
Excess Vitamins or Imbalance of Folic Acid and Choline in the Gestational Diet Alter the Gut Microbiota and Obesogenic Effects in Wistar Rat Offspring


Abstract
:1. Introduction
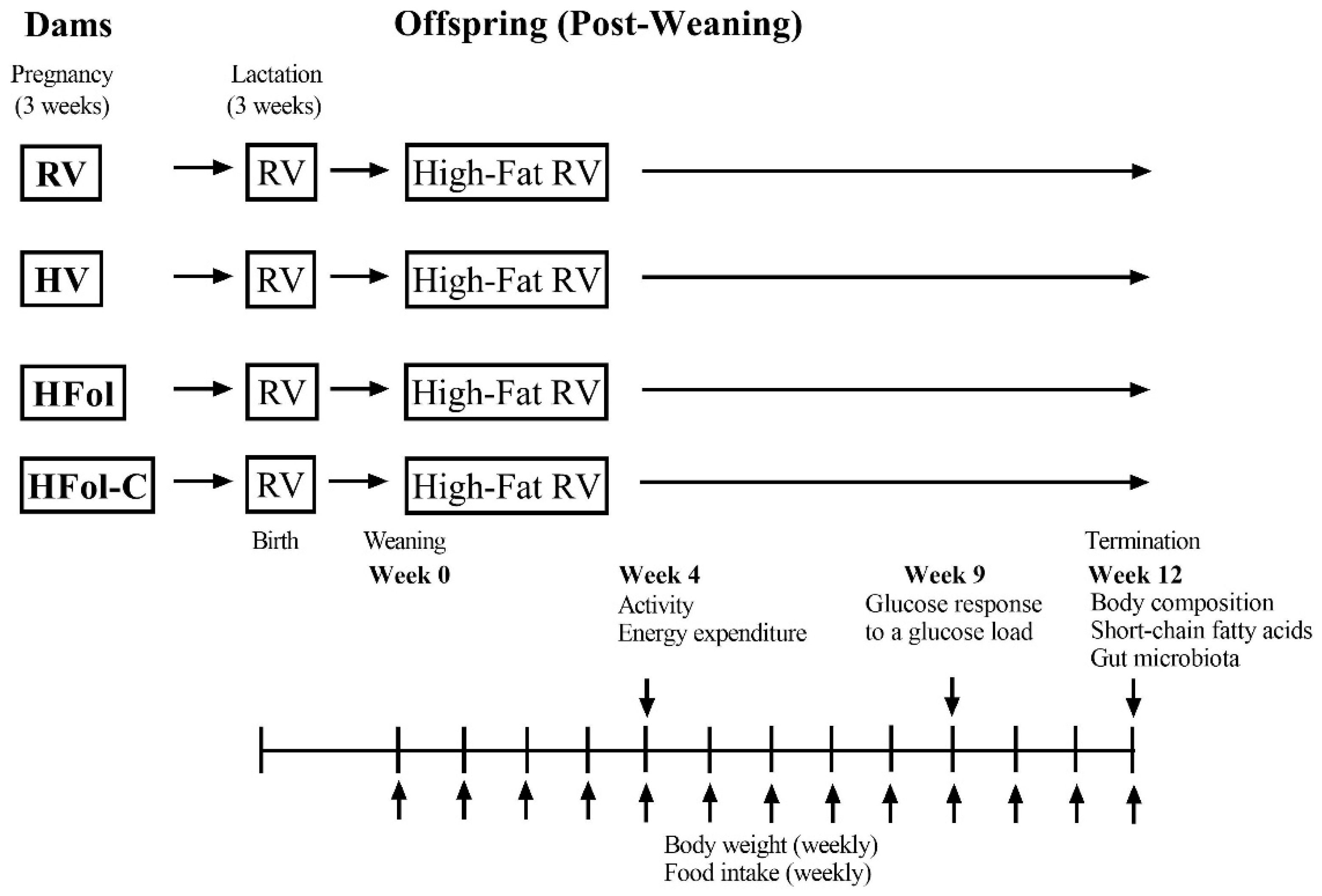
2. Materials and Methods
2.1. Animals and Diets
2.2. Body Weight and Food Intake
2.3. Activity and Energy Expenditure
2.4. Glucose Response to a Glucose Load
2.5. Body Composition and Fecal Sample Collection
2.6. Short-Chain Fatty Acid Analysis
2.7. 16S rRNA Gene Sequencing
2.8. Statistical Analyses
3. Results
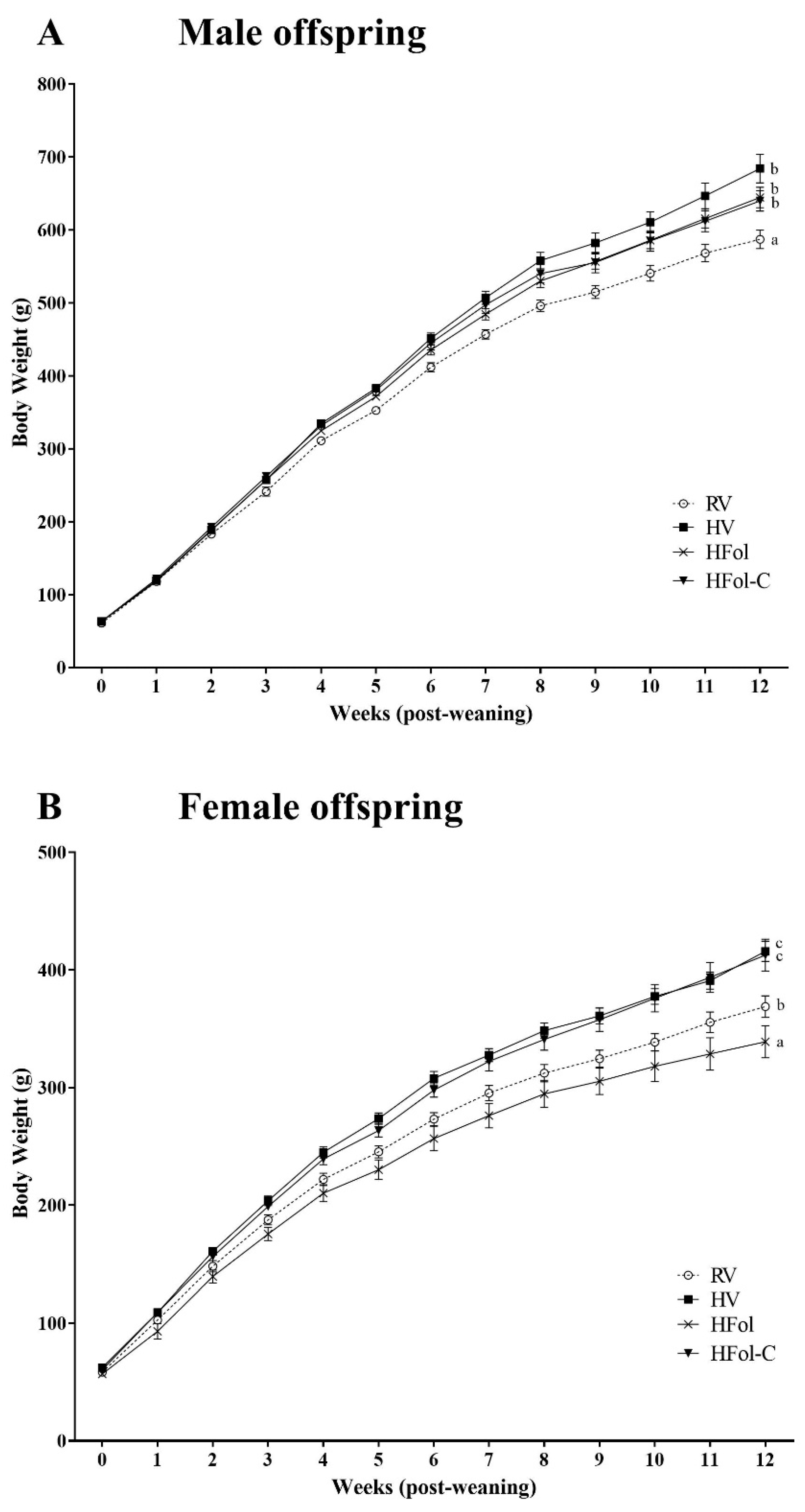
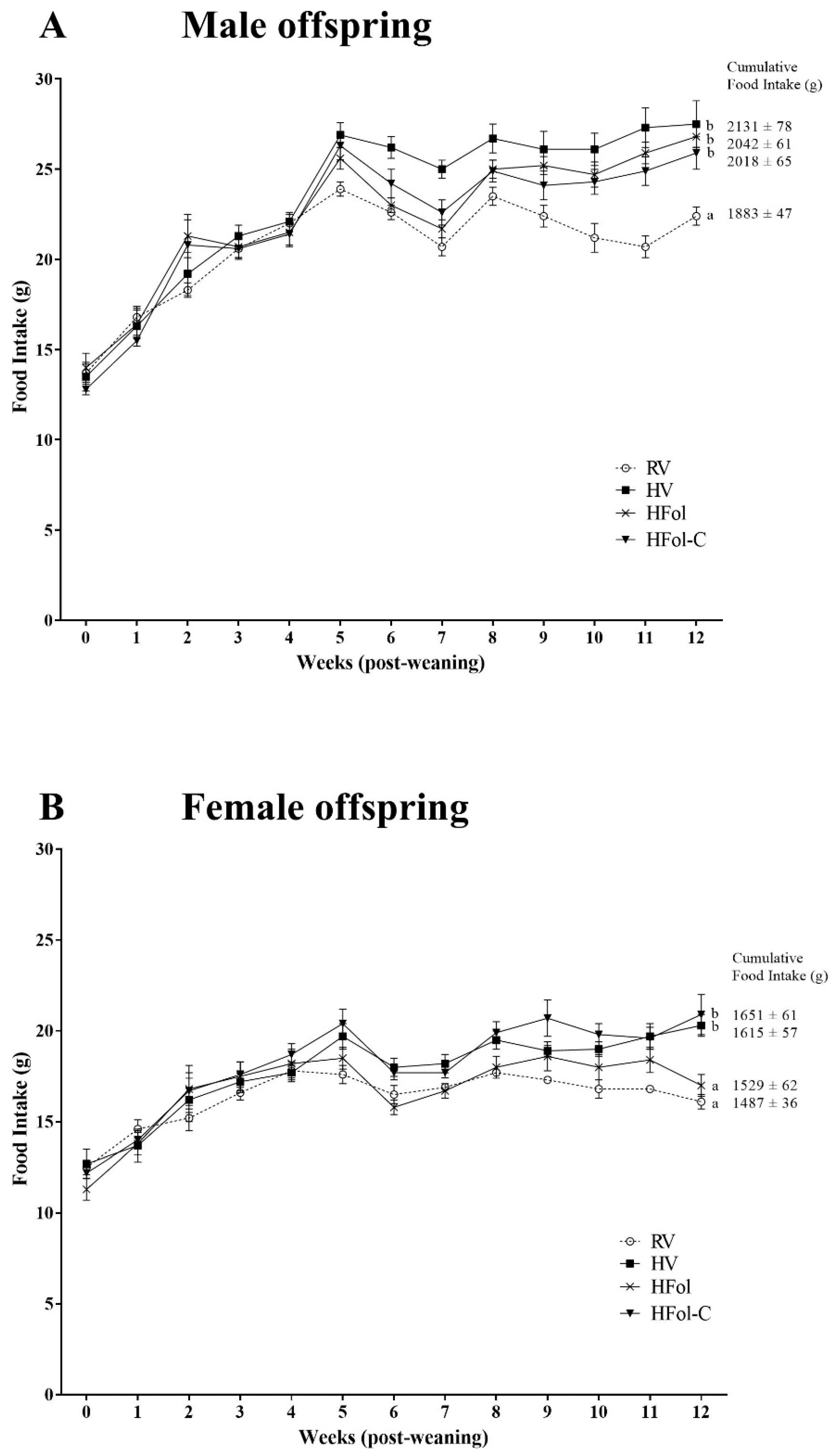
3.1. Body Weight and Food Intake
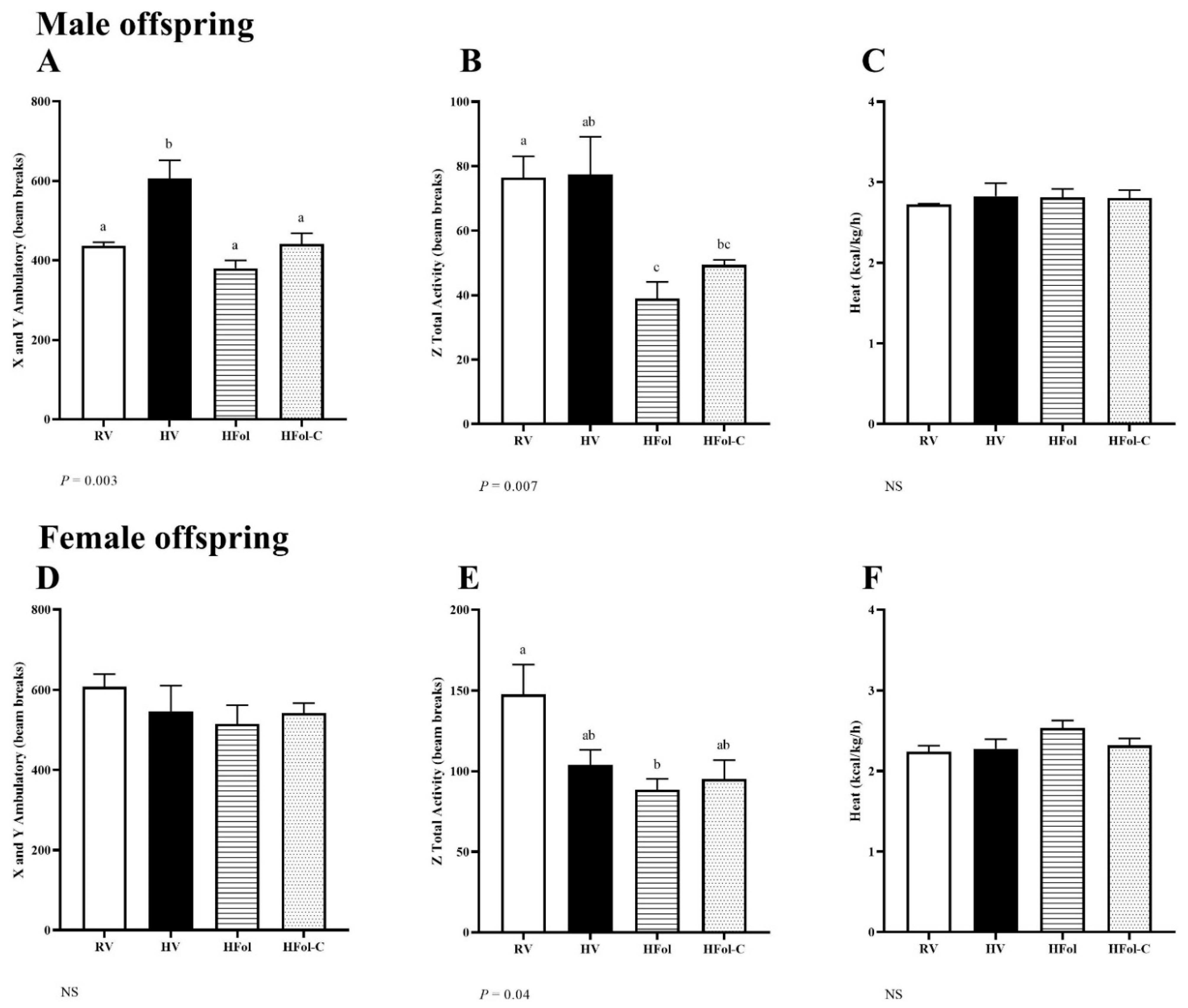
3.2. Activity and Energy Expenditure
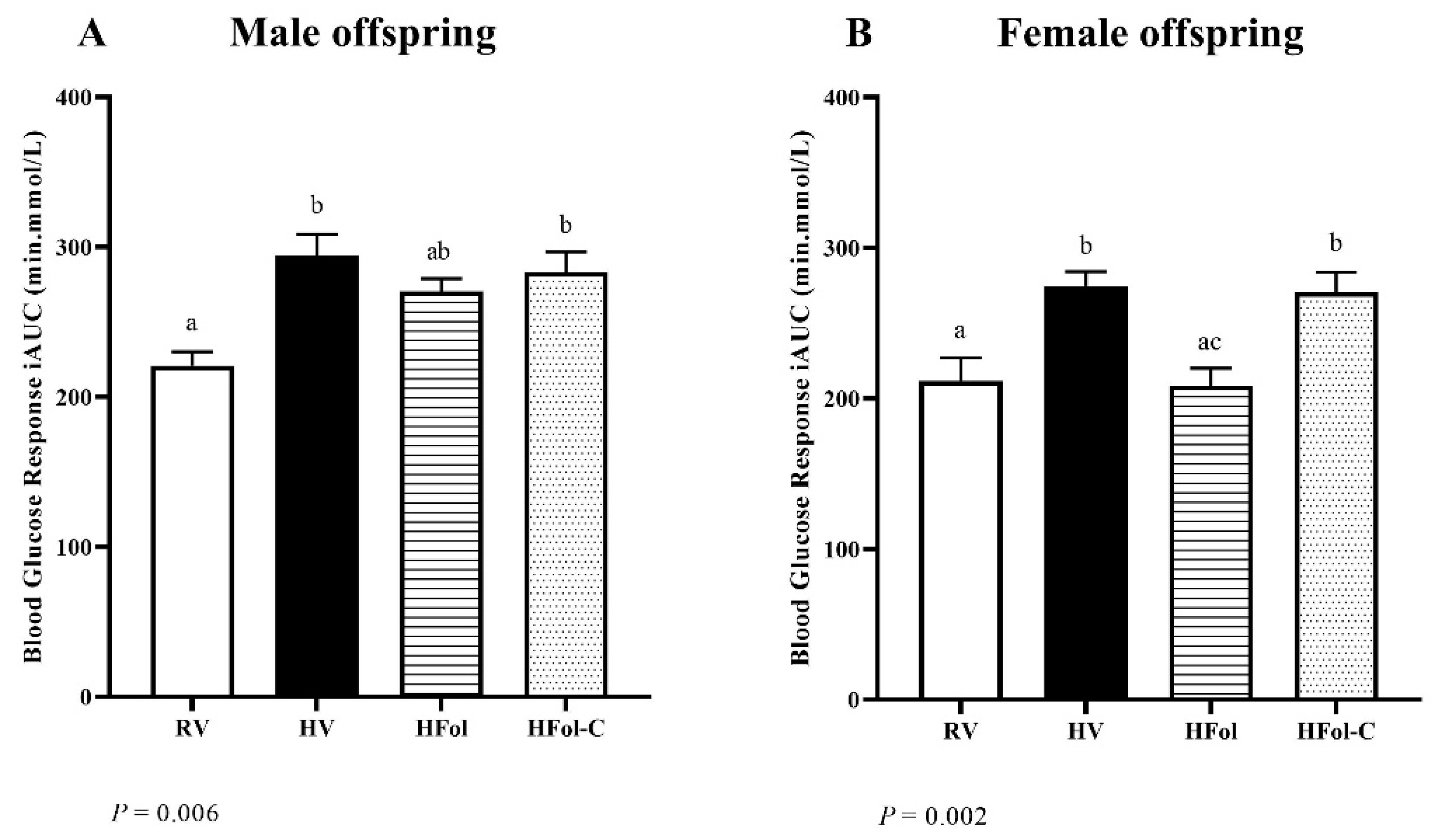
3.3. Glucose Response to a Glucose Load
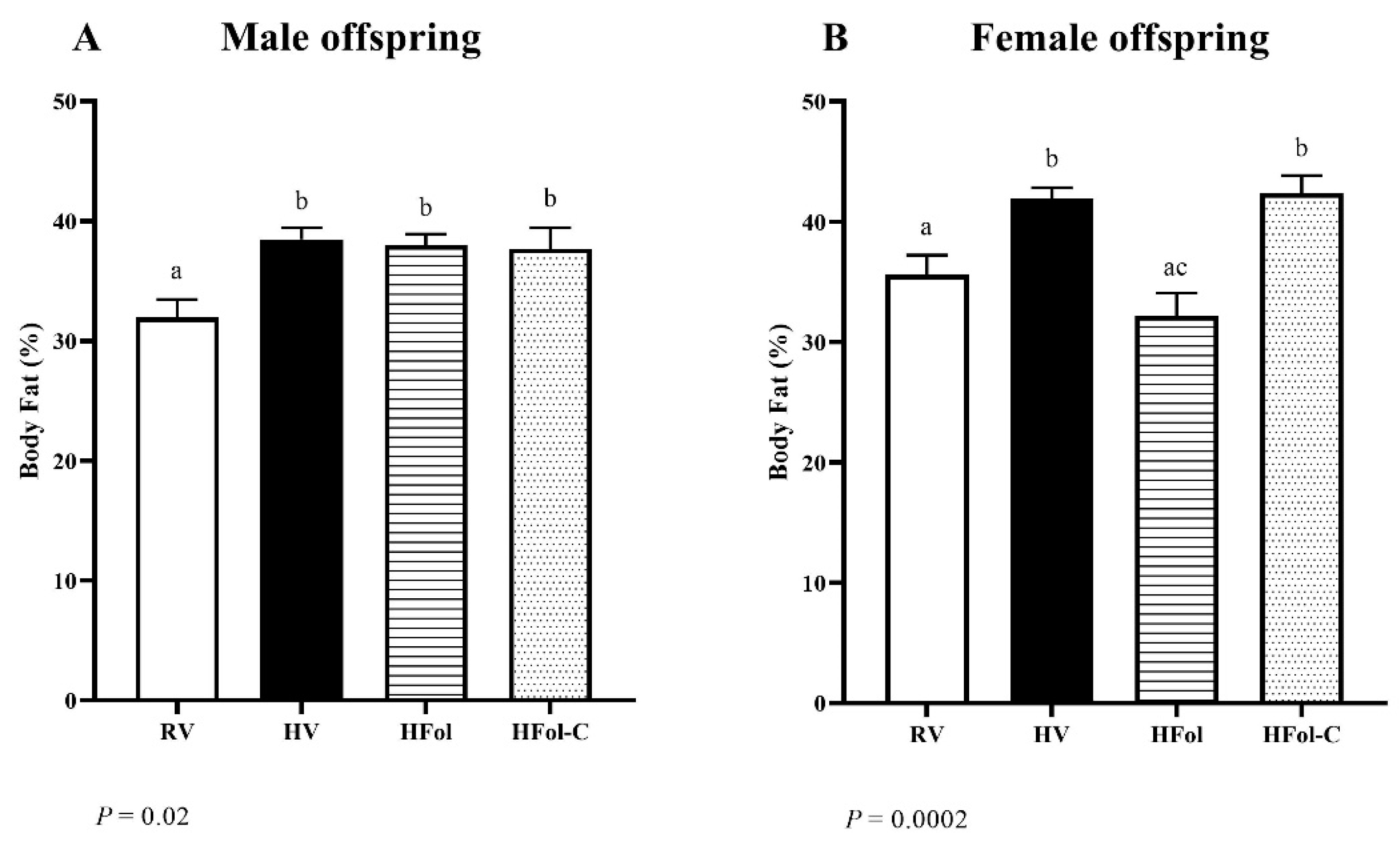
3.4. Body Composition
3.5. Short-Chain Fatty Acid Concentrations
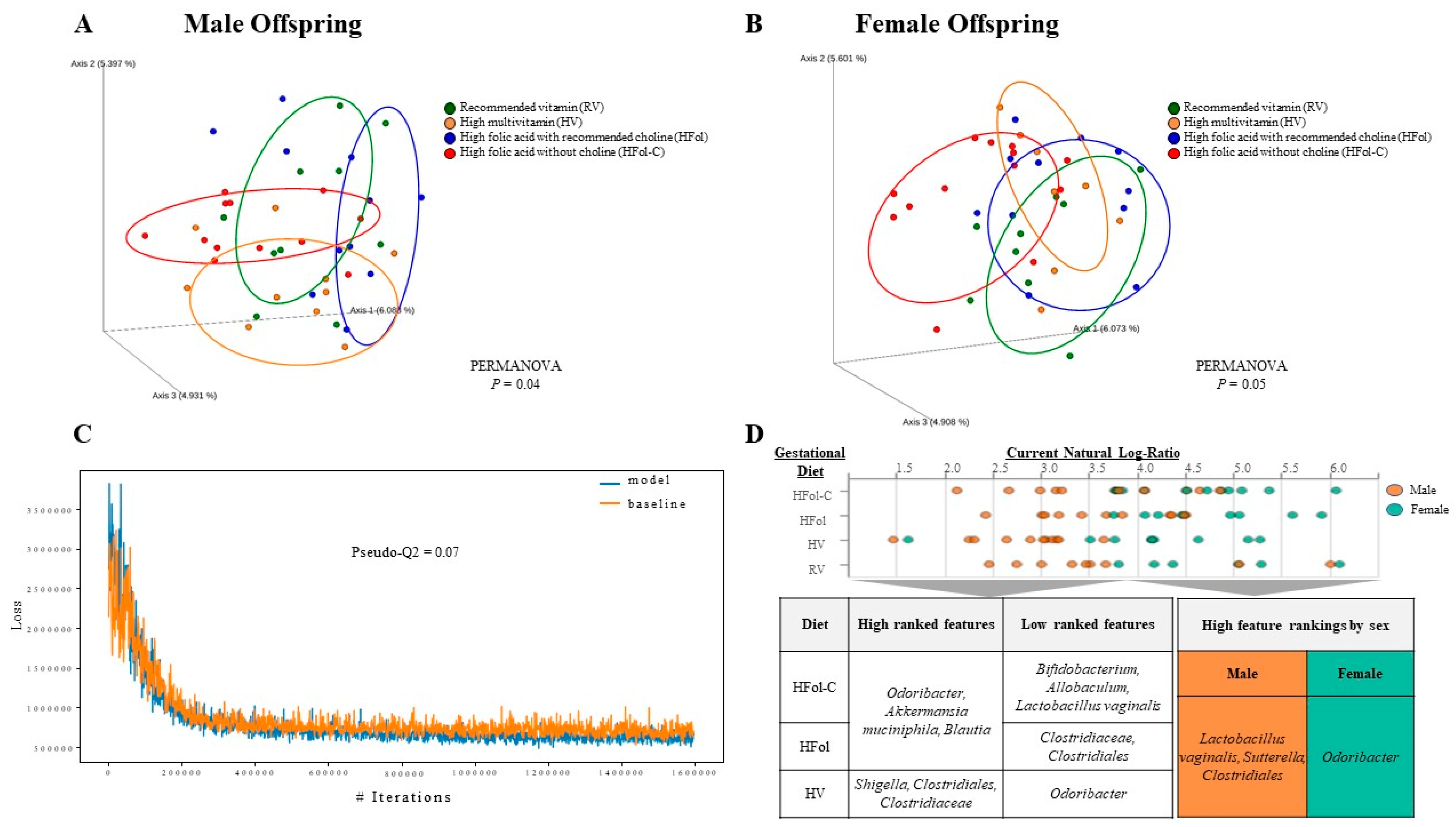
3.6. Gut Microbiota Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Tarry-Adkins, J.L.; Ozanne, S.E. Nutrition in early life and age-associated diseases. Ageing Res. Rev. 2017, 39, 96–105. [Google Scholar] [CrossRef] [Green Version]
- Blackmore, H.L.; Ozanne, S.E. Maternal diet-induced obesity and offspring cardiovascular health. J. Dev. Orig. Health Dis. 2013, 4, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.A.; Gluckman, P.D. Early Developmental Conditioning of Later Health and Disease: Physiology or Pathophysiology? Physiol. Rev. 2014, 94, 1027–1076. [Google Scholar] [CrossRef]
- Jun, S.; Gahche, J.J.; Potischman, N.; Dwyer, J.T.; Guenther, P.M.; Sauder, K.A.; Bailey, R.L. Dietary Supplement Use and Its Micronutrient Contribution During Pregnancy and Lactation in the United States. Obstet. Gynecol. 2020, 135, 623–633. [Google Scholar] [CrossRef] [Green Version]
- Bailey, R.L.; Gahche, J.J.; Lentino, C.V.; Dwyer, J.T.; Engel, J.S.; Thomas, P.R.; Betz, J.M.; Sempos, C.T.; Picciano, M.F. Dietary Supplement Use in the United States, 2003–2006. J. Nutr. 2010, 141, 261–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- E Sacco, J.; Dodd, K.W.; Kirkpatrick, S.; Tarasuk, V. Voluntary food fortification in the United States: Potential for excessive intakes. Eur. J. Clin. Nutr. 2013, 67, 592–597. [Google Scholar] [CrossRef]
- Bailey, R.L.; Pac, S.G.; Fulgoni, V.L., III; Reidy, K.C.; Catalano, P.M. Estimation of Total Usual Dietary Intakes of Pregnant Women in the United States. JAMA Netw. Open 2019, 2, e195967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, C.E.; Hernandez, D.S.; A Reza-López, S.; Huot, P.; Kim, Y.-I.; Anderson, G.H. Obesogenic phenotype of offspring of dams fed a high multivitamin diet is prevented by a post-weaning high multivitamin or high folate diet. Int. J. Obes. 2013, 37, 1177–1182. [Google Scholar] [CrossRef] [Green Version]
- Szeto, I.M.Y.; Das, P.J.; Aziz, A.; Anderson, G.H. Multivitamin supplementation of Wistar rats during pregnancy accelerates the development of obesity in offspring fed an obesogenic diet. Int. J. Obes. 2009, 33, 364–372. [Google Scholar] [CrossRef] [Green Version]
- Cho, C.E.; Pannia, E.; Huot, P.S.P.; Sánchez-Hernández, D.; Kubant, R.; Dodington, D.W.; Ward, W.E.; Bazinet, R.P.; Anderson, G.H. Methyl vitamins contribute to obesogenic effects of a high multivitamin gestational diet and epigenetic alterations in hypothalamic feeding pathways in Wistar rat offspring. Mol. Nutr. Food Res. 2014, 59, 476–489. [Google Scholar] [CrossRef]
- Huot, P.S.P.; Dodington, D.W.; Mollard, R.C.; Reza-López, S.A.; Sanchez-Hernandez, D.; Cho, C.E.; Kuk, J.; Ward, W.E.; Anderson, G.H. High Folic Acid Intake during Pregnancy Lowers Body Weight and Reduces Femoral Area and Strength in Female Rat Offspring. J. Osteoporos. 2013, 2013, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H.; Blusztajn, J.K. Choline and Human Nutrition. Annu. Rev. Nutr. 1994, 14, 269–296. [Google Scholar] [CrossRef] [PubMed]
- Wallace, T.C.; Fulgoni, V.L. Assessment of Total Choline Intakes in the United States. J. Am. Coll. Nutr. 2015, 35, 108–112. [Google Scholar] [CrossRef]
- Corbin, K.D.; Zeisel, S.H. The Nutrigenetics and Nutrigenomics of the Dietary Requirement for Choline. Prog. Mol. Biol. Transl. Sci. 2012, 108, 159–177. [Google Scholar] [CrossRef]
- Selhub, J.; Seyoum, E.; A Pomfret, E.; Zeisel, S.H. Effects of choline deficiency and methotrexate treatment upon liver folate content and distribution. Cancer Res. 1991, 51, 16–21. [Google Scholar] [PubMed]
- Varela-Moreiras, G.; Ragel, C.; de Miguelsanz, J.P. Choline deficiency and methotrexate treatment induces marked but reversible changes in hepatic folate concentrations, serum homocysteine and DNA methylation rates in rats. J. Am. Coll. Nutr. 1995, 14, 480–485. [Google Scholar] [CrossRef]
- Horne, D.W.; Cook, R.J.; Wagner, C. Effect of Dietary Methyl Group Deficiency on Folate Metabolism in Rats. J. Nutr. 1989, 119, 618–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinert, R.E.; Lee, Y.-K.; Sybesma, W. Vitamins for the Gut Microbiome. Trends Mol. Med. 2019, 26, 137–140. [Google Scholar] [CrossRef]
- Gurwara, S.; Ajami, N.J.; Jang, A.; Hessel, F.C.; Chen, L.; Plew, S.; Wang, Z.; Graham, D.Y.; Hair, C.; White, D.L.; et al. Dietary Nutrients Involved in One-Carbon Metabolism and Colonic Mucosa-Associated Gut Microbiome in Individuals with an Endoscopically Normal Colon. Nutrients 2019, 11, 613. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, A.V.A.; Oliveira, S.B.D.C.; Di Rienzi, S.C.; Brown-Steinke, K.; Dehan, L.M.; Rood, J.K.; Carreira, V.S.; Le, H.; A Maier, E.; Betz, K.J.; et al. Murine Methyl Donor Deficiency Impairs Early Growth in Association with Dysmorphic Small Intestinal Crypts and Reduced Gut Microbial Community Diversity. Curr. Dev. Nutr. 2018, 3. [Google Scholar] [CrossRef]
- Sharma, M.; Li, Y.; Stoll, M.L.; Tollefsbol, T.O. The Epigenetic Connection Between the Gut Microbiome in Obesity and Diabetes. Front. Genet. 2020, 10, 1329. [Google Scholar] [CrossRef] [Green Version]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Chu, D.M.; Meyer, K.M.; Prince, A.L.; Aagaard, K.M. Impact of maternal nutrition in pregnancy and lactation on offspring gut microbial composition and function. Gut Microbes 2016, 7, 459–470. [Google Scholar] [CrossRef] [Green Version]
- Matamoros, S.; Guen, C.G.-L.; Le Vacon, F.; Potel, G.; de La Cochetiere, M.-F. Development of intestinal microbiota in infants and its impact on health. Trends Microbiol. 2013, 21, 167–173. [Google Scholar] [CrossRef]
- E Beaudin, A.; Abarinov, E.V.; Malysheva, O.; A Perry, C.; Caudill, M.; Stover, P.J. Dietary folate, but not choline, modifies neural tube defect risk in Shmt1 knockout mice. Am. J. Clin. Nutr. 2011, 95, 109–114. [Google Scholar] [CrossRef]
- Reeves, P.G. Components of the AIN-93 Diets as Improvements in the AIN-76A Diet. J. Nutr. 1997, 127, 838S–841S. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.E.; Benninghoff, A.D.; Healy, B.J.; Li, M.; Vagu, B.; Hintze, K.J. Consumption of the total Western diet differentially affects the response to green tea in rodent models of chronic disease compared to the AIN93G diet. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.E.; Aardema, N.D.J.; Bunnell, M.L.; Larson, D.P.; Aguilar, S.S.; Bergeson, J.R.; Malysheva, O.V.; Caudill, M.A.; Lefevre, M. Effect of Choline Forms and Gut Microbiota Composition on Trimethylamine-N-Oxide Response in Healthy Men. Nutrients 2020, 12, 2220. [Google Scholar] [CrossRef]
- Illumina. 16S Metagenomic Sequencing Library Preparation. 2013. Available online: https://support.illumina.com/content/dam/illumina-support/documents/documentation/chemistry_documentation/16s/16s-metagenomic-library-prep-guide-15044223-b.pdf (accessed on 14 September 2020).
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a Dual-Index Sequencing Strategy and Curation Pipeline for Analyzing Amplicon Sequence Data on the MiSeq Illumina Sequencing Platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozupone, C.; Knight, R. UniFrac: A New Phylogenetic Method for Comparing Microbial Communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vázquez-Baeza, Y.; Pirrung, M.; Gonzalez, A.; Knight, R. EMPeror: A tool for visualizing high-throughput microbial community data. GigaScience 2013, 2, 2047-217X-2-16. [Google Scholar] [CrossRef] [Green Version]
- Morton, J.T.; Marotz, C.; Washburne, A.; Silverman, J.; Zaramela, L.S.; Edlund, A.; Zengler, K.; Knight, R. Establishing microbial composition measurement standards with reference frames. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Fedarko, M.W.; Martino, C.; Morton, J.T.; González, A.; Rahman, G.; A Marotz, C.; Minich, J.J.; E Allen, E.; Knight, R. Visualizing ’omic feature rankings and log-ratios using Qurro. NAR Genom. Bioinform. 2020, 2, lqaa023. [Google Scholar] [CrossRef]
- Szeto, I.M.Y.; Aziz, A.; Das, P.J.; Taha, A.Y.; Okubo, N.; Reza-Lopez, S.; Giacca, A.; Anderson, G.H. High multivitamin intake by Wistar rats during pregnancy results in increased food intake and components of the metabolic syndrome in male offspring. Am. J. Physiol. Integr. Comp. Physiol. 2008, 295, R575–R582. [Google Scholar] [CrossRef]
- Cho, C.E.; Sánchez-Hernández, D.; Reza-López, S.A.; Huot, P.S.; Kim, Y.-I.; Anderson, G.H. High folate gestational and post-weaning diets alter hypothalamic feeding pathways by DNA methylation in Wistar rat offspring. Epigenetics 2013, 8, 710–719. [Google Scholar] [CrossRef] [Green Version]
- Hammoud, R.; Pannia, E.; Kubant, R.; Wasek, B.; Bottiglieri, T.; Malysheva, O.V.; A Caudill, M.; Anderson, G.H. Choline and Folic Acid in Diets Consumed during Pregnancy Interact to Program Food Intake and Metabolic Regulation of Male Wistar Rat Offspring. J. Nutr. 2021, 151, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Savva, C.; Helguero, L.A.; González-Granillo, M.; Couto, D.; Melo, T.; Li, X.; Angelin, B.; Domingues, M.R.; Kutter, C.; Korach-André, M. Obese mother offspring have hepatic lipidic modulation that contributes to sex-dependent metabolic adaptation laterin life. Commun. Biol. 2021, 4, 14. [Google Scholar] [CrossRef]
- Bloor, I.D.; Sebert, S.; Saroha, V.; Gardner, D.S.; Keisler, D.; Budge, H.; Symonds, M.E.; Mahajan, R.P. Sex Differences in Metabolic and Adipose Tissue Responses to Juvenile-Onset Obesity in Sheep. Endocrinology 2013, 154, 3622–3631. [Google Scholar] [CrossRef] [Green Version]
- Andrich, D.E.; Melbouci, L.; Ou, Y.; Leduc-Gaudet, J.-P.; Chabot, F.; Lalonde, F.; Lira, F.S.; Gaylinn, B.D.; Gouspillou, G.; Danialou, G.; et al. Altered Feeding Behaviors and Adiposity Precede Observable Weight Gain in Young Rats Submitted to a Short-Term High-Fat Diet. J. Nutr. Metab. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hammoud, R.; Pannia, E.; Kubant, R.; Liao, C.-S.; Ho, M.; Yang, N.V.; Chatterjee, D.; Caudill, M.A.; Malysheva, O.V.; Pausova, Z.; et al. Maternal Choline Intake Programs Hypothalamic Energy Regulation and Later-Life Phenotype of Male Wistar Rat Offspring. Mol. Nutr. Food Res. 2020, 64, e1901178. [Google Scholar] [CrossRef]
- Coppola, S.; Avagliano, C.; Calignano, A.; Canani, R.B. The Protective Role of Butyrate against Obesity and Obesity-Related Diseases. Molecules 2021, 26, 682. [Google Scholar] [CrossRef] [PubMed]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S.; et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 2014, 5, 3611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, R.J.; Peng, L.; Barry, N.A.; Cline, G.W.; Zhang, D.; Cardone, R.L.; Petersen, K.F.; Kibbey, R.G.; Goodman, A.L.; Shulman, G.I. Acetate mediates a microbiome–brain–β-cell axis to promote metabolic syndrome. Nature 2016, 534, 213–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Rubaye, H.; Adamson, C.C.; Jadavji, N.M. The role of maternal diet on offspring gut microbiota development: A review. J. Neurosci. Res. 2020, 99, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Schaible, T.D.; Harris, R.; Dowd, S.; Smith, C.W.; Kellermayer, R. Maternal methyl-donor supplementation induces prolonged murine offspring colitis susceptibility in association with mucosal epigenetic and microbiomic changes. Hum. Mol. Genet. 2011, 20, 1687–1696. [Google Scholar] [CrossRef] [Green Version]
- Pannia, E.; Cho, C.E.; Kubant, R.; Sánchez-Hernández, D.; Huot, P.S.; Chatterjee, D.; Fleming, A.; Anderson, G.H. A high multivitamin diet fed to Wistar rat dams during pregnancy increases maternal weight gain later in life and alters homeostatic, hedonic and peripheral regulatory systems of energy balance. Behav. Brain Res. 2014, 278, 1–11. [Google Scholar] [CrossRef]
- Hiippala, K.; Barreto, G.; Burrello, C.; Diaz-Basabe, A.; Suutarinen, M.; Kainulainen, V.; Bowers, J.R.; Lemmer, D.; Engelthaler, D.M.; Eklund, K.K.; et al. Novel Odoribacter splanchnicus Strain and Its Outer Membrane Vesicles Exert Immunoregulatory Effects in vitro. Front. Microbiol. 2020, 11, 575455. [Google Scholar] [CrossRef]
- Lim, M.Y.; You, H.J.; Yoon, H.S.; Kwon, B.; Lee, J.Y.; Lee, S.; Song, Y.-M.; Lee, K.; Sung, J.; Ko, G. The effect of heritability and host genetics on the gut microbiota and metabolic syndrome. Gut 2017, 66, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Dao, M.C.; Everard, A.; Aron-Wisnewsky, J.; Sokolovska, N.; Prifti, E.; Verger, E.O.; Kayser, B.D.; Levenez, F.; Chilloux, J.; Hoyles, L.; et al. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: Relationship with gut microbiome richness and ecology. Gut 2016, 65, 426–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benítez-Páez, A.; del Pugar, E.M.G.; López-Almela, I.; Moya-Pérez, A.; Codoñer-Franch, P.; Sanz, Y. Depletion of Blautia Species in the Microbiota of Obese Children Relates to Intestinal Inflammation and Metabolic Phenotype Worsening. mSystems 2020, 5. [Google Scholar] [CrossRef] [Green Version]
- Peters, B.A.; Shapiro, J.A.; Church, T.R.; Miller, G.; Trinh-Shevrin, C.; Yuen, E.; Friedlander, C.; Hayes, R.B.; Ahn, J. A taxonomic signature of obesity in a large study of American adults. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Cerdó, T.; García-Santos, J.A.; Bermúdez, M.G.; Campoy, C. The Role of Probiotics and Prebiotics in the Prevention and Treatment of Obesity. Nutrients 2019, 11, 635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravussin, Y.; Koren, O.; Spor, A.; LeDuc, C.; Gutman, R.; Stombaugh, J.; Knight, R.; Ley, R.E.; Leibel, R.L. Responses of Gut Microbiota to Diet Composition and Weight Loss in Lean and Obese Mice. Obesity 2012, 20, 738–747. [Google Scholar] [CrossRef]
- Li, Q.; Liu, F.; Liu, J.; Liao, S.; Zou, Y. Mulberry Leaf Polyphenols and Fiber Induce Synergistic Antiobesity and Display a Modulation Effect on Gut Microbiota and Metabolites. Nutrients 2019, 11, 1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, H.A.; Bomhof, M.R.; Vogel, H.J.; Reimer, R.A. Diet-induced changes in maternal gut microbiota and metabolomic profiles influence programming of offspring obesity risk in rats. Sci. Rep. 2016, 6, 20683. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RV (D10012G) | HV (D18072701) | HFol (D18072702) | HFol-C (D18072703) | |
|---|---|---|---|---|
| g% diet (kcal%) | ||||
| Protein | 20 (20) | 20 (20) | 20 (20) | 20 (20) |
| Carbohydrate | 64 (64) | 64 (64) | 64 (64) | 64 (64) |
| Fat | 7 (16) | 7 (16) | 7 (16) | 7 (16) |
| Ingredient | g/kg diet (kcal/kg diet) | |||
| Casein | 200 (800) | 200 (800) | 200 (800) | 200 (800) |
| L-cystine | 3 (12) | 3 (12) | 3 (12) | 3 (12) |
| Cornstarch | 397 (1590) | 397 (1590) | 397 (1590) | 397 (1590) |
| Maltodextrin 10 | 132 (528) | 132 (528) | 132 (528) | 132 (528) |
| Sucrose | 100 (400) | 10 (40) | 100 (400) | 100 (400) |
| Sucrose from vitamin mix | 9.75 | 97.5 | 9.75 | 9.75 |
| Cellulose, BW200 | 50 (0) | 50 (0) | 50 (0) | 50 (0) |
| Soybean oil | 70 (630) | 70 (630) | 70 (630) | 70 (630) |
| Tert-butylhydroquinone | 0.014 (0) | 0.014 (0) | 0.014 (0) | 0.014 (0) |
| Mineral mix S10022G | 35 (0) | 35 (0) | 35 (0) | 35 (0) |
| Vitamin mix V10037 | 10 (40) | 100 (400) | 10 (40) | 10 (40) |
| Folic acid from vitamin mix | 0.002 (0) | 0.02 (0) | 0.002 (0) | 0.002 (0) |
| Folic acid added | 0.018 (0) | 0.018 (0) | ||
| Choline bitartrate | 2.5 (0) | 2.5 (0) | 2.5 (0) | 0 (0) |
| Total | 1000 (4000) | 1000 (4000) | 1000 (4000) | 998 (4000) |
| (µmol/g) | Acetic Acid | Propionic Acid | Isobutyric Acid | Butyric Acid | Isovaleric Acid | Valeric Acid | Caproic Acid |
|---|---|---|---|---|---|---|---|
| A Male offspring | |||||||
| RV | 4.91 ± 0.40 | 0.78 ± 0.04 | 0.13 ± 0.01 | 0.82 ± 0.08 a | 0.12 ± 0.01 | 0.13 ± 0.01 | 0.09 ± 0.01 |
| HV | 5.05 ± 0.39 | 0.93 ± 0.07 | 0.15 ± 0.01 | 0.86 ± 0.07 a | 0.13 ± 0.01 | 0.15 ± 0.01 | 0.08 ± 0.00 |
| HFol | 4.25 ± 0.24 | 0.85 ± 0.03 | 0.13 ± 0.01 | 0.58 ± 0.03 b | 0.12 ± 0.01 | 0.13 ± 0.00 | 0.08 ± 0.00 |
| HFol-C | 4.17 ± 0.35 | 0.76 ± 0.06 | 0.13 ± 0.01 | 0.49 ± 0.04 b | 0.12 ± 0.01 | 0.13 ± 0.01 | 0.08 ± 0.00 |
| p value | NS | NS | NS | p = 0.0004 | NS | NS | NS |
| B Female offspring | |||||||
| RV | 4.21 ± 0.25 a | 0.74 ± 0.03 | 0.13 ± 0.01 | 0.69 ± 0.08 | 0.11 ± 0.01 | 0.13 ± 0.01 | 0.07 ± 0.00 |
| HV | 5.50 ± 0.32 b | 0.81 ± 0.04 | 0.13 ± 0.00 | 0.78 ± 0.09 | 0.11 ± 0.01 | 0.13 ± 0.00 | 0.08 ± 0.00 |
| HFol | 3.73 ± 0.16 a | 0.77 ± 0.07 | 0.11 ± 0.01 | 0.54 ± 0.06 | 0.10 ± 0.01 | 0.11 ± 0.01 | 0.07 ± 0.00 |
| HFol-C | 5.47 ± 0.19 b | 0.82 ± 0.06 | 0.13 ± 0.01 | 0.70 ± 0.06 | 0.11 ± 0.01 | 0.13 ± 0.01 | 0.07 ± 0.00 |
| p value | p < 0.0001 | NS | NS | NS | NS | NS | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mjaaseth, U.N.; Norris, J.C.; Aardema, N.D.J.; Bunnell, M.L.; Ward, R.E.; Hintze, K.J.; Cho, C.E. Excess Vitamins or Imbalance of Folic Acid and Choline in the Gestational Diet Alter the Gut Microbiota and Obesogenic Effects in Wistar Rat Offspring. Nutrients 2021, 13, 4510. https://doi.org/10.3390/nu13124510
Mjaaseth UN, Norris JC, Aardema NDJ, Bunnell ML, Ward RE, Hintze KJ, Cho CE. Excess Vitamins or Imbalance of Folic Acid and Choline in the Gestational Diet Alter the Gut Microbiota and Obesogenic Effects in Wistar Rat Offspring. Nutrients. 2021; 13(12):4510. https://doi.org/10.3390/nu13124510
Chicago/Turabian StyleMjaaseth, Ulrik N., Jackson C. Norris, Niklas D. J. Aardema, Madison L. Bunnell, Robert E. Ward, Korry J. Hintze, and Clara E. Cho. 2021. "Excess Vitamins or Imbalance of Folic Acid and Choline in the Gestational Diet Alter the Gut Microbiota and Obesogenic Effects in Wistar Rat Offspring" Nutrients 13, no. 12: 4510. https://doi.org/10.3390/nu13124510
APA StyleMjaaseth, U. N., Norris, J. C., Aardema, N. D. J., Bunnell, M. L., Ward, R. E., Hintze, K. J., & Cho, C. E. (2021). Excess Vitamins or Imbalance of Folic Acid and Choline in the Gestational Diet Alter the Gut Microbiota and Obesogenic Effects in Wistar Rat Offspring. Nutrients, 13(12), 4510. https://doi.org/10.3390/nu13124510