Obesity Measures and Dietary Parameters as Predictors of Gut Microbiota Phyla in Healthy Individuals

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Study Subjects
2.2.1. Dietary Assessment
2.2.2. Body Composition Measurements
2.2.3. Overall Health Status and Quality of Life
2.2.4. Gut Microbiota Composition
2.3. Data Analysis
2.3.1. Cluster Analysis of Gut Microbiota
2.3.2. Predicting Microbiota Clusters Using Support Vector Machine
3. Results
3.1. Characteristics of the Study Subjects
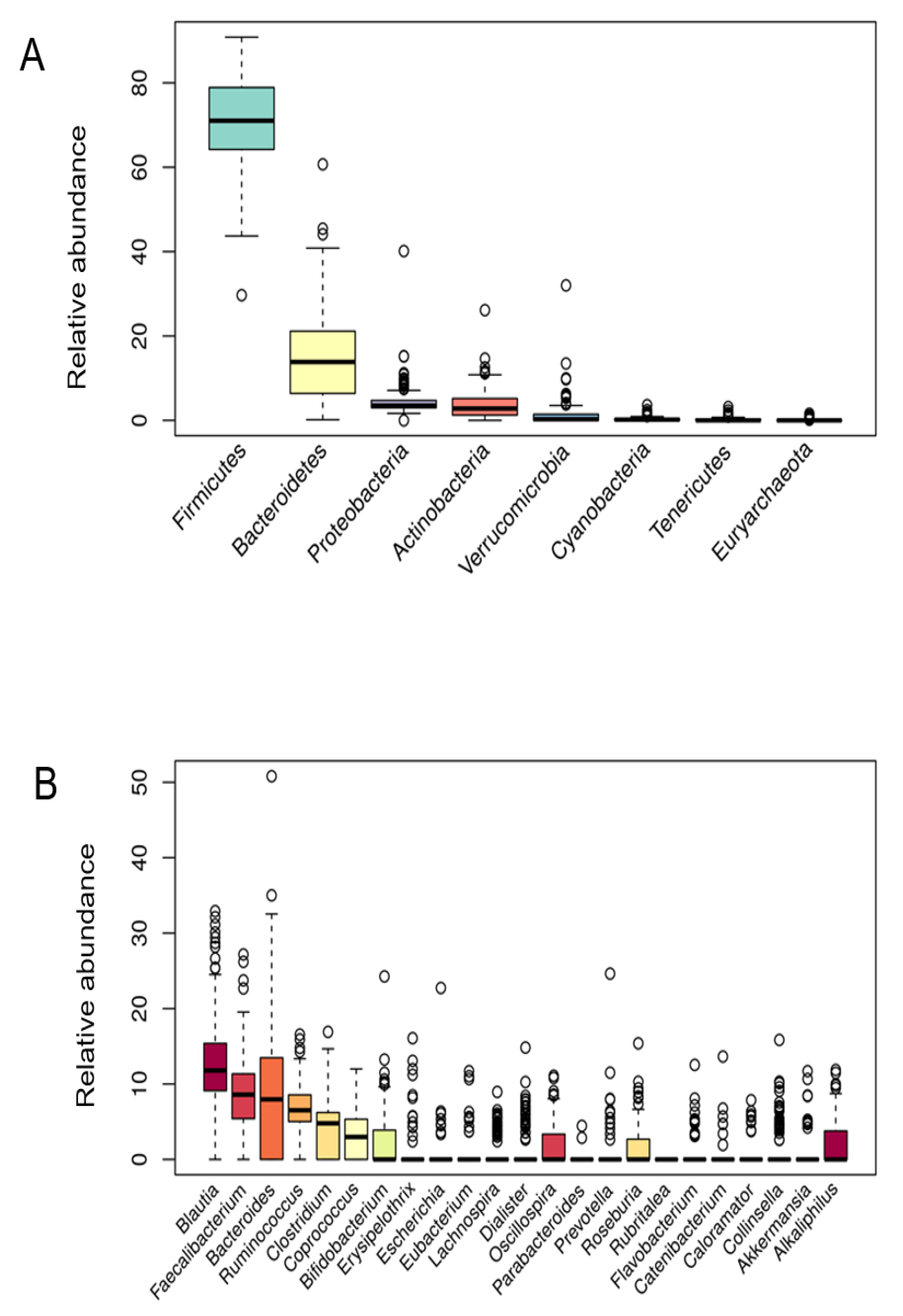
3.2. Microbiota Composition
3.3. Microbiota Cluster Analysis
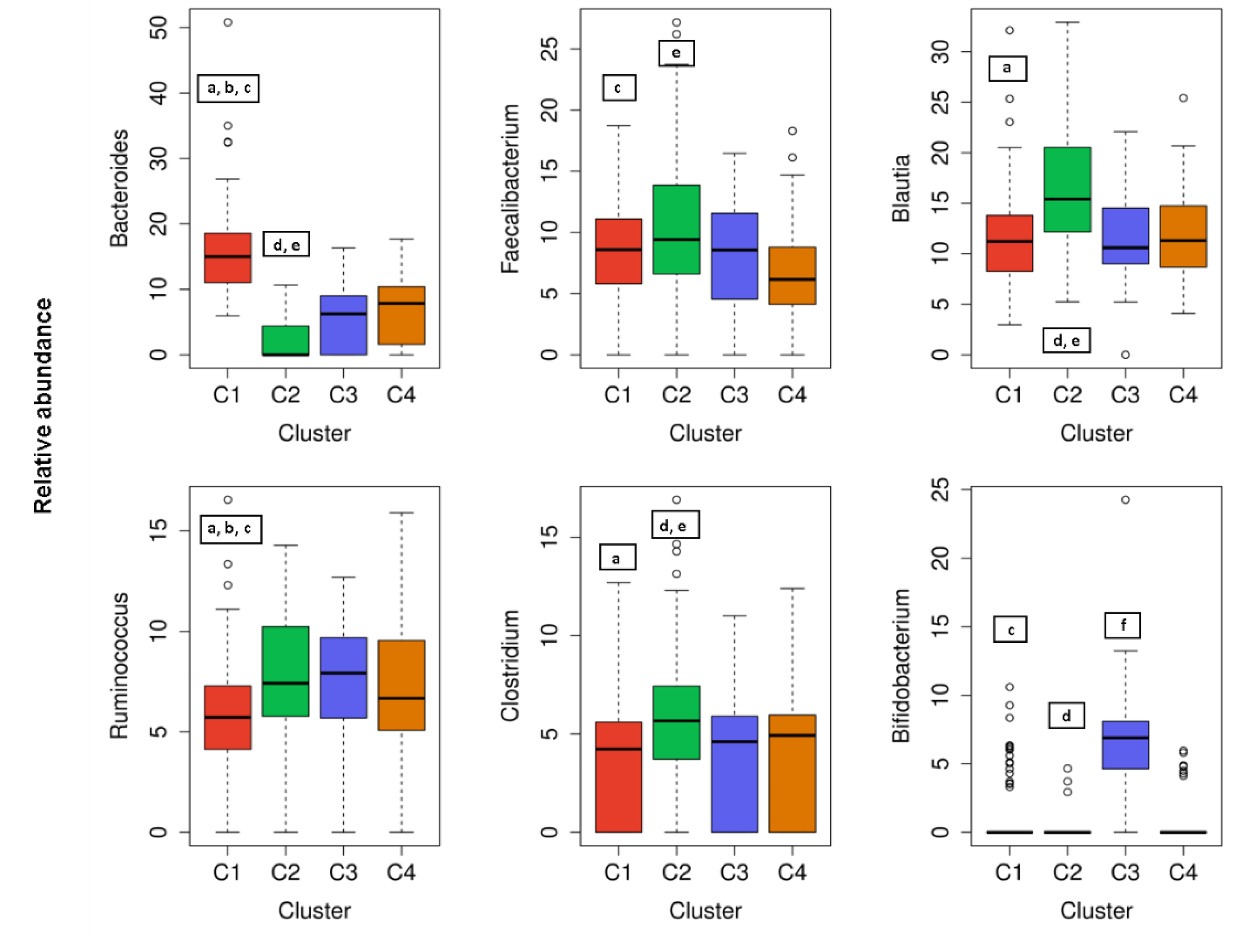
3.3.1. Microbial Abundance over Clusters
3.3.2. The Most Important Variables for Classifying Subjects into Clusters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hillman, E.T.; Lu, H.; Yao, T.; Nakatsu, C.H. Microbial Ecology along the Gastrointestinal Tract. Microbes Environ. 2017, 32, 300–313. [Google Scholar] [CrossRef] [Green Version]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.M.; Finlay, B.B. Gut microbiota in health and disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, N.; Li, N.; Duan, X.; Niu, H. Interaction between the gut microbiome and mucosal immune system. Mil. Med. Res. 2017, 4, 14. [Google Scholar] [CrossRef] [PubMed]
- Wells, J.M.; Brummer, R.J.; Derrien, M.; MacDonald, T.T.; Troost, F.; Cani, P.D.; Theodorou, V.; Dekker, J.; Méheust, A.; de Vos, W.M.; et al. Homeostasis of the gut barrier and potential biomarkers. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 312, G171–G193. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, A.; Mazmanian, S.K. Disruption of the gut microbiome as a risk factor for microbial infections. Curr. Opin. Microbiol. 2013, 16, 221–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, A.M.; Sun, E.W.; Rogers, G.B.; Keating, D.J. The Influence of the Gut Microbiome on Host Metabolism through the Regulation of Gut Hormone Release. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.; DeCoffe, D.; Molcan, E.; Gibson, D.L. Diet-Induced Dysbiosis of the Intestinal Microbiota and the Effects on Immunity and Disease. Nutrients 2012, 4, 1095–1119. [Google Scholar] [CrossRef] [Green Version]
- Arora, T.; Bäckhed, F. The gut microbiota and metabolic disease: Current understanding and future perspectives. J. Intern. Med. 2016, 280, 339–349. [Google Scholar] [CrossRef]
- Grochowska, M.; Laskus, T.; Radkowski, M. Gut Microbiota in Neurological Disorders. Arch. Immunol. Ther. Exp. 2019, 67, 375–383. [Google Scholar] [CrossRef] [Green Version]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.-M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Castaño-Rodríguez, N.; Underwood, A.P.; Merif, J.; Riordan, S.M.; Rawlinson, W.D.; Mitchell, H.M.; Kaakoush, N.O. Gut Microbiome Analysis Identifies Potential Etiological Factors in Acute Gastroenteritis. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [Green Version]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Mantrana, I.; Selma-Royo, M.; Alcantara, C.; Collado, M.C. Shifts on Gut Microbiota Associated to Mediterranean Diet Adherence and Specific Dietary Intakes on General Adult Population. Front. Microbiol. 2018, 9, 890. [Google Scholar] [CrossRef]
- Jackson, M.A.; Verdi, S.; Maxan, M.-E.; Shin, C.M.; Zierer, J.; Bowyer, R.C.E.; Martin, T.; Williams, F.M.K.; Menni, C.; Bell, J.T.; et al. Gut microbiota associations with common diseases and prescription medications in a population-based cohort. Nat. Commun. 2018, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Zhernakova, A.; Kurilshikov, A.; Bonder, M.J.; Tigchelaar, E.F.; Schirmer, M.; Vatanen, T.; Mujagic, Z.; Vila, A.V.; Falony, G.; Vieira-Silva, S.; et al. Population-based metagenomics analysis reveals markers for gut microbiome composition and diversity. Science 2016, 352, 565–569. [Google Scholar] [CrossRef] [Green Version]
- Scepanovic, P.; Hodel, F.; Mondot, S.; Partula, V.; Byrd, A.; Hammer, C.; Alanio, C.; Bergstedt, J.; Patin, E.; Touvier, M.; et al. A comprehensive assessment of demographic, environmental, and host genetic associations with gut microbiome diversity in healthy individuals. Microbiome 2019, 7, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Statnikov, A.; Henaff, M.; Narendra, V.; Konganti, K.; Li, Z.; Yang, L.; Pei, Z.; Blaser, M.J.; Aliferis, C.F.; Alekseyenko, A.V. A comprehensive evaluation of multicategory classification methods for microbiomic data. Microbiome 2013, 1, 11. [Google Scholar] [CrossRef] [PubMed]
- Bizjak, M.; Jenko-Pražnikar, Z.; Koroušić Seljak, B. Development and validation of an electronic FFQ to assess food intake in the Slovene population. Public Health Nutr. 2014, 17, 1729–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-González, M.A.; García-Arellano, A.; Toledo, E.; Salas-Salvadó, J.; Buil-Cosiales, P.; Corella, D.; Covas, M.I.; Schröder, H.; Arós, F.; Gómez-Gracia, E.; et al. A 14-Item Mediterranean Diet Assessment Tool and Obesity Indexes among High-Risk Subjects: The PREDIMED Trial. PLoS ONE 2012, 7, e43134. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Villegas, A.; De Irala, J.; Martínez-González, M.A. Research team of the SUN (University of Navarre Follow-Up Study) project [The Mediterranean diet and cardiovascular disease: Results of a pilot study from the SUN project (University of Navarre Follow-Up Study)]. Rev. Med. Univ. Navar. 2002, 46, 9–16. [Google Scholar]
- Martínez-Gómez, D.; Eisenmann, J.C.; Gómez-Martínez, S.; Veses, A.; Marcos, A.; Veiga, O.L. Sedentarismo, adiposidad y factores de riesgo cardiovascular en adolescentes. Estudio AFINOS. Rev. Esp. Cardiol. 2010, 63, 277–285. [Google Scholar] [CrossRef]
- Veiga, O.L.; Gómez-Martínez, S.; Martínez-Gómez, D.; Villagra, A.; Calle, M.E.; Marcos, A. AFINOS Study Group Physical activity as a preventive measure against overweight, obesity, infections, allergies and cardiovascular disease risk factors in adolescents: AFINOS Study protocol. BMC Public Health 2009, 9, 475. [Google Scholar] [CrossRef] [Green Version]
- Redondo-Useros, N.; Gheorghe, A.; Díaz-Prieto, L.E.; Villavisencio, B.; Marcos, A.; Nova, E. Associations of Probiotic Fermented Milk (PFM) and Yogurt Consumption with Bifidobacterium and Lactobacillus Components of the Gut Microbiota in Healthy Adults. Nutrients 2019, 11, 651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago, A.; Panda, S.; Mengels, G.; Martinez, X.; Azpiroz, F.; Dore, J.; Guarner, F.; Manichanh, C. Processing faecal samples: A step forward for standards in microbial community analysis. BMC Microbiol. 2014, 14, 112. [Google Scholar] [CrossRef] [Green Version]
- Illumina Read Classification. Available online: https://support.illumina.com/help/BaseSpace_App_16S_Metagenomics_help/16S_Metagenomics_App_Help.htm#Vault/Informatics/Sequencing_Analysis/Apps/swSEQ_mAPP_16S_Read_Classification.htm%3FToc (accessed on 20 August 2020).
- Zhong, H.; Penders, J.; Shi, Z.; Ren, H.; Cai, K.; Fang, C.; Ding, Q.; Thijs, C.; Blaak, E.E.; Stehouwer, C.D.A.; et al. Impact of early events and lifestyle on the gut microbiota and metabolic phenotypes in young school-age children. Microbiome 2019, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Falony, G.; Joossens, M.; Vieira-Silva, S.; Wang, J.; Darzi, Y.; Faust, K.; Kurilshikov, A.; Bonder, M.J.; Valles-Colomer, M.; Vandeputte, D.; et al. Population-level analysis of gut microbiome variation. Science 2016, 352, 560–564. [Google Scholar] [CrossRef]
- Zoetendal, E.G.; Rajilić-Stojanović, M.; Vos, W.M. de High-throughput diversity and functionality analysis of the gastrointestinal tract microbiota. Gut 2008, 57, 1605–1615. [Google Scholar] [CrossRef] [PubMed]
- Koliada, A.; Syzenko, G.; Moseiko, V.; Budovska, L.; Puchkov, K.; Perederiy, V.; Gavalko, Y.; Dorofeyev, A.; Romanenko, M.; Tkach, S.; et al. Association between body mass index and Firmicutes/Bacteroidetes ratio in an adult Ukrainian population. BMC Microbiol. 2017, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haro, C.; Rangel-Zúñiga, O.A.; Alcalá-Díaz, J.F.; Gómez-Delgado, F.; Pérez-Martínez, P.; Delgado-Lista, J.; Quintana-Navarro, G.M.; Landa, B.B.; Navas-Cortés, J.A.; Tena-Sempere, M.; et al. Intestinal Microbiota Is Influenced by Gender and Body Mass Index. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silveira-Nunes, G.; Durso, D.F.; de Oliveira, L.R.A., Jr.; Cunha, E.H.M.; Maioli, T.U.; Vieira, A.T.; Speziali, E.; Corrêa-Oliveira, R.; Martins-Filho, O.A.; Teixeira-Carvalho, A.; et al. Hypertension Is Associated With Intestinal Microbiota Dysbiosis and Inflammation in a Brazilian Population. Front. Pharmacol. 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; Li, E.; Ahmari, N.; Carvajal, J.M.; Zadeh, M.; Gong, M.; Qi, Y.; Zubcevic, J.; et al. Gut microbiota dysbiosis is linked to hypertension. Hypertension 2015, 65, 1331–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, H.-J.; Nguyen, T.T.B.; Kim, H.-J.; Hong, S.-T. Gut Microbiota as a Missing Link Between Nutrients and Traits of Human. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graf, D.; Di Cagno, R.; Fåk, F.; Flint, H.J.; Nyman, M.; Saarela, M.; Watzl, B. Contribution of diet to the composition of the human gut microbiota. Microb. Ecol. Health Dis. 2015, 26, 26164. [Google Scholar] [CrossRef]
- Martínez-González, M.Á.; Hershey, M.S.; Zazpe, I.; Trichopoulou, A. Transferability of the Mediterranean Diet to Non-Mediterranean Countries. What Is and What Is Not the Mediterranean Diet. Nutrients 2017, 9, 1226. [Google Scholar] [CrossRef] [Green Version]
- Nagpal, R.; Shively, C.A.; Register, T.C.; Craft, S.; Yadav, H. Gut microbiome-Mediterranean diet interactions in improving host health. F1000Res 2019, 8, 699. [Google Scholar] [CrossRef] [Green Version]
- Wolters, M.; Ahrens, J.; Romaní-Pérez, M.; Watkins, C.; Sanz, Y.; Benítez-Páez, A.; Stanton, C.; Günther, K. Dietary fat, the gut microbiota, and metabolic health—A systematic review conducted within the MyNewGut project. Clin. Nutr. 2019, 38, 2504–2520. [Google Scholar] [CrossRef] [Green Version]
- Partula, V.; Mondot, S.; Torres, M.J.; Kesse-Guyot, E.; Deschasaux, M.; Assmann, K.; Latino-Martel, P.; Buscail, C.; Julia, C.; Galan, P.; et al. Associations between usual diet and gut microbiota composition: Results from the Milieu Intérieur cross-sectional study. Am. J. Clin. Nutr. 2019, 109, 1472–1483. [Google Scholar] [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Watson, H.; Mitra, S.; Croden, F.C.; Taylor, M.; Wood, H.M.; Perry, S.L.; Spencer, J.A.; Quirke, P.; Toogood, G.J.; Lawton, C.L.; et al. A randomised trial of the effect of omega-3 polyunsaturated fatty acid supplements on the human intestinal microbiota. Gut 2018, 67, 1974–1983. [Google Scholar] [CrossRef] [PubMed]
- Bindels, L.B.; Delzenne, N.M.; Cani, P.D.; Walter, J. Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.L.; Erickson, J.M.; Lloyd, B.B.; Slavin, J.L. Health Effects and Sources of Prebiotic Dietary Fiber. Curr. Dev. Nutr. 2018, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [Green Version]
- Zimmer, J.; Lange, B.; Frick, J.-S.; Sauer, H.; Zimmermann, K.; Schwiertz, A.; Rusch, K.; Klosterhalfen, S.; Enck, P. A vegan or vegetarian diet substantially alters the human colonic faecal microbiota. Eur. J. Clin. Nutr. 2012, 66, 53–60. [Google Scholar] [CrossRef]
- Volokh, O.; Klimenko, N.; Berezhnaya, Y.; Tyakht, A.; Nesterova, P.; Popenko, A.; Alexeev, D. Human Gut Microbiome Response Induced by Fermented Dairy Product Intake in Healthy Volunteers. Nutrients 2019, 11, 547. [Google Scholar] [CrossRef] [Green Version]
- Hansen, T.H.; Thomassen, M.T.; Madsen, M.L.; Kern, T.; Bak, E.G.; Kashani, A.; Allin, K.H.; Hansen, T.; Pedersen, O. The effect of drinking water pH on the human gut microbiota and glucose regulation: Results of a randomized controlled cross-over intervention. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Engen, P.A.; Green, S.J.; Voigt, R.M.; Forsyth, C.B.; Keshavarzian, A. The Gastrointestinal Microbiome: Alcohol Effects on the Composition of Intestinal Microbiota. Alcohol. Res. 2015, 37, 223–236. [Google Scholar] [PubMed]
- Liu, J.; Liu, X.; Xiong, X.-Q.; Yang, T.; Cui, T.; Hou, N.-L.; Lai, X.; Liu, S.; Guo, M.; Liang, X.-H.; et al. Effect of vitamin A supplementation on gut microbiota in children with autism spectrum disorders—A pilot study. BMC Microbiol. 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Gurwara, S.; Ajami, N.J.; Jang, A.; Hessel, F.C.; Chen, L.; Plew, S.; Wang, Z.; Graham, D.Y.; Hair, C.; White, D.L.; et al. Dietary Nutrients Involved in One-Carbon Metabolism and Colonic Mucosa-Associated Gut Microbiome in Individuals with an Endoscopically Normal Colon. Nutrients 2019, 11, 613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, S.; Saunier, K.; Hanisch, C.; Norin, E.; Alm, L.; Midtvedt, T.; Cresci, A.; Silvi, S.; Orpianesi, C.; Verdenelli, M.C.; et al. Differences in Fecal Microbiota in Different European Study Populations in Relation to Age, Gender, and Country: A Cross-Sectional Study. Appl. Environ. Microbiol. 2006, 72, 1027–1033. [Google Scholar] [CrossRef] [Green Version]
- Dominianni, C.; Sinha, R.; Goedert, J.J.; Pei, Z.; Yang, L.; Hayes, R.B.; Ahn, J. Sex, body mass index, and dietary fiber intake influence the human gut microbiome. PLoS ONE 2015, 10, e0124599. [Google Scholar] [CrossRef] [Green Version]
- Salazar, N.; Arboleya, S.; Fernández-Navarro, T.; de los Reyes-Gavilán, C.G.; Gonzalez, S.; Gueimonde, M. Age-Associated Changes in Gut Microbiota and Dietary Components Related with the Immune System in Adulthood and Old Age: A Cross-Sectional Study. Nutrients 2019, 11, 1765. [Google Scholar] [CrossRef] [Green Version]
- Odamaki, T.; Kato, K.; Sugahara, H.; Hashikura, N.; Takahashi, S.; Xiao, J.; Abe, F.; Osawa, R. Age-related changes in gut microbiota composition from newborn to centenarian: A cross-sectional study. BMC Microbiol. 2016, 16. [Google Scholar] [CrossRef] [Green Version]
- Lan, Y.; Kriete, A.; Rosen, G.L. Selecting age-related functional characteristics in the human gut microbiome. Microbiome 2013, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Zhu, H.; Qiu, P. Aging progression of human gut microbiota. BMC Microbiol. 2019, 19, 236. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.P.; Easson, C.; Lyle, S.M.; Kapoor, R.; Donnelly, C.P.; Davidson, E.J.; Parikh, E.; Lopez, J.V.; Tartar, J.L. Gut microbiome diversity is associated with sleep physiology in humans. PLoS ONE 2019, 14, e0222394. [Google Scholar] [CrossRef]
- Benedict, C.; Vogel, H.; Jonas, W.; Woting, A.; Blaut, M.; Schürmann, A.; Cedernaes, J. Gut microbiota and glucometabolic alterations in response to recurrent partial sleep deprivation in normal-weight young individuals. Mol. Metab. 2016, 5, 1175–1186. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | |
|---|---|
| Age years (mean ± SD) | 35.4 ± 7.0 |
| Gender | |
| Female | 117 (58.5%) |
| Male | 83 (41.5%) |
| Level of education | |
| Primary education | 5 (2.5%) |
| Secondary education or high school | 68 (34%) |
| University degree (Bachelor’s, Master’s, Doctor’s) | 127 (63.5%) |
| BMI kg/m2 (mean ± SD) | 24.2 ± 3.5 |
| Weight (kg) (mean ± SD) | 73.1 ± 15.4 |
| WHR = waist (cm)/hip (cm) (mean ± SD) | 0.87 ± 0.07 |
| Visceral fat index | 4.7 ± 2.9 |
| Blood pressure | |
| Systolic | 125.8 ± 13.2 |
| Diastolic | 81.3 ± 9.8 |
| Hours of sleep (mean ± SD) | 7.7 ± 0.8 |
| Smoker | |
| Yes | 54 (27%) |
| No | 146 (73%) |
| Alcohol consumption habits (mean g/day ± SD) | 14.5 ± 13.6 |
| Up to 5 g/day | 78 (39%) |
| 5–12 g/day (W) and 5–20 g/day (M) | 60 (30%) |
| >12 g/day (W) and >20 g/day (M) | 62 (31%) |
| Dietary intake | |
| EER kcal (mean ± SD) | 3093 ± 776 |
| Energy intake (kcal/day) | 1892 ± 1000 |
| Total fat (%) | 32.07 ± 9.96 |
| Carbohydrate (%) | 49.05 ± 10.18 |
| Protein (%) | 18.14 ± 6.48 |
| Fiber (g/day) | 19.37 ± 13.67 |
| MEDAS (mean ± SD) | 7.7 ± 2.4 |
| 0–6 points | 60 (30%) |
| 7–10 points | 118 (59%) |
| 11–14 points | 22 (11%) |
| Physical activity (kcal/week) | 4219 ± 335 |
| Clusters of Subjects | ||||||||
|---|---|---|---|---|---|---|---|---|
| C1 | C2 | C3 | C4 | |||||
| Predictor and Relative Value of Importance | ||||||||
| 1 | Total fiber intake | 86.1 | BMI | 88.2 | Fat intake | 91.0 | Water intake | 100 |
| 2 | BMI | 81.2 | Blood pressure | 81.7 | Water intake | 90.2 | Total fiber intake | 86.1 |
| 3 | Water intake | 81.1 | Total body water | 77.7 | BMI | 81.3 | Visceral fat index | 77.7 |
| 4 | Blood pressure | 77.4 | Waist/hip ratio | 76.5 | Energy intake | 81.1 | Energy intake | 77.4 |
| 5 | Energy intake | 76.4 | Visceral fat index | 73.8 | Visceral fat index | 77.7 | Olive oil intake | 74.8 |
| 6 | Olive oil intake | 74.1 | Fat intake | 68.7 | Vitamin A intake | 76.7 | Milk and milk products intake | 74.4 |
| 7 | Visceral fat index | 73.5 | Saturated fatty acids intake | 68.7 | Vitamin B9 intake | 75.2 | BMI | 68.8 |
| 8 | Waist/hip ratio | 68.0 | Waist/hip ratio | 64.9 | Sleeping hours | 73.5 | Carbohydrates intake | 68.4 |
| 9 | Milk and milk products intake | 67.9 | Gender | 64.7 | Age | 68.0 | Fat intake | 67.9 |
| 10 | Fat intake | 64.0 | Energy requirements | 64.7 | Gender | 67.7 | Age | 60.5 |
| 11 | Saturated fatty acids intake | 63.3 | Legume intake | 63.5 | Saturated fatty acids intake | 64.3 | MEDAS | 57.7 |
| 12 | Gender | 62.1 | Protein intake | 60.8 | Monounsaturated fatty acids intake | 63.0 | Monounsaturated fatty acids intake | 57.7 |
| 13 | Energy requirements | 58.2 | Vegetable fiber intake | 60.2 | MEDAS | 62.1 | Protein intake | 57.3 |
| 14 | Carbohydrates intake | 57.7 | Metabolic age | 59.8 | Vegetable fiber intake | 60.3 | Vegetable fiber intake | 56.6 |
| 15 | Age | 57.7 | Carbohydrates intake | 59.3 | Vitamin E intake | 59.9 | Energy requirements | 56.2 |
| Clusters | ||||
|---|---|---|---|---|
| Predictors | C1 (n = 80) | C2 (n = 55) | C3 (n = 34) | C4 (n = 31) |
| Gender: n (F/M); % (F/M) | 54/26; 68/32 a,b | 21/34; 38/62 d,e | 19/15; 55/45 f | 22/9; 71/29 |
| Age (years) | 36.0 ± 7.4 b | 36.4 ± 6.2 d | 32.6 ± 6.1 f | 35.8 ± 6.2 |
| Obesity parameters | ||||
| BMI (kg/m2) | 23.1 ± 3.3 a,b,c | 25.5 ± 3.4 d,e | 24.7 ± 3.3 | 24.0 ± 3.3 |
| BMI > 25 kg/m2: n; % | 20; 20% a,b,c | 31; 56% d,e | 14; 41% | 12; 39% |
| Waist to hip ratio | 0.85 ± 0.08 a | 0.89 ± 0.06 e | 0.87 ± 0.07 | 0.85 ± 0.05 |
| Visceral fat index | 3.9 ± 2.8 a,b,c | 5.7 ± 2.9 d,e | 4.8 ± 2.7 | 4.6 ± 3.0 |
| Total body water (%) | 55.6 ± 4.1 c | 55.8 ± 5.8 e | 54.2 ± 6.1 | 52.5 ± 4.3 |
| Blood pressure (mmHg) | 122 ± 12/78 ± 9 a,c | 130 ± 13/84 ± 10 d | 125 ± 12/81 ± 9 | 128 ± 13/84 ± 9 |
| Presence of two metabolic syndrome components: n; % | 6; 7.5% a | 11; 20% d,e | 1; 2.9% | 3; 9.7% |
| Nutrition | ||||
| Energy intake (kcal/day) | 1700 ± 433 | 1720 ± 486 | 1590 ± 1165 | 1666 ± 614 |
| Water intake (g) | 1080 ± 435 b | 1029 ± 322 d | 840 ± 553 f | 1097 ± 615 |
| Fat intake (%) | 30.7 ± 9.6 a | 36.2 ± 10.1 d,e | 28.7 ± 9.3 | 31.8 ± 8.7 |
| Saturated fat intake (%) | 8.3 ± 4.6 a,b | 8.8 ± 4.8 d,e | 7.2 ± 4.0 f | 8.3 ± 4.8 |
| Carbohydrates intake (%) | 50.0 ± 9.0 | 46.4 ± 11.4 d | 52.2 ± 10.4 | 48.2 ± 8.2 |
| Total fiber intake (g/day) | 21.0 ± 14.0 a,b | 17.0 ± 8.8 e | 16.4 ± 12.2 f | 22.1 ± 13.1 |
| Vitamin A (µg/day) | 1352 ± 1837 b | 1420 ± 1585 d | 960 ± 630 | 1243 ± 1178 |
| Vitamin B9 (µg/day) | 408 ± 249 a | 338 ± 283 d,e | 412 ± 222 | 374 ± 208 |
| Olive oil intake (g/day) | 26 ± 24 a,b | 19 ± 21 e | 18 ± 13 f | 28 ± 23 |
| Milk and milk products | 108 ± 101 a,b | 162 ± 149 e | 174 ± 129 f | 124 ± 119 |
| Other parameters | ||||
| Sleeping hours (hours/day) | 7.7 ± 0.7 | 7.5 ± 0.7 d | 8.0 ± 0.7 | 8.0 ± 0.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bezek, K.; Petelin, A.; Pražnikar, J.; Nova, E.; Redondo, N.; Marcos, A.; Jenko Pražnikar, Z. Obesity Measures and Dietary Parameters as Predictors of Gut Microbiota Phyla in Healthy Individuals. Nutrients 2020, 12, 2695. https://doi.org/10.3390/nu12092695
Bezek K, Petelin A, Pražnikar J, Nova E, Redondo N, Marcos A, Jenko Pražnikar Z. Obesity Measures and Dietary Parameters as Predictors of Gut Microbiota Phyla in Healthy Individuals. Nutrients. 2020; 12(9):2695. https://doi.org/10.3390/nu12092695
Chicago/Turabian StyleBezek, Katja, Ana Petelin, Jure Pražnikar, Esther Nova, Noemi Redondo, Ascensión Marcos, and Zala Jenko Pražnikar. 2020. "Obesity Measures and Dietary Parameters as Predictors of Gut Microbiota Phyla in Healthy Individuals" Nutrients 12, no. 9: 2695. https://doi.org/10.3390/nu12092695
APA StyleBezek, K., Petelin, A., Pražnikar, J., Nova, E., Redondo, N., Marcos, A., & Jenko Pražnikar, Z. (2020). Obesity Measures and Dietary Parameters as Predictors of Gut Microbiota Phyla in Healthy Individuals. Nutrients, 12(9), 2695. https://doi.org/10.3390/nu12092695