
The Role of Fetal, Infant, and Childhood Nutrition in the Timing of Sexual Maturation

 ,
,  ,
,  ,
,  ,
,  and
and 
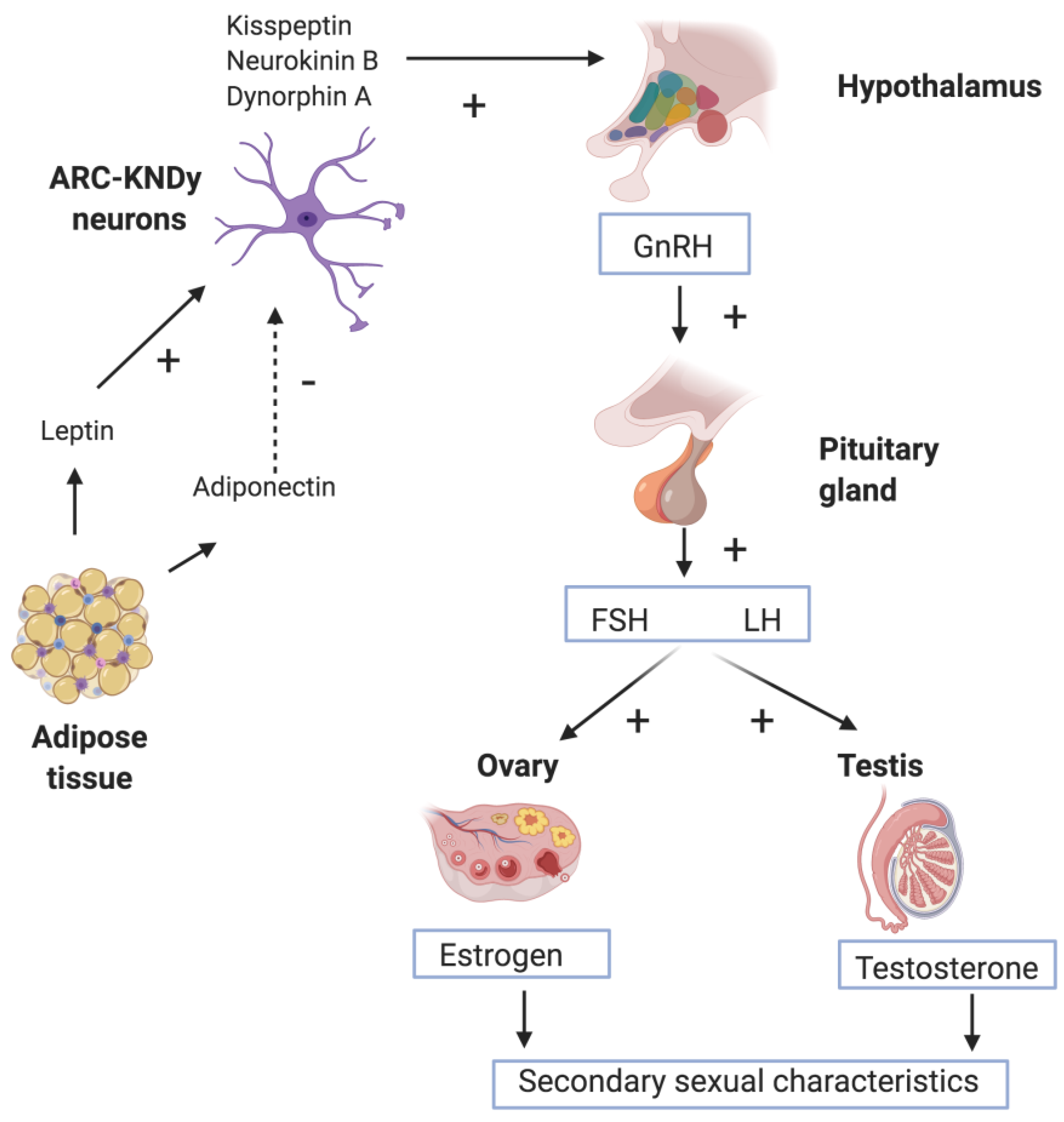{kind=link}
Abstract
:1. Introduction
2. Methods
3. Physiology of Puberty
4. Fetal Growth and Timing of Puberty
4.1. Small for Gestational Age and Pubertal Timing
4.2. IUGR Born SGA or AGA and Timing of Puberty
5. Neonatal and Early Infancy Nutrition and Puberty
6. Nutrient Intake during Childhood and Timing of Puberty
6.1. Macronutrient Intake
6.2. Micronutrient Intake
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Clayton, P.E.; Cianfarani, S.; Czernichow, P.; Johannsson, G.; Rapaport, R.; Rogol, A. Management of the child born small for gestational age through to adulthood: A consensus statement of the International Societies of Pediatric Endocrinology and the Growth Hormone Research Society. J. Clin. Endocrinol. Metab. 2007, 92, 804–810. [Google Scholar] [CrossRef] [Green Version]
- Abreu, A.P.; Kaiser, U.B. Pubertal development and regulation. Lancet Diabetes Endocrinol. 2016, 4, 254–264. [Google Scholar] [CrossRef] [Green Version]
- Barker, D.J.; Clark, P.M. Fetal undernutrition and disease in later life. Rev. Reprod. 1997, 2, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.W.; Ehrhardt, R.A. Regulation of placental nutrient transport and implications for fetal growth. Nutr. Res. Rev. 2002, 15, 211–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Pond, W.G.; Flynn, S.P.; Ott, T.L.; Bazer, F.W. Maternal dietary protein deficiency decreases nitric oxide synthase and ornithine decarboxylase activities in placenta and endometrium of pigs during early gestation. J. Nutr. 1998, 128, 2395–2402. [Google Scholar] [CrossRef] [PubMed]
- Sugden, M.C.; Holness, M.J. Gender-specific programming of insulin secretion and action. J. Endocrinol. 2002, 175, 757–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uenoyama, Y.; Inoue, N.; Nakamura, S.; Tsukamura, H. Central Mechanism Controlling Pubertal Onset in Mammals: A Triggering Role of Kisspeptin. Front. Endocrinol. 2019, 10, 312. [Google Scholar] [CrossRef] [Green Version]
- Kale, A.; Deardorff, J.; Lahiff, M.; Laurent, C.; Greenspan, L.C.; Hiatt, R.A.; Windham, G.; Galvez, M.P.; Biro, F.M.; Pinney, S.M.; et al. Breastfeeding versus formula-feeding and girls’ pubertal development. Matern. Child Health J. 2015, 19, 519–527. [Google Scholar] [CrossRef] [Green Version]
- Waterland, R.A.; Jirtle, R.L. Early nutrition, epigenetic changes at transposons and imprinted genes, and enhanced susceptibility to adult chronic diseases. Nutrition 2004, 20, 63–68. [Google Scholar] [CrossRef]
- Farello, G.; Altieri, C.; Cutini, M.; Pozzobon, G.; Verrotti, A. Review of the Literature on Current Changes in the Timing of Pubertal Development and the Incomplete Forms of Early Puberty. Front. Pediatr. 2019, 7, 147. [Google Scholar] [CrossRef] [Green Version]
- Marshall, W.A.; Tanner, J.M. Variations in pattern of pubertal changes in girls. Arch. Dis. Child. 1969, 44, 291–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaltiala-Heino, R.; Marttunen, M.; Rantanen, P.; Rimpelä, M. Early puberty is associated with mental health problems in middle adolescence. Soc. Sci. Med. 2003, 57, 1055–1064. [Google Scholar] [CrossRef]
- Copeland, W.; Shanahan, L.; Miller, S.; Costello, E.J.; Angold, A.; Maughan, B. Outcomes of early pubertal timing in young women: A prospective population-based study. Am. J. Psychiatry. 2010, 167, 1218–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klopack, E.T.; Sutton, T.E.; Simons, R.L.; Simons, L.G. Disentangling the Effects of Boys’ Pubertal Timing: The Importance of Social Context. J. Youth Adolesc. 2020, 49, 1393–1405. [Google Scholar] [CrossRef] [PubMed]
- Carel, J.C.; Lahlou, N.; Roger, M.; Chaussain, J.L. Precocious puberty and statural growth. Hum. Reprod. Update 2014, 10, 135–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasa, T.; Matsuzaki, T.; Murakami, M.; Fujisawa, S.; Kinouchi, R.; Gereltsetseg, G.; Kuwahara, A.; Yasui, T.; Irahara, M. Effects of intrauterine undernutrition on hypothalamic Kiss1 expression and the timing of puberty in female rats. J. Physiol. 2010, 588, 821–829. [Google Scholar] [CrossRef]
- Ojeda, S.R.; Dubay, C.; Lomniczi, A.; Kaidar, G.; Matagne, V.; Sandau, U.S.; Dissen, G.A. Gene networks and the neuroendocrine regulation of puberty. Mol. Cell Endocrinol. 2010, 324, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Connor, N.E. Impact of fetal and neonatal malnutrition on the onset of puberty and associated noncommunicable disease risks. Adolesc. Health Med. Ther. 2011, 2, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Matsui, H.; Takatsu, Y.; Kumano, S.; Matsumoto, H.; Ohtaki, T. Peripheral administration of metastin induces marked gonadotropin release and ovulation in the rat. Biochem. Biophys. Res. Commun. 2004, 320, 383–388. [Google Scholar] [CrossRef]
- Messager, S.; Chatzidaki, E.E.; Ma, D.; Hendrick, A.G.; Zahn, D.; Dixon, J.; Thresher, R.R.; Malinge, I.; Lomet, D.; Carlton, M.B.; et al. Kisspeptin directly stimulates gonadotropin-releasing hormone release via G protein-coupled receptor 54. Proc. Natl. Acad. Sci. USA 2005, 102, 1761–1766. [Google Scholar] [CrossRef] [Green Version]
- Ohkura, S.; Takase, K.; Matsuyama, S.; Mogi, K.; Ichimaru, T.; Wakabayashi, Y.; Uenoyama, Y.; Mori, Y.; Steiner, R.A.; Tsukamura, H.; et al. Gonadotrophin-releasing hormone pulse generator activity in the hypothalamus of the goat. J. Neuroendocrinol. 2009, 21, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Naniwa, Y.; Nakatsukasa, K.; Setsuda, S.; Oishi, S.; Fujii, N.; Matsuda, F.; Uenoyama, Y.; Tsukamura, H.; Maeda, K.; Ohkura, S. Effects of full-length kisspeptin administration on follicular development in Japanese Black beef cows. J. Reprod. Dev. 2013, 59, 588–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahab, M.; Mastronardi, C.; Seminara, S.B.; Crowley, W.F.; Ojeda, S.R.; Plant, T.M. Increased hypothalamic GPR54 signaling: A potential mechanism for initiation of puberty in primates. Proc. Natl. Acad. Sci. USA 2005, 102, 2129–2134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funes, S.; Hedrick, J.A.; Vassileva, G.; Markowitz, L.; Abbondanzo, S.; Golovko, A.; Yang, S.; Monsma, F.J.; Gustafson, E.L. The KiSS-1 receptor GPR54 is essential for the development of the murine reproductive system. Biochem. Biophys. Res. Commun. 2003, 312, 1357–1363. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, D.M.; Blache, D.; Hoggard, N.; Brookes, E.; Wooding, F.B.; Fowden, A.L.; Forhead, A.J. Developmental control of plasma leptin and adipose leptin messenger ribonucleic acid in the ovine fetus during late gestation: Role of glucocorticoids and thyroid hormones. Endocrinology 2007, 148, 3750–3757. [Google Scholar] [CrossRef] [Green Version]
- Quinton, N.D.; Smith, R.F.; Clayton, P.E.; Gill, M.S.; Shalet, S.; Justice, S.K.; Simon, S.A.; Walters, S.; Postel-Vinay, M.C.; Blakemore, A.I.; et al. Leptin binding activity changes with age: The link between leptin and puberty. J. Clin. Endocrinol. Metab. 1999, 84, 2336–2341. [Google Scholar] [CrossRef]
- Soliman, A.; De Sanctis, V.; Elalaily, R. Nutrition and pubertal development. Indian J. Endocrinol. Metab. 2014, 18 (Suppl. S1), S39–S47. [Google Scholar] [CrossRef]
- Mantzoros, C.S.; Flier, J.S.; Rogol, A.D. A longitudinal assessment of hormonal and physical alterations during normal puberty in boys. V. Rising leptin levels may signal the onset of puberty. J. Clin. Endocrinol. Metab. 1997, 82, 1066–1070. [Google Scholar] [CrossRef]
- Gruszfeld, D.; Kułaga, Z.; Wierzbicka, A.; Rzehak, P.; Grote, V.; Martin, F.; Poncelet, P.; Closa-Monasterolo, P.; Escribano, J.; Verduci, E.; et al. EU Childhood Obesity Study Group. Leptin and Adiponectin Serum Levels from Infancy to School Age: Factors Influencing Tracking. Child. Obes. 2016, 12, 179–187. [Google Scholar] [CrossRef]
- Fandiño, J.; Toba, L.; González-Matías, L.C.; Diz-Chaves, Y.; Mallo, F. Perinatal Undernutrition, Metabolic Hormones, and Lung Development. Nutrients 2019, 11, 2870. [Google Scholar] [CrossRef] [Green Version]
- Alkalay, A.L.; Graham, J.M., Jr.; Pomerance, J.J. Evaluation of neonates born with intrauterine growth retardation: Review and practice guidelines. J. Perinatol. 1998, 18, 142–151. [Google Scholar] [PubMed]
- Schlaudecker, E.P.; Munoz, F.M.; Bardají, A.; Boghossian, N.S.; Khalil, A.; Mousa, H.; Nesin, M.; Nisar, M.I.; Pool, V.; Spiegel, H.M.L.; et al. For The Brighton Collaboration Small for Gestational Age Working Group. Small for gestational age: Case definition & guidelines for data collection, analysis, and presentation of maternal immunization safety data. Vaccine 2017, 5, 6518–6528. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Sánchez, B.N.; Goodrich, J.M.; Dolinoy, D.C.; Cantoral, A.; Mercado-Garcia, A.; Ruiz-Narváez, E.A.; Téllez-Rojo, M.M.; Peterson, K.E. Dietary exposures, epigenetics and pubertal tempo. Environ. Epigenet. 2019, 5, dvz002. [Google Scholar] [CrossRef] [PubMed]
- Cutland, C.L.; Lackritz, E.; Alonso, A.B.; Bodjick, T.; Chandrasekaran, R.; Lahariya, C.; Nisar, M.I.; Tapia, M.D.; Pathirana, J.; Kochhar, S.; et al. Low birth weight: Case definition & guidelines for data collection, analysis, and presentation of maternal immunisation safety data. Vaccine 2016, 35, 6492. [Google Scholar]
- Tam, C.S.; de Zegher, F.; Garnett, S.P.; Baur, L.A.; Cowell, C.T. Opposing influences of prenatal and postnatal growth on the timing of menarche. J. Clin. Endocrinol. Metab. 2006, 91, 4369–4373. [Google Scholar] [CrossRef]
- Wang, Y.; Dinse, G.E.; Rogan, W.J. Birth weight, early weight gain and pubertal maturation: A longitudinal study. Pediatr. Obes. 2012, 7, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Koziel, S.; Jankowska, E.A. Effect of low versus normal birthweight on menarche in 14-year-old Polish girls. J. Paediatr. Child Health 2002, 38, 268–271. [Google Scholar] [CrossRef]
- Adair, L.S. Size at birth predicts age at menarche. Pediatrics 2001, 107, E59. [Google Scholar] [CrossRef] [Green Version]
- Sloboda, D.M.; Hart, R.; Doherty, D.A.; Pennell, C.E.; Hickey, M. Age at menarche: Influences of prenatal and postnatal growth. Int. J. Clin. Endocrinol. Metab. 2007, 92, 46–50. [Google Scholar] [CrossRef]
- Terry, M.B.; Ferris, J.S.; Tehranifar, P.; Wei, Y.; Flom, J.D. Birth weight, postnatal growth, and age at menarche. Am. J. Epidemiol. 2009, 170, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Epplein, M.; Novotny, R.; Daida, Y.; Vijayadeva, V.; Onaka, A.T.; Le Marchand, L. Association of maternal and intrauterine characteristics with ageatmenarche in a multiethnicpopulation in Hawaii. Cancer Cause Control. 2010, 21, 259–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos Silva, I.; De Stavola, B.L.; Mann, V.; Kuh, D.; Hardy, R.; Wadsworth, M.E.J. Prenatal factors, childhoodgrowthtrajectories and ageatmenarche. Int. J. Epidemiol. 2002, 31, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Maisonet, M.; Christensen, K.Y.; Rubin, C.; Holmes, A.; Flanders, W.D.; Heron, J.; Ong, K.K.; Golding, J.; McGeehin, M.A.; Marcus, M. Role of prenatal characteristics and early growth on pubertal attainment of British girls. Pediatrics 2010, 126, e591–e600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dossus, L.; Kvaskoff, M.; Bijon, A.; Fervers, B.; Boutron-Ruault, M.C.; Mesrine, S.; Clavel-Chapelon, F. Determinants of age at menarche and time to menstrual cycle regularity in the French E3N cohort. Ann. Epidemiol. 2012, 22, 723–730. [Google Scholar] [CrossRef] [PubMed]
- D’Aloisio, A.A.; DeRoo, L.A.; Baird, D.D.; Weinberg, C.R.; Sandler, D.P. Prenatal and infant exposures and age at menarche. Epidemiology 2013, 24, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Raaijmakers, A.; Jacobs, L.; Rayyan, M.; Van Tienoven, T.P.; Ortibus, E.; Levtchenko, E.; Staessen, J.A.; Allegaert, K. Catch-up growth in the first two years of life in Extremely Low Birth Weight (ELBW) infants is associated with lower body fat in young adolescence. PLoS ONE 2017, 12, e0173349. [Google Scholar] [CrossRef] [PubMed]
- Hvidt, J.J.; Brix, N.; Ernst, A.; Lauridsen, L.L.B.; Ramlau-Hansen, C.H. Size at birth, infant growth, and age at pubertal development in boys and girls. Clin. Epidemiol. 2019, 11, 873–883. [Google Scholar] [CrossRef] [Green Version]
- Persson, I.; Ahlsson, F.; Ewald, U.; Tuvemo, T.; Qingyuan, M.; von Rosen, D.; Proos, L. Influence of perinatal factors on the onset of puberty in boys and girls: Implications for interpretation of link with risk of long term diseases. Am. J. Epidemiol. 1999, 150, 747–755. [Google Scholar] [CrossRef]
- Lazar, L.; Pollak, U.; Kalter-Leibovici, O.; Pertzelan, A.; Phillip, M. Pubertal course of persistently short children born small for gestational age (SGA) compared with idiopathic short children born appropriate for gestational age (AGA). Eur. J. Endocrinol. 2003, 149, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Bhargava, S.K.; Ramji, S.; Srivastava, U.; Sachdev, H.P.; Kapani, V.; Datta, V.; Satyanarayana, L. Growth and sexual maturation of low birth weight children: A 14 year follow up. Indian Pediatr. 1995, 32, 963–970. [Google Scholar]
- Adair, L.S. Size at birth and growth trajectories to young adulthood. Am. J. Hum. Biol. 2007, 19, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Ilbertsson-Wikland, K.; Karlberg, J. Natural growth in children born small for gestational age with and without catch-up growth. Acta Paediatr. 1994, 399, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Karlberg, J.; Kwan, C.W.; Gelander, L.; Albertsson-Wikland, K. Pubertal growth assessment. Horm. Res. 2003, 60 (Suppl. S1), 27–35. [Google Scholar] [CrossRef]
- Ibáñez, L.; Jiménez, R.; de Zegher, F. Early Puberty-Menarche after Precocious Pubarche: Relation to Prenatal Growth. Pediatrics 2006, 117, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Neville, K.A.; Walker, J.L. Precocious pubarche is associated with SGA, prematurity, weight gain, and obesity. Arch. Dis. Child. 2005, 90, 258–261. [Google Scholar] [CrossRef] [Green Version]
- Cooper, C.; Kuh, D.; Egger, P.; Wadsworth, M.; Barker, D. Childhood growth and age at menarche. Br. J. Obs. Gynaecol. 1996, 103, 814–817. [Google Scholar] [CrossRef]
- Blell, M.; Pollard, T.M.; Pearce, M.S. Predictors of age at menarche in the new castle thousand families study. J. Biosoc. Sci. 2008, 40, 563–575. [Google Scholar] [CrossRef] [Green Version]
- Behie, A.M.; O’Donnell, M.H. Prenatal smoking and ageatmenarche: Influence of the prenatal environment on the timing of puberty. Hum Reprod. 2015, 30, 957–962. [Google Scholar] [CrossRef] [Green Version]
- Morris, D.H.; Jones, M.E.; Schoemaker, M.J.; Ashworth, A.; Swerdlow, A.J. Determinants of age at menarche in the UK: Analyses from the break through generations study. Br. J. Cancer 2010, 103, 1760–1764. [Google Scholar] [CrossRef] [Green Version]
- Ong, K.K.; Potau, N.; Petry, C.J.; Jones, R.; Ness, A.R.; Honour, J.W.; de Zegher, F.; Ibáñez, L.; Dunger, D.B. Avon Longitudinal Study of Parents and Children Study Team. Opposing influences of prenatal and postnatal weight gain on adrenarche in normal boys and girls. J. Clin. Endocrinol. Metab. 2004, 89, 2647–2651. [Google Scholar] [CrossRef] [Green Version]
- Ibáñez, L.; Potau, N.; Zampolli, M.; Riqué, S.; Saenger, P.; Carrascosa, A. Hyperinsulinemia and decreased insulin-like growth factor-binding protein-1 are common features in prepubertal and pubertal girls with a history of premature pubarche. J. Clin. Endocrinol. Metab. 1997, 82, 2283–2288. [Google Scholar] [CrossRef] [PubMed]
- Hernández, M.I.; Mericq, V. Impact of being born small for gestational age on onset and progression of puberty. Best Pract. Res. Clin. Endocrinol. Metab. 2008, 22, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Francois, I.; de Zegher, F.; Spiessens, C.; D’Hooghe, T.; Vanderschueren, D. Low birth weight and subsequent male subfertility. Pediatr. Res. 1997, 42, 899–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nardozza, L.M.; Caetano, A.C.; Zamarian, A.C.; Mazzola, J.B.; Silva, C.P.; Marçal, V.M.; Lobo, T.F.; Peixoto, A.B.; Araujo Júnior, E. Fetal growth restriction: Current knowledge. Arch. Gynecol. Obstet. 2017, 295, 1061–1077. [Google Scholar] [CrossRef] [PubMed]
- Saenger, P.; Czernichow, P.; Hughes, I.; Reiter, E.O. Small for gestational age: Short stature and beyond. Endocr. Rev. 2007, 28, 219–251. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Sharma, P.; Shastri, S. Genetic, metabolic and endocrine aspect of intrauterine growth restriction: An update. J. Matern. Fetal Neonatal Med. 2017, 30, 2263–2275. [Google Scholar] [CrossRef]
- Barker, D.J.; Eriksson, J.G.; Forsén, T.; Osmond, C. Fetal origins of adult disease: Strength of effects and biological basis. Int. J. Epidemiol. 2002, 31, 1235–1239. [Google Scholar] [CrossRef] [Green Version]
- Hanson, M.A.; Gluckman, P.D. Early developmental conditioning of later health and disease: Physiology or pathophysiology? Physiol. Rev. 2014, 94, 1027–1076. [Google Scholar] [CrossRef]
- Sagiv, S.K.; Tolbert, P.E.; Altshul, L.M.; Korrick, S.A. Organochlorine exposures during pregnancy and infant size at birth. Epidemiology 2007, 18, 120–129. [Google Scholar] [CrossRef]
- Sancakli, O.; Darendeliler, F.; Bas, F.; Gokcay, G.; Disci, R.; Aki, S.; Eskiyurt, N. Insulin, adiponectin, IGFBP-1 levels and body composition in small for gestational age born non-obese children during prepubertal ages. Clin. Endocrinol. 2008, 69, 88–92. [Google Scholar] [CrossRef]
- Ibáñez, L.; Ong, K.; Dunger, D.B.; de Zegher, F. Early development of adiposity and insulin resistance after catch-up weight gain in small-for-gestational-age children. J. Clin. Endocrinol. Metab. 2006, 91, 2153–2158. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, M.L.; Ong, K.K.; Dunger, D.B. Childhood obesity and the timing of puberty. Trends Endocrinol. Metab. 2009, 20, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Kota, S.K.; Gayatri, K.; Jammula, S.; Meher, L.K.; Kota, S.K.; Krishna, S.V.; Modi, K.D. Fetal endocrinology. Indian J. Endocrinol. Metab. 2013, 17, 568–579. [Google Scholar] [CrossRef] [PubMed]
- Engelbregt, M.J.; Houdijk, M.E.; Popp-Snijders, C.; Delemarre-van de Waal, H.A. The effects of intra-uterine growth retardation and postnatal undernutrition on onset of puberty in male and female rats. Pediatr. Res. 2000, 48, 803–807. [Google Scholar] [CrossRef] [PubMed]
- Engelbregt, M.J.; van Weissenbruch, M.M.; Popp-Snijders, C.; Delemarre-van de Waal, H.A. Delayed first cycle in intrauterine growth-retarded and postnatally undernourished female rats: Follicular growth and ovulation after stimulation with pregnant mare serum gonadotropin at first cycle. J. Endocrinol. 2002, 173, 297–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, R.; Sloboda, D.M.; Doherty, D.A.; Norman, R.J.; Atkinson, H.C.; Newnham, J.P.; Dickinson, J.E.; Hickey, M. Prenatal determinants of uterine volume and ovarian reserve in adolescence. J. Clin. Endocrinol. Metab. 2009, 94, 4931–4937. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez, L.; Potau, N.; Francois, I.; de Zegher, F. Precocious pubarche, hyperinsulinism, and ovarian hyperandrogenism in girls: Relation to reduced fetal growth. J. Clin. Endocrinol. Metab. 1998, 83, 3558–3562. [Google Scholar] [CrossRef]
- Hoek, A.; Schoemaker, J.; Drexhage, H.A. Premature ovarian failure and ovarian autoimmunity. Endocr. Rev. 1997, 18, 107–134. [Google Scholar] [CrossRef] [Green Version]
- Cebu Study Team. Underlying and Proximate Determinants of Child Health: The Cebu Longitudinal Health and Nutrition Study. Am. J. Epidemiol. 1991, 133, 185–201. [Google Scholar] [CrossRef]
- Da Silva, P.; Aitken, R.P.; Rhind, S.M.; Racey, P.A.; Wallace, J.M. Influence of placentally mediated fetal growth restriction on the onset of puberty in male and female lambs. Reproduction 2001, 122, 375–383. [Google Scholar] [CrossRef]
- Victora, C.G.; Horta, B.L.; Loret de Mola, C.; Quevedo, L.; Pinheiro, R.T.; Gigante, D.P.; Gonçalves, H.; Barros, F.C. Association between breastfeeding and intelligence, educational attainment, and income at 30 years of age: A prospective birth cohort study from Brazil. Lancet Glob. Health 2015, 3, e199–e205. [Google Scholar] [CrossRef] [Green Version]
- Owen, C.G.; Martin, R.M.; Whincup, P.H.; Smith, G.D.; Cook, D.G. Effect of infant feeding on the risk of obesity across the life course: A quantitative review of published evidence. Pediatrics 2005, 115, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Kaingade, P.; Somasundaram, I.; Nikam, A.; Behera, P.; Kulkarni, S.; Patel, J. Breast milk cell components and its beneficial effects on neonates: Need for breast milk cell banking. J. Pediatr. Neonatal Individ. Med. 2017, 6, e060115. [Google Scholar] [CrossRef]
- Kramer, M.S.; Kakuma, R. Optimal duration of exclusive breastfeeding. Cochrane Database Syst. Rev. 2012, 2012, CD003517. [Google Scholar] [CrossRef]
- World Health Organization. Global Strategy for Infant and Young Child Feeding. In The Optimal Duration of Exclusive Breastfeeding; WHO: Geneva, Switzerland, 2001. [Google Scholar]
- Karaolis-Danckert, N.; Buyken, A.E.; Sonntag, A.; Kroke, A. Birth and early life influences on the timing of puberty onset: Results from the DONALD (Dortmund Nutritional and Anthropometric Longitudinally Designed) Study. Am. J. Clin. Nutr. 2009, 90, 1559–1565. [Google Scholar] [CrossRef] [Green Version]
- Kwok, M.K.; Leung, G.M.; Lam, T.H.; Schooling, C.M. Breastfeeding, childhood milk consumption, and onset of puberty. Pediatrics 2012, 130, e631–e639. [Google Scholar] [CrossRef] [Green Version]
- Schooling, C.M.; Hui, L.L.; Ho, L.M.; Lam, T.H.; Leung, G.M. Cohort profile: ‘children of 1997′: A Hong Kong Chinese birth cohort. Int. J. Epidemiol. 2012, 41, 611–620. [Google Scholar] [CrossRef]
- Aghaee, S.; Deardorff, J.; Greenspan, L.C.; Quesenberry, C.P., Jr.; Kushi, L.H.; Kubo, A. Breastfeeding and timing of pubertal onset in girls: A multiethnic population-based prospective cohort study. BMC Pediatr. 2019, 19, 277. [Google Scholar] [CrossRef]
- Al-Sahab, B.; Adair, L.; Hamadeh, M.J.; Ardern, C.I.; Tamim, H. Impact of breastfeeding duration on age at menarche. Am. J. Epidemiol. 2011, 173, 971–977. [Google Scholar] [CrossRef] [Green Version]
- Kwok, M.K.; Schooling, C.M.; Lam, T.H.; Leung, G.M. Does breastfeeding protect against childhood overweight? Hong Kong’s ’Children of 1997’ birth cohort. Int. J. Epidemiol. 2010, 39, 297–305. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.A.; Kim, Y.J.; Lee, H.; Gwak, H.S.; Hong, Y.S.; Kim, H.S.; Park, E.A.; Cho, S.J.; Ha, E.H.; Park, H. The preventive effect of breast-feeding for longer than 6 months on early pubertal development among children aged 7–9 years in Korea. Public Health Nutr. 2015, 18, 3300–3307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maclure, M.; Travis, L.B.; Willett, W.; MacMahon, B. A prospective cohort study of nutrient intake and age at menarche. Am. J. Clin. Nutr. 1991, 54, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Vickers, M.H.; Krechowec, S.O.; Gluckman, P.D.; Breier, B.H.; Gertler, A. The Role of Leptin during Early Life in Imprinting Later Metabolic Responses. In Leptin Leptin Antagon; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Emmett, P.M.; Jones, L.R. Diet, growth, and obesity development throughout childhood in the Avon Longitudinal Study of Parents and Children. Nutr. Rev. 2015, 73 (Suppl. S3), 175–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moisan, J.; Meyer, F.; Gingras, S. A nested case-control study of the correlates of early menarche. Am. J. Epidemiol. 1990, 132, 953–961. [Google Scholar] [CrossRef] [PubMed]
- Lynch, C.; Kristjansdottir, A.G.; Te Velde, S.J.; Lien, N.; Roos, E.; Thorsdottir, I.; Krawinkel, M.; de Almeida, M.D.; Papadaki, A.; Hlastan Ribic, C.; et al. Fruit and vegetable consumption in a sample of 11-year-old children in ten European countries--the PRO GREENS cross-sectional survey. Public Health Nutr. 2014, 17, 2436–2444. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Lamb, K.E.; Grimes, C.; Laws, R.; Bolton, K.; Ong, K.K.; Campbell, K. Rapid weight gain during infancy and subsequent adiposity: A systematic review and meta-analysis of evidence. Obes. Rev. 2018, 19, 321–332. [Google Scholar] [CrossRef]
- Yan, J.; Liu, L.; Zhu, Y.; Huang, G.; Wang, P.P. The association between breastfeeding and childhood obesity: A meta-analysis. BMC Public Health 2014, 14, 1267. [Google Scholar] [CrossRef] [Green Version]
- Pluymen, L.P.M.; Wijga, A.H.; Gehring, U.; Koppelman, G.H.; Smit, H.A.; van Rossem, L. Early introduction of complementary foods and childhood overweight in breastfed and formula-fed infants in the Netherlands: The PIAMA birth cohort study. Eur. J. Nutr. 2018, 57, 1985–1993. [Google Scholar] [CrossRef] [Green Version]
- Smith-Brown, P.; Morrison, M.; Krause, L.; Davies, P.S.W. Microbiota and Body Composition During the Period of Complementary Feeding. J. Pediatr. Gastroenterol. Nutr. 2019, 69, 726–732. [Google Scholar] [CrossRef]
- Stewart, C.J.; Ajami, N.J.; O’Brien, J.L.; Hutchinson, D.S.; Smith, D.P.; Wong, M.C.; Ross, M.C.; Lloyd, R.E.; Doddapaneni, H.; Metcalf, G.A.; et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature 2018, 562, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.T.K.; Fan, H.Y.; Tsai, M.C.; Tung, T.H.; Huynh, Q.T.V.; Huang, S.Y.; Chen, Y.C. Nutrient Intake through Childhood and Early Menarche Onset in Girls: Systematic Review and Meta-Analysis. Nutrients 2020, 12, 2544. [Google Scholar] [CrossRef] [PubMed]
- Remer, T.; Shi, L.; Buyken, A.E.; Maser-Gluth, C.; HartmAnn, M.F.; Wudy, S.A. Prepubertal adrenarchal androgens and animal protein intake independently and differentially influence pubertal timing. J. Clin. Endocrinol. Metab. 2010, 95, 3002–3009. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.J. The adiposity rebound in the 21st century children: Meaning for what? Korean J. Pediatr. 2018, 61, 375–380. [Google Scholar] [CrossRef]
- Wiley, A.S. Milk intake and total dairy consumption: Associations with early menarche in NHANES 1999–2004. PLoS ONE 2011, 6, e14685. [Google Scholar] [CrossRef] [Green Version]
- Günther, A.L.; Karaolis-Danckert, N.; Kroke, A.; Remer, T.; Buyken, A.E. Dietary protein intake throughout childhood is associated with the timing of puberty. J. Nutr. 2010, 140, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Ramezani Tehrani, F.; Moslehi, N.; Asghari, G.; Gholami, R.; Mirmiran, P.; Azizi, F. Intake of dairy products, calcium, magnesium, and phosphorus in childhood and age at menarche in the Tehran Lipid and Glucose Study. PLoS ONE 2013, 8, e57696. [Google Scholar] [CrossRef] [PubMed]
- Boyd, A.; Golding, J.; Macleod, J.; Lawlor, D.A.; Fraser, A.; Henderson, J.; Molloy, L.; Ness, A.; Ring, S.; Davey Smith, G. Cohort Profile: The ‘children of the 90s’—The index offspring of the Avon Longitudinal Study of Parents and Children. Int. J. Epidemiol. 2013, 42, 111–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carwile, J.L.; Willett, W.C.; Wang, M.; Rich-Edwards, J.; Frazier, A.L.; Michels, K.B. Milk Consumption after Age 9 Years Does Not Predict Age at Menarche. J. Nutr. 2015, 145, 1900–1908. [Google Scholar] [CrossRef] [PubMed]
- Iannotti, L.L. The benefits of animal products for child nutrition in developing countries. Rev. Sci. Tech. 2018, 37, 37–46. [Google Scholar] [CrossRef]
- Givens, D.I. MILK Symposium review: The importance of milk and dairy foods in the diets of infants, adolescents, pregnant women, adults, and the elderly. J. Dairy Sci. 2020, 103, 9681–9699. [Google Scholar] [CrossRef] [PubMed]
- Carwile, J.L.; Willett, W.C.; Spiegelman, D.; Hertzmark, E.; Rich-Edwards, J.; Frazier, A.L.; Michels, K.B. Sugar-sweetened beverage consumption and age at menarche in a prospective study of US girls. Hum. Reprod. 2015, 30, 675–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, E.; Powell, L.M. Consumption patterns of sugar-sweetened beverages in the United States. J. Acad. Nutr. Diet 2013, 113, 43–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brand-Miller, J.C.; Barclay, A.W. Declining consumption of added sugars and sugar- sweetened beverages in Australia: A challenge for obesity prevention. Am. J. Clin. Nutr. 2017, 105, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Pimienta, T.G.; Batis, C.; Lutter, C.K.; Rivera, J.A. Sugar-Sweetened Beverages Are the Main Sources of Added Sugar Intake in the Mexican Population. J. Nutr. 2016, 146, 1888S–1896S. [Google Scholar] [CrossRef] [Green Version]
- Gui, Z.H.; Zhu, Y.N.; Cai, L.; Sun, F.H.; Ma, Y.H.; Jing, J.; Chen, Y.J. Sugar-Sweetened Beverage Consumption and Risks of Obesity and Hypertension in Chinese Children and Adolescents: A National Cross-Sectional Analysis. Nutrients 2017, 9, 1302. [Google Scholar] [CrossRef] [Green Version]
- Dereń, K.; Weghuber, D.; Caroli, M.; Koletzko, B.; Thivel, D.; Frelut, M.L.; Socha, P.; Grossman, Z.; Hadjipanayis, A.; Wyszyńska, J.; et al. Consumption of Sugar-Sweetened Beverages in Paediatric Age: A Position Paper of the European Academy of Paediatrics and the European Childhood Obesity Group. Ann. Nutr. Metab. 2019, 74, 296–302. [Google Scholar] [CrossRef]
- Merzenich, H.; Boeing, H.; Wahrendorf, J. Dietary fat and sports activity as determinants for age at menarche. Am. J. Epidemiol. 1993, 138, 217–224. [Google Scholar] [CrossRef]
- Berkey, C.S.; Gardner, J.D.; Frazier, A.L.; Colditz, G.A. Relation of childhood diet and body size to menarche and adolescent growth in girls. Am. J. Epidemiol. 2000, 152, 446–452. [Google Scholar] [CrossRef] [Green Version]
- Koprowski, C.; Ross, R.K.; Mack, W.J.; Henderson, B.E.; Bernstein, L. Diet, body size and menarche in a multiethnic cohort. Br. J. Cancer 1999, 79, 1907–1911. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.L.; Raubenheimer, D.; Steinbeck, K.; Baur, L.; Garnett, S. New insights into the association of mid-childhood macronutrient intake to pubertal development in adolescence using nutritional geometry. Br. J. Nutr. 2019, 122, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Olmedilla, B.; Granado, F. Growth and micronutrient needs of adolescents. Eur. J. Clin. Nutr. 2000, 54 (Suppl. S1), S11–S15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alimujiang, A.; Colditz, G.A.; Gardner, J.D.; Park, Y.; Berkey, C.S.; Sutcliffe, S. Childhood diet and growth in boys in relation to timing of puberty and adult height: The Longitudinal Studies of Child Health and Development. Cancer Causes Control 2018, 29, 915–926. [Google Scholar] [CrossRef] [PubMed]
- Frisch, R.E. The right weight: Body fat, menarche and fertility. Proc. Nutr. Soc. 1994, 53, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Odongkara Mpora, B.; Piloya, T.; Awor, S.; Ngwiri, T.; Laigong, P.; Mworozi, E.A.; Hochberg, Z. Age at menarche in relation to nutritional status and critical life events among rural and urban secondary school girls in post-conflict northern Uganda. BMC Womens Health 2014, 14, 66. [Google Scholar] [CrossRef] [Green Version]
- Villamor, E.; Marín, C.; Mora-Plazas, M.; Oliveros, H. Micronutrient status in middle childhood and age at menarche: Results from the Bogotá School Children Cohort. Br. J. Nutr. 2017, 118, 1097–1105. [Google Scholar] [CrossRef] [Green Version]
- Devi, S.; Pasanna, R.M.; Shamshuddin, Z.; Bhat, K.; Sivadas, A.; Mandal, A.K.; Kurpad, A.V. Measuring vitamin B-12 bioavailability with [13C]-cyanocobalamin in humans. Am. J. Clin. Nutr. 2020, 25, nqaa221. [Google Scholar] [CrossRef]
- Grundmann, M.; von Versen-Höynck, F. Vitamin D—Roles in women’s reproductive health? Reprod. Biol. Endocrinol. 2011, 9, 146. [Google Scholar] [CrossRef] [Green Version]
- Villamor, E.; Marin, C.; Mora-Plazas, M.; Baylin, A. Vitamin D deficiency and age atmenarche: A prospective study. Am. J. Clin. Nutr. 2011, 94, 1020–1025. [Google Scholar] [CrossRef]
- Zhu, K.; Zhang, Q.; Foo, L.H.; Trube, A.; Ma, G.; Hu, X.; Du, X.; Cowell, C.T.; Fraser, D.R.; Greenfield, H. Growth, bone mass, and vitamin D status of Chinese adolescent girls 3 y after withdrawal of milk supplementation. Am. J. Clin. Nutr. 2006, 83, 714–721. [Google Scholar] [CrossRef] [Green Version]
- Cheng, G.; Buyken, A.E.; Shi, L.; Karaolis-Danckert, N.; Kroke, A.; Wudy, S.A.; Degen, G.H.; Remer, T. Beyond overweight: Nutrition as an important lifestyle factor influencing timing of puberty. Nutr. Rev. 2012, 70, 133–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calcaterra, V.; Cena, H.; Regalbuto, C.; Vinci, F.; Porri, D.; Verduci, E.; Mameli, C.; Zuccotti, G.V. The Role of Fetal, Infant, and Childhood Nutrition in the Timing of Sexual Maturation. Nutrients 2021, 13, 419. https://doi.org/10.3390/nu13020419
Calcaterra V, Cena H, Regalbuto C, Vinci F, Porri D, Verduci E, Mameli C, Zuccotti GV. The Role of Fetal, Infant, and Childhood Nutrition in the Timing of Sexual Maturation. Nutrients. 2021; 13(2):419. https://doi.org/10.3390/nu13020419
Chicago/Turabian StyleCalcaterra, Valeria, Hellas Cena, Corrado Regalbuto, Federica Vinci, Debora Porri, Elvira Verduci, Chiara Mameli, and Gian Vincenzo Zuccotti. 2021. "The Role of Fetal, Infant, and Childhood Nutrition in the Timing of Sexual Maturation" Nutrients 13, no. 2: 419. https://doi.org/10.3390/nu13020419
APA StyleCalcaterra, V., Cena, H., Regalbuto, C., Vinci, F., Porri, D., Verduci, E., Mameli, C., & Zuccotti, G. V. (2021). The Role of Fetal, Infant, and Childhood Nutrition in the Timing of Sexual Maturation. Nutrients, 13(2), 419. https://doi.org/10.3390/nu13020419