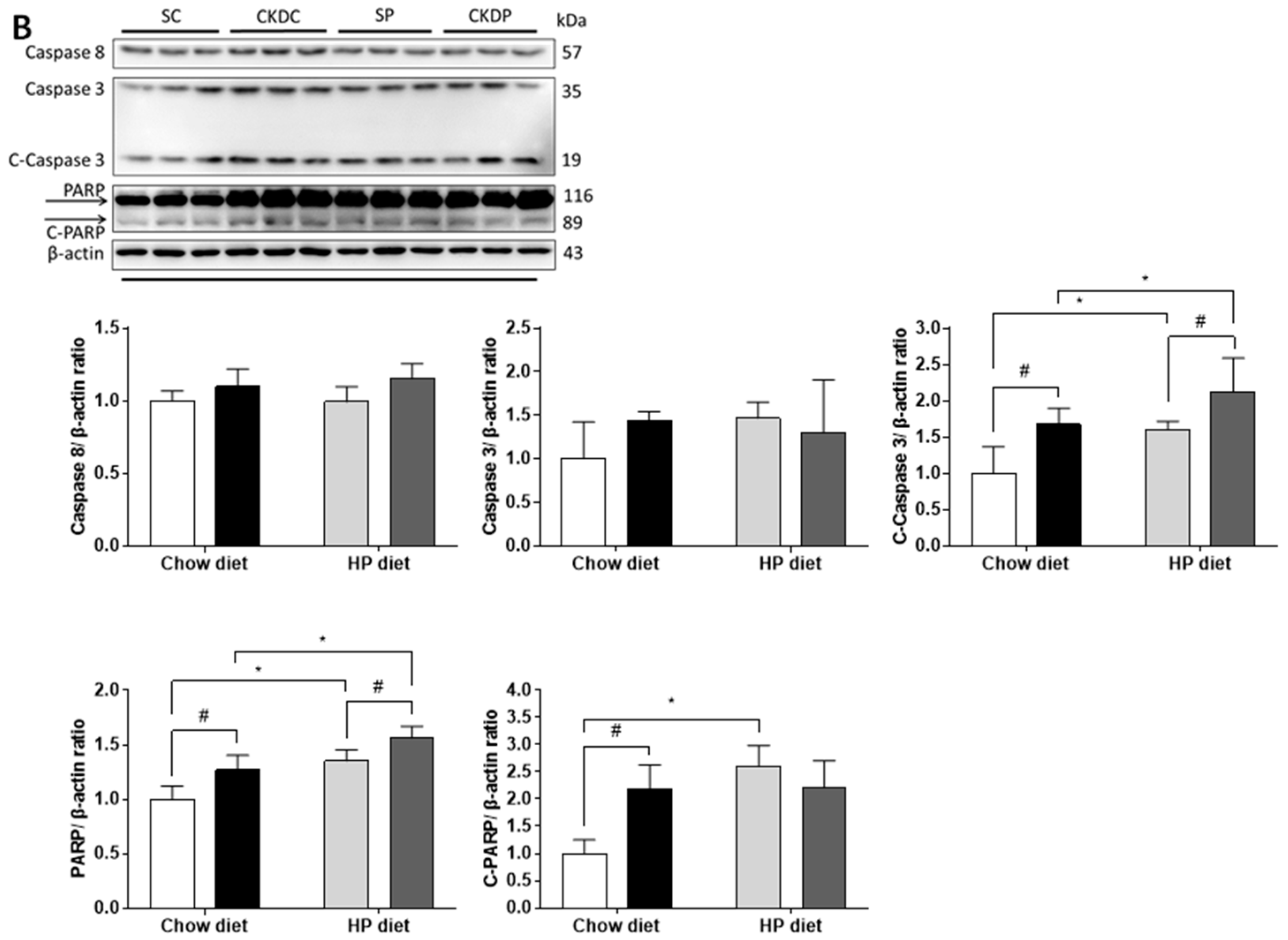
3.6. Protein Expressions of Apoptosis Markers in the Testis
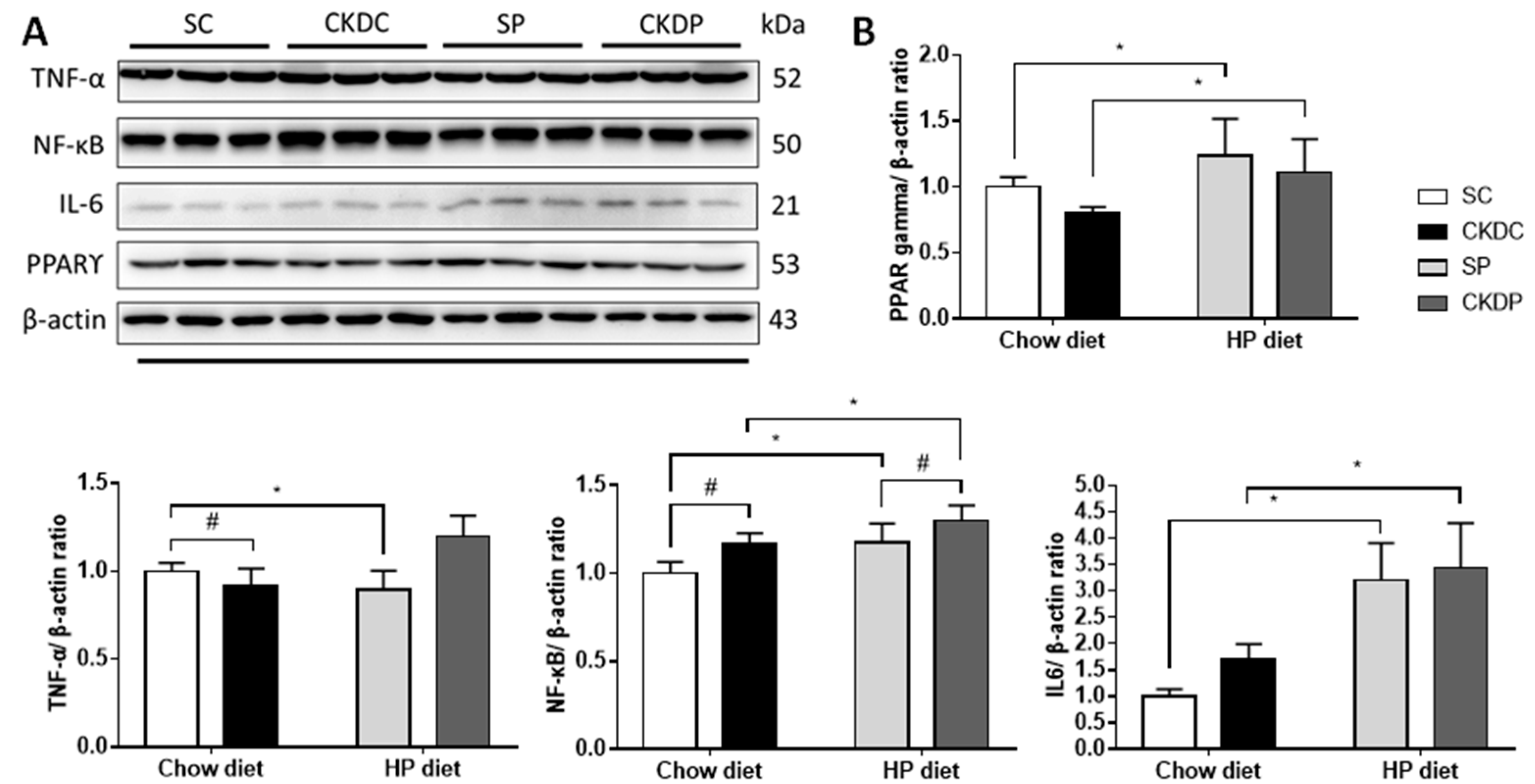
3.7. Protein Expressions of Inflammation Markers in the Testis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tsai, M.H.; Hsu, C.Y.; Lin, M.Y.; Yen, M.F.; Chen, H.H.; Chiu, Y.H.; Hwang, S.J. Incidence, prevalence, and duration of chronic kidney disease in Taiwan: Results from a community-based screening program of 106,094 individuals. Nephron 2018, 140, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Hill, N.R.; Fatoba, S.T.; Oke, J.L.; Hirst, J.A.; O’Callaghan, C.A.; Lasserson, D.S.; Hobbs, F.D.R. Global prevalence of chronic kidney disease—A systematic review and meta-analysis. PLoS ONE 2016, 11, e0158765. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.-Y.; Tseng, H.-F.; Tan, H.-F.; Chien, Y.-S.; Chang, C.-C. End-stage renal disease in Taiwan: A case-control study. J. Epidemiol. 2009, 19, 169–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, A.J.; Foley, R.N.; Gilbertson, D.T.; Chen, S.-C. United States Renal Data System public health surveillance of chronic kidney disease and end-stage renal disease. Kidney Int. Suppl. 2015, 5, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Goretti Penido, M.; Alon, U.S. Phosphate homeostasis and its role in bone health. Pediatric Nephrol. 2012, 27, 2039–2048. [Google Scholar] [CrossRef] [Green Version]
- Shaikh, A.; Berndt, T.; Kumar, R. Regulation of phosphate homeostasis by the phosphatonins and other novel mediators. Pediatric Nephrol. 2008, 23, 1203–1210. [Google Scholar] [CrossRef] [Green Version]
- Prasad, N.; Bhadauria, D. Renal phosphate handling: Physiology. Indian J. Endocrinol. Metab. 2013, 17, 620–627. [Google Scholar] [CrossRef]
- Craver, L.; Marco, M.P.; Martinez, I.; Rue, M.; Borras, M.; Martin, M.L.; Sarro, F.; Valdivielso, J.M.; Fernandez, E. Mineral metabolism parameters throughout chronic kidney disease stages 1–5—Achievement of K/DOQI target ranges. Nephrol. Dial. Transplant. 2007, 22, 1171–1176. [Google Scholar] [CrossRef] [Green Version]
- Qunibi, W.Y. Consequences of hyperphosphatemia in patients with end-stage renal disease (ESRD). Kidney Int. 2004, 66, S8–S12. [Google Scholar] [CrossRef] [Green Version]
- Friedman, E.A. Consequences and management of hyperphosphatemia in patients with renal insufficiency. Kidney Int. 2005, 67, S1–S7. [Google Scholar] [CrossRef] [Green Version]
- Shaman, A.M.; Kowalski, S.R. Hyperphosphatemia management in patients with chronic kidney disease. Saudi Pharm. J. 2016, 24, 494–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, S.K.; Choi, K.; Kim, S.K.; Lee, G.I.; Cho, I.-C. Phosphorus as predictive factor for erectile dysfunction in middle aged men: A cross sectional study in Korea. Investig. Clin. Urol. 2016, 57, 442–448. [Google Scholar] [CrossRef]
- Lehtihet, M.; Hylander, B. Semen quality in men with chronic kidney disease and its correlation with chronic kidney disease stages. Andrologia 2015, 47, 1103–1108. [Google Scholar] [CrossRef]
- Edey, M.M. Male sexual dysfunction and chronic kidney disease. Front. Med. 2017, 4, 32. [Google Scholar] [CrossRef] [Green Version]
- Banjoko, S.O.; Adeseolu, F.O. Seminal plasma pH, inorganic phosphate, total and ionized calcium concentrations in the assessment of human spermatozoa function. J. Clin. Diagn. Res. 2013, 7, 2483–2486. [Google Scholar] [CrossRef]
- Suzuki, Y.; Ichihara, G.; Sahabudeen, S.M.; Kato, A.; Yamaguchi, T.; Imanaka-Yoshida, K.; Yoshida, T.; Yamada, Y.; Ichihara, S. Rats with metabolic syndrome resist the protective effects of N-acetyl l-cystein against impaired spermatogenesis induced by high-phosphorus/zinc-free diet. Exp. Toxicol. Pathol. 2013, 65, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, S.; Pergola, P.E.; Zager, R.A.; Vaziri, N.D. Targeting the transcription factor Nrf2 to ameliorate oxidative stress and inflammation in chronic kidney disease. Kidney Int. 2013, 83, 1029–1041. [Google Scholar] [CrossRef] [Green Version]
- Mihai, S.; Codrici, E.; Popescu, I.D.; Enciu, A.-M.; Albulescu, L.; Necula, L.G.; Mambet, C.; Anton, G.; Tanase, C. Inflammation-related mechanisms in chronic kidney disease prediction, progression, and outcome. J. Immunol. Res. 2018, 2018, 16. [Google Scholar] [CrossRef] [PubMed]
- Daenen, K.; Andries, A.; Mekahli, D.; Van Schepdael, A.; Jouret, F.; Bammens, B. Oxidative stress in chronic kidney disease. Pediatric Nephrol. 2019, 34, 975–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, N.; Elsayed, E. Dietary phosphate: What do we know about its toxicity? J. Nephrol. 2013, 26, 856–864. [Google Scholar] [CrossRef]
- Yamada, S.; Tokumoto, M.; Tatsumoto, N.; Taniguchi, M.; Noguchi, H.; Nakano, T.; Masutani, K.; Ooboshi, H.; Tsuruya, K.; Kitazono, T. Phosphate overload directly induces systemic inflammation and malnutrition as well as vascular calcification in uremia. Am. J. Physiol. Renal Physiol. 2014, 306, F1418–F1428. [Google Scholar] [CrossRef] [PubMed]
- Bain, J. The many faces of testosterone. Clin. Interv. Aging 2007, 2, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, V.; Scordo, M.; Yoon, R.S.; Liporace, F.A.; Greene, L.W. Revisiting the role of testosterone: Are we missing something? Rev. Urol. 2017, 19, 16–24. [Google Scholar] [CrossRef]
- Tripathi, U.K.; Chhillar, S.; Kumaresan, A.; Aslam, M.K.M.; Rajak, S.K.; Nayak, S.; Manimaran, A.; Mohanty, T.K.; Yadav, S. Morphometric evaluation of seminiferous tubule and proportionate numerical analysis of Sertoli and spermatogenic cells indicate differences between crossbred and purebred bulls. Vet. World 2015, 8, 645–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnsen, S.G. Testicular biopsy score count—A method for registration of spermatogenesis in human testes: Normal values and results in 335 hypogonadal males. Horm. Res. Paediatr. 1970, 1, 2–25. [Google Scholar] [CrossRef] [PubMed]
- Kujal, P.; Vernerová, Z. 5/6 nephrectomy as an experimental model of chronic renal failure and adaptation to reduced nephron number. Ceskoslovenská Fysiol. 2008, 57, 104–109. [Google Scholar]
- Hamzaoui, M.; Djerada, Z.; Brunel, V.; Mulder, P.; Richard, V.; Bellien, J.; Guerrot, D. 5/6 nephrectomy induces different renal, cardiac and vascular consequences in 129/Sv and C57BL/6JRj mice. Sci. Rep. 2020, 10, 1524. [Google Scholar] [CrossRef] [Green Version]
- Souza, A.C.P.; Tsuji, T.; Baranova, I.N.; Bocharov, A.V.; Wilkins, K.J.; Street, J.M.; Alvarez-Prats, A.; Hu, X.; Eggerman, T.; Yuen, P.S.T.; et al. TLR4 mutant mice are protected from renal fibrosis and chronic kidney disease progression. Physiol. Rep. 2015, 3, e12558. [Google Scholar] [CrossRef]
- Gava, A.; Freitas, F.; Balarini, C.; Vasquez, E.; Meyrelles, S. Effects of 5/6 nephrectomy on renal function and blood pressure in mice. Int. J. Physiol. Pathophysiol. Pharmacol. 2012, 4, 167–173. [Google Scholar]
- Tan, R.-Z.; Zhong, X.; Li, J.-C.; Zhang, Y.-W.; Yan, Y.; Liao, Y.; Wen, D.; Diao, H.; Wang, L.; Shen, H.-C. An optimized 5/6 nephrectomy mouse model based on unilateral kidney ligation and its application in renal fibrosis research. Ren. Fail. 2019, 41, 555–566. [Google Scholar] [CrossRef]
- Shimada, M.; Shutto-Uchita, Y.; Yamabe, H. Lack of awareness of dietary sources of phosphorus is a clinical concern. In Vivo 2019, 33, 11–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Abbadi, M.M.; Pai, A.S.; Leaf, E.M.; Yang, H.-Y.; Bartley, B.A.; Quan, K.K.; Ingalls, C.M.; Liao, H.W.; Giachelli, C.M. Phosphate feeding induces arterial medial calcification in uremic mice: Role of serum phosphorus, fibroblast growth factor-23, and osteopontin. Kidney Int. 2009, 75, 1297–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, W.L.; Linnes, M.; Chu, E.Y.; Foster, B.L.; Bartley, B.A.; Somerman, M.J.; Giachelli, C.M. High phosphate feeding promotes mineral and bone abnormalities in mice with chronic kidney disease. Nephrol. Dial. Transplant. 2013, 28, 62–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, W.L.; Leaf, E.M.; Hu, M.C.; Takeno, M.M.; Kuro-o, M.; Moe, O.W.; Giachelli, C.M. Vitamin D receptor agonists increase klotho and osteopontin while decreasing aortic calcification in mice with chronic kidney disease fed a high phosphate diet. Kidney Int. 2012, 82, 1261–1270. [Google Scholar] [CrossRef] [Green Version]
- Mawhinney, M.; Mariotti, A. Physiology, pathology and pharmacology of the male reproductive system. Periodontology 2000 2013, 61, 232–251. [Google Scholar] [CrossRef]
- Li, G.; Xin, Z.; Yuan, Y.; Yang, X.; Xia, T.; Liu, W.; Fu, J.; Tian, L.; Na, Y. Seminiferous tubule scores used for quantitative assessment of spermatogenic function of patients with azoospermia. Natl. J. Androl. 2004, 10, 94–99. [Google Scholar]
- Rigi Manesh, M.; Arab, M.R.; Sargolzaei Aval, F.; Mashhadi, M.A.; Sargolzaei, N.; Mir, M.; Shahraki Salar, M. Cisplatin induce changes of cell surface glycoconjugates in germinal epithelium of seminiferous tubules. Anat. Sci. J. 2017, 14, 31–36. [Google Scholar]
- Slominski, A.T.; Li, W.; Kim, T.-K.; Semak, I.; Wang, J.; Zjawiony, J.K.; Tuckey, R.C. Novel activities of CYP11A1 and their potential physiological significance. J. Steroid Biochem. Mol. Biol. 2015, 151, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Xie, T.; Hu, G.; Dong, B.; Yan, Y.; Liu, M.; Yao, X.; Zheng, J.; Xu, Y. Roscovitine protects murine Leydig cells from lipopolysaccharide-induced inflammation. Exp. Ther. Med. 2017, 13, 2169–2176. [Google Scholar] [CrossRef]
- McVary, K.T. Clinical practice. Erectile dysfunction. N. Engl. J. Med. 2007, 357, 2472–2481. [Google Scholar] [CrossRef]
- Manna, P.R.; Stetson, C.L.; Slominski, A.T.; Pruitt, K. Role of the steroidogenic acute regulatory protein in health and disease. Endocrine 2016, 51, 7–21. [Google Scholar] [CrossRef] [Green Version]
- Razzaque, M.S. Phosphate toxicity: New insights into an old problem. Clin. Sci. 2011, 120, 91–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohnishi, M.; Razzaque, M.S. Dietary and genetic evidence for phosphate toxicity accelerating mammalian aging. FASEB J. 2010, 24, 3562–3571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative stress and male infertility. Nat. Rev. Urol. 2017, 14, 470–485. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Rana, M.; Qiu, E.; AlBunni, H.; Bui, A.D.; Henkel, R. Role of oxidative stress, infection and inflammation in male infertility. Andrologia 2018, 50, e13126. [Google Scholar] [CrossRef]
- Oberg, B.P.; McMenamin, E.; Lucas, F.L.; McMonagle, E.; Morrow, J.; Ikizler, T.A.; Himmelfarb, J. Increased prevalence of oxidant stress and inflammation in patients with moderate to severe chronic kidney disease. Kidney Int. 2004, 65, 1009–1016. [Google Scholar] [CrossRef] [Green Version]
- Granata, S.; Zaza, G.; Simone, S.; Villani, G.; Latorre, D.; Pontrelli, P.; Carella, M.; Schena, F.P.; Grandaliano, G.; Pertosa, G. Mitochondrial dysregulation and oxidative stress in patients with chronic kidney disease. BMC Genom. 2009, 10, 388. [Google Scholar] [CrossRef] [Green Version]
- Gyurászová, M.; Kovalčíková, A.G.; Renczés, E.; Kmeťová, K.; Celec, P.; Bábíčková, J.; Tóthová, Ľ. Oxidative stress in animal models of acute and chronic renal failure. Dis. Markers 2019, 2019, 8690805. [Google Scholar] [CrossRef] [Green Version]
- Ling, X.C.; Kuo, K.-L. Oxidative stress in chronic kidney disease. Ren. Replace. Ther. 2018, 4, 53. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Vaziri, N.D. Contribution of impaired Nrf2-Keap1 pathway to oxidative stress and inflammation in chronic renal failure. Am. J. Physiol. Renal Physiol. 2010, 298, F662–F671. [Google Scholar] [CrossRef] [Green Version]
- Ratliff, B.B.; Abdulmahdi, W.; Pawar, R.; Wolin, M.S. Oxidant mechanisms in renal injury and disease. Antioxid. Redox Signal. 2016, 25, 119–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, C.T.; Chiang, C.K. Uremic toxins, oxidative stress, and renal fibrosis: An interwined complex. J. Renal Nutr. 2015, 25, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Carrero, J.J.; Qureshi, A.R.; Nakashima, A.; Arver, S.; Parini, P.; Lindholm, B.; Bárány, P.; Heimbürger, O.; Stenvinkel, P. Prevalence and clinical implications of testosterone deficiency in men with end-stage renal disease. Nephrol. Dial. Transplant. 2011, 26, 184–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Print, C.G.; Loveland, K.L. Germ cell suicide: New insights into apoptosis during spermatogenesis. BioEssays 2000, 22, 423–430. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Moreno, J.M.; Herencia, C.; de Oca, A.M.; Díaz-Tocados, J.M.; Vergara, N.; Gómez-Luna, M.J.; López-Argüello, S.D.; Camargo, A.; Peralbo-Santaella, E.; Rodríguez-Ortiz, M.E.; et al. High phosphate induces a pro-inflammatory response by vascular smooth muscle cells and modulation by vitamin D derivatives. Clin. Sci. 2017, 131, 1449–1463. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient (%) | Chow | HP |
|---|---|---|
| Protein | 24.6 | 18.2 |
| Fat (ether extract) | 5.0 | 7.1 |
| Fat (acid hydrolysis) | 6.4 | 7.1 |
| Fiber | 4.2 | 5.0 |
| Nitrogen-free extract | 50.0 | 58.5 |
| Starch | 29.4 | 43.0 |
| Sucrose | 1.2 | 12.4 |
| Calcium | 1.0 | 0.5 |
| Phosphorus | 0.8 | 2.0 |
| Energy (%) | Chow | HP |
| Carbohydrate | 58 | 63 |
| Fat | 13 | 17 |
| Protein | 29 | 20 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsao, C.-W.; Hsu, Y.-J.; Chang, T.-C.; Wu, S.-T.; Cha, T.-L.; Liu, C.-Y. A High Phosphorus Diet Impairs Testicular Function and Spermatogenesis in Male Mice with Chronic Kidney Disease. Nutrients 2020, 12, 2624. https://doi.org/10.3390/nu12092624
Tsao C-W, Hsu Y-J, Chang T-C, Wu S-T, Cha T-L, Liu C-Y. A High Phosphorus Diet Impairs Testicular Function and Spermatogenesis in Male Mice with Chronic Kidney Disease. Nutrients. 2020; 12(9):2624. https://doi.org/10.3390/nu12092624
Chicago/Turabian StyleTsao, Chih-Wei, Yu-Juei Hsu, Ting-Chia Chang, Sheng-Tang Wu, Tai-Lung Cha, and Chin-Yu Liu. 2020. "A High Phosphorus Diet Impairs Testicular Function and Spermatogenesis in Male Mice with Chronic Kidney Disease" Nutrients 12, no. 9: 2624. https://doi.org/10.3390/nu12092624
APA StyleTsao, C. -W., Hsu, Y. -J., Chang, T. -C., Wu, S. -T., Cha, T. -L., & Liu, C. -Y. (2020). A High Phosphorus Diet Impairs Testicular Function and Spermatogenesis in Male Mice with Chronic Kidney Disease. Nutrients, 12(9), 2624. https://doi.org/10.3390/nu12092624