
Potential Health Benefits Associated with Lunasin Concentration in Dietary Supplements and Lunasin-Enriched Soy Extract

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Lunasin-Based Commercial Dietary Supplement Samples
2.3. Pilot-Plant Production of Lunasin-Enriched Soy Extract
2.4. Analysis of Lunasin during Pilot-Plant Based Production of Lunasin-Enriched Soy Extract
2.5. Lunasin Purification Anion Exchange Chromatography
2.6. Protein Concentration Measurement by Detergent-Compatible (DC) Protein Assay
2.7. Enzyme-Linked Immunosorbent Assay
2.8. Antioxidant Capacity
2.9. Histone Acetyltransferase (HAT) Inhibitory Screening Assay
2.10. Cell Cytotoxicity
2.11. Protein Kinase B (Akt) Pathway Expression
2.12. Statistical Analysis
3. Results and Discussion
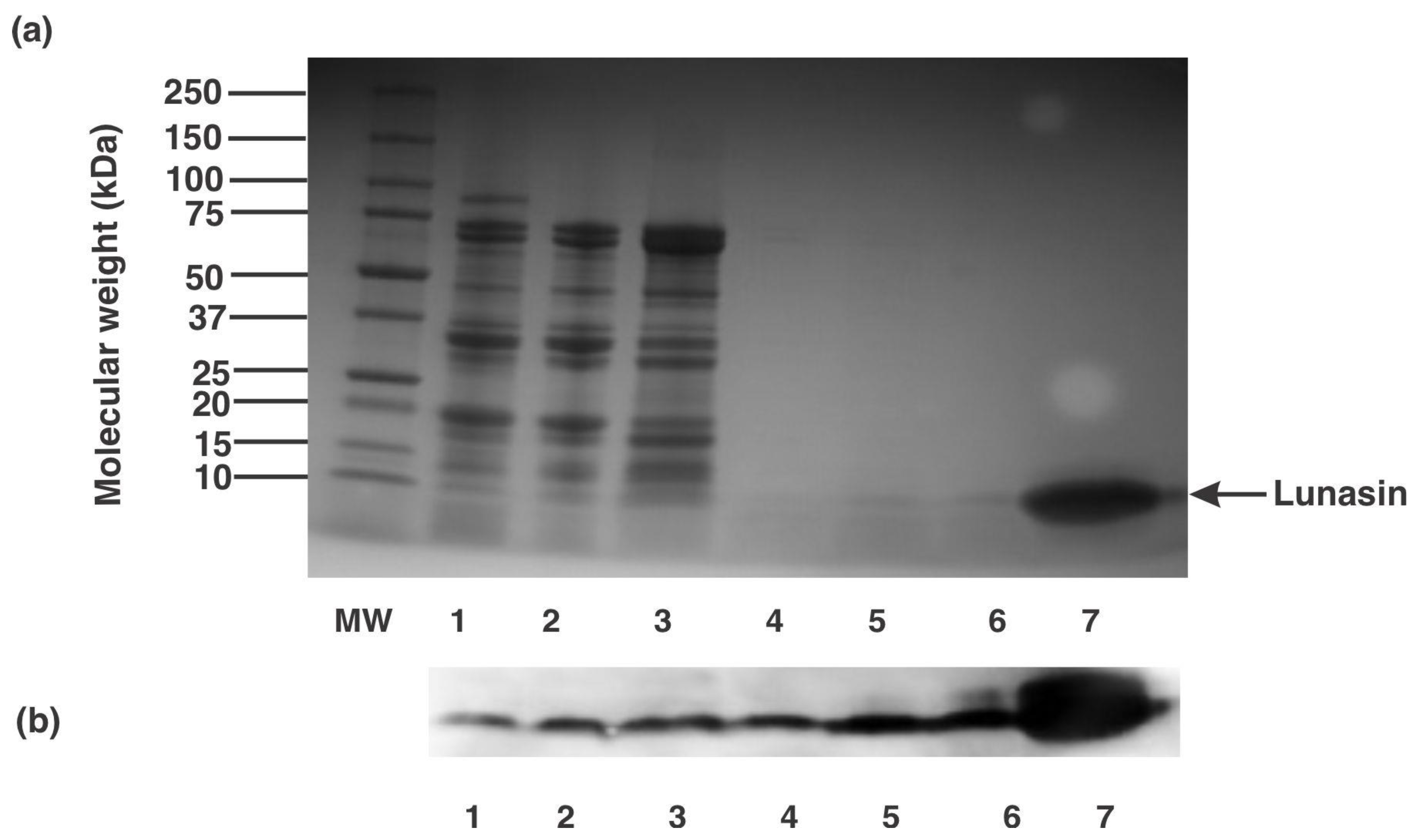
3.1. Analysis of Lunasin Concentrations in Lunasin-Based Commercial Dietary Supplements
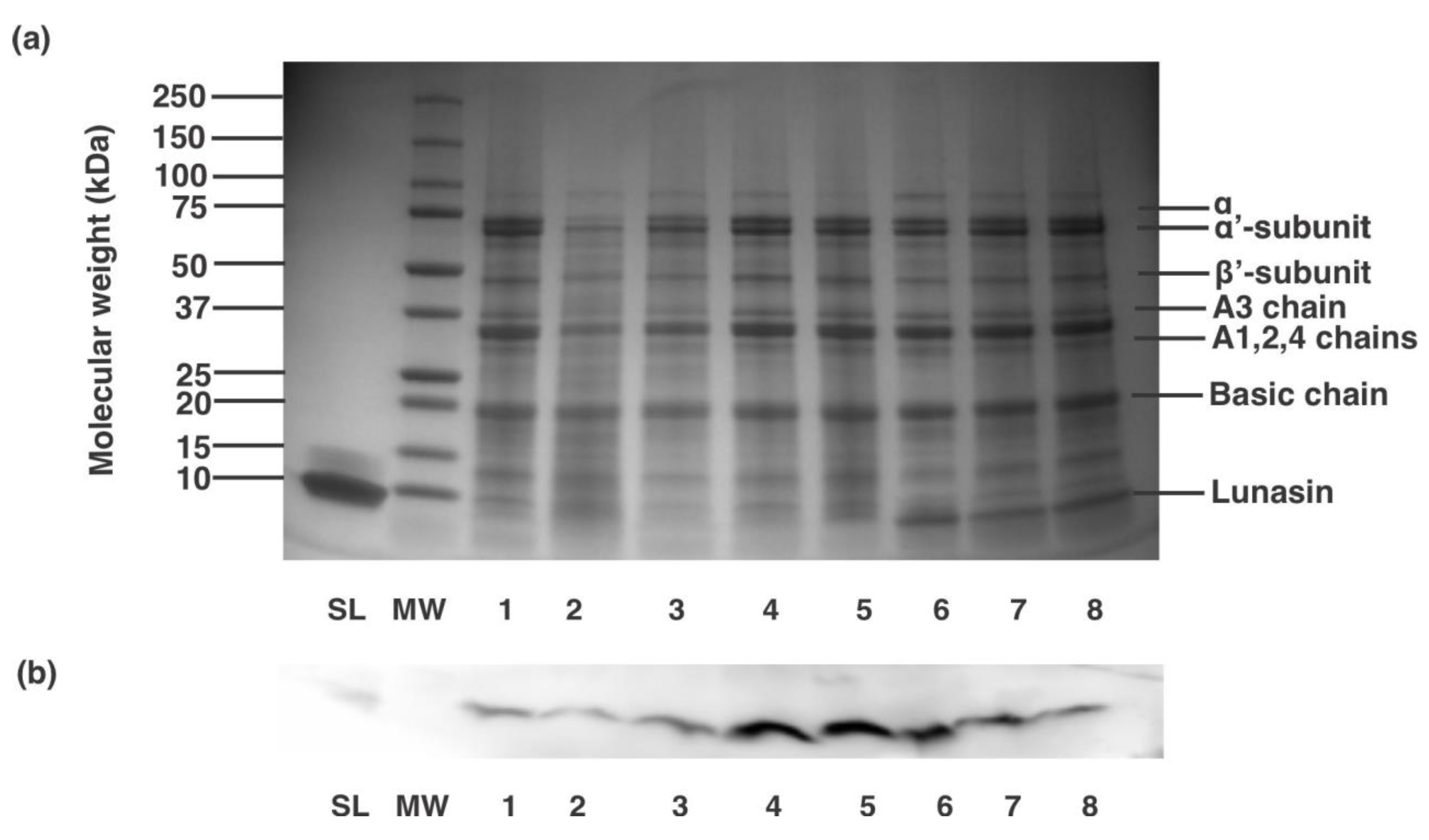
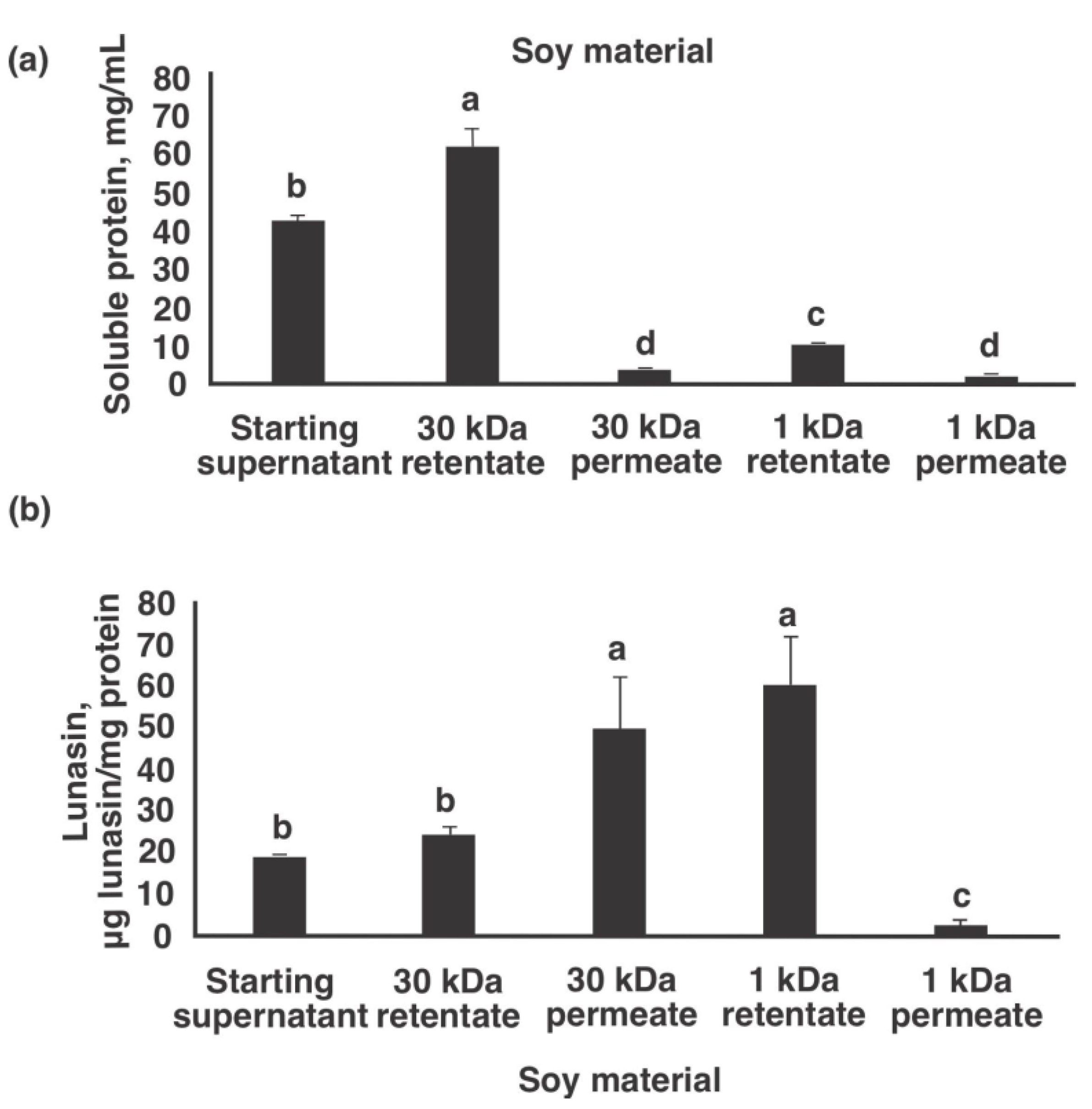
3.2. Pilot-Plant Production of Lunasin-Enriched Soy Extract
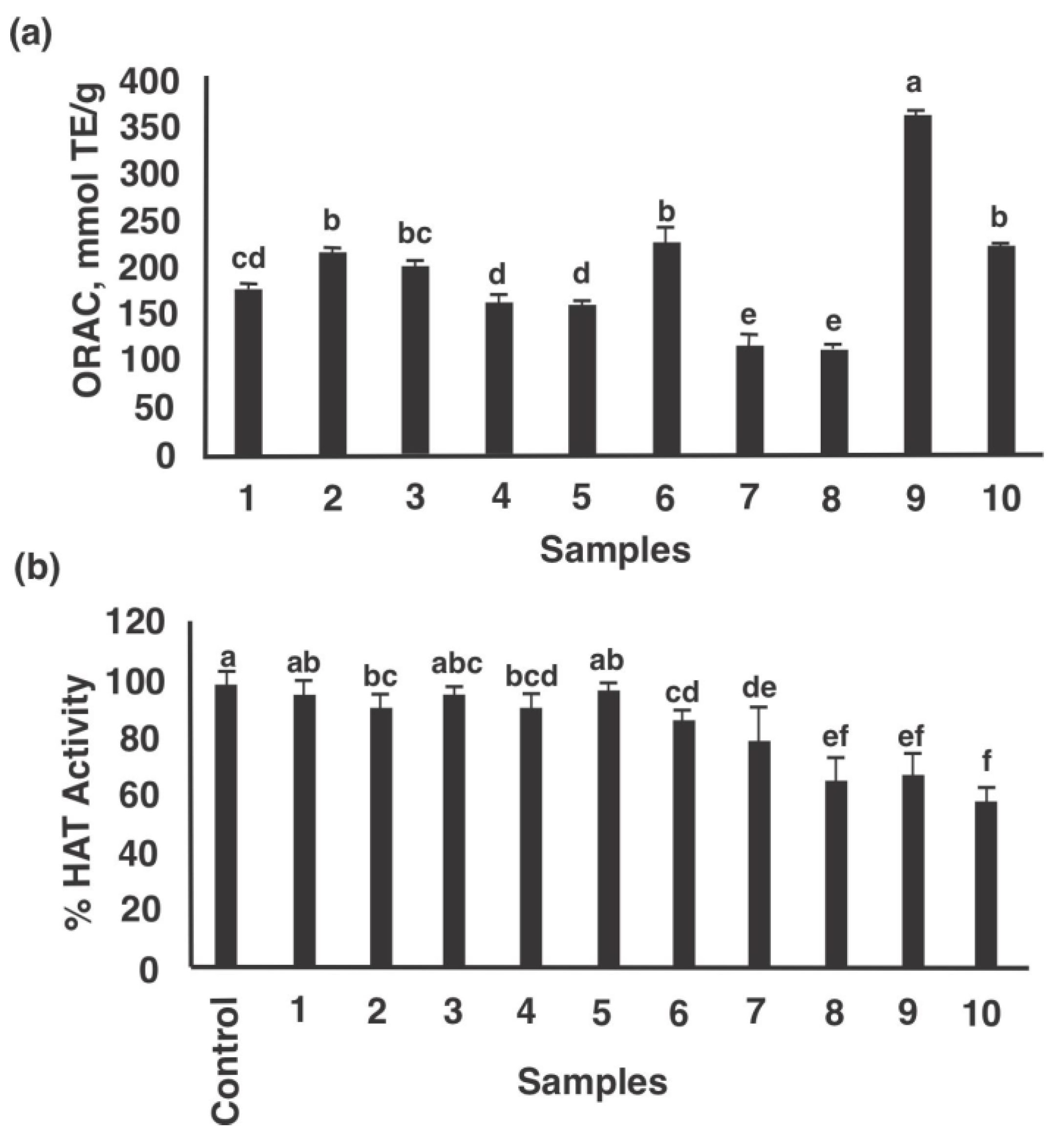
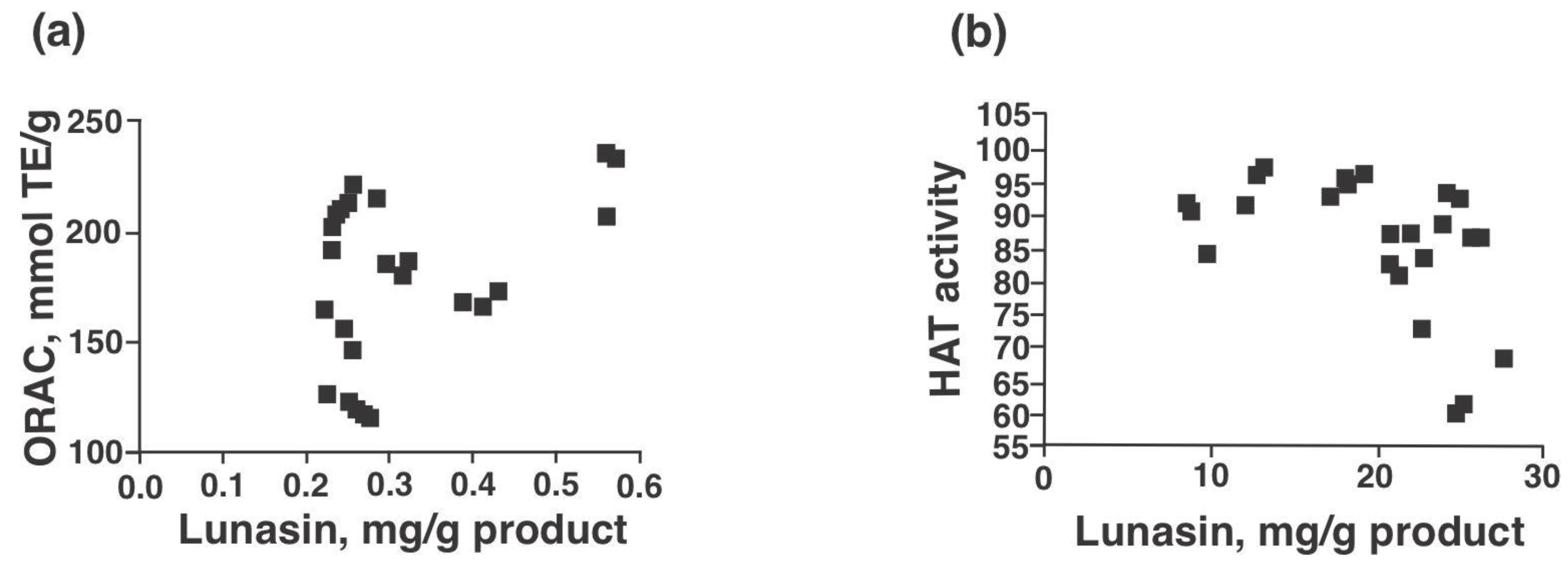
3.3. Antioxidant Capacity and Inhibition Potential of Histone Acetyltransferase (HAT)
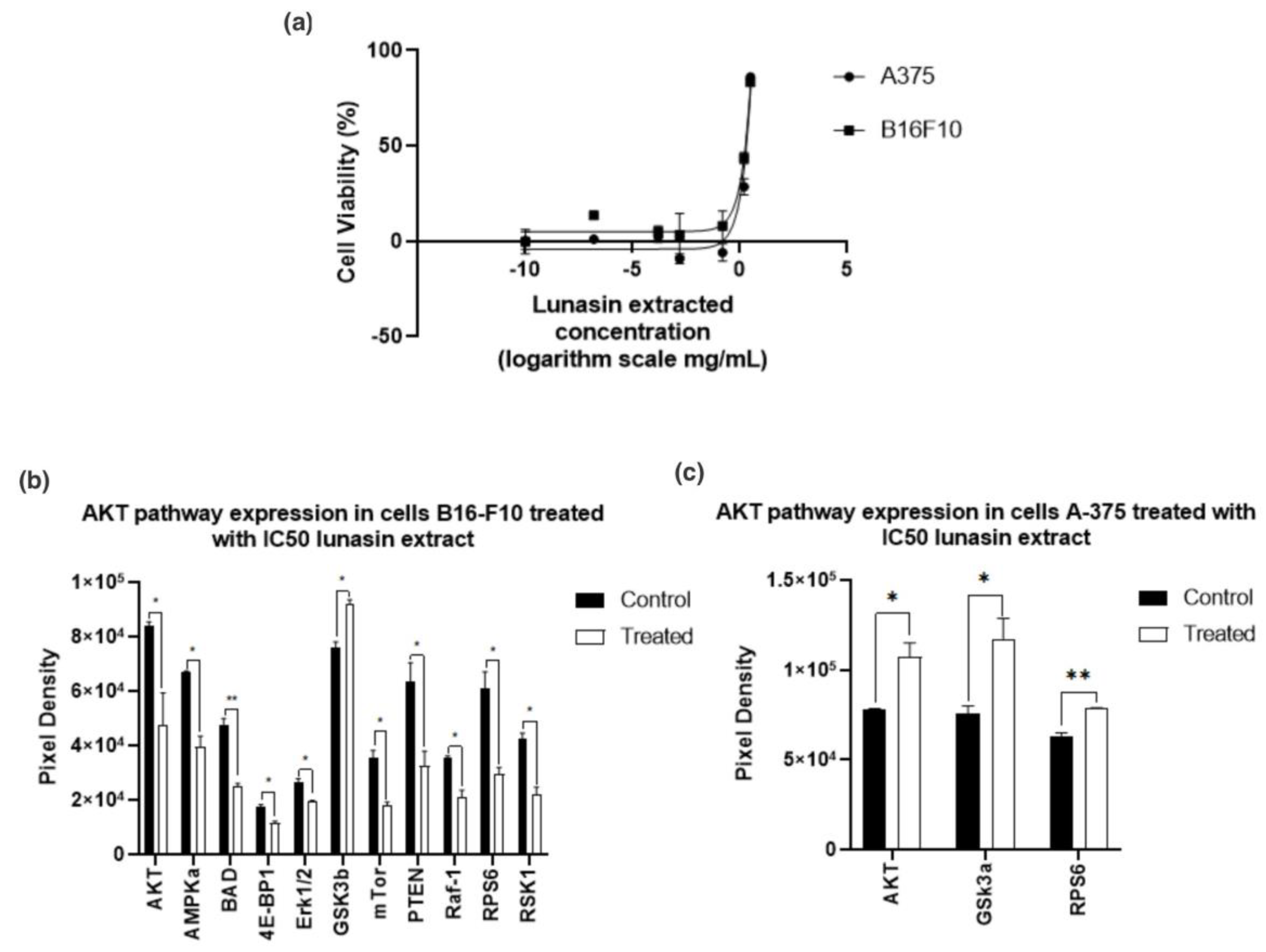
3.4. Cell Cytotoxicity
3.5. Protein Kinase B (Akt) Pathway Expression
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dia, V.P.; Wang, W.; Oh, V.L.; de Lumen, B.O.; Gonzalez de Mejia, E. Isolation, Purification and Characterisation of Lunasin from Defatted Soybean Flour and in Vitro Evaluation of Its Anti-Inflammatory Activity. Food Chem. 2009, 114, 108–115. [Google Scholar] [CrossRef]
- Pabona, J.M.P.; Dave, B.; Su, Y.; Montales, M.T.E.; de Lumen, B.O.; Gonzalez de Mejia, E.; Rahal, O.M.; Simmen, R.C.M. The Soybean Peptide Lunasin Promotes Apoptosis of Mammary Epithelial Cells via Induction of Tumor Suppressor PTEN: Similarities and Distinct Actions from Soy Isoflavone Genistein. Genes Nutr. 2013, 8, 79–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Indiano-Romacho, P.; Fernández-Tomé, S.; Amigo, L.; Hernández-Ledesma, B. Multifunctionality of Lunasin and Peptides Released during Its Simulated Gastrointestinal Digestion. Food Res. Int. 2019, 125, 108513. [Google Scholar] [CrossRef]
- Chatterjee, C.; Gleddie, S.; Xiao, C.W. Soybean Bioactive Peptides and Their Functional Properties. Nutrients 2018, 10, 1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Ledesma, B.; Hsieh, C.C.; de Lumen, B.O. Chemopreventive Properties of Peptide Lunasin: A Review. Protein Pept. Lett. 2013, 20, 424–432. [Google Scholar] [CrossRef]
- Fernández-Tomé, S.; Ramos, S.; Cordero-Herrera, I.; Recio, I.; Goya, L.; Hernández-Ledesma, B. In Vitro Chemo-Protective Effect of Bioactive Peptide Lunasin against Oxidative Stress in Human HepG2 Cells. Food Res. Int. 2014, 62, 793–800. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.C.; Lewis, D.; Tung, C.Y.; Han, L.; Henriquez, S.M.P.; Voiles, L.; Lupov, I.P.; Pelloso, D.; Sinn, A.L.; Pollok, K.E.; et al. Soypeptide Lunasin in Cytokine Immunotherapy for Lymphoma. Cancer Immunol. Immunother. 2014, 63, 283–295. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Li, H.; Wang, X. Lunasin Abrogates Monocytes to Endothelial Cells. Mol. Immunol. 2017, 92, 146–150. [Google Scholar] [CrossRef]
- Fernández-Tomé, S.; Sanchón, J.; Recio, I.; Hernández-Ledesma, B. Transepithelial Transport of Lunasin and Derived Peptides: Inhibitory Effects on the Gastrointestinal Cancer Cells Viability. J. Food Compos. Anal. 2018, 68, 101–110. [Google Scholar] [CrossRef]
- Bachala, D.; El-Refai, N.; Greenfield, E.; Aminoshariae, A.; Mickel, A. The Effect of Lunasin on Receptor Activator of Nuclear Factor Kappa-B Ligand−mediated Osteoclast Formation from RAW 264.7 Cells. J. Endod. 2018, 44, 997–999. [Google Scholar] [CrossRef]
- Vuyyuri, S.B.; Shidal, C.; Davis, K.R. Development of the Plant-Derived Peptide Lunasin as an Anticancer Agent. Curr. Opin. Pharmacol. 2018, 41, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Tomé, S.; Hernández-Ledesma, B. Current State of Art after Twenty Years of the Discovery of Bioactive Peptide Lunasin. Food Res. Int. 2019, 116, 71–78. [Google Scholar] [CrossRef]
- Cam, A.; Sivaguru, M.; Gonzalez de Mejia, E. Endocytic Mechanism of Internalization of Dietary Peptide Lunasin into Macrophages in Inflammatory Condition Associated with Cardiovascular Disease. PLoS ONE 2013, 8, e72115. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez de Mejia, E.; Dia, V.P. Lunasin and Lunasin-like Peptides Inhibit Inflammation through Suppression of NF-ΚB Pathway in the Macrophage. Peptides 2009, 30, 2388–2398. [Google Scholar] [CrossRef]
- García-Nebot, M.J.; Recio, I.; Hernández-Ledesma, B. Antioxidant Activity and Protective Effects of Peptide Lunasin against Oxidative Stress in Intestinal Caco-2 Cells. Food Chem. Toxicol. 2014, 65, 155–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, C.C.; Hernández-Ledesma, B.; de Lumen, B.O. Lunasin-Aspirin Combination Against NIH/3T3 Cells Transformation Induced by Chemical Carcinogens. Plant Foods Hum. Nutr. 2011, 66, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.C.; Hernández-Ledesma, B.; Jeong, H.J.; Park, J.H.; de Lumen, B.O. Complementary Roles in Cancer Prevention: Protease Inhibitor Makes the Cancer Preventive Peptide Lunasin Bioavailable. PLoS ONE 2010, 5, e8890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galvez, A.F.; Chen, N.; Macasieb, J.; de Lumen, B.O. Chemopreventive Property of a Soybean Peptide (Lunasin) That Binds to Deacetylated Histones and Inhibits Acetylation. Cancer Res. 2001, 61, 7473–7478. [Google Scholar]
- Dia, V.P.; Gonzalez de Mejia, E. Lunasin Potentiates the Effect of Oxaliplatin Preventing Outgrowth of Colon Cancer Metastasis, Binds to A5β1 Integrin and Suppresses FAK/ERK/NF-ΚB Signaling. Cancer Lett. 2011, 313, 167–180. [Google Scholar] [CrossRef]
- Tung, C.Y.; Kyazike, S.; Lewis, D.; Han, L.; Kolb, A.; Pelloso, D.; Sinn, A.; Pollok, K.E.; Srivastava, S.; Robertson, M.J.; et al. Activation Of Natural Killer Cells By Soypeptide Lunasin and Cytokine: Implication In Cancer Immunotherapy For Lymphoma. Blood 2013, 122, 1042. [Google Scholar] [CrossRef]
- Jeong, H.J.; Jeong, J.B.; Kim, D.S.; de Lumen, B.O. Inhibition of Core Histone Acetylation by the Cancer Preventive Peptide Lunasin. J. Agric. Food Chem. 2007, 55, 632–637. [Google Scholar] [CrossRef]
- Galvez, A.F. Products and Methods Using Soy Peptides to Lower Total and LDL Cholesterol Levels. U.S. Patent 8598111 B2, 3 December 2013. [Google Scholar]
- Galvez, A.F.; Schmidt, R.; Hastings, C. Products and Methods Using Lunasin-Enriched Soy Extract Mixtures to Reduce Free Fatty Acid Levels, Increase Leptin Levels and Increase Adiponectin Levels in Plasma. Patent WO2014145086 A2, 18 September 2014. [Google Scholar]
- Gonzalez de Mejia, E.; Vásconez, M.; de Lumen, B.O.; Nelson, R. Lunasin Concentration in Different Soybean Genotypes, Commercial Soy Protein, and Isoflavone Products. J. Agric. Food Chem. 2004, 52, 5882–5887. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ledesma, B.; Hsieh, C.C.; de Lumen, B.O. Antioxidant and Anti-Inflammatory Properties of Cancer Preventive Peptide Lunasin in RAW 264.7 Macrophages. Biochem. Biophys. Res. Commun. 2009, 390, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Cavazos, A.; Morales, E.; Dia, V.P.; Gonzalez de Mejia, E. Analysis of Lunasin in Commercial and Pilot Plant Produced Soybean Products and an Improved Method of Lunasin Purification. J. Food Sci. 2012, 77, C539–C545. [Google Scholar] [CrossRef]
- Davis, K.; Barnett, B.; Cai, J.; Mcconnell, E. Lunasin-Containing Complex and Purification of Lunasin from Plants. Patent WO2011060181A1, 19 May 2011. [Google Scholar]
- Prior, R.L.; Hoang, H.; Gu, L.; Wu, X.; Bacchiocca, M.; Howard, L.; Hampsch-Woodill, M.; Huang, D.; Ou, B.; Jacob, R. Assays for Hydrophilic and Lipophilic Antioxidant Capacity (Oxygen Radical Absorbance Capacity (ORACFL)) of Plasma and Other Biological and Food Samples. J. Agric. Food Chem. 2003, 51, 3273–3279. [Google Scholar] [CrossRef] [PubMed]
- Darmawan, R.; Bringe, N.A.; Gonzalez de Mejia, E. Antioxidant Capacity of Alcalase Hydrolysates and Protein Profiles of Two Conventional and Seven Low Glycinin Soybean Cultivars. Plant Foods Hum. Nutr. 2010, 65, 233–240. [Google Scholar] [CrossRef]
- Hernández-Ledesma, B.; Hsieh, C.C.; de Lumen, B.O. Lunasin and Bowman-Birk Protease Inhibitor (BBI) in US Commercial Soy Foods. Food Chem. 2009, 115, 574–580. [Google Scholar] [CrossRef]
- Gonzalez de Mejia, E.; Wang, W.; Dia, V.P. Lunasin, with an Arginine-Glycine-Aspartic Acid Motif, Causes Apoptosis to L1210 Leukemia Cells by Activation of Caspase-3. Mol. Nutr. Food Res. 2010, 54, 406–414. [Google Scholar] [CrossRef]
- Dia, V.P.; Torres, S.; De Lumen, B.O.; Erdman, J.W.; Gonzalez de Mejia, E. Presence of Lunasin in Plasma of Men after Soy Protein Consumption. J. Agric. Food Chem. 2009, 57, 1260–1266. [Google Scholar] [CrossRef]
- Seber, L.E.; Barnett, B.W.; McConnell, E.J.; Hume, S.D.; Cai, J.; Boles, K.; Davis, K.R. Scalable Purification and Characterization of the Anticancer Lunasin Peptide from Soybean. PLoS ONE 2012, 7, e35409. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Jeong, H.J.; De Lumen, B.O. In Vitro Digestibility of the Cancer-Preventive Soy Peptides Lunasin and BBI. J. Agric. Food Chem. 2007, 55, 10703–10706. [Google Scholar] [CrossRef]
- Krishnan, H.B.; Wang, T.T.Y. An Effective and Simple Procedure to Isolate Abundant Quantities of Biologically Active Chemopreventive Lunasin Protease Inhibitor Concentrate (LPIC) from Soybean. Food Chem. 2015, 177, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ledesma, B.; Hsieh, C.-C.; de Lumen, B.O. Relationship between Lunasin’s Sequence and Its Inhibitory Activity of Histones H3 and H4 Acetylation. Mol. Nutr. Food Res. 2011, 55, 989–998. [Google Scholar] [CrossRef]
- Shidal, C.; Al-Rayyan, N.; Yaddanapudi, K.; Davis, K.R. Lunasin Is a Novel Therapeutic Agent for Targeting Melanoma Cancer Stem Cells. Oncotarget 2016, 7, 84128–84141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruzzolini, J.; Peppicelli, S.; Andreucci, E.; Bianchini, F.; Scardigli, A.; Romani, A.; La Marca, G.; Nediani, C.; Calorini, L. Oleuropein, the Main Polyphenol of Olea Europaea Leaf Extract, Has an Anti-Cancer Effect on Human BRAF Melanoma Cells and Potentiates the Cytotoxicity of Current Chemotherapies. Nutrients 2018, 10, 1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dia, V.P.; Gonzalez de Mejia, E. Lunasin Induces Apoptosis and Modifies the Expression of Genes Associated with Extracellular Matrix and Cell Adhesion in Human Metastatic Colon Cancer Cells. Mol. Nutr. Food Res. 2011, 55, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Bringe, N.A.; Berhow, M.A.; Gonzalez de Mejia, E. β-Conglycinins among Sources of Bioactives in Hydrolysates of Different Soybean Varieties That Inhibit Leukemia Cells in Vitro. J. Agric. Food Chem. 2008, 56, 4012–4020. [Google Scholar] [CrossRef]
- Jia, S.; Zhang, S.; Yuan, H.; Chen, N. Lunasin Inhibits Cell Proliferation via Apoptosis and Reduces the Production of Proinflammatory Cytokines in Cultured Rheumatoid Arthritis Synovial Fibroblasts. Biomed Res. Int. 2015, 2015, 346839. [Google Scholar] [CrossRef]
- Zaidi, M.R.; Fisher, D.E.; Rizos, H. Biology of Melanocytes and Primary Melanoma. In Cutaneous Melanoma; Balch, C.M., Atkins, M.B., Garbe, C., Gershenwald, J.E., Halpern, A.C., Kirkwood, J.M., McArthur, G.A., Thompson, J.F., Sober, A.J., Eds.; Springer International Publishing: Cham, Germany, 2020; pp. 3–40. ISBN 978-3-030-05070-2. [Google Scholar]
- Castañeda-Reyes, E.D.; Perea-Flores, M.D.J.; Davila-Ortiz, G.; Lee, Y.; De Mejia, E.G. Development, Characterization and Use of Liposomes as Amphipathic Transporters of Bioactive Compounds for Melanoma Treatment and Reduction of Skin Inflammation: A Review. Int. J. Nanomed. 2020, 15, 7627–7650. [Google Scholar] [CrossRef]
- Jiang, Q.; Pan, Y.; Cheng, Y.; Li, H.; Liu, D.; Li, H. Lunasin Suppresses the Migration and Invasion of Breast Cancer Cells by Inhibiting Matrix Metalloproteinase-2/-9 via the FAK/Akt/ERK and NF-ΚB Signaling Pathways. Oncol. Rep. 2016, 36, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Lv, D.; Guo, L.; Zhang, T.; Huang, L. PRAS40 Signaling in Tumor. Oncotarget 2017, 8, 69076–69085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J. Glycogen Synthase Kinase 3β (GSK3β) in Tumorigenesis and Cancer Chemotherapy. Cancer Lett. 2009, 273, 194–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, G.; Liu, B.; Meng, Z.; Liu, Y.; Li, X.; Wu, X.; Zhou, Q.; Xu, K. MiR-26a Enhances Invasive Capacity by Suppressing GSK3β in Human Lung Cancer Cells. Exp. Cell Res. 2017, 352, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Chao, X.J.; Wu, J.F.; Cheng, B.C.Y.; Su, T.; Fu, X.Q.; Li, T.; Guo, H.; Tse, A.K.W.; Kwan, H.Y.; et al. ERK/GSK3β Signaling Is Involved in Atractylenolide I-Induced Apoptosis and Cell Cycle Arrest in Melanoma Cells. Oncol. Rep. 2015, 34, 1543–1548. [Google Scholar] [CrossRef] [PubMed]
- John, J.K.; Paraiso, K.H.T.; Rebecca, V.W.; Cantini, L.P.; Abel, E.V.; Pagano, N.; Meggers, E.; Mathew, R.; Krepler, C.; Izumi, V.; et al. GSK3β Inhibition Blocks Melanoma Cell/Host Interactions by Downregulating N-Cadherin Expression and Decreasing FAK Phosphorylation. J. Investig. Dermatol. 2012, 132, 2818–2827. [Google Scholar] [CrossRef] [Green Version]
- Jheng, H.-F.; Hirotsuka, M.; Goto, T.; Shibata, M.; Matsumura, Y.; Kawada, T. Dietary Low-Fat Soy Milk Powder Retards Diabetic Nephropathy Progression via Inhibition of Renal Fibrosis and Renal Inflammation. Mol. Nutr. Food Res. 2017, 61, 1600461. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Product | Lunasin, mg/100 g Powder | Lunasin, mg/Serving | Protein *, g/Serving | Serving Size *, g (mL) | Other Ingredients |
|---|---|---|---|---|---|
| Soy Sentials® | 31.1 ± 1.4 c | 6.8 ± 0.3 ab | 10 | 22 (240) | 20 mg isoflavones, 2.5 g protective blend, LunaRich soy powder, red clover, wild Mexican yam, green tea extract (marketed as a protective supplement for women) |
| Slimplicity® | 26.3 ± 1.8 cd | 7.1 ± 0.5 ab | 10 | 27 (240) | Conjugated linoleic acid, LunaRich soy powder, Advantra Z, L-carnitine, inulin, ChromeMate, CitriMax, Optizinc (marketed as meal replacement shake) |
| Now® | 23.6 ± 0.6 d | 4.4 ± 0.1 c | 7 | 18.76 (240) | 197 mg proprietary blend, LunaRich soy powder, vitamins and minerals |
| Provantage® | 23.3 ± 1.1 d | 6.1 ± 0.3 b | 14 | 26 (240) | 694 mg amino acid blend, 1220 mg performance blend, LunaRich soy powder, Tonalin, medium-chain triglycerides, creatine, CoQ10 (marketed as a dietary supplement for sports nutrition) |
| LunaRich X™ | 41.0 ± 2.1 b | 0.2 ± 0.0 d | --- | 0.4 (capsule) | 125 mg soy bioactive lunasin peptide (concentrated form of lunasin) |
| GlucAffect® | 56.4 ± 0.6 a | 6.8 ± 0.1 ab | 5 | 12 (240) | 15 mg pycnogenol, 2221 mg proprietary blend, LunaRich soy powder, omega-3-fish oils, Salacia extract, Glucohelp (marketed as a dietary supplement for maintaining healthy blood sugar) |
| Now® for Kids (Vanilla) | 26.2 ± 1.1 d | 7.6 ± 0.3 a | 5 | 29 (240) | 408 mg proprietary blend, LunaRich soy powder, omega-3 fatty acid, phosphatidylserine, phosphatidylcholine, grape seed extract (marketed to help kids boost energy and mental performance) |
| Now® for Kids (Chocolate) | 25.1 ± 3.4 d | 7.3 ± 1.0 a | 5 | 29 (240) | 408 mg proprietary blend, LunaRich soy powder, omega-3 fatty acid, phosphatidylserine, phosphatidylcholine, grape seed extract (marketed to help kids boost energy and mental performance) |
| Abbreviation | Full Name | Expression (%) | Epitope |
|---|---|---|---|
| B16-F10 | |||
| AKT | Protein B kinase | 57 | Ser473 |
| AMPKa | AMP-activated protein kinase | 59 | Thr172 |
| BAD | BCL2-associated agonist of cell death | 51 | Ser112 |
| 4E-BP1 | 4E-binding protein 1 | 68 | Thr36 |
| ERK 1/2 | Extracellular signal-regulated kinase | 72 | ERK1 Thr202/Tyr204 ERK2 Tyr185/Tyr187 |
| GSK3b | Glycogen synthase kinase 3b | 119 | Ser9 |
| mTOR | Mammalian target of rapamycin | 54 | Ser2448 |
| PTEN | Phosphatase and tensin homolog | 56 | Ser380 |
| Raf-1 | Rapidly accelerated fibrosarcoma-1 | 59 | Ser301 |
| RPS6 | Ribosomal protein S6 | 52 | Ser235/Ser236 |
| RSK1 | Ribosomal S6 kinase 1 | 50 | Ser380 |
| A-375 | |||
| Akt | Protein B kinase | 137 | Ser 473 |
| GSK3a | Glycogen synthase kinase 3a | 155 | Ser21 |
| RPS6 | Ribosomal protein S6 | 124 | Ser380 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez de Mejia, E.; Castañeda-Reyes, E.D.; Mojica, L.; Dia, V.; Wang, H.; Wang, T.; Johnson, L.A. Potential Health Benefits Associated with Lunasin Concentration in Dietary Supplements and Lunasin-Enriched Soy Extract. Nutrients 2021, 13, 1618. https://doi.org/10.3390/nu13051618
Gonzalez de Mejia E, Castañeda-Reyes ED, Mojica L, Dia V, Wang H, Wang T, Johnson LA. Potential Health Benefits Associated with Lunasin Concentration in Dietary Supplements and Lunasin-Enriched Soy Extract. Nutrients. 2021; 13(5):1618. https://doi.org/10.3390/nu13051618
Chicago/Turabian StyleGonzalez de Mejia, Elvira, Erick Damian Castañeda-Reyes, Luis Mojica, Vermont Dia, Hui Wang, Toni Wang, and Lawrence A. Johnson. 2021. "Potential Health Benefits Associated with Lunasin Concentration in Dietary Supplements and Lunasin-Enriched Soy Extract" Nutrients 13, no. 5: 1618. https://doi.org/10.3390/nu13051618
APA StyleGonzalez de Mejia, E., Castañeda-Reyes, E. D., Mojica, L., Dia, V., Wang, H., Wang, T., & Johnson, L. A. (2021). Potential Health Benefits Associated with Lunasin Concentration in Dietary Supplements and Lunasin-Enriched Soy Extract. Nutrients, 13(5), 1618. https://doi.org/10.3390/nu13051618