
Enteral Feeding Interventions in the Prevention of Necrotizing Enterocolitis: A Systematic Review of Experimental and Clinical Studies

 , , ,
, , , 
Abstract
:1. Introduction
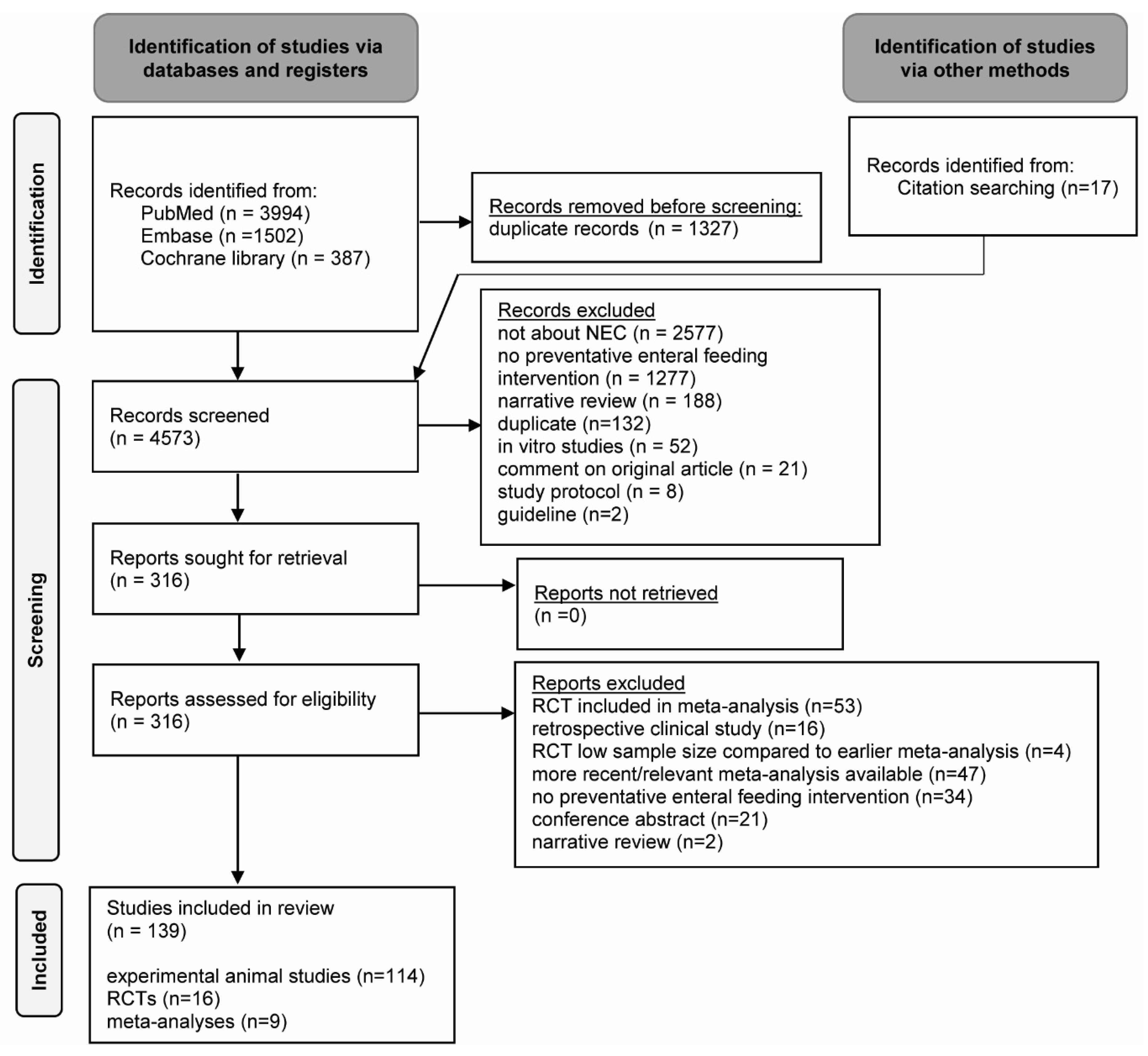
2. Materials and Methods
2.1. Search Strategy
2.2. Selection Criteria
2.3. Selection Process
2.4. Data Extraction
2.5. Risk of Bias Assessment
2.6. Certainty of Evidence Assessment
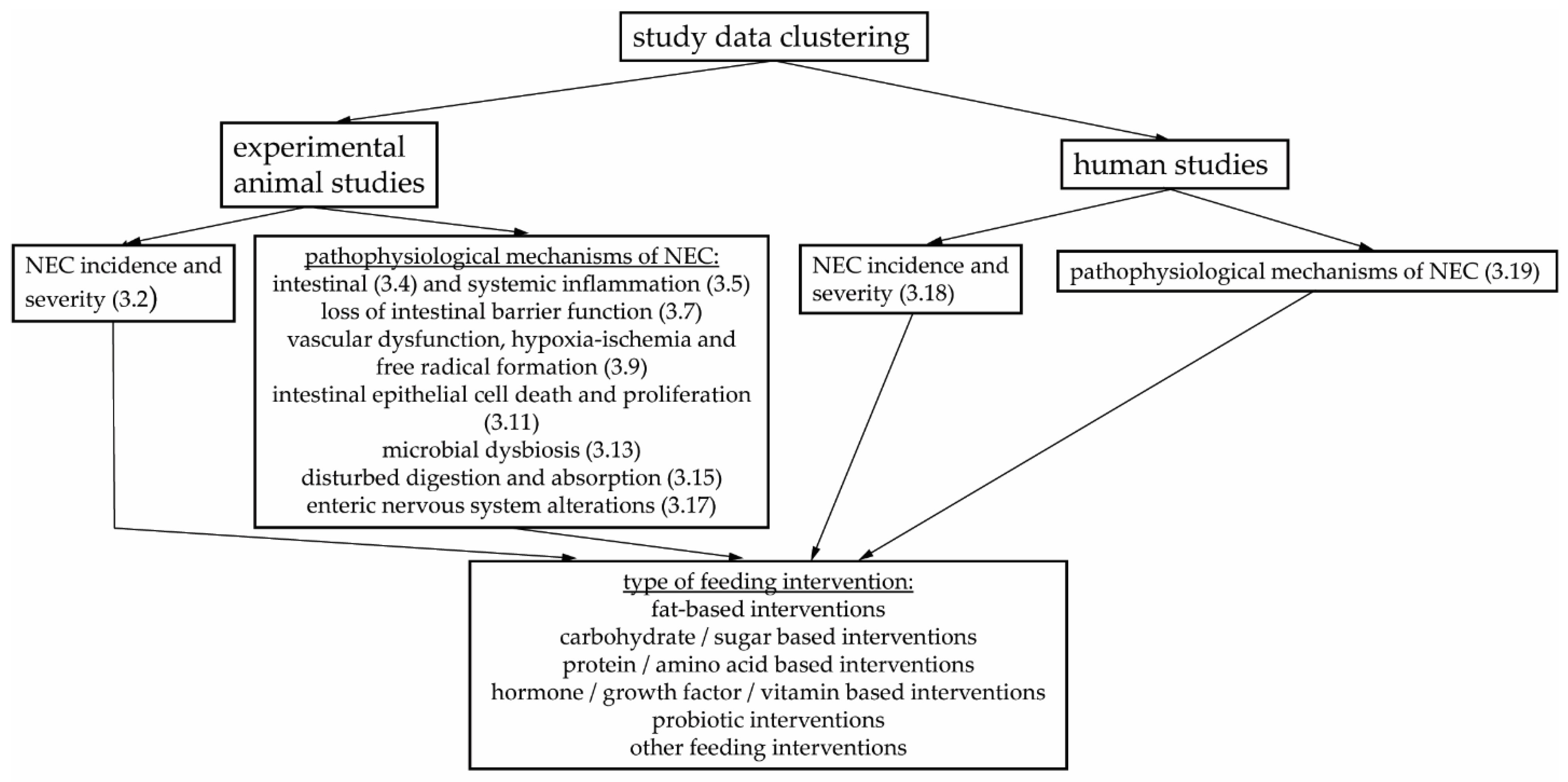
3. Results
3.1. Study Characteristics
3.2. Enteral Feeding Interventions Affecting NEC Incidence and Severity in Animal Studies
3.3. NEC Pathophysiology: Intestinal and Systemic Inflammation
3.4. Enteral Feeding and Intestinal Inflammation in Animal Models of NEC
3.4.1. Fat-Based Feeding Interventions
3.4.2. Carbohydrate or Sugar-Based Feeding Interventions
3.4.3. Protein or Amino Acid-Based Feeding Interventions
3.4.4. Hormone, Growth Factor or Vitamin-Based Feeding Interventions
3.4.5. Probiotic Feeding Interventions
3.4.6. Other Enteral Feeding Interventions

{kind=link}
{kind=link}
| Enteral Feeding Intervention | Effect on Intestinal Inflammation (Compared to NEC Protocol Exposure without Feeding Intervention) |
|---|---|
| Fat-based interventions | |
| Fish oil (n-3 PUFA) | Intestinal PAF (protein) ↓ [82] Intestinal leukotriene B4 (protein) ↓ [82] Intestinal IL6 (protein) ↓ [83] Intestinal TNFα (protein) ↓ [83] |
| AA + DHA | Duodenal, jejunal and ileal TLR 4 (mRNA) ↓ [44] Intestinal TLR2 (mRNA) = [44] Intestinal PLA2-II (mRNA) ↓ [45] Intestinal PLA2-II (mRNA) = [44] Ileal, colonic and intestinal PAFR (mRNA) ↓ [44,45] |
| DHA | Intestinal TLR4 (mRNA) = [44] Intestinal TLR2 (mRNA) = [44] Intestinal PLA2-II (mRNA) = [44] Ileum and colon PAFR (mRNA) ↓ [44] |
| DHA (maternal intervention) | Ileal DHA ↑ [43] Ileal EPA ↑ [43] Small intestinal iκbα (mRNA) ↓ [43] Small intestinal IκBβ (mRNA) ↓ [43] Small intestinal PPARϒ (mRNA) ↓ [43] |
| EPA (maternal intervention) | Ileal DHA ↑ [43] Ileal EPA ↑ [43] Small intestinal IκBα (mRNA) ↓ [43] Small intestinal IκBβ (mRNA) ↓ [43] Small intestinal PPARϒ (mRNA) ↓ [43] |
| Egg phospholipids | Intestinal TLR 4 (mRNA) ↓ [44] Ileal TLR2 (mRNA) ↓ [44] Intestinal PLA2 (mRNA) = [44] Ileal and colonic PAFR (mRNA) ↓ [44] |
| BCFA | Ileal IL10 (mRNA) ↑ [46] Ileal IL10 (protein) ↑ [46] |
| Pomegranate seed oil | Ileal IL6 (mRNA) ↓ [47] Ileal IL8 (mRNA) ↓ [47] Ileal IL12 (mRNA) ↓ [47] Ileal IL23 (mRNA) ↓ [47] Ileal TNFα (mRNA) ↓ [47] |
| Pre-digested fat (less long chain triacylglycerol, not dependent on intestinal lipases) | Intestinal IL1β (mRNA) ↓ [85] Intestinal TNFα (mRNA) ↓ [85] |
| Very low-fat diet | Intestinal IL1β (mRNA) ↓ [85] Intestinal TNFα (mRNA) ↓ [85] |
| MFGM | Ileal IL6 (mRNA) ↓ [48] Ileal IL1β (mRNA) ↓ [48] Ileal TNFα (mRNA) ↓ [48] Ileal TLR4 (protein) ↓ [48] |
| MPL | Intestinal IL10 (protein) ↑ [84] Intestinal TNFα (protein) ↓ [84] Intestinal IL6 (protein) ↓ [84] Intestinal IL8 (protein) ↓ [84] Intestinal TLR4 (protein) ↓ [84] Intestinal p65 (protein) ↓ [84] Intestinal p50 (protein) ↓ [84] |
| Carbohydrate/sugar-based interventions | |
| HMO | Ileal IL6 (mRNA) ↓ [49] Ileal IL8 (mRNA) ↓ [49] Ileal IL1β (mRNA) ↓ [49] Ileal TLR4 (mRNA) ↓ [49] Ileal IL6 (protein) ↓ [49] Ileal IL8 (protein) ↓ [49,88] Ileal phosphorylated NFκβ (protein) ↓ [49] Ileal phosphorylated IκBα (protein) ↓ [49] Ileal TLR4 (protein) ↓ [49] |
| Mixture of four HMO | Small intestinal IL10 (mRNA) ↑ [134] Small intestinal IL12 (mRNA) ↑ [134] Small intestinal TGF-β (mRNA) ↑ [134] Small intestinal TLR4 (mRNA) ↑ [134] Small intestinal IL8 (mRNA) = [134] Small intestinal IFNϒ (mRNA) = [134] Small intestinal TNFα (mRNA) = [134] Small intestinal TLR2 (mRNA) = [134] |
| 2′-FL | Intestinal IL6 (mRNA) ↓ [91] (Small) intestinal IL1β (mRNA) ↓ [91] [92] Small intestinal TNFα (mRNA) ↓ [92] (Small) intestinal TLR4 (mRNA) ↓ [91] [92] |
| 6′-SL | Small intestinal IL1β (mRNA) ↓ [92] Small intestinal TNFα (mRNA) ↓ [92] Small intestinal TLR4 (mRNA) ↓ [92] |
| 2′-FL + 6′-SL | Small intestinal IL1β (mRNA) ↓ [92] Small intestinal TNFα (mRNA) ↓ [92] Small intestinal TLR4 (mRNA) ↓ [92] |
| Sialylated HMO | Ileal mast cell counts ↓ [93] Ileal DPPI activity ↓ [93] Ileal IL6 (protein) ↓ [93] Ileal TNFα (protein) ↓ [93] |
| GOS/FOS | Terminal ileum IL1β (protein) ↓ [136] Terminal ileum TNFα (protein) ↓ [136] Terminal ileum IL1β (mRNA) ↓ [136] Terminal ileum TNFα (mRNA) ↓ [136] Terminal ileum IL6 (mRNA) ↓ [136] |
| GD3 | Ileal TNFα (protein) ↓ [50] Ileal IL6 (protein) ↓ [50] Ileal CCL5 (protein) ↓ [50] Ileal L-selectin (protein) ↓ [50] Ileal TIMP1 (protein) ↑ [50] Ileal IL1ra (protein) ↑ [50] Ileal IL10 (protein) ↑ [50] Ileal Foxp3 (protein) ↑ [50] Ileal Foxp3 cellcount ↑ [50] |
| Protein/amino acid-based interventions | |
| IAP | Ileal endogenous IAP (mRNA) ↑ [160] Ileal TNFα (mRNA) ↓ [160] |
| L-Glutamine/glutamine | Intestinal TNFα (protein) ↓ [95] Intestinal IL10 (protein) ↑ [84] Intestinal TNFα (protein) ↓ [84] Intestinal IL6 (protein) ↓ [84] Intestinal IL8 (protein) ↓ [84] Intestinal TLR4 (protein) ↓ [84] Intestinal p65 (protein) ↓ [84] Intestinal p50 (protein) ↓ [84] Jejununal, ileal and colonic TLR4 (protein) ↓ [97] Jejununal, ileal and colonic TLR4 (mRNA) ↓ [97] Jejununal, ileal and colonic TLR2 (protein) ↓ [97] Jejununal, ileal and colonic TLR2 (mRNA) ↓ [97] |
| Arginine | Ileal IL6 (mRNA) ↓ [98] Ileal TNFα (mRNA) ↓ [98] |
| N-Acetylcysteine | Intestinal IL1β (mRNA) ↓ [85] Intestinal TNFα (mRNA) ↓ [85] |
| Lactoferrin | Ileal IL6 (mRNA) ↓ [101] Ileal TNFα (mRNA) ↓ [101] |
| Bovine lactoferrin | Proximal small intestinal IL1β (protein) ↓ [35] |
| Hormone/growth factor/vitamin based interventions | |
| EGF | Ileal IL18 (mRNA) ↓ [53] Ileal IL10 (mRNA) ↑ [53] Ileal Sp1 (mRNA) ↑ [53] |
| Recombinant EGF from soybean extract | Ileal COX2 (mRNA) ↓ [104] |
| HB-EGF | Intestinal M1 macrophages cellcount (CD86) ↓ [57] Intestinal % M1 macrophages/total macrophages (CD86/CD68) ↓ [57] Intestinal M2 macrophages cellcount (CD206) ↑ [57] Intestinal % M1 macrophages/total macrophages (CD206/CD68) ↑ [57] |
| TGF-β1 | Ileal Smad2 activation/phosphorylation ↑ [64] Ileal phosphorylated NFκβ positive intestinal epithelial cells ↓ [64] Ileal IκBα (protein) ↑ [64] |
| IGF1 | Ileal TLR4 (mRNA) ↓ [65] Ileal NFκβ (mRNA) ↓ [65] Ileal IL6 (protein) ↓ [65] |
| Vitamin A | Intestinal IL6 (protein) ↓ [111] Intestinal TNFα (protein) ↓ [111] |
| ATRA | Foxp3 (mRNA) in CD4+ T cells from lamina propria ↑ [109] IL17 (mRNA) in CD4+ T cells from lamina propria ↓ [109] FoxP3+ CD4+ T cells from lamina propria (FACs) ↑ [110] CD4+ Th17 cells from lamina propria (FACs) ↓ [110] Ileal IL1β (mRNA) ↓ [109] Ileal IL6 (mRNA) ↓ [109,110] Ileal IL17 (mRNA) ↓ [110] |
| Vitamin D | Intestinal ÌL6 (mRNA) ↓ [112] Intestinal IL1β (mRNA) ↓ [112] Intestinal TNFα (mRNA) ↓ [112] Intestinal ÌL6 (protein) ↓ [112] Intestinal IL1β (protein) ↓ [112] Intestinal TNFα (protein) ↓ [112] |
| Probiotic interventions | |
| Lactobacillus reuteri DSM 17938 | Intestinal % CD4+ Foxp3+ Treg ↑ [67,69,132] Mesenteric lymph nodes % CD4+ Foxp3+ Treg ↑ [69] Terminal ileum % Foxp3+ CD4+CD8+ Treg cells ↑ [132] Mesenteric lymph nodes % Foxp3+ CD4+CD8+ Treg cells ↓ [132] Intestinal % tolerogenic DC ↑ [67] Intestinal % activated CD4+ Teff ↓ [67] Intestinal % activated effector/memory T cells (CD44+CD45RBlo) ↓ [69] Intestinal % transitional effector T cells (CD44+CD45RBhi) ↓ [69] Ileal IL10 (mRNA) ↑ [68] Ileal IL6 (mRNA) ↓ [68] Ileal TNFα (mRNA) ↓ [68,70] Ileal TLR4 (mRNA) ↓ [68] Ileal TLR1 (mRNA) ↓ [68] Ileal NFκβ (mRNA) ↓ [68] Ileal IL1β (mRNA) ↓ [69,70] Ileal IL1β (protein) ↓ [67,68,69,70] Ileal IFNϒ (protein) ↓ [67] Ileal TNFα (protein) ↓ [68,70] Ileal TLR4 (protein) ↑[68] Ileal phosphorylated Iκβ (protein) ↑[68] Ileal mitogen-activated protein kinase 8 interaction protein 3 (mRNA) ↓ [68] Ileal NFκβ inhibitor-β (mRNA) ↑ [68] |
| Lactobacillus reuteri ATCC PTA 4659 | Ileal IL6 (mRNA) ↓ [68] Ileal TNFα (mRNA) ↓ [68] Ileal TLR4 (mRNA) ↓ [68] Ileal TLR1 (mRNA) ↓ [68] Ileal NFκβ (mRNA) ↓ [68] Ileal TNFα (protein) ↓ [68] Ileal IL1β (protein) ↓ [68] Ileal TLR4 (protein) ↑ [68] Ileal phosphorylated Iκβ (protein) ↑ [68] Ileal IL10 (mRNA) ↑ [68] Ileal myelin and lymphocyte protein (mRNA) ↓ [68] |
| Lactobacillus rhamnosus GG | Ileal TRL4 (mRNA) ↓ [161] Ileal SIGIRR (mRNA) ↑ [161] Ileal SIGIRR (protein) ↑ [161] Ileal A20 (mRNA) ↑ [161] Ileal p-IKKb (protein) ↓ [161] Ileal p-p65 (protein) ↓ [161] Ileal ICAM-1 (protein) ↓ [161] Ileal ICAM-1 (mRNA) ↓ [161] Ileal IL1β (protein) ↓ [161] Ileal IL1β (mRNA) ↓ [161] Ileal IL8 (mRNA) ↓ [161] |
| Lactobacillus reuteri DSM 20016 | Small intestinal IL6 (mRNA) = [72] small intestinal IL1β (mRNA) = [72] Small intestinal CCL2 (mRNA) = [72] Small intestinal CXCL1 (mRNA) = [72] Small intestinal IL10 (mRNA) = [72] |
| Lactobacillus reuteri biofilm on sucrose loaded microspheres | Small intestinal IL6 (mRNA) ↓ [72] small intestinal IL1β (mRNA) ↓ [72] Small intestinal CCL2 (mRNA) ↓ [72] Small intestinal CXCL1 (mRNA) ↓ [72] Small intestinal IL10 (mRNA) ↓ [72] |
| Bifidobacterium bifidum OLB6378 | Ileal IL6 (mRNA) ↓ [73] |
| Bifidobacterium infantis | Intestinal PLA2 II (mRNA) ↓ [75] |
| Bifidobacterium microcapsules | Ileal TLR4 (protein) ↓ [114] Ileal TLR2 (protein) ↓ [114] Ileal NFκβ p65 (protein) ↓ [114] |
| Bifidobacterium adolescentis | Ileal TLR4 (mRNA) ↓ [76] Ileal TOLLIP (mRNA) ↑ [76] Ileal SIGIRR (mRNA) ↑ [76] |
| Bifidobacterium breve M-16V | Ileal TLR4 (mRNA) ↓ [117] Ileal IL1β (mRNA) ↓ [117] Ileal IL6 (mRNA) ↓[117] Ileal TNFα (mRNA) ↓ [117] Ileal IL10 (mRNA) ↓ [117] Ileal TLR2 (mRNA) ↑ [117] Ileal MIP1α (protein) ↓ [117] Ileal IL1β (protein) ↓ [117] |
| Bacteroides fragilis ZY-312 | Intestinal IL1β (protein) ↓ [113] |
| Saccharomyces Boulardii | Terminal ileum IL1β (protein) ↓ [136] Terminal ileum IL6 (protein) ↓ [136] Terminal ileum TNFα (protein) ↓ [136] Terminal ileum IFNβ (mRNA) ↓ [136] Terminal ileum TNFα (mRNA) ↓ [136] |
| CpG-DNA | Ileal IL6 (mRNA) ↓ [39] |
| Other interventions | |
| Ginger | Intestinal IL1β (protein) ↓ [121] Intestinal IL6 (protein) ↓ [121] Intestinal TNFα (protein) ↓ [121] Intestinal MPO (protein) ↓ [121] |
| Fennel seed extracts | Intestinal IL6 (protein) ↓ [122] Intestinal TNFα (protein) ↓ [122] Intestinal MPO (protein) ↓ [122] |
| Bovine milk exosomes | Distal ileal MPO (protein) ↓ [118] |
| Human milk exosomes | Ileal IL6 (mRNA) ↓ [119] Ileal MPO (protein) ↓ [119] |
| Amniotic fluid | Ileal CXCL2 (mRNA) ↓ [63] Ileal CXCL5 (mRNA) ↓ [63] Ileal CCL2 (mRNA) ↓ [63] Ileal CCL5 (mRNA) ↓ [63] Ileal IFNγ (mRNA) ↓ [63] Distal small intestinal IFNγ (mRNA) ↓ [123] Distal small intestinal IL1α (mRNA) ↓ [123] Distal small intestinal TNFα (mRNA) ↓ [123] Middle small intestinal IL1α (mRNA) ↓ [123] Middle small intestinal TNFα (mRNA) ↓ [123] Middle small intestinal IL6 (mRNA) ↓ [123] Middle small intestinal IL8 (mRNA) ↓ [123] |
| Curcumin | Intestinal IL1β (protein) ↓ [124] Intestinal IL6 (protein) ↓ [124] Intestinal IL18 (protein) ↓ [124] Intestinal TNFα (protein) ↓ [124] Intestinal TLR4 (protein) ↓ [124] Intestinal SIRT1 (protein) ↑ [124] Intestinal NRF2 (protein) ↑ [124] Intestinal TLR4 (mRNA) ↓ [124] Intestinal SIRT1 (mRNA) ↑ [124] Intestinal NRF2 (mRNA) ↑ [124] |
| Surfactant protein A | Ileal IL1β (protein) ↓ [80] Ileal TNFα (protein) ↓ [80] Ileal IFNϒ (protein) ↓ [80] Ileal TLR4 (protein) ↓ [80] |
| Human β-defensin-3 | Ileal TNFα (mRNA) ↓ [81] Ileal IL6 (mRNA) ↓ [81] Ileal IL10 (mRNA) ↓ [81] |
| Berberine | Ileal TLR4 (protein) ↓ [79] Ileal IL6 (protein) ↓ [79] Ileal IL10 (protein) ↓ [79] Ileal TLR4 (mRNA) ↓ [79] Ileal NFκβ (mRNA) ↓ [79] Ileal TNFα (mRNA) ↓ [79] |
| Astragaloside IV | Distal ileal TNFα (mRNA) ↓ [126] Distal ileal IL1β (mRNA) ↓ [126] Distal ileal IL6 (mRNA) ↓ [126] Distal ileal NFκβ p65 (mRNA) ↓ [126] Distal ileal MPO (protein) ↓ [126] Distal ileal p-NFκβ p65/ NFκβ p65 (protein) ↓ [126] Distal ileal p-IκBα/ IκBα (protein) ↓ [126] Distal ileal p-IκBα (protein) ↓ [126] Distal ileal p-NFκβ p65 (protein)↓ [126] Distal ileal NFκβ p65 (protein) ↓ [126] Distal ileal IκBα (protein) ↑ [126] |
3.5. Enteral Feeding and Systemic Inflammation in Animal Models of NEC
3.6. NEC Pathophysiology: Loss of Intestinal Barrier Function
3.7. Enteral Feeding and Loss of Intestinal Barrier Function in Animal Models of NEC
3.7.1. Fat-Based Feeding Interventions
3.7.2. Carbohydrate or Sugar-Based Feeding Interventions
3.7.3. Protein or Amino Acid-Based Feeding Interventions
3.7.4. Hormone, Growth Factor or Vitamin-Based Feeding Interventions
3.7.5. Probiotic Feeding Interventions
3.7.6. Other Enteral Feeding Interventions
| Enteral Feeding Intervention | Effect on Intestinal Barrier Function (Compared to NEC Protocol Exposure without Feeding Intervention) |
|---|---|
| Fat-based interventions | |
| PUFA | Endotoxemia (plasma) ↓ [45] |
| DHA | Ileal effective diffusivity amine modified particles ↓ [131] Ileal effective diffusivity carboxyl modified particles ↓ [131] Ileal linear movements E. coli through intestinal mucus ↓ [131] Ileal movement speed E. coli through intestinal mucus ↓ [131] Ileal sialic acid content mucus ↓ [131] Ileal mucus structure (confocal imaging/SEM) = [131] |
| Carbohydrate/sugar-based interventions | |
| Hyaluronan 35 kD | Intestinal permeability (functional orogastric FITC-dextran assay) ↓ [94] Small intestinal occludin (protein) ↑ [94] Small intestinal claudin-4 (protein) ↑ [94] Small intestinal claudin-3 (protein) ↑ [94] Small intestinal claudin-2 (protein) ↑ [94] Small intestinal ZO-1 (protein) ↑ [94] Small intestinal occludin localization [94] Small intestinal claudin-3 localization [94] |
| HMO | Intestinal permeability (functional orogastric FITC-dextran assay) ↓ [38] Ileal number Muc2-positive cells ↑ [38,86] Ileal Muc2 (mRNA) ↑ (trend) [38] Ileal TFF3 (mRNA) ↑ (trend) [38] |
| Mixture of four HMOs | Small intestinal bacterial adhesion and tissue invasion = [134] Small intestinal Muc1 (mRNA) = [134] Small intestinal Muc2 (mRNA) = [134] |
| Protein/amino acid-based interventions | |
| IAP | Ileal intestinal permeability (ex-vivo FITC-dextran assay) ↓ [103] Ileal claudin-1 (protein) ↓ [103] Ileal claudin-3 (protein) ↑ [103] Ileal occludin (protein) = [103] Ileal ZO-1 (protein) = [103] Ileal claudin-1 (mRNA) = [103] Ileal claudin-3 (mRNA) = [103] Ileal occludin (mRNA) = [103] Ileal ZO-1 (mRNA) = [103] Endotoxemia (plasma) ↓ [160] |
| Bovine lactoferrin | Lactulose/mannitol recovery ratio in urine ↑ (only in animals with NEC) [35] |
| Lysozyme | Ileal effective diffusivity amine modified particles ↓ [131] Ileal effective diffusivity carboxyl modified particles ↓ [131] Ileal linear movements e coli through intestinal mucus ↓ [131] Ileal movement speed e coli through intestinal mucus ↓ [131] Ileal sialic acid content mucus ↓ [131] Ileal mucus structure (confocal imaging/SEM) changed [131] |
| Lactadherin | Intestinal permeability (ex-vivo FITC-dextran assay) ↓ [51] Ileal organization of cell junctions, anchoring of the TJ complexes and space between adjacent Cells improved (transmission electron microscropy) [51] Ileal claudin-3 (mRNA) ↓[51] Ileal JAM-A (mRNA) ↓ [51] Ileal claudin-3 (protein) ↓ [51] Ileal JAM-A (protein) ↓ [51] Ileal E-cadherin (protein) ↓ [51] Ileal claudin-3 distribution towards cell membranes along crypt-villus junction (normalization) [51] Ileal occludin distribution towards cell membranes along villus (normalization) [51] Ileal E-cadherin distribution towards cell membranes of villus and basolateral region of crypt cells [51] Ileal JAM-A distribution = [51] |
| Hormone/growth factor/vitamin- based interventions | |
| EGF | Paracellular intestinal permeability (functional orogastric [3H]lactulose assay) ↓ [137] Transcellular intestinal permeability (functional orogastric [3H]rhamnose assay) = [137] Ileal occludin (mRNA) ↓ [137] Jejunal and ileal claudin-3 (mRNA) ↓ [137] Ileal occludin (protein) ↓ [137] Jejunal and ileal claudin-3 (protein) ↓ [137] Ileal occludin distribution towards apical and basolateral membrane of crypt-villus axis [137] Ileal claudin-3 distribution towards apical and basolateral membrane of crypt-villus axis [137] Ileal number of goblet cells (Muc2 protein) ↑ [137] Ileal mucus layer on top villi tips ↑ [137] Ileal Muc2 (mRNA) ↑ [137] Jejunal and ileal Math1 (mRNA) ↑ [137] Ileal goblet cell phenotype normalized (scanning electron microscopy) [137] |
| Recombinant EGF from soybean extract | Ileal ZO-1 more sharply expressed, better incorporation in TJ (IF) [104] Ileal JAM-A more sharply expressed, better incorporation in TJ (IF) [104] |
| HB-EGF | Intestinal permeability (functional orogastric FITC-dextran assay) ↓ [58,61,105,175] Ileal Muc2 (mRNA) ↑ [37] Jejunal goblet cell number (AB/PAS) ↑ [176] Ileal bacterial adherence to intestinal villi ↓ (scanning electron microscope) [59] |
| IGF1 | Ileal secretory IgA (protein) ↑ [65] Ileal Muc2 (protein) ↑ [65] |
| EPO | Intestinal permeability (functional orogastric FITC-dextran assay) ↓ [66] Intestinal ZO-1 loss from TJ intact villi (protein) ↓ [66] Intestinal caudin-1 (protein) = [66] Intestinal caudin-3 (protein) = [66] Intestinal E-cadherin (protein) = [66] Intestinal β-catenin (protein) = [66] Intestinal p-Akt (protein) ↑ [66] |
| Vitamin A | Intestinal claudin-1 (protein) ↑ [111] Intestinal occludin (protein) ↑ [111] Intestinal ZO-1 (protein) ↑ [111] |
| Probiotic interventions | |
| Bifidobacterium mixture | Ileal β defensin (protein) ↑ [115] Ileal β defensin (mRNA) ↑ [115] |
| Bifidobacterium infantis | Endotoxemia (plasma) ↓ [75] Intestinal permeability (functional orogastric FITC-dextran assay) ↓ [116] Small intestinal internalization of claudin-4 in enterocyte cytoplasm (protein) ↓ [116] Small intestinal claudin-4 expression in TJ complex (protein) ↑ [116] Small intestinal occludin expression in TJ complex (protein) ↑ [116] Small intestinal co-fractioning of claudins-2 and -4 and caveolin 1 (protein) ↑ [116] intestinal permeability (functional orogastric FITC-dextran assay) ↓ [116] |
| Bifidobacterium bifidum OLB6378 | Ileal occludin (protein) ↓ [73] Ileal claudin-3 (protein) ↓ [73] Ileal occludin distribution towards crypts (normalization) [73] Ileal claudin-3 distribution towards crypts and cell membrane (normalization) [73] Ileal α-catenin (protein) = [73] Ileal β-catenin (protein) = [73] Ileal e-cadherin (protein) = [73] Ileal α-catenin distribution towards complete villus length and cell membrane (normalization) [73] Ileal β-catenin distribution towards complete villus length except for villi tips and cell membrane (normalization) [73] Ileal e-cadherin distribution towards crypts and cell membrane (normalization) [73] Ileal muc2 (mRNA) ↓ [73] Ileal Muc3 (mRNA) ↓ [73] Ileal TFF3 (mRNA) = [73] Ileal number of goblet cells (Muc2 protein) = [73] Ileal number of TFF3 positive cells (TFF3 protein) ↓ [73] |
| Bifidobacterium breve M-16V | Ileal ZO-1 (mRNA) ↓ [117] Ileal claudin-1 (mRNa) ↓ [117] Ileal occludin (mRNA) ↓ [117] |
| Bacteroides fragilis strain ZY-312 | Intestinal permeability (functional orogastric FITC-dextran assay) ↓ [113] Intestinal ZO-1 (protein) ↑ [113] Intestinal secretory IgA (protein) ↑ [113] |
| Lactobacillus reuteri DSM 20016 | Intestinal permeability (functional orogastric FITC-dextran assay) = [71,72] |
| Lactobacillus reuteri biofilm on unloaded microspheres | Intestinal permeability (functional orogastric FITC-dextran assay) ↓ [71] |
| Lactobacillus reuteri biofilm on MRS loaded microspheres | Intestinal permeability (functional orogastric FITC-dextran assay) ↓ [71] |
| Lactobacillus reuteri biofilm on sucrose loaded microspheres | Intestinal permeability (functional orogastric FITC-dextran assay) ↓ [72] |
| Lactobacillus reuteri biofilm on maltose loaded microspheres | Intestinal permeability (functional orogastric FITC-dextran assay) ↓ [72] |
| Lactobacillus rhamnosus GG | Colonic mucosal infiltration of bacteria (EUB338 staining) ↓ [161] |
| Other interventions | |
| Bovine milk exosomes | Distal ileal number of goblet cells (Muc2 protein) ↑ [118] Distal ileal number of goblet cells (AB-PAS) ↑ [118] Distal ileal number of GRP93 positive cells (protein) ↑ [118] |
| Human breast milk exosomes | Distal ileal number of goblet cells (Muc2 protein) ↑ [119] Distal ileal Muc2 (mRNA) ↑ [119] |
| Berberine | Distal ileal Muc2 (protein) ↑ [79] Distal ileal secretory IgA (protein) ↑ [79] |
| Human β-defensin-3 | Terminal ileal ZO-1 (protein) ↑ [81] |
3.8. NEC Pathophysiology: Vascular Dysfunction, Hypoxia-Ischemia and Free Radical Formation
3.9. Enteral Feeding and Vascular Dysfunction, Hypoxia-Ischemia and Free Radical Formation in Animal Models of NEC
3.9.1. Fat-Based Feeding Interventions
3.9.2. Carbohydrate or Sugar-Based Feeding Interventions
3.9.3. Protein or Amino Acid-Based Feeding Interventions
3.9.4. Hormone or Growth Factor-Based Feeding Interventions
3.9.5. Probiotic Feeding Interventions
3.9.6. Other Enteral Feeding Interventions
| Enteral Feeding Intervention | Effect on Vascular Dysfunction, Hypoxia and Free Radical Formation (Compared to NEC Protocol Exposure without Feeding Intervention) |
|---|---|
| Fat-based interventions | |
| PUFA | Intestinal iNOS (mRNA) = [45] |
| Pre-digested fat (less long chain triacylglycerol, not dependent on intestinal lipases | Ileal ROS accumulation ↓ (DHE staining) [85] Ileal MDA (protein) ↓ [85] |
| Very low-fat diet | Ileal ROS accumulation ↓ (DHE staining) [85] Ileal MDA (protein) ↓ [85] |
| MFGM | Ileal iNOS (mRNA) ↓ [48] Intestinal MDA (protein) ↓ [48] Intestinal SOD (protein) ↑ [48] |
| Carbohydrate/sugar-based interventions | |
| 2′-FL | Mesenteric blood flow ↑ (mesenteric micro-angiography) (eNOS dependent) [91] Intestinal iNOS (mRNA) ↓ [91] Small intestinal free nitrogen species, 3-nitrotyrosine (protein) ↓ [92] |
| 6′-SL | Small intestinal free nitrogen species, 3-nitrotyrosine (protein) ↓ [92] |
| 2′-FL + 6′-SL | Small intestinal free nitrogen species, 3-nitrotyrosine (protein) ↓ [92] |
| GOS/FOS | Terminal ileum SOD-1 (mRNA) ↑ [135] Terminal ileum SOD-3 (mRNA) ↑ [136] Terminal ileum GSH-Px-1 ↑ [135] Terminal ileum GSH-Px-7 ↑ [135] Terminal ileum CAT ↑ [135] |
| Protein/amino acid-based interventions | |
| L-Arginine | Intestinal thiobarbituric acid reactive substances ↓ [99] Intestinal SOD (protein) = [99] Intestinal CAT (protein) = [99] Intestinal nitrate (stable metabolite of NO) ↑ (NS) [99] Intestinal hypoxia ↓ (pimonidazole) [98] Postprandial arterial diameter intestinal microcirculation ↑ [98] |
| L-Carnitine | Intestinal thiobarbituric acid reactive substances ↓ [99] Intestinal SOD (protein) = [99] Intestinal CAT (protein) = [99] |
| Glutamine | Terminal ileal NO production = [96] |
| IAP | Ileal iNOS (MMA) ↓ [160] Ileal free nitrogen species, 3-nitrotyrosine (protein) ↓ [160] |
| N-Acetylcysteine | Ileal ROS accumulation ↓ (DHE staining) [85] Ileal MDA (protein) ↓ [85] |
| Hormone/growth factor/Vitamin-based interventions | |
| Recombinant EGF from soybean extract | ileal iNOS (mRNA) ↓ [104] |
| HB-EGF | Villus microvascular blood flow ↑ (angiography) [106] Villus microvascular structure preserved (angiography, scanning electron microscopy) [106] Submucosal intestinal blood flow ↑ (angiography) [106] |
| Relaxin | Ileal blood flow ↑ (laser Doppler flowmetry) [107] |
| Vitamin D | Intestinal MDA (protein) ↓ [112] Intestinal GSH-Px (protein) ↑ [112] |
| Probiotic interventions | |
| Bacteroides fragilis strain ZY-312 | Intestinal iNOS (protein) ↓ [113] |
| Lactobacillus rhamnosus HN001 (alive) | Terminal ileal iNOS (mRNA) ↓ (TLR9 dependent) [39] |
| Lactobacillus rhamnosus HN001 (dead, UV-radiated) | Terminal ileal iNOS (mRNA) ↓ [39] |
| Lactobacillus rhamnosus HN001 isolated microbial DNA | Terminal ileal iNOS (mRNA) ↓ [39] |
| CpG-DNA | Terminal ileal iNOS (mRNA) ↓ [39] |
| Lactobacillus reuteri DSM 17938 | Terminal ileal SOD activity (U/mg protein) ↑ [70] Terminal ileal SOD inhibition rate (%) ↑ [70] Terminal ileal GSSG concentration (protein) ↓ [70] Terminal ileal GSH concentration (protein) ↑ [70] Terminal ileal GSSG/GSH ratio (protein) ↓ [70] Terminal ileal MDA concentration (protein) ↓ [70] |
| Saccharomyces Boulardii | Terminal ileal SOD-1 (mRNA) ↑ [135] Terminal ileal SOD-3 (mRNA) ↑ [136] Terminal ileal GSH-Px-1 ↑ [135] Terminal ileal GSH-Px-3 ↑ [135] Terminal ileal GSH-Px-4 ↑ [135] Terminal ileal GSH-Px-7 ↑ [135] Terminal ileal CAT ↑ [135] |
| Other interventions | |
| Ginger | Intestinal SOD (protein) ↑ [121] Intestinal GSH-Px (protein) ↑ [121] Intestinal MDA (protein) ↓ [121] Intestinal XO (protein) ↓ [121] |
| Sesamol | Intestinal SOD (protein) ↑ [125] Intestinal GSH-Px (protein) ↑ (NS) [125] Intestinal MDA (protein) ↓ [125] |
| Fennel seed extracts | Intestinal total oxidant status (µmol H2O2 equivalent/g protein) ↓ [122] Intestinal oxidative stress index (total oxidant status/total antioxidant status) ↓ [122] intestinal advanced oxidation protein products (ng/mg protein) ↓ [122] intestinal lipid hydroperoxide (nmol/L) ↓ [122] intestinal 8-hydroxydeoxyguanosine (8-OhdG, ng/mL) ↓ [122] intestinal total antioxidant status (mmol Trolox equivalent/g protein) ↑ [122] |
| Amniotic fluid | intestinal iNOS (mRNA) ↓ [63] distal small intestinal/terminal ileum iNOS (mRNA) ↓ [40,123] terminal ileum iNOS (protein) ↓ [40] |
| Berberine | ileal iNOS (mRNA) ↓ [79] |
| Astragaloside IV | distal ileum GSH (protein) ↑ [126] distal ileum SOD (protein) ↑ [126] distal ileum MDA (protein) ↓ [126] |
| Resveratrol | ileum iNOS (protein) ↓ [127] |
3.10. NEC Pathophysiology: Intestinal Epithelial Cell Death and Proliferation
3.11. Enteral Feeding and Intestinal Epithelial Cell Death and Proliferation in Animal Models of NEC
3.11.1. Fat-Based Feeding Interventions
3.11.2. Carbohydrate or Sugar Based Feeding Interventions
3.11.3. Protein or Amino Acid-Based Feeding Interventions
3.11.4. Hormone, Growth Factor or Vitamin-Based Feeding Interventions
3.11.5. Probiotic Feeding Interventions
3.11.6. Other Enteral Feeding Interventions
| Enteral Feeding Intervention | Effect on Intestinal Epithelial Cell Death and Proliferation (Compared to NEC Protocol Exposure without Feeding Intervention) |
|---|---|
| Fat-based interventions | |
| Fish oil (rich in n-3 PUFA) | Small intestinal BiP (protein) ↓ [83] Small intestinal caspase 12 (protein) ↓ [83] |
| PUFA | Intestinal apoptosis (TUNEL) = [45] |
| MPL | Small intestinal apoptosis (TUNEL) ↓ [84] Small intestinal Bax (protein) ↓ [84] Small intestinal Bcl-2 (protein) ↑ [84] Small intestinal caspase 9 (protein) ↓ [84] Small intestinal caspase 3 protein) ↓ [84] |
| Pomegranate seed oil | Mean ileal villus length ↑ [47] Ileal epithelial cell proliferation (PCNA) ↑ [47] |
| Carbohydrate/sugar-based interventions | |
| HMO | (Terminal) ileal Ki67-positive cells ↑ [49,86,88] Ileal Sox9-positive cells ↑ [49] Terminal ileal TUNEL (protein) ↓ [86] Ileal cleaved caspase 3 (protein) ↓ [88] Ileal HIF1α (protein) ↓ [88] |
| Mixture of four HMOs | Small intestinal PCNA (mRNA) = [134] |
| 2′-FL | Small intestinal apoptosis (TUNEL) ↓ [92] |
| 6′-SL | Small intestinal apoptosis (TUNEL) ↓ [92] |
| 2′-FL + 6′-SL | Small intestinal apoptosis (TUNEL) ↓ [92] |
| Protein/amino acid-based interventions | |
| Lactoferrin | Proximal intestinal villus length/crypt depth ratio ↓ [36] Middle intestinal Bax-to-Bcl-2 ratio (protein) ↑ [36] Middle intestinal HIF-1α (protein) ↑ [36] Middle intestinal pro-caspase 3 (protein) = [36] Middle intestinal CC3 (protein) = [36] Distal ileal Ki67 (protein) ↑ [101] Distal ileal β-catenin (protein) ↑ [101] Distal ileal LGR5 (mRNA) ↑ [101] |
| L-Glutamine/glutamine | Small intestinal apoptosis (TUNEL) ↓ [84] Small intestinal Bax (protein) ↓ [84] Small intestinal Bcl-2 (protein) ↑ [84] Small intestinal caspase 9 (protein) ↓ [84] Small intestinal caspase 3 (protein) ↓ [84] Jejunum, ileum and colon caspase 3 (protein) ↓ [97] |
| Hormone/growth factor/vitamin-based interventions | |
| EGF | Ileal villus length ↑ [193] Ileal epithelial proliferation (PCNA) = [193] Ileal Bax (mRNA) ↓ [193] Ileal Bax (protein) ↓ [193] Ileal Bcl-2 (mRNA) ↑ [193] Ileal Bcl-2 (protein) ↑ [193] Ileal Bax-to-Bcl-2 ratio (mRNA) ↓ [193] Ileal CC3 villus tips (protein) ↓ [193] Ileal Bax-to-Bcl-2 ratio (protein) ↓ [37,193] Ileal Beclin 1 (protein) ↓ [55] Ileal LC3II (protein) ↓ [55] Ileal LC3II/LCRI ratio (protein) ↓ [55] Ileal p62 (protein) ↑ [55] Ileal autophagy signs (autophagosomes, autophagolysosomes, vacuoles) (transmission electron microscopy) ↓ [55] |
| HB-EGF | Intestinal TUNEL score (protein) ↓ [60] Intestinal CC3 score (protein) ↓ [60] Ileal cell migration (BrdU-positive cells) ↑ [59,61] Small intestinal integrin subunit α5 (mRNA) ↑ [61] Small intestinal integrin subunit β1 (mRNA) ↑ [61] Small intestinal integrin subunit α1 (mRNA) = [61] Small intestinal integrin subunit α2 (mRNA) = [61] Small intestinal integrin subunit α3 (mRNA) = [61] Small intestinal integrin subunit α6 (mRNA) = [61] Small intestinal integrin subunit α5 (protein) ↑ [61] Small intestinal integrin subunit β1 (protein) ↑ [61] Ileal epithelial cell proliferation (number of BrdU-positive cells) ↑ [59] Ileal Bax-to-Bcl-2 ratio (protein) = [37] Jejunal crypt epithelial cell proliferation (PCNA) ↑ [176] Jejunal number of enterocytes per villus ↑ [176] Jejunal number of LGR5+/prominin-1+ stem cells ↑ [176] |
| EPO | Ileal Beclin 1 immunoreactivity ↓ [194] Ileal LC3 immunoreactivity ↓ [194] Small intestinal Beclin 1 (protein) ↓ [194] Small intestinal LC3II (protein) ↓ [194] Ileal p62 immunoreactivity ↑ [194] Small intestinal p62 (protein) ↑ [194] Ileal CC3 immunoreactivity ↓ [194] Small intestinal Bcl-2 (protein) ↑ [194] |
| ATRA | Terminal ileal apoptosis intestinal crypts (TUNEL) ↓ [109] Terminal ileal proliferation crypt intestinal epithelial cells (Ki67, BrdU) ↑ [109] |
| Vitamin D | Intestinal cleaved caspase 3 (protein) ↓ [112] Intestinal Bcl-2 (protein) ↑ [112] Intestinal Ki67 (protein) ↑ [112] |
| Probiotic interventions | |
| Bacteroides fragilis strain ZY-312 | Intestinal CC3 (protein) ↓ [113] Intestinal Bax (protein) ↓ [113] Intestinal Bcl-2 (protein) ↓ [113] Intestinal caspase 1 (protein) ↓ [113] Intestinal ASC (protein) ↓ [113] Intestinal NLRP3 (protein) ↓ [113] Intestinal IL1β (protein) ↓ [113] Intestinal gasdermin-D (protein) ↓ [113] |
| Lactobacillus rhamnosus GG | Ileal CC3 (protein) ↓ [161] Ileal apoptotic index (TUNEL) ↓ [161] |
| Bifidobacterium bifidum OLB6378 | Ileal Bax (protein) ↓ [74] Ileal Bcl-w (protein) ↑ [74] Ileal Bax/Bcl-w ratio ↓ [74] Ileal CC3-positive cell number ↓ [74] |
| Bifidobacterium breve M-16V | Ileal caspase 3 (mRNA) ↓ [117] |
| Other interventions | |
| Amniotic fluid | Terminal ileal PCNA immunoreactivity ↑ [40] |
| Ginger | Intestinal TUNEL-positive cell number ↓ [121] Intestinal C3-positive cell number ↓ [121] Intestinal C8-positive cell number ↓ [121] Intestinal caspase 3 (protein) ↓ [121] |
| Fennel seed extracts | Terminal ileal C3-positive cells number ↓ [122] Terminal ileal C8-positive cells number ↓ [122] Terminal ileal C9-positive cells number ↓ [122] Intestinal C3 concentration (protein) ↓ [122] |
| Preterm human breast milk exosomes | Intestinal enterocyte proliferation (BrdU) ↑ [120] |
| Sesamol | Intestinal Bcl-2-positive cell number ↓ [125] Intestinal caspase-3 positive cell number ↓ [125] |
| Curcumin | Intestinal caspase 1 (protein) ↓ [124] Intestinal NLRP3 (protein) ↓ [124] Intestinal caspase 1 (mRNA) ↓ [124] Intestinal NLRP3 (mRNA) ↓ [124] |
3.12. NEC Pathophysiology: Microbial Dysbiosis
3.13. Enteral Feeding and Microbial Dysbiosis in Animal Models of NEC
3.13.1. Fat-Based Feeding Interventions
3.13.2. Carbohydrate or Sugar-Based Feeding Interventions
3.13.3. Protein or Amino Acid-Based Feeding Interventions
3.13.4. Hormone, Growth Factor or Vitamin-Based Feeding Interventions
3.13.5. Probiotic Feeding Interventions
3.13.6. Other Enteral Feeding Interventions
| Enteral Feeding Intervention | Effect on Microbial Dysbiosis (Compared to NEC Protocol Exposure without Feeding Intervention) |
|---|---|
| Fat-based interventions | |
| BCFA | Cecal Bacillaeceae (family) abundance ↑ [46] Cecal Pseusomonadaceae (family) abundance ↑ [46] Cecal Bacillus subtilis (species) abundance ↑ [46] Cecal Pseudomonas aeruginosa (species) abundance ↑ [46] |
| Carbohydrate/sugar-based interventions | |
| 2′-FL | Fecal Enterobacteriaceae (family) abundance = [91] Fecal Lactobacillaceae (family) abundance = [91] Fecal microbiota β-diversity ↓ [91] Cecal microbial colonization density (FISH) = [133] α-Diversity cecal tissue = [133] α-Diversity cecal content = [133] Proportion Enterococcus (genus) in cecal content ↑ [133] |
| Mixture of four HMOs | Colonic microbial diversity (number of OTU per sample) = [134] Colonic microbial clustering = [134] Colonic relative abundance genera OTU = [202] Colonic number of Fusobaceterium (genus) on individual level ↓ [134] |
| Mixture of >25 HMO components | Colonic relative abundance genera OTU = [202] |
| Protein/amino acid-based interventions | |
| OPN | Colonic microbial α diversity = [129] Colonic microbial abundance of genera = [129] |
| CGMP | Colonic microbial α diversity = [129] Colonic microbial abundance of genera = [129] |
| Vitamin-based interventions | |
| Vitamin A | Fecal Proteobacteria (phylum) abundance ↓ [111] Fecal Escherichia-Shigella (genus) abundance ↓ [111] Fecal Lactobacillus (genus) abundance ↓ [111] Fecal Acinetobacter (genus) abundance ↓ [111] Fecal Gemella (genus) abundance ↓ [111] Fecal Bacteroidetes (phylum) abundance ↑ [111] Fecal Bacteroides (genus) abundance ↑ [111] Fecal Romboutsia (genus) abundance ↑ [111] Fecal Parabacteroides (genus) abundance ↑ [111] |
| Probiotic interventions | |
| Bifidobacterium infantis-longum strain CUETM 89-215 | Cecal Clostridium perfringens (species) count ↓ [130] Cecal Clostridium difficile (species) count ↓ [130] |
| Bacteroides fragilis strain ZY-312 | Fecal number of OTU ↑ [113] Fecal relative abundance Bacteroidetes (phylum) ↑ [113] Fecal relative abundance Proteobacteria (phylum) ↓ [113] |
| Lactobacillus reuteri DSM 20016 | Fecal Lactobacillus (genus) abundance ↑ [72] Fecal Enterobacter (genus) abundance ↓ [72] |
| Lactobacillus reuteri biofilm on maltose loaded microspheres | Shift of fecal microbiome towards breastfed controls (16S sRNA sequencing) [72] Fecal Lactobacillus (genus) abundance ↑ [72] Fecal Enterobacter (genus) abundance ↓ [72] |
| Probiotic mixture (Bifidobacterium animalis DSM15954, Lactobacillus acidophilus DSM13241, Lactobacillus casei ATCC55544, Lactobacillus pentosus DSM14025 and Lactobacillus plantarum DSM13367 | Distal small intestinal general colonization pattern (T-RFLP analysis) changed [77] Colonic general colonization pattern (T-RFLP analysis) changed [77] Distal small intestinal colonization density of Clostridium perfringens ↓ [77] Distal small intestinal homogenate relative proportion Clostridium (genus) ↓ [77] Colonic content relative proportion Clostridium (genus) ↓ [77] Distal small intestinal homogenate relative proportion Enterococcus (genus) ↓ [77] Stomach content relative proportion Enterococcus (genus) ↓ [77] Colon content relative proportion lactic acid bacteria ↑ [77] Stomach content relative proportion lactic acid bacteria ↑ [77] Distal small intestinal homogenate relative proportion Lactobacillus (genus) ↑ [77] Stomach content relative proportion Lactobacillus (genus) ↑ [77] Colon content relative proportion total anaerobes ↑ [77] |
| Other interventions | |
| Amniotic fluid | Distal small intestinal bacterial colonization (general eubacterial probe) ↓ [123] Colonic bacterial colonization normalized (PCA of T-RFLP analysis) [123] |
3.14. NEC Pathophysiology: Disturbed Digestion and Absorption
3.15. Enteral Feeding and Disturbed Digestion and Absorption in Animal Models of NEC
3.15.1. Carbohydrate or Sugar-Based Feeding Interventions
3.15.2. Protein or Amino Acid-Based Feeding Interventions
3.15.3. Probiotic Feeding Interventions
3.15.4. Other Enteral Feeding Interventions
| Enteral Feeding Intervention | Effect on Digestion and Absorption (Compared to NEC Protocol Exposure without Feeding Intervention) |
|---|---|
| Carbohydrate/sugar-based interventions | |
| Gangliosides | Intestinal enzyme activity small intestine = [100] Intestinal hexose absorption (galactose lactose absorption test) = [100] |
| SL | Intestinal enzyme activity small intestine = [100] Intestinal hexose absorption (galactose lactose absorption test) = [100] |
| HMO mixture of four components | Intestinal hexose absorption (galactose lactose absorption test) = [134] Intestinal enzyme activity small intestine = [134] Colonic butyric acid (protein) ↑ [134] Small intestinal sucrase (mRNA) = [134] Small intestinal lactase (mRNA) = [134] Small intestinal alkaline phosphatase (mRNA) = [134] small intestinal SGLT1 (mRNA) = [134] |
| HMO mixture >25 components | Intestinal hexose absorption (galactose lactose absorption test) = [134] Distal small intestinal lactase enzyme activity ↑ [134] Distal small intestinal aminopeptidase A enzyme activity ↑ [134] Distal small intestinal aminopeptidase N enzyme activity ↑ [134] Distal small intestinal dipeptidyl peptidase IV enzyme activity ↑ [134] |
| 2′-FL | Galactose absorptive capacity = (galactose mannitol absorption test) [133] Proximal/middle/distal small intestinal enzyme activity (sucrose, maltase, lactase, ApN, ApA, DPPIV) = [133] Colon small intestinal enzyme activity (sucrose, maltase, lactase, ApN, ApA, DPPIV) = [133] |
| Protein/amino acid-based interventions | |
| OPN | Intestinal enzyme activity small intestine = [100,129] Intestinal hexose absorption (galactose lactose absorption test) = [100,129] |
| bovine lactoferrin | Small intestine enzyme activity = [35] Intestinal hexose absorption (galactose lactose absorption test) = [35] |
| CGMP | Middle small intestinal lactase enzyme activity ↑ [129] Proximal small intestinal lactase enzyme activity = [129] Distal small intestinal lactase enzyme activity = [129] Intestinal hexose absorption (galactose lactose absorption test) = [129] |
| Probiotic interventions | |
| Probiotic mixture (Bifidobacterium animalis DSM15954, Lactobacillus acidophilus DSM13241, Lactobacillus casei ATCC55544, Lactobacillus pentosus DSM14025 and Lactobacillus plantarum DSM13367) | Distal small intestinal lactase enzyme activity = [77] Distal small intestinal maltase enzyme activity = [77] Distal small intestinal ApA enzyme activity ↑ [77] Distal small intestinal ApN enzyme activity ↑ [77] |
| Other interventions | |
| Amniotic fluid | Proximal small intestine maltase enzyme activity ↑ [123] Middle small intestinal maltase enzyme activity ↑ [123] |
3.16. NEC Pathophysiology: Enteric Nervous System Alterations
3.17. Enteral Feeding and Enteric Nervous System Alterations in Animal Models of NEC
3.18. Enteral Feeding Interventions Affecting NEC Incidence and Severity in Human Studies
3.18.1. Fat-Based Feeding Interventions
3.18.2. Carbohydrate or Sugar-Based Feeding Interventions
3.18.3. Protein or Amino Acid-Based Feeding Interventions
3.18.4. Hormone, Growth Factor or Vitamin-Based Feeding Interventions
3.18.5. Probiotic Feeding Interventions
3.18.6. Other Feeding Interventions
| Enteral Feeding Intervention | Effect on NEC Incidence | ||||
|---|---|---|---|---|---|
| Intervention | Study, N | Observed Relative Effect | Anticipated Absolute Effects | Certainty of Evidence (GRADE) | |
| Risk with No Enteral Feeding Intervention | Risk with Enteral Feeding Intervention (95% CI) | ||||
| Fat-Based Interventions | |||||
| n-3 PUFA | Meta-analysis, 11 RCTs, N = 1753 neonates | RR 1.17 (95% CI 0.77–1.79) (incidence all neonates) [223] | 28 per 1000 | 5 more per 1000 more (from 6 less per 1000 to 50 more per 1000) | Low ⊕⊕⊝⊝ (risk of bias, imprecision) |
| RR 0.50 (95% CI 0.23–1.10) (incidence neonates ≤32 weeks) [223] | 58 per 1000 | 29 less per 1000 (from 45 less per 1000 to 6 more per 1000) | Low ⊕⊕⊝⊝ (risk of bias, imprecision) | ||
| DHA | RCT, N = 1205 neonates | RR1.16 (95% CI 0.79–1.69) (incidence) [224] | 71 per 1000 | 11 more per 1000 (from 15 less per 1000 to 49 more per 1000) | Low ⊕⊕⊝⊝ (imprecision) |
| Carbohydrate or sugar-based interventions | |||||
| Prebiotics | Meta-analysis, 6 RCTs, N = 737 neonates | RR 0.79 (95% CI 0.44–1.44) (incidence) [225] | 112 per 1000 | 24 less per 1000 (from 63 less per 1000 to 49 more per 1000) | Low ⊕⊕⊝⊝ (inconsistency, imprecision) |
| Protein or amino acid-based interventions | |||||
| Lactoferrin | Meta-analysis, 7 RCTs, N = 4874 neonates | RR 0.90 (95% CI 0.69–1.17) (incidence stage II or III NEC) [226] | 47 per 1000 | 5 per 1000 less (from 15 per 1000 less to 8 per 1000 more) | Low ⊕⊕⊝⊝ (risk of bias, imprecision) |
| Arginine | Meta-analysis, 3 RCTs, N = 285 neonates | RR 0.38 (95% CI 0.23–0.64) (incidence) [227] | 303 per 1000 | 188 per 1000 less (from 233 less per 1000 to 109 less per 1000) | Moderate ⊕⊕⊕⊝ (imprecision) |
| RR 0.18 (95% CI 0.03–1.00) (death due to NEC) [227] | 55 per 1000 | 45 less per 1000 (from 53 less per 1000 to 0 less per 1000 | Low ⊕⊕⊝⊝ (imprecision) | ||
| Glutamine | Meta-analysis, 7 RCTs, N = 1172 neonates | RR 0.73 (95% CI 0.49–1.08) (incidence) [228] | 90 per 1000 | 24 less per 1000 (from 46 less per 1000 to 8 more per 1000) | Low ⊕⊕⊝⊝ (imprecision, publication bias) |
| IgG or IgG + IgA | meta-analysis, 3 clinical trials, N = 2095 neonates | RR 0.84 (95% CI 0.57–1.25) (incidence) [229] | 55 per 1000 | 9 less per 1000 (from 24 less per 1000 to 14 more per 1000) | Low ⊕⊕⊝⊝ (risk of bias) |
| RR 0.21 (95% CI 0.02–1.75) (NEC related surgery) [229] | 25 per 1000 | 20 less per 1000 (from 25 less per 1000 to 19 more per 1000) | Very low ⊕⊝⊝⊝ (risk of bias, imprecision) | ||
| RR 1.10 (95% CI 0.47–2.59) (death due to NEC) [229] | 10 per 1000 | 1 per 1000 more (from 5 less per 1000 to 16 more per 1000) | Very low ⊕⊝⊝⊝ (risk of bias, imprecision) | ||
| Hormone/growth factor/vitamin-based interventions | |||||
| EPO | Meta-analysis, 2 RCTs, N = 110 neonates | RR 0.62 (95% CI 0.15–2.59) (incidence stage II or III NEC) (NS) [230] | 61 per 1000 | 24 less per 1000 (from 52 less per 1000 to 97 more per 1000) | Very low ⊕⊝⊝⊝ (inconsistency imprecision) |
| RCT, N = 120 neonates | 2.8pp increase (11.1% control group, 13.9% EPO group (NS) (incidence) [232] | 111 per 1000 | 28 less per 1000 (no CI reported) | Very low ⊕⊝⊝⊝ (risk of bias, inconsistency imprecision) | |
| RCT, N = 100 neonates | 2 pp reduction (8% control group, 6% EPO group) (NS) (incidence) [231] | 80 per 1000 | 20 less per 1000 (no CI reported) | ||
| EPO + G-CSF | RCT, N = 50 neonates | 10 pp reduction (10% control group, 0% EPO group) (NS) (incidence) [233] | 100 per 1000 | 100 per 1000 less (no CI reported) | Low ⊕⊕⊝⊝ (imprecision) |
| G-CSF | RCT, N = 50 neonates | 10 pp reduction (10% control group, 0% EPO group) (NS) (incidence) [233] | 100 per 1000 | 100 per 1000 less (no CI reported) | Low ⊕⊕⊝⊝ (imprecision) |
| G-CSF (in artificial amniotic fluid) | RCT, N = 100 neonates | 2 pp reduction (8% control group, 6% EPO group) (NS) (incidence) [231] | 80 per 1000 | 20 per 1000 less (no CI reported) | Very low ⊕⊝⊝⊝ (risk of bias, imprecision) |
| Vitamin A | RCT, N = 154 neonates | RR 1.14 (95% CI 0.66–1.66) (incidence) [234] | 91 per 1000 | 13 per 1000 more (from 31 per 1000 less to 60 per 1000 more) | Low ⊕⊕⊝⊝ (risk of bias, imprecision) |
| RCT, N = 262 neonates | RR 0.69 (95% CI 0.27–1.76) (incidence) [235] | 77 per 1000 | 24 per 1000 less (from 56 per 1000 less to 59 per 1000 more) | ||
| Probiotic interventions | |||||
| Probiotics | (network) meta-analysis, 56 RCTs, N = 12,738 neonates | ||||
| Lactobacillus spp. and Bifidobacterium spp. 11 RCTs, N = 1878 neonates | OR 0.35 (95% CI 0.20–0.59) (incidence stage II or III NEC) [238] | 63 per 1000 | 41 per 1000 less (from 51 per 1000 less to 26 per 1000 less) | Moderate ⊕⊕⊕⊝ (indirectness) | |
| Bifodobacterium animalis subsp. Lactis 5 RCTs, 628 neonates | OR 0.31 (95% CI 0.13–0.74) (incidence stage II or III NEC) [238] | 94 per 1000 | 44 per 1000 less (from 56 per 1000 less to 16 per 1000 less) | Moderate ⊕⊕⊕⊝ (imprecision) | |
| Lactobacillus reuteri 5 RCTs, N = 1388 neonates | OR 0.55 (95% CI 0.34–0.91) (incidence stage II or III NEC) [238] | 71 per 1000 | 28 per 1000 less (from 42 per 1000 less to 5 per 1000 less) | Low ⊕⊕⊝⊝ (imprecision, indirectness) | |
| Lactobacillus rhamnosus 5 RCTs, N = 839 neonates | OR 0.44 (95% CI 0.21–0.90) (incidence stage II or III NEC) [238] | 60 per 1000 | 35 per 1000 less (from 50 per 1000 less to 5 per 1000 less) | Moderate ⊕⊕⊕⊝ (imprecision) | |
| Combination of Lactobacillus ssp., Bifidobacterium spp. and Enterococcus ssp. 7 RCTs, N = 1950 neonates | OR 0.28 (95% CI 0.16–0.49) (incidence stage II or III NEC) [238] | 63 per 1000 | 46 per 1000 less (from 54 per 1000 less to 32 per 1000 less) | Low ⊕⊕⊝⊝ (risk of bias) | |
| Combination of Bacillus spp. and Enterococcus spp. 1 RCT, N = 355 neonates | OR 0.23 (95% 0.08–0.63) (incidence stage II or III NEC) [238] | 110 per 1000 | 49 per 1000 less (from 59 per 1000 less to 23 per 1000 less) | Moderate ⊕⊕⊕⊝ (imprecision) | |
| Network meta-analysis 43 RCTs, N = 10,651 neonates | |||||
| Bifidobacterium lactis Bb-12 or B-94 5 RCTs, N = 828 neonates | RR 0.25 (95% CI 0.10–0.56) (incidence) [239] | 100 per 10000 | 75 per 1000 less (from 90 per 1000 less to 44 per 1000 less | Moderate ⊕⊕⊕⊝ (imprecision) | |
| Lactobacillus reuteri ATCC55730 or DSM17938 4 RCTs, N = 1459 neonates | RR 0.43 (95% CI 0.16–0.98) (incidence) [239] | 92 per 1000 | 52 per 1000 less (from 77 per 1000 less to 2 per 1000 less) | Low ⊕⊕⊝⊝ (imprecision, indirectness) | |
| Lactobacillus rhamnosus GG 6 RCTs, N = 1507 neonates | RR 0.24 (95% CI 0.064–0.67) (incidence) [239] | 24 per 1000 | 18 per 1000 less (from 22 per 1000 less to 8 per 1000 less) | Low ⊕⊕⊝⊝ (imprecision, indirectness) | |
| Combination of Bifidobacterium bifidum, Bifidobacterium infantis, Bifidobacteirum longum and Lactobacillus acidophilus 2 RCTs, N = 247 neonates | RR 0.25 (95% CI 0.051–0.89) (incidence) [239] | 129 per 1000 | 97 per 1000 less (from 122 per 1000 less to 14 per 1000 less) | Low ⊕⊕⊝⊝ (imprecision) | |
| Combination of Bifidobacterium infantis Bb-02, Bifidobacterium lactis Bb-12 and Streptococcus thermophilus TH-4 2 RCTs, N = 1244 neonates | RR 0.29 (95% CI 0.073–0.78) (incidence) [239] | 54 per 1000 | 38 per 1000 less (from 50 per 1000 less to 12 per 1000 less) | Low ⊕⊕⊝⊝ (inconsistency, imprecision) | |
| Bifidobacterium longum 35624 and Lacobacillus rhamnosus GG 2 RCTs, N = 285 neonates | RR 0.18 (95% CI 0.020–0.89) (incidence) [239] | 42 per 1000 | 34 per 1000 less (from 41 per 1000 less to 5 per 1000 less) | Low ⊕⊕⊝⊝ (imprecision) | |
| Other interventions | |||||
| Carotenoids | RCT, N = 229 neonates | OR 0.34 (95% CI 0.07–1.66) (incidence stage II or III NEC) [241] | 52 per 1000 | 34 per 1000 less (from 48 per 1000 less to 60 per 1000 more) | Low ⊕⊕⊝⊝ (imprecision) |
| Mixture of probiotics, prebiotics and lactoferrin | RCT, N = 208 neonates | RR 0.16 (95% CI 0.03–0.77) (incidence) [242] | 106 per 1000 | 89 per 1000 less (from 103 per 1000 less to 24 per 1000 less) | Low ⊕⊕⊝⊝ (imprecision) |
| RR 0.56 (95% CI 0.47–0.67) (incidence stage II or III NEC) [242] | 38 per 1000 | 17 per 1000 less (from 20 per 1000 less to 13 per 1000 less) | Low ⊕⊕⊝⊝ (imprecision) | ||
3.19. Enteral Feeding Interventions Affecting Pathophysiological Mechanisms of NEC in Human Studies
3.19.1. Carbohydrate or Sugar Based Feeding Interventions
3.19.2. Protein or Amino Acid-Based Feeding Interventions
3.19.3. Probiotic Feeding Interventions
3.19.4. Other Feeding Interventions
| Enteral Feeding Intervention | Effect on Pathophysiological Mechanism (Compared to Placebo/No Intervention) |
|---|---|
| Carbohydrate or sugar-based interventions | |
| (SC) GOS + (LC) FOS | % Gastric slow wave propagation (electrogastrography) ↑ [243] Gastric half emptying time (echography) ↓ [243] Intestinal transit time (passage carmine red) ↑ [244] Stool pH ↓ [244] Stool viscosity ↑ [244] |
| SC GOS + LC FOS + AOS | Fecal IL8 = [245] Fecal calprotectin = [245] |
| Protein or amino acid-based interventions | |
| Lactoferrin | Whole blood CD4+ CD25high Foxp3+ Treg cell number ↑ [246] |
| L-Glutamine | Lactulose recovery in urine day 7 ↓ [247] Lactulose recovery in urine day 30 ↓ [247] Lactulose/mannitol recovery ratio in urine day 7 ↓ [247] Lactulose/mannitol recovery ratio in urine day 30 ↓ [247] |
| Probiotic interventions | |
| Lactobacillus reuteri | % Gastric slow wave propagation (electrogastrography) ↑ [243] Gastric half emptying time (echography) ↓ [243] |
| Bifidobacterium lactis | Lactulose/mannitol recovery ratio in urine day 30 ↓ [248] |
| Other interventions | |
| Mixture of probiotics, prebiotics and lactoferrin | Serum IL5 (protein) at 0, 14 and 28 days = [249] Serum IL10 (protein) at 0, 14 and 28 days (protein) = [249] Serum IL17 (protein) at 0, 14 and 28 days = [249] Serum IFNϒ (protein) 0, 14 days = [249] Serum IFNϒ (protein) 28 days ↑ [249] |
3.20. Interaction between Feeding Components and NEC
3.21. Enteral Feeding Strategies and NEC: Feeding Regimens, Fortifiers and More
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Neu, J.; Walker, W.A. Necrotizing enterocolitis. N. Engl. J. Med. 2011, 364, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Fitzgibbons, S.C.; Ching, Y.; Yu, D.; Carpenter, J.; Kenny, M.; Weldon, C.; Lillehei, C.; Valim, C.; Horbar, J.D.; Jaksic, T. Mortality of necrotizing enterocolitis expressed by birth weight categories. J. Pediatr. Surg. 2009, 44, 1072–1075. [Google Scholar] [CrossRef]
- Alsaied, A.; Islam, N.; Thalib, L. Global incidence of Necrotizing Enterocolitis: A systematic review and Meta-analysis. BMC Pediatr. 2020, 20, 344. [Google Scholar] [CrossRef] [PubMed]
- Han, S.M.; Hong, C.R.; Knell, J.; Edwards, E.M.; Morrow, K.A.; Soll, R.F.; Modi, B.P.; Horbar, J.D.; Jaksic, T. Trends in incidence and outcomes of necrotizing enterocolitis over the last 12 years: A multicenter cohort analysis. J. Pediatr. Surg. 2020, 55, 998–1001. [Google Scholar] [CrossRef] [PubMed]
- Heida, F.H.; Stolwijk, L.; Loos, M.H.; van den Ende, S.J.; Onland, W.; van den Dungen, F.A.; Kooi, E.M.; Bos, A.F.; Hulscher, J.B.; Bakx, R. Increased incidence of necrotizing enterocolitis in the Netherlands after implementation of the new Dutch guideline for active treatment in extremely preterm infants: Results from three academic referral centers. J. Pediatr. Surg. 2017, 52, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.W.; Stoll, B.J. Necrotising enterocolitis. Lancet 2006, 368, 1271–1283. [Google Scholar] [CrossRef]
- Blakely, M.L.; Lally, K.P.; McDonald, S.; Brown, R.L.; Barnhart, D.C.; Ricketts, R.R.; Thompson, W.R.; Scherer, L.R.; Klein, M.D.; Letton, R.W.; et al. Postoperative outcomes of extremely low birth-weight infants with necrotizing enterocolitis or isolated intestinal perforation: A prospective cohort study by the NICHD Neonatal Research Network. Ann. Surg. 2005, 241, 984–989. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.; Kamitsuka, M.; Clark, R.H.; Kelleher, A.S.; Spitzer, A.R. Etiologies of NICU deaths. Pediatrics 2015, 135, e59–e65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, C.R.; Fullerton, B.S.; Mercier, C.E.; Morrow, K.A.; Edwards, E.M.; Ferrelli, K.R.; Soll, R.F.; Modi, B.P.; Horbar, J.D.; Jaksic, T. Growth morbidity in extremely low birth weight survivors of necrotizing enterocolitis at discharge and two-year follow-up. J. Pediatr. Surg. 2018, 53, 1197–1202. [Google Scholar] [CrossRef]
- Mutanen, A.; Wales, P.W. Etiology and prognosis of pediatric short bowel syndrome. Semin. Pediatr. Surg. 2018, 27, 209–217. [Google Scholar] [CrossRef]
- Duggan, C.P.; Jaksic, T. Pediatric Intestinal Failure. N. Engl. J. Med. 2017, 377, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Schulzke, S.M.; Deshpande, G.C.; Patole, S.K. Neurodevelopmental outcomes of very low-birth-weight infants with necrotizing enterocolitis: A systematic review of observational studies. Arch. Pediatr. Adolesc. Med. 2007, 161, 583–590. [Google Scholar] [CrossRef] [Green Version]
- Mowitz, M.E.; Dukhovny, D.; Zupancic, J.A.F. The cost of necrotizing enterocolitis in premature infants. Semin. Fetal Neonatal Med. 2018, 23, 416–419. [Google Scholar] [CrossRef] [PubMed]
- Bisquera, J.A.; Cooper, T.R.; Berseth, C.L. Impact of necrotizing enterocolitis on length of stay and hospital charges in very low birth weight infants. Pediatrics 2002, 109, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Ganapathy, V.; Hay, J.W.; Kim, J.H.; Lee, M.L.; Rechtman, D.J. Long term healthcare costs of infants who survived neonatal necrotizing enterocolitis: A retrospective longitudinal study among infants enrolled in Texas Medicaid. BMC Pediatr. 2013, 13, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christian, V.J.; Polzin, E.; Welak, S. Nutrition Management of Necrotizing Enterocolitis. Nutr. Clin. Pr. 2018, 33, 476–482. [Google Scholar] [CrossRef]
- Vaks, Y.; Birnie, K.L.; Carmichael, S.L.; Hernandez-Boussard, T.; Benitz, W.E. Temporal Relationship of Onset of Necrotizing Enterocolitis and Introduction of Enteric Feedings and Powdered Milk Fortifier. Am. J. Perinatol. 2018, 35, 616–623. [Google Scholar] [CrossRef]
- Kirtsman, M.; Yoon, E.W.; Ojah, C.; Cieslak, Z.; Lee, S.K.; Shah, P.S. Nil-per-os days and necrotizing enterocolitis in extremely preterm infants. Am. J. Perinatol. 2015, 32, 785–794. [Google Scholar] [CrossRef]
- Thai, J.D.; Gregory, K.E. Bioactive Factors in Human Breast Milk Attenuate Intestinal Inflammation during Early Life. Nutrients 2020, 12, 581. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishna, K.P.; Hand, T.W. Influence of Maternal Milk on the Neonatal Intestinal Microbiome. Nutrients 2020, 12, 823. [Google Scholar] [CrossRef] [Green Version]
- Lucas, A.; Cole, T.J. Breast milk and neonatal necrotising enterocolitis. Lancet 1990, 336, 1519–1523. [Google Scholar] [CrossRef]
- Altobelli, E.; Angeletti, P.M.; Verrotti, A.; Petrocelli, R. The Impact of Human Milk on Necrotizing Enterocolitis: A Systematic Review and Meta-Analysis. Nutrients 2020, 12, 1322. [Google Scholar] [CrossRef]
- Quigley, M.; Embleton, N.D.; McGuire, W. Formula versus donor breast milk for feeding preterm or low birth weight infants. Cochrane Database Syst. Rev. 2019, 7, Cd002971. [Google Scholar] [CrossRef] [PubMed]
- Tarnow-Mordi, W.O.; Abdel-Latif, M.E.; Martin, A.; Pammi, M.; Robledo, K.; Manzoni, P.; Osborn, D.; Lui, K.; Keech, A.; Hague, W.; et al. The effect of lactoferrin supplementation on death or major morbidity in very low birthweight infants (LIFT): A multicentre, double-blind, randomised controlled trial. Lancet. Child Adolesc. Health 2020, 4, 444–454. [Google Scholar] [CrossRef]
- Cui, X.; Shi, Y.; Gao, S.; Xue, X.; Fu, J. Effects of Lactobacillus reuteri DSM 17938 in preterm infants: A double-blinded randomized controlled study. Ital. J. Pediatr. 2019, 45, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maamouri, G.; Zegheibizadeh, F.; Khatam, F.; Boskabadi, H. The Impact of Oral Glutamine Supplementation on Prevention of Nosocomial Infections in Preterm Infants. IJN Iran. J. Neonatol. 2016, 7, 19–24. [Google Scholar] [CrossRef]
- Gómez-Rodríguez, G.; Amador-Licona, N.; Daza-Benítez, L.; Barbosa-Sabanero, G.; Carballo-Magdaleno, D.; Aguilar-Padilla, R.; González-Ramirez, E. Single strain versus multispecies probiotic on necrotizing enterocolitis and faecal IgA levels in very low birth weight preterm neonates: A randomized clinical trial. Pediatr. Neonatol. 2019, 60, 564–569. [Google Scholar] [CrossRef] [Green Version]
- Ouzzani, M.; Hammady, H.; Fedorowicz, Z.; Elmagarmid, A. Rayyan-a web and mobile app for systematic reviews. Syst. Rev. 2016, 5, 210. [Google Scholar] [CrossRef] [Green Version]
- Hooijmans, C.R.; Rovers, M.M.; de Vries, R.B.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jadad, A.R.; Moore, R.A.; Carroll, D.; Jenkinson, C.; Reynolds, D.J.; Gavaghan, D.J.; McQuay, H.J. Assessing the quality of reports of randomized clinical trials: Is blinding necessary? Control. Clin. Trials 1996, 17, 1–12. [Google Scholar] [CrossRef]
- Shea, B.J.; Grimshaw, J.M.; Wells, G.A.; Boers, M.; Andersson, N.; Hamel, C.; Porter, A.C.; Tugwell, P.; Moher, D.; Bouter, L.M. Development of AMSTAR: A measurement tool to assess the methodological quality of systematic reviews. BMC Med. Res. Methodol. 2007, 7, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schünemann, H.; Brożek, J.; Guyatt, G.; Oxman, A. GRADE Handbook for Grading Quality of Evidence and Strength of Recommendations. Updated October 2013; The GRADE Working Group: Hamilton, ON, Canada, 2013. [Google Scholar]
- Wei, D.; Tang, K.; Wang, Q.; Estill, J.; Yao, L.; Wang, X.; Chen, Y.; Yang, K. The use of GRADE approach in systematic reviews of animal studies. J. Evid. Based Med. 2016, 9, 98–104. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.N.; Li, Y.; Sangild, P.T.; Bering, S.B.; Chatterton, D.E. Effects of bovine lactoferrin on the immature porcine intestine. Br. J. Nutr. 2014, 111, 321–331. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.N.; Jiang, P.; Stensballe, A.; Bendixen, E.; Sangild, P.T.; Chatterton, D.E. Bovine lactoferrin regulates cell survival, apoptosis and inflammation in intestinal epithelial cells and preterm pig intestine. J. Proteom. 2016, 139, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, B.; Khailova, L.; Clark, J.A.; Hosseini, D.M.; Arganbright, K.M.; Reynolds, C.A.; Halpern, M.D. Comparison of epidermal growth factor and heparin-binding epidermal growth factor-like growth factor for prevention of experimental necrotizing enterocolitis. J. Pediatr. Gastroenterol. Nutr. 2008, 47, 11–18. [Google Scholar] [CrossRef]
- Wu, R.Y.; Li, B.; Koike, Y.; Maattanen, P.; Miyake, H.; Cadete, M.; Johnson-Henry, K.C.; Botts, S.R.; Lee, C.; Abrahamsson, T.R.; et al. Human Milk Oligosaccharides Increase Mucin Expression in Experimental Necrotizing Enterocolitis. Mol. Nutr. Food Res. 2019, 63, e1800658. [Google Scholar] [CrossRef] [PubMed]
- Good, M.; Sodhi, C.P.; Ozolek, J.A.; Buck, R.H.; Goehring, K.C.; Thomas, D.L.; Vikram, A.; Bibby, K.; Morowitz, M.J.; Firek, B.; et al. Lactobacillus rhamnosus HN001 decreases the severity of necrotizing enterocolitis in neonatal mice and preterm piglets: Evidence in mice for a role of TLR9. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G1021–G1032. [Google Scholar] [CrossRef] [Green Version]
- Good, M.; Siggers, R.H.; Sodhi, C.P.; Afrazi, A.; Alkhudari, F.; Egan, C.E.; Neal, M.D.; Yazji, I.; Jia, H.; Lin, J.; et al. Amniotic fluid inhibits Toll-like receptor 4 signaling in the fetal and neonatal intestinal epithelium. Proc. Natl. Acad. Sci. USA 2012, 109, 11330–11335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radulescu, A.; Zorko, N.A.; Yu, X.; Besner, G.E. Preclinical neonatal rat studies of heparin-binding EGF-like growth factor in protection of the intestines from necrotizing enterocolitis. Pediatr. Res. 2009, 65, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Jantscher-Krenn, E.; Zherebtsov, M.; Nissan, C.; Goth, K.; Guner, Y.S.; Naidu, N.; Choudhury, B.; Grishin, A.V.; Ford, H.R.; Bode, L. The human milk oligosaccharide disialyllacto-N-tetraose prevents necrotising enterocolitis in neonatal rats. Gut 2012, 61, 1417–1425. [Google Scholar] [CrossRef]
- Ohtsuka, Y.; Okada, K.; Yamakawa, Y.; Ikuse, T.; Baba, Y.; Inage, E.; Fujii, T.; Izumi, H.; Oshida, K.; Nagata, S.; et al. omega-3 fatty acids attenuate mucosal inflammation in premature rat pups. J. Pediatr. Surg. 2011, 46, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Jilling, T.; Li, D.; Caplan, M.S. Polyunsaturated fatty acid supplementation alters proinflammatory gene expression and reduces the incidence of necrotizing enterocolitis in a neonatal rat model. Pediatr. Res. 2007, 61, 427–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caplan, M.S.; Russell, T.; Xiao, Y.; Amer, M.; Kaup, S.; Jilling, T. Effect of polyunsaturated fatty acid (PUFA) supplementation on intestinal inflammation and necrotizing enterocolitis (NEC) in a neonatal rat model. Pediatr. Res. 2001, 49, 647–652. [Google Scholar] [CrossRef] [Green Version]
- Ran-Ressler, R.R.; Khailova, L.; Arganbright, K.M.; Adkins-Rieck, C.K.; Jouni, Z.E.; Koren, O.; Ley, R.E.; Brenna, J.T.; Dvorak, B. Branched chain fatty acids reduce the incidence of necrotizing enterocolitis and alter gastrointestinal microbial ecology in a neonatal rat model. PLoS ONE 2011, 6, e29032. [Google Scholar] [CrossRef] [Green Version]
- Coursodon-Boyiddle, C.F.; Snarrenberg, C.L.; Adkins-Rieck, C.K.; Bassaganya-Riera, J.; Hontecillas, R.; Lawrence, P.; Brenna, J.T.; Jouni, Z.E.; Dvorak, B. Pomegranate seed oil reduces intestinal damage in a rat model of necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G744–G751. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Wen, J.; Zhou, J.; Cai, W.; Qian, L. Milk Fat Globule Membrane Ameliorates Necrotizing Enterocolitis in Neonatal Rats and Suppresses Lipopolysaccharide-Induced Inflammatory Response in IEC-6 Enterocytes. JPEN J. Parenter. Enter. Nutr. 2019. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, M.; Guo, H.; Yan, J.; Liu, F.; Chen, J.; Li, Y.; Ren, F. Human Milk Oligosaccharides Protect Against Necrotizing Enterocolitis by Inhibiting Intestinal Damage Via Increasing the Proliferation of Crypt Cells. Mol. Nutr. Food Res. 2019. [Google Scholar] [CrossRef]
- Xu, J.; Anderson, V.; Schwarz, S.M. Dietary GD3 ganglioside reduces the incidence and severity of necrotizing enterocolitis by sustaining regulatory immune responses. J. Pediatr. Gastroenterol. Nutr. 2013, 57, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Lei, Y.; He, X.; Liu, D.; He, Z. Role of lactadherin in intestinal barrier integrity in experimental neonatal necrotizing enterocolitis. J. Cell. Biochem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Caplan, M.S.; Lickerman, M.; Adler, L.; Dietsch, G.N.; Yu, A. The role of recombinant platelet-activating factor acetylhydrolase in a neonatal rat model of necrotizing enterocolitis. Pediatr. Res. 1997, 42, 779–783. [Google Scholar] [CrossRef] [Green Version]
- Halpern, M.D.; Dominguez, J.A.; Dvorakova, K.; Holubec, H.; Williams, C.S.; Meza, Y.G.; Ruth, M.C.; Dvorak, B. Ileal cytokine dysregulation in experimental necrotizing enterocolitis is reduced by epidermal growth factor. J. Pediatr. Gastroenterol. Nutr. 2003, 36, 126–133. [Google Scholar] [CrossRef]
- Dvorak, B.; Halpern, M.D.; Holubec, H.; Williams, C.S.; McWilliam, D.L.; Dominguez, J.A.; Stepankova, R.; Payne, C.M.; McCuskey, R.S. Epidermal growth factor reduces the development of necrotizing enterocolitis in a neonatal rat model. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 282, G156–G164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maynard, A.A.; Dvorak, K.; Khailova, L.; Dobrenen, H.; Arganbright, K.M.; Halpern, M.D.; Kurundkar, A.R.; Maheshwari, A.; Dvorak, B. Epidermal growth factor reduces autophagy in intestinal epithelium and in the rat model of necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G614–G622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khailova, L.; Dvorak, K.; Arganbright, K.M.; Williams, C.S.; Halpern, M.D.; Dvorak, B. Changes in hepatic cell junctions structure during experimental necrotizing enterocolitis: Effect of EGF treatment. Pediatr. Res. 2009, 66, 140–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Besner, G.E. M1 to M2 macrophage polarization in heparin-binding epidermal growth factor-like growth factor therapy for necrotizing enterocolitis. J. Surg. Res. 2015, 197, 126–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Watkins, D.; Chen, C.L.; Bhushan, B.; Zhou, Y.; Besner, G.E. Heparin-binding epidermal growth factor-like growth factor and mesenchymal stem cells act synergistically to prevent experimental necrotizing enterocolitis. J. Am. Coll. Surg. 2012, 215, 534–545. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Besner, G.E. Heparin-binding epidermal growth factor-like growth factor promotes enterocyte migration and proliferation in neonatal rats with necrotizing enterocolitis. J. Pediatr. Surg. 2007, 42, 214–220. [Google Scholar] [CrossRef]
- Feng, J.; El-Assal, O.N.; Besner, G.E. Heparin-binding epidermal growth factor-like growth factor reduces intestinal apoptosis in neonatal rats with necrotizing enterocolitis. J. Pediatr. Surg. 2006, 41, 742–747. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Yang, J.; Besner, G.E. HB-EGF promotes intestinal restitution by affecting integrin-extracellular matrix interactions and intercellular adhesions. Growth Factors 2013, 31, 39–55. [Google Scholar] [CrossRef]
- Feng, J.; El-Assal, O.N.; Besner, G.E. Heparin-binding EGF-like growth factor (HB-EGF) and necrotizing enterocolitis. Semin. Pediatr. Surg. 2005, 14, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.K.; Baggerman, E.W.; Mohankumar, K.; Namachivayam, K.; Jagadeeswaran, R.; Reyes, V.E.; Maheshwari, A. Amniotic fluid-borne hepatocyte growth factor protects rat pups against experimental necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G361–G369. [Google Scholar] [CrossRef] [Green Version]
- Shiou, S.R.; Yu, Y.; Guo, Y.; Westerhoff, M.; Lu, L.; Petrof, E.O.; Sun, J.; Claud, E.C. Oral administration of transforming growth factor-beta1 (TGF-beta1) protects the immature gut from injury via Smad protein-dependent suppression of epithelial nuclear factor kappaB (NF-kappaB) signaling and proinflammatory cytokine production. J. Biol. Chem. 2013, 288, 34757–34766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, F.; Liu, G.R.; Li, N.; Yuan, G. Insulin-like growth factor I reduces the occurrence of necrotizing enterocolitis by reducing inflammatory response and protecting intestinal mucosal barrier in neonatal rats model. Eur. Rev. Med. Pharm. Sci. 2017, 21, 4711–4719. [Google Scholar]
- Shiou, S.R.; Yu, Y.; Chen, S.; Ciancio, M.J.; Petrof, E.O.; Sun, J.; Claud, E.C. Erythropoietin protects intestinal epithelial barrier function and lowers the incidence of experimental neonatal necrotizing enterocolitis. J. Biol. Chem. 2011, 286, 12123–12132. [Google Scholar] [CrossRef] [Green Version]
- Hoang, T.K.; He, B.; Wang, T.; Tran, D.Q.; Rhoads, J.M.; Liu, Y. Protective effect of Lactobacillus reuteri DSM 17938 against experimental necrotizing enterocolitis is mediated by Toll-like receptor 2. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G231–G240. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Fatheree, N.Y.; Mangalat, N.; Rhoads, J.M. Lactobacillus reuteri strains reduce incidence and severity of experimental necrotizing enterocolitis via modulation of TLR4 and NF-kappaB signaling in the intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G608–G617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Tran, D.Q.; Fatheree, N.Y.; Marc Rhoads, J. Lactobacillus reuteri DSM 17938 differentially modulates effector memory T cells and Foxp3+ regulatory T cells in a mouse model of necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G177–G186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.; Guo, C.; Gong, F. Protective effect of Lactobacillus reuteri against oxidative stress in neonatal mice with necrotizing enterocolitis. Nan Fang Yi Ke Da Xue Xue Bao 2019, 39, 1221–1226. [Google Scholar] [CrossRef]
- Olson, J.K.; Rager, T.M.; Navarro, J.B.; Mashburn-Warren, L.; Goodman, S.D.; Besner, G.E. Harvesting the benefits of biofilms: A novel probiotic delivery system for the prevention of necrotizing enterocolitis. J. Pediatr. Surg. 2016, 51, 936–941. [Google Scholar] [CrossRef]
- Olson, J.K.; Navarro, J.B.; Allen, J.M.; McCulloh, C.J.; Mashburn-Warren, L.; Wang, Y.; Varaljay, V.A.; Bailey, M.T.; Goodman, S.D.; Besner, G.E. An enhanced Lactobacillus reuteri biofilm formulation that increases protection against experimental necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G408–G419. [Google Scholar] [CrossRef]
- Khailova, L.; Dvorak, K.; Arganbright, K.M.; Halpern, M.D.; Kinouchi, T.; Yajima, M.; Dvorak, B. Bifidobacterium bifidum improves intestinal integrity in a rat model of necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G940–G949. [Google Scholar] [CrossRef] [Green Version]
- Khailova, L.; Mount Patrick, S.K.; Arganbright, K.M.; Halpern, M.D.; Kinouchi, T.; Dvorak, B. Bifidobacterium bifidum reduces apoptosis in the intestinal epithelium in necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G1118–G1127. [Google Scholar] [CrossRef] [Green Version]
- Caplan, M.S.; Miller-Catchpole, R.; Kaup, S.; Russell, T.; Lickerman, M.; Amer, M.; Xiao, Y.; Thomson, R., Jr. Bifidobacterial supplementation reduces the incidence of necrotizing enterocolitis in a neonatal rat model. Gastroenterology 1999, 117, 577–583. [Google Scholar] [CrossRef]
- Wu, W.; Wang, Y.; Zou, J.; Long, F.; Yan, H.; Zeng, L.; Chen, Y. Bifidobacterium adolescentis protects against necrotizing enterocolitis and upregulates TOLLIP and SIGIRR in premature neonatal rats. BMC Pediatr. 2017, 17, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siggers, R.H.; Siggers, J.; Boye, M.; Thymann, T.; Mølbak, L.; Leser, T.; Jensen, B.B.; Sangild, P.T. Early administration of probiotics alters bacterial colonization and limits diet-induced gut dysfunction and severity of necrotizing enterocolitis in preterm pigs. J. Nutr. 2008, 138, 1437–1444. [Google Scholar] [CrossRef] [Green Version]
- Pisano, C.; Galley, J.; Elbahrawy, M.; Wang, Y.; Farrell, A.; Brigstock, D.; Besner, G.E. Human Breast Milk-Derived Extracellular Vesicles in the Protection Against Experimental Necrotizing Enterocolitis. J. Pediatr. Surg. 2020, 55, 54–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, Y.; Peng, F.; Shan, Y.; Jiang, J. Berberine reduces the occurrence of neonatal necrotizing enterocolitis by reducing the inflammatory response. Exp. Med. 2018, 16, 5280–5285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintanilla, H.D.; Liu, Y.; Fatheree, N.Y.; Atkins, C.L.; Hashmi, S.S.; Floros, J.; McCormack, F.X.; Rhoads, J.M.; Alcorn, J.L. Oral administration of surfactant protein-a reduces pathology in an experimental model of necrotizing enterocolitis. J. Pediatr. Gastroenterol. Nutr. 2015, 60, 613–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, Q.; Lv, Z.; Cai, W.; Song, H.; Qian, L.; Mu, H.; Shi, J.; Wang, X. Human β-defensin-3 promotes intestinal epithelial cell migration and reduces the development of necrotizing enterocolitis in a neonatal rat model. Pediatr. Res. 2014, 76, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Akisu, M.; Baka, M.; Coker, I.; Kultursay, N.; Huseyinov, A. Effect of dietary n-3 fatty acids on hypoxia-induced necrotizing enterocolitis in young mice. n-3 fatty acids alter platelet-activating factor and leukotriene B4 production in the intestine. Biol. Neonate 1998, 74, 31–38. [Google Scholar] [CrossRef]
- Zhu, X.; Cui, N.; Yu, L.; Cheng, P.; Cui, M.; Zhu, X.; Wang, J. Potential role of endoplasmic reticulum stress is involved in the protection of fish oil on neonatal rats with necrotizing enterocolitis. Sci. Rep. 2020, 10, 6448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Zhang, T.; Zhou, G.; Jiang, X.; Tao, M.; Zhang, J.; Zeng, X.; Wu, Z.; Pan, D.; Guo, Y. Prevention of Necrotizing Enterocolitis through Milk Polar Lipids Reducing Intestinal Epithelial Apoptosis. J. Agric. Food Chem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, C.P.; Fulton, W.B.; Good, M.; Vurma, M.; Das, T.; Lai, C.S.; Jia, H.; Yamaguchi, Y.; Lu, P.; Prindle, T.; et al. Fat composition in infant formula contributes to the severity of necrotising enterocolitis. Br. J. Nutr. 2018, 120, 665–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Pierro, A. Human milk oligosaccharides protect against necrotizing enterocolitis by activating intestinal cell differentiation. Mol. Nutr. Food Res. 2020, e2000519. [Google Scholar] [CrossRef] [PubMed]
- Autran, C.A.; Schoterman, M.H.; Jantscher-Krenn, E.; Kamerling, J.P.; Bode, L. Sialylated galacto-oligosaccharides and 2’-fucosyllactose reduce necrotising enterocolitis in neonatal rats. Br. J. Nutr. 2016, 116, 294–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Zhang, M.; Guo, H.; Yan, J.; Chen, L.; Teng, W.; Ren, F.; Li, Y.; Wang, X.; Luo, J.; et al. Human Milk Oligosaccharides Activate Epidermal Growth Factor Receptor and Protect Against Hypoxia-Induced Injuries in the Mouse Intestinal Epithelium and Caco2 Cells. J. Nutr. 2020, 150, 756–762. [Google Scholar] [CrossRef]
- Yu, H.; Lau, K.; Thon, V.; Autran, C.A.; Jantscher-Krenn, E.; Xue, M.; Li, Y.; Sugiarto, G.; Qu, J.; Mu, S.; et al. Synthetic disialyl hexasaccharides protect neonatal rats from necrotizing enterocolitis. Angew. Chem. Int. Ed. Engl. 2014, 53, 6687–6691. [Google Scholar] [CrossRef]
- Yu, H.; Yan, X.; Autran, C.A.; Li, Y.; Etzold, S.; Latasiewicz, J.; Robertson, B.M.; Li, J.; Bode, L.; Chen, X. Enzymatic and Chemoenzymatic Syntheses of Disialyl Glycans and Their Necrotizing Enterocolitis Preventing Effects. J. Org. Chem. 2017, 82, 13152–13160. [Google Scholar] [CrossRef] [Green Version]
- Good, M.; Sodhi, C.P.; Yamaguchi, Y.; Jia, H.; Lu, P.; Fulton, W.B.; Martin, L.Y.; Prindle, T.; Nino, D.F.; Zhou, Q.; et al. The human milk oligosaccharide 2’-fucosyllactose attenuates the severity of experimental necrotising enterocolitis by enhancing mesenteric perfusion in the neonatal intestine. Br. J. Nutr. 2016, 116, 1175–1187. [Google Scholar] [CrossRef] [Green Version]
- Sodhi, C.P.; Wipf, P.; Yamaguchi, Y.; Fulton, W.B.; Kovler, M.; Nino, D.F.; Zhou, Q.; Banfield, E.; Werts, A.D.; Ladd, M.R.; et al. The human milk oligosaccharides 2’-fucosyllactose and 6’-sialyllactose protect against the development of necrotizing enterocolitis by inhibiting toll-like receptor 4 signaling. Pediatr. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- He-Yang, J.; Zhang, W.; Liu, J.; Xue, P.; Zhou, X. Human breast milk oligosaccharides attenuate necrotizing enterocolitis in rats by suppressing mast cell accumulation, DPPI activity and TLR4 expression in ileum tissue, and regulating mitochondrial damage of Caco-2 cells. Int. Immunopharmacol. 2020, 88, 106881. [Google Scholar] [CrossRef] [PubMed]
- Gunasekaran, A.; Eckert, J.; Burge, K.; Zheng, W.; Yu, Z.; Kessler, S.; de la Motte, C.; Chaaban, H. Hyaluronan 35 kDa enhances epithelial barrier function and protects against the development of murine necrotizing enterocolitis. Pediatr. Res. 2019. [Google Scholar] [CrossRef]
- Akisu, M.; Baka, M.; Huseyinov, A.; Kultursay, N. The role of dietary supplementation with L-glutamine in inflammatory mediator release and intestinal injury in hypoxia/reoxygenation-induced experimental necrotizing enterocolitis. Ann. Nutr. Metab. 2003, 47, 262–266. [Google Scholar] [CrossRef]
- Dilsiz, A.; Ciftci, I.; Aktan, T.M.; Gurbilek, M.; Karagozoglu, E. Enteral glutamine supplementation and dexamethasone attenuate the local intestinal damage in rats with experimental necrotizing enterocolitis. Pediatr. Surg. Int. 2003, 19, 578–582. [Google Scholar] [CrossRef]
- Zhou, W.; Li, W.; Zheng, X.H.; Rong, X.; Huang, L.G. Glutamine downregulates TLR-2 and TLR-4 expression and protects intestinal tract in preterm neonatal rats with necrotizing enterocolitis. J. Pediatr. Surg. 2014, 49, 1057–1063. [Google Scholar] [CrossRef]
- Chen, Y.; Koike, Y.; Chi, L.; Ahmed, A.; Miyake, H.; Li, B.; Lee, C.; Delgado-Olguin, P.; Pierro, A. Formula feeding and immature gut microcirculation promote intestinal hypoxia, leading to necrotizing enterocolitis. Dis. Model. Mech. 2019, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akisu, M.; Ozmen, D.; Baka, M.; Habif, S.; Yalaz, M.; Arslanoglu, S.; Kultursay, N.; Bayindir, O. Protective effect of dietary supplementation with l-arginine and l-carnitine on hypoxia/reoxygenation-induced necrotizing enterocolitis in young mice. Biol. Neonate 2002, 81, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Moller, H.K.; Thymann, T.; Fink, L.N.; Frokiaer, H.; Kvistgaard, A.S.; Sangild, P.T. Bovine colostrum is superior to enriched formulas in stimulating intestinal function and necrotising enterocolitis resistance in preterm pigs. Br. J. Nutr. 2011, 105, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhu, H.T.; Li, B.; Robinson, S.C.; Lee, C.; O’Connell, J.S.; Bindi, E.; Zheng, S.; Sherman, P.M.; Pierro, A. Lactoferrin Reduces Necrotizing Enterocolitis Severity by Upregulating Intestinal Epithelial Proliferation. Eur. J. Pediatr. Surg. 2019. [Google Scholar] [CrossRef]
- Whitehouse, J.S.; Riggle, K.M.; Purpi, D.P.; Mayer, A.N.; Pritchard, K.A., Jr.; Oldham, K.T.; Gourlay, D.M. The protective role of intestinal alkaline phosphatase in necrotizing enterocolitis. J. Surg. Res. 2010, 163, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Rentea, R.M.; Liedel, J.L.; Welak, S.R.; Cassidy, L.D.; Mayer, A.N.; Pritchard, K.A., Jr.; Oldham, K.T.; Gourlay, D.M. Intestinal alkaline phosphatase administration in newborns is protective of gut barrier function in a neonatal necrotizing enterocolitis rat model. J. Pediatr. Surg. 2012, 47, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Isani, M.; Illingworth, L.; Herman, E.; Schmidt, M.; Barron, L.; Bowling, J.; Elizee, M.; Bai, I.; Gayer, C.; Grishin, A.; et al. Soybean-derived recombinant human epidermal growth factor protects against experimental necrotizing enterocolitis. J. Pediatr. Surg. 2018, 53, 1203–1207. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; El-Assal, O.N.; Besner, G.E. Heparin-binding epidermal growth factor-like growth factor decreases the incidence of necrotizing enterocolitis in neonatal rats. J. Pediatr. Surg. 2006, 41, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Radulescu, A.; Zorko, N.; Besner, G.E. Heparin-binding EGF-like growth factor increases intestinal microvascular blood flow in necrotizing enterocolitis. Gastroenterology 2009, 137, 221–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matheson, P.J.; Walker, S.K.; Maki, A.C.; Shaheen, S.P.; Neal Garrison, R.; Downard, C.D. Oral relaxin maintains intestinal blood flow in a rat model of NEC. J. Pediatr. Surg. 2014, 49, 961–964; discussion 964–965. [Google Scholar] [CrossRef]
- Maheshwari, A.; Kelly, D.R.; Nicola, T.; Ambalavanan, N.; Jain, S.K.; Murphy-Ullrich, J.; Athar, M.; Shimamura, M.; Bhandari, V.; Aprahamian, C.; et al. TGF-beta2 suppresses macrophage cytokine production and mucosal inflammatory responses in the developing intestine. Gastroenterology 2011, 140, 242–253. [Google Scholar] [CrossRef] [Green Version]
- Nino, D.F.; Sodhi, C.P.; Egan, C.E.; Zhou, Q.; Lin, J.; Lu, P.; Yamaguchi, Y.; Jia, H.; Martin, L.Y.; Good, M.; et al. Retinoic Acid Improves Incidence and Severity of Necrotizing Enterocolitis by Lymphocyte Balance Restitution and Repopulation of LGR5+ Intestinal Stem Cells. Shock 2017, 47, 22–32. [Google Scholar] [CrossRef]
- Egan, C.E.; Sodhi, C.P.; Good, M.; Lin, J.; Jia, H.; Yamaguchi, Y.; Lu, P.; Ma, C.; Branca, M.F.; Weyandt, S.; et al. Toll-like receptor 4-mediated lymphocyte influx induces neonatal necrotizing enterocolitis. J. Clin. Investig. 2016, 126, 495–508. [Google Scholar] [CrossRef] [Green Version]
- Xiao, S.; Li, Q.; Hu, K.; He, Y.; Ai, Q.; Hu, L.; Yu, J. Vitamin A and Retinoic Acid Exhibit Protective Effects on Necrotizing Enterocolitis by Regulating Intestinal Flora and Enhancing the Intestinal Epithelial Barrier. Arch. Med. Res. 2018, 49, 1–9. [Google Scholar] [CrossRef]
- Lyu, C.; Jiang, S.; Kong, M.; Chen, X.; Zhang, L. Vitamin D protects against necrotising enterocolitis in newborn mice by activating the ERK signalling pathway. Mol. Med. Rep. 2020, 22, 2107–2114. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Chen, Z.; Lin, R.; Liu, Y.; Wu, X.; Puthiyakunnon, S.; Wang, Y.; Zhu, B.; Zhang, Q.; Bai, Y.; et al. Bacteroides fragilis Strain ZY-312 Defense against Cronobacter sakazakii-Induced Necrotizing Enterocolitis In Vitro and in a Neonatal Rat Model. mSystems 2019, 4. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Lv, H.; Li, M.X.; Su, H.; Huang, L.G.; Li, J.; Yuan, W.M. Protective effects of bifidobacteria on intestines in newborn rats with necrotizing enterocolitis and its regulation on TLR2 and TLR4. Genet. Mol. Res. 2015, 14, 11505–11514. [Google Scholar] [CrossRef]
- Lu, W.C.; Zheng, X.; Liu, J.F.; Wu, W.C.; Chen, X.Y.; Wei, H.B.; Li, C.L.; Lin, M.J. Effect of Bifidobacterium on the expression of β-defensin-2 in intestinal tissue of neonatal rats with necrotizing enterocolitis. Zhongguo Dang Dai Er Ke Za Zhi 2018, 20, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, K.R.; Liu, S.X.; Tian, R.; Kushnir, A.; Turner, J.R.; Li, H.L.; Chou, P.M.; Weber, C.R.; De Plaen, I.G. Bifidobacteria stabilize claudins at tight junctions and prevent intestinal barrier dysfunction in mouse necrotizing enterocolitis. Am. J. Pathol. 2013, 182, 1595–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, T.; Izumi, H.; Iwabuchi, N.; Odamaki, T.; Namba, K.; Abe, F.; Xiao, J.Z. Bifidobacterium breve prevents necrotising enterocolitis by suppressing inflammatory responses in a preterm rat model. Benef. Microbes 2016, 7, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Hock, A.; Wu, R.Y.; Minich, A.; Botts, S.R.; Lee, C.; Antounians, L.; Miyake, H.; Koike, Y.; Chen, Y.; et al. Bovine milk-derived exosomes enhance goblet cell activity and prevent the development of experimental necrotizing enterocolitis. PLoS ONE 2019, 14, e0211431. [Google Scholar] [CrossRef]
- Miyake, H.; Lee, C.; Chusilp, S.; Bhalla, M.; Li, B.; Pitino, M.; Seo, S.; O’Connor, D.L.; Pierro, A. Human breast milk exosomes attenuate intestinal damage. Pediatr. Surg. Int. 2019. [Google Scholar] [CrossRef]
- Wang, X.; Yan, X.; Zhang, L.; Cai, J.; Zhou, Y.; Liu, H.; Hu, Y.; Chen, W.; Xu, S.; Liu, P.; et al. Identification and Peptidomic Profiling of Exosomes in Preterm Human Milk: Insights Into Necrotizing Enterocolitis Prevention. Mol. Nutr. Food Res. 2019, e1801247. [Google Scholar] [CrossRef] [PubMed]
- Cakir, U.; Tayman, C.; Serkant, U.; Yakut, H.I.; Cakir, E.; Ates, U.; Koyuncu, I.; Karaogul, E. Ginger (Zingiber officinale Roscoe) for the treatment and prevention of necrotizing enterocolitis. J. Ethnopharmacol. 2018, 225, 297–308. [Google Scholar] [CrossRef]
- Yakut, H.I.; Koyuncu, E.; Cakir, U.; Tayman, C.; Koyuncu, İ.; Taskin Turkmenoglu, T.; Cakir, E.; Ozyazici, A.; Aydogan, S.; Zenciroglu, A. Preventative and therapeutic effects of fennel (Foeniculum vulgare) seed extracts against necrotizing enterocolitis. J. Food Biochem. 2020, e13284. [Google Scholar] [CrossRef]
- Siggers, J.; Ostergaard, M.V.; Siggers, R.H.; Skovgaard, K.; Mølbak, L.; Thymann, T.; Schmidt, M.; Møller, H.K.; Purup, S.; Fink, L.N.; et al. Postnatal amniotic fluid intake reduces gut inflammatory responses and necrotizing enterocolitis in preterm neonates. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G864–G875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Wu, X.; Peng, B.; Zou, H.; Li, S.; Wang, J.; Cao, J. Curcumin improves necrotising microscopic colitis and cell pyroptosis by activating SIRT1/NRF2 and inhibiting the TLR4 signalling pathway in newborn rats. Innate Immun. 2020. [Google Scholar] [CrossRef]
- Cigsar, E.B.; Ali Karadag, C.; Tanik, C.; Fatih Aydin, A.; Ihsan Dokucu, A. The protective effects of sesamol in a neonatal rat model of necrotizing enterocolitis. J. Matern. Fetal Neonatal Med. 2018, 1–6. [Google Scholar] [CrossRef]
- Cai, Z.; Liu, J.; Bian, H.; Cai, J. Astragaloside IV ameliorates necrotizing enterocolitis by attenuating oxidative stress and suppressing inflammation via the vitamin D3-upregulated protein 1/NF-κB signaling pathway. Exp. Med. 2016, 12, 2702–2708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ergün, O.; Ergün, G.; Oktem, G.; Selvi, N.; Doğan, H.; Tunçyürek, M.; Saydam, G.; Erdener, A. Enteral resveratrol supplementation attenuates intestinal epithelial inducible nitric oxide synthase activity and mucosal damage in experimental necrotizing enterocolitis. J. Pediatr. Surg. 2007, 42, 1687–1694. [Google Scholar] [CrossRef] [PubMed]
- Catala, I.; Butel, M.J.; Bensaada, M.; Popot, F.; Tessedre, A.C.; Rimbault, A.; Szylit, O. Oligofructose contributes to the protective role of bifidobacteria in experimental necrotising enterocolitis in quails. J. Med. Microbiol. 1999, 48, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Ren, S.; Hui, Y.; Goericke-Pesch, S.; Pankratova, S.; Kot, W.; Pan, X.; Thymann, T.; Sangild, P.T.; Nguyen, D.N. Gut and immune effects of bioactive milk factors in preterm pigs exposed to prenatal inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2019. [Google Scholar] [CrossRef]
- Butel, M.J.; Roland, N.; Hibert, A.; Popot, F.; Favre, A.; Tessedre, A.C.; Bensaada, M.; Rimbault, A.; Szylit, O. Clostridial pathogenicity in experimental necrotising enterocolitis in gnotobiotic quails and protective role of bifidobacteria. J. Med. Microbiol. 1998, 47, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Lock, J.Y.; Carlson, T.L.; Yu, Y.; Lu, J.; Claud, E.C.; Carrier, R.L. Impact of Developmental Age, Necrotizing Enterocolitis Associated Stress, and Oral Therapeutic Intervention on Mucus Barrier Properties. Sci. Rep. 2020, 10, 6692. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Fatheree, N.Y.; Dingle, B.M.; Tran, D.Q.; Rhoads, J.M. Lactobacillus reuteri DSM 17938 changes the frequency of Foxp3+ regulatory T cells in the intestine and mesenteric lymph node in experimental necrotizing enterocolitis. PLoS ONE 2013, 8, e56547. [Google Scholar] [CrossRef]
- Cilieborg, M.S.; Bering, S.B.; Ostergaard, M.V.; Jensen, M.L.; Krych, L.; Newburg, D.S.; Sangild, P.T. Minimal short-term effect of dietary 2’-fucosyllactose on bacterial colonisation, intestinal function and necrotising enterocolitis in preterm pigs. Br. J. Nutr. 2016, 116, 834–841. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, S.O.; Martin, L.; Ostergaard, M.V.; Rudloff, S.; Roggenbuck, M.; Nguyen, D.N.; Sangild, P.T.; Bering, S.B. Human milk oligosaccharide effects on intestinal function and inflammation after preterm birth in pigs. J. Nutr. Biochem. 2017, 40, 141–154. [Google Scholar] [CrossRef]
- D’Souza, A.; Fordjour, L.; Ahmad, A.; Cai, C.; Kumar, D.; Valencia, G.; Aranda, J.V.; Beharry, K.D. Effects of probiotics, prebiotics, and synbiotics on messenger RNA expression of caveolin-1, NOS, and genes regulating oxidative stress in the terminal ileum of formula-fed neonatal rats. Pediatr. Res. 2010, 67, 526–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Souza, A.; Cai, C.L.; Kumar, D.; Cai, F.; Fordjour, L.; Ahmad, A.; Valencia, G.; Aranda, J.V.; Beharry, K.D. Cytokines and Toll-like receptor signaling pathways in the terminal ileum of hypoxic/hyperoxic neonatal rats: Benefits of probiotics supplementation. Am. J. Transl. Res. 2012, 4, 187–197. [Google Scholar] [PubMed]
- Clark, J.A.; Doelle, S.M.; Halpern, M.D.; Saunders, T.A.; Holubec, H.; Dvorak, K.; Boitano, S.A.; Dvorak, B. Intestinal barrier failure during experimental necrotizing enterocolitis: Protective effect of EGF treatment. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 291, G938–G949. [Google Scholar] [CrossRef] [PubMed]
- MohanKumar, K.; Namachivayam, K.; Ho, T.T.B.; Torres, B.A.; Ohls, R.K.; Maheshwari, A. Cytokines and growth factors in the developing intestine and during necrotizing enterocolitis. Semin. Perinatol. 2017, 41, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Viscardi, R.M.; Lyon, N.H.; Sun, C.C.; Hebel, J.R.; Hasday, J.D. Inflammatory cytokine mRNAs in surgical specimens of necrotizing enterocolitis and normal newborn intestine. Pediatr. Pathol. Lab. Med. 1997, 17, 547–559. [Google Scholar] [CrossRef]
- Miyake, H.; Lee, C.; Seo, S.; Li, B.; Pierro, A. Liver Organoids Generated from Mice with Necrotizing Enterocolitis Have Reduced Regenerative Capacity. Eur. J. Pediatr. Surg. 2019. [Google Scholar] [CrossRef]
- MohanKumar, K.; Kaza, N.; Jagadeeswaran, R.; Garzon, S.A.; Bansal, A.; Kurundkar, A.R.; Namachivayam, K.; Remon, J.I.; Bandepalli, C.R.; Feng, X.; et al. Gut mucosal injury in neonates is marked by macrophage infiltration in contrast to pleomorphic infiltrates in adult: Evidence from an animal model. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G93–G102. [Google Scholar] [CrossRef] [Green Version]
- Vincent, D.; Klinke, M.; Eschenburg, G.; Trochimiuk, M.; Appl, B.; Tiemann, B.; Bergholz, R.; Reinshagen, K.; Boettcher, M. NEC is likely a NETs dependent process and markers of NETosis are predictive of NEC in mice and humans. Sci. Rep. 2018, 8, 12612. [Google Scholar] [CrossRef] [Green Version]
- Weitkamp, J.H.; Koyama, T.; Rock, M.T.; Correa, H.; Goettel, J.A.; Matta, P.; Oswald-Richter, K.; Rosen, M.J.; Engelhardt, B.G.; Moore, D.J.; et al. Necrotising enterocolitis is characterised by disrupted immune regulation and diminished mucosal regulatory (FOXP3)/effector (CD4, CD8) T cell ratios. Gut 2013, 62, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Ma, F.; Li, S.; Gao, X.; Zhou, J.; Zhu, X.; Wang, D.; Cai, Y.; Li, F.; Yang, Q.; Gu, X.; et al. Interleukin-6-mediated CCR9(+) interleukin-17-producing regulatory T cells polarization increases the severity of necrotizing enterocolitis. EBioMedicine 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, S.X.; Berger, P.J.; Nold-Petry, C.A.; Nold, M.F. The immunological landscape in necrotising enterocolitis. Expert Rev. Mol. Med. 2016, 18, e12. [Google Scholar] [CrossRef] [Green Version]
- Hackam, D.J.; Sodhi, C.P. Toll-Like Receptor-Mediated Intestinal Inflammatory Imbalance in the Pathogenesis of Necrotizing Enterocolitis. Cell. Mol. Gastroenterol. Hepatol. 2018, 6, 229–238.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sodhi, C.P.; Neal, M.D.; Siggers, R.; Sho, S.; Ma, C.; Branca, M.F.; Prindle, T., Jr.; Russo, A.M.; Afrazi, A.; Good, M.; et al. Intestinal epithelial Toll-like receptor 4 regulates goblet cell development and is required for necrotizing enterocolitis in mice. Gastroenterology 2012, 143, 708–718.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leaphart, C.L.; Cavallo, J.; Gribar, S.C.; Cetin, S.; Li, J.; Branca, M.F.; Dubowski, T.D.; Sodhi, C.P.; Hackam, D.J. A critical role for TLR4 in the pathogenesis of necrotizing enterocolitis by modulating intestinal injury and repair. J. Immunol. 2007, 179, 4808–4820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gribar, S.C.; Sodhi, C.P.; Richardson, W.M.; Anand, R.J.; Gittes, G.K.; Branca, M.F.; Jakub, A.; Shi, X.H.; Shah, S.; Ozolek, J.A.; et al. Reciprocal expression and signaling of TLR4 and TLR9 in the pathogenesis and treatment of necrotizing enterocolitis. J. Immunol. 2009, 182, 636–646. [Google Scholar] [CrossRef] [Green Version]
- Nanthakumar, N.; Meng, D.; Goldstein, A.M.; Zhu, W.; Lu, L.; Uauy, R.; Llanos, A.; Claud, E.C.; Walker, W.A. The mechanism of excessive intestinal inflammation in necrotizing enterocolitis: An immature innate immune response. PLoS ONE 2011, 6, e17776. [Google Scholar] [CrossRef] [Green Version]
- Cuna, A.; Sampath, V. Genetic alterations in necrotizing enterocolitis. Semin. Perinatol. 2017, 41, 61–69. [Google Scholar] [CrossRef]
- Sampath, V.; Le, M.; Lane, L.; Patel, A.L.; Cohen, J.D.; Simpson, P.M.; Garland, J.S.; Hines, R.N. The NFKB1 (g.-24519delATTG) variant is associated with necrotizing enterocolitis (NEC) in premature infants. J. Surg. Res. 2011, 169, e51–e57. [Google Scholar] [CrossRef]
- Wolfs, T.G.; Derikx, J.P.; Hodin, C.M.; Vanderlocht, J.; Driessen, A.; de Bruine, A.P.; Bevins, C.L.; Lasitschka, F.; Gassler, N.; van Gemert, W.G.; et al. Localization of the lipopolysaccharide recognition complex in the human healthy and inflamed premature and adult gut. Inflamm. Bowel Dis. 2010, 16, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Jenke, A.C.; Zilbauer, M.; Postberg, J.; Wirth, S. Human beta-defensin 2 expression in ELBW infants with severe necrotizing enterocolitis. Pediatr. Res. 2012, 72, 513–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Härtel, C.; Hartz, A.; Pagel, J.; Rupp, J.; Stein, A.; Kribs, A.; Müller, A.; Haase, R.; Gille, C.; Böttger, R.; et al. NOD2 Loss-of-Function Mutations and Risks of Necrotizing Enterocolitis or Focal Intestinal Perforation in Very Low-birth-weight Infants. Inflamm. Bowel Dis. 2016, 22, 249–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caplan, M.S.; Sun, X.M.; Hseuh, W.; Hageman, J.R. Role of platelet activating factor and tumor necrosis factor-alpha in neonatal necrotizing enterocolitis. J. Pediatrics 1990, 116, 960–964. [Google Scholar] [CrossRef]
- Maheshwari, A.; Schelonka, R.L.; Dimmitt, R.A.; Carlo, W.A.; Munoz-Hernandez, B.; Das, A.; McDonald, S.A.; Thorsen, P.; Skogstrand, K.; Hougaard, D.M.; et al. Cytokines associated with necrotizing enterocolitis in extremely-low-birth-weight infants. Pediatr. Res. 2014, 76, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Benkoe, T.; Baumann, S.; Weninger, M.; Pones, M.; Reck, C.; Rebhandl, W.; Oehler, R. Comprehensive evaluation of 11 cytokines in premature infants with surgical necrotizing enterocolitis. PLoS ONE 2013, 8, e58720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelson, M.B.; Bagwell, C.E.; Rozycki, H.J. Circulating pro- and counterinflammatory cytokine levels and severity in necrotizing enterocolitis. Pediatrics 1999, 103, 766–771. [Google Scholar] [CrossRef]
- Rentea, R.M.; Liedel, J.L.; Fredrich, K.; Pritchard, K., Jr.; Oldham, K.T.; Simpson, P.M.; Gourlay, D.M. Enteral intestinal alkaline phosphatase administration in newborns decreases iNOS expression in a neonatal necrotizing enterocolitis rat model. J. Pediatr. Surg. 2013, 48, 124–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuna, A.; Yu, W.; Menden, H.L.; Feng, L.; Srinivasan, P.; Chavez-Bueno, S.; Ahmed, I.; Umar, S.; Sampath, V. NEC-like intestinal injury is ameliorated by Lactobacillus rhamnosus GG in parallel with SIGIRR and A20 induction in neonatal mice. Pediatr. Res. 2020. [Google Scholar] [CrossRef]
- Rentea, R.M.; Liedel, J.L.; Fredrich, K.; Welak, S.R.; Pritchard, K.A., Jr.; Oldham, K.T.; Simpson, P.M.; Gourlay, D.M. Intestinal alkaline phosphatase administration in newborns decreases systemic inflammatory cytokine expression in a neonatal necrotizing enterocolitis rat model. J. Surg. Res. 2012, 177, 228–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez de Medina, F.; Romero-Calvo, I.; Mascaraque, C.; Martínez-Augustin, O. Intestinal Inflammation and Mucosal Barrier Function. Inflamm. Bowel Dis. 2014, 20, 2394–2404. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.M.; Van Itallie, C.M. Physiology and function of the tight junction. Cold Spring Harb. Perspect. Biol. 2009, 1, a002584. [Google Scholar] [CrossRef]
- Halpern, M.D.; Denning, P.W. The role of intestinal epithelial barrier function in the development of NEC. Tissue Barriers 2015, 3, e1000707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, R.J.; Leaphart, C.L.; Mollen, K.P.; Hackam, D.J. The role of the intestinal barrier in the pathogenesis of necrotizing enterocolitis. Shock 2007, 27, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Schaart, M.W.; de Bruijn, A.C.; Bouwman, D.M.; de Krijger, R.R.; van Goudoever, J.B.; Tibboel, D.; Renes, I.B. Epithelial functions of the residual bowel after surgery for necrotising enterocolitis in human infants. J. Pediatr. Gastroenterol. Nutr. 2009, 49, 31–41. [Google Scholar] [CrossRef] [PubMed]
- McElroy, S.J.; Prince, L.S.; Weitkamp, J.H.; Reese, J.; Slaughter, J.C.; Polk, D.B. Tumor necrosis factor receptor 1-dependent depletion of mucus in immature small intestine: A potential role in neonatal necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G656–G666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salzman, N.H.; Polin, R.A.; Harris, M.C.; Ruchelli, E.; Hebra, A.; Zirin-Butler, S.; Jawad, A.; Martin Porter, E.; Bevins, C.L. Enteric defensin expression in necrotizing enterocolitis. Pediatr. Res. 1998, 44, 20–26. [Google Scholar] [CrossRef] [Green Version]
- Markasz, L.; Wanders, A.; Szekely, L.; Lilja, H.E. Diminished DEFA6 Expression in Paneth Cells Is Associated with Necrotizing Enterocolitis. Gastroenterol. Res. Pr. 2018, 2018, 7345426. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishna, K.P.; Macadangdang, B.R.; Rogers, M.B.; Tometich, J.T.; Firek, B.A.; Baker, R.; Ji, J.; Burr, A.H.P.; Ma, C.; Good, M.; et al. Maternal IgA protects against the development of necrotizing enterocolitis in preterm infants. Nat. Med. 2019. [Google Scholar] [CrossRef]
- Moore, S.A.; Nighot, P.; Reyes, C.; Rawat, M.; McKee, J.; Lemon, D.; Hanson, J.; Ma, T.Y. Intestinal barrier dysfunction in human necrotizing enterocolitis. J. Pediatr. Surg. 2016, 51, 1907–1913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bein, A.; Eventov-Friedman, S.; Arbell, D.; Schwartz, B. Intestinal tight junctions are severely altered in NEC preterm neonates. Pediatr. Neonatol. 2018, 59, 464–473. [Google Scholar] [CrossRef] [Green Version]
- Ares, G.; Buonpane, C.; Sincavage, J.; Yuan, C.; Wood, D.R.; Hunter, C.J. Caveolin 1 is Associated with Upregulated Claudin 2 in Necrotizing Enterocolitis. Sci. Rep. 2019, 9, 4982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Zhou, Y.; Besner, G.E. Heparin-binding EGF-like growth factor and enteric neural stem cell transplantation in the prevention of experimental necrotizing enterocolitis in mice. Pediatr. Res. 2015, 78, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.L.; Yu, X.; James, I.O.; Zhang, H.Y.; Yang, J.; Radulescu, A.; Zhou, Y.; Besner, G.E. Heparin-binding EGF-like growth factor protects intestinal stem cells from injury in a rat model of necrotizing enterocolitis. Lab. Investig. 2012, 92, 331–344. [Google Scholar] [CrossRef] [Green Version]
- Nair, J.; Lakshminrusimha, S. Role of NO and other vascular mediators in the etiopathogenesis of necrotizing enterocolitis. Front. Biosci. 2019, 11, 9–28. [Google Scholar] [CrossRef]
- Murdoch, E.M.; Sinha, A.K.; Shanmugalingam, S.T.; Smith, G.C.; Kempley, S.T. Doppler flow velocimetry in the superior mesenteric artery on the first day of life in preterm infants and the risk of neonatal necrotizing enterocolitis. Pediatrics 2006, 118, 1999–2003. [Google Scholar] [CrossRef]
- Nowicki, P.T.; Caniano, D.A.; Hammond, S.; Giannone, P.J.; Besner, G.E.; Reber, K.M.; Nankervis, C.A. Endothelial nitric oxide synthase in human intestine resected for necrotizing enterocolitis. J. Pediatr. 2007, 150, 40–45. [Google Scholar] [CrossRef]
- Grishin, A.; Bowling, J.; Bell, B.; Wang, J.; Ford, H.R. Roles of nitric oxide and intestinal microbiota in the pathogenesis of necrotizing enterocolitis. J. Pediatr. Surg. 2016, 51, 13–17. [Google Scholar] [CrossRef]
- Ford, H.; Watkins, S.; Reblock, K.; Rowe, M. The role of inflammatory cytokines and nitric oxide in the pathogenesis of necrotizing enterocolitis. J. Pediatr. Surg. 1997, 32, 275–282. [Google Scholar] [CrossRef]
- Nowicki, P.T.; Reber, K.M.; Giannone, P.J.; Nankervis, C.A.; Hammond, S.; Besner, G.E.; Caniano, D.A. Intestinal O2 Consumption in Necrotizing Enterocolitis: Role of Nitric Oxide. Pediatr. Res. 2006, 59, 500–505. [Google Scholar] [CrossRef] [Green Version]
- Upperman, J.S.; Potoka, D.; Grishin, A.; Hackam, D.; Zamora, R.; Ford, H.R. Mechanisms of nitric oxide-mediated intestinal barrier failure in necrotizing enterocolitis. Semin. Pediatr. Surg. 2005, 14, 159–166. [Google Scholar] [CrossRef]
- Nowicki, P.T.; Dunaway, D.J.; Nankervis, C.A.; Giannone, P.J.; Reber, K.M.; Hammond, S.B.; Besner, G.E.; Caniano, D.A. Endothelin-1 in human intestine resected for necrotizing enterocolitis. J. Pediatr. 2005, 146, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Günther, C.; Neumann, H.; Neurath, M.F.; Becker, C. Apoptosis, necrosis and necroptosis: Cell death regulation in the intestinal epithelium. Gut 2013, 62, 1062–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werts, A.D.; Fulton, W.B.; Ladd, M.R.; Saad-Eldin, A.; Chen, Y.X.; Kovler, M.L.; Jia, H.; Banfield, E.C.; Buck, R.; Goerhing, K.; et al. A Novel Role for Necroptosis in the Pathogenesis of Necrotizing Enterocolitis. Cell. Mol. Gastroenterol. Hepatol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Ates, U.; Gollu, G.; Kucuk, G.; Billur, D.; Bingol-Kologlu, M.; Yılmaz, Y.; Ozkan-Ulu, H.; Bayram, P.; Bagriacik, E.; Dindar, H. Increase in pro-apoptotic Bax expression and decrease in anti-apoptotic Bcl-2 expression in newborns with necrotizing enterocolitis. Arch. Argent. Pediatr. 2016, 114, 243–247. [Google Scholar] [CrossRef]
- Neal, M.D.; Sodhi, C.P.; Dyer, M.; Craig, B.T.; Good, M.; Jia, H.; Yazji, I.; Afrazi, A.; Richardson, W.M.; Beer-Stolz, D.; et al. A critical role for TLR4 induction of autophagy in the regulation of enterocyte migration and the pathogenesis of necrotizing enterocolitis. J. Immunol. 2013, 190, 3541–3551. [Google Scholar] [CrossRef]
- Haq, S.; Grondin, J.; Banskota, S.; Khan, W.I. Autophagy: Roles in intestinal mucosal homeostasis and inflammation. J. Biomed. Sci. 2019, 26, 19. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.; Struijs, M.C.; Mei, J.; Witte-Bouma, J.; Korteland-van Male, A.M.; de Bruijn, A.C.; van Goudoever, J.B.; Renes, I.B. Endoplasmic reticulum stress, unfolded protein response and altered T cell differentiation in necrotizing enterocolitis. PLoS ONE 2013, 8, e78491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afrazi, A.; Branca, M.F.; Sodhi, C.P.; Good, M.; Yamaguchi, Y.; Egan, C.E.; Lu, P.; Jia, H.; Shaffiey, S.; Lin, J.; et al. Toll-like receptor 4-mediated endoplasmic reticulum stress in intestinal crypts induces necrotizing enterocolitis. J. Biol. Chem. 2014, 289, 9584–9599. [Google Scholar] [CrossRef] [Green Version]
- Vieten, D.; Corfield, A.; Ramani, P.; Spicer, R. Proliferative response in necrotising enterocolitis is insufficient to prevent disease progression. Pediatr. Surg. Int. 2006, 22, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.A.; Lane, R.H.; Maclennan, N.K.; Holubec, H.; Dvorakova, K.; Halpern, M.D.; Williams, C.S.; Payne, C.M.; Dvorak, B. Epidermal growth factor reduces intestinal apoptosis in an experimental model of necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G755–G762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Shiou, S.-R.; Guo, Y.; Lu, L.; Westerhoff, M.; Sun, J.; Petrof, E.O.; Claud, E.C. Erythropoietin protects epithelial cells from excessive autophagy and apoptosis in experimental neonatal necrotizing enterocolitis. PLoS ONE 2013, 8, e69620. [Google Scholar] [CrossRef] [Green Version]
- Fundora, J.B.; Guha, P.; Shores, D.R.; Pammi, M.; Maheshwari, A. Intestinal dysbiosis and necrotizing enterocolitis: Assessment for causality using Bradford Hill criteria. Pediatr. Res. 2019. [Google Scholar] [CrossRef]
- Pammi, M.; Cope, J.; Tarr, P.I.; Warner, B.B.; Morrow, A.L.; Mai, V.; Gregory, K.E.; Kroll, J.S.; McMurtry, V.; Ferris, M.J.; et al. Intestinal dysbiosis in preterm infants preceding necrotizing enterocolitis: A systematic review and meta-analysis. Microbiome 2017, 5, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olm, M.R.; Bhattacharya, N.; Crits-Christoph, A.; Firek, B.A.; Baker, R.; Song, Y.S.; Morowitz, M.J.; Banfield, J.F. Necrotizing enterocolitis is preceded by increased gut bacterial replication, Klebsiella, and fimbriae-encoding bacteria. Sci. Adv. 2019, 5, eaax5727. [Google Scholar] [CrossRef] [Green Version]
- Stinson, L.F.; Boyce, M.C.; Payne, M.S.; Keelan, J.A. The Not-so-Sterile Womb: Evidence That the Human Fetus Is Exposed to Bacteria Prior to Birth. Front. Microbiol. 2019, 10, 1124. [Google Scholar] [CrossRef]
- Westerbeek, E.A.; van den Berg, A.; Lafeber, H.N.; Knol, J.; Fetter, W.P.; van Elburg, R.M. The intestinal bacterial colonisation in preterm infants: A review of the literature. Clin. Nutr. 2006, 25, 361–368. [Google Scholar] [CrossRef]
- Penders, J.; Thijs, C.; Vink, C.; Stelma, F.F.; Snijders, B.; Kummeling, I.; van den Brandt, P.A.; Stobberingh, E.E. Factors influencing the composition of the intestinal microbiota in early infancy. Pediatrics 2006, 118, 511–521. [Google Scholar] [CrossRef] [Green Version]
- Niemarkt, H.J.; De Meij, T.G.; van Ganzewinkel, C.-J.; de Boer, N.K.H.; Andriessen, P.; Hütten, M.C.; Kramer, B.W. Necrotizing Enterocolitis, Gut Microbiota, and Brain Development: Role of the Brain-Gut Axis. Neonatology 2019, 115, 423–431. [Google Scholar] [CrossRef]
- Rudloff, S.; Kuntz, S.; Ostenfeldt Rasmussen, S.; Roggenbuck, M.; Sprenger, N.; Kunz, C.; Sangild, P.T.; Brandt Bering, S. Metabolism of Milk Oligosaccharides in Preterm Pigs Sensitive to Necrotizing Enterocolitis. Front. Nutr. 2019, 6, 23. [Google Scholar] [CrossRef] [PubMed]
- Heyman, M.B. Lactose intolerance in infants, children, and adolescents. Pediatrics 2006, 118, 1279–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Book, L.S.; Herbst, J.J.; Jung, A.L. Carbohydrate malabsorption in necrotizing enterocolitis. Pediatrics 1976, 57, 201–204. [Google Scholar]
- Misselwitz, B.; Butter, M.; Verbeke, K.; Fox, M.R. Update on lactose malabsorption and intolerance: Pathogenesis, diagnosis and clinical management. Gut 2019, 68, 2080–2091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kien, C.L. Colonic fermentation of carbohydrate in the premature infant: Possible relevance to necrotizing enterocolitis. J. Pediatr. 1990, 117, S52–S58. [Google Scholar] [CrossRef]
- Thymann, T.; Moller, H.K.; Stoll, B.; Stoy, A.C.; Buddington, R.K.; Bering, S.B.; Jensen, B.B.; Olutoye, O.O.; Siggers, R.H.; Molbak, L.; et al. Carbohydrate maldigestion induces necrotizing enterocolitis in preterm pigs. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G1115–G1125. [Google Scholar] [CrossRef]
- Sangild, P.T. Gut responses to enteral nutrition in preterm infants and animals. Exp. Biol. Med. 2006, 231, 1695–1711. [Google Scholar] [CrossRef]
- Buddington, R.K.; Sangild, P.T.; Hance, B.; Huang, E.Y.; Black, D.D. Prenatal gastrointestinal development in the pig and responses after preterm birth. J. Anim. Sci. 2012, 90, 290–298. [Google Scholar] [CrossRef]
- Kulkarni, S.; Ganz, J.; Bayrer, J.; Becker, L.; Bogunovic, M.; Rao, M. Advances in Enteric Neurobiology: The “Brain” in the Gut in Health and Disease. J. Neurosci. 2018, 38, 9346–9354. [Google Scholar] [CrossRef] [Green Version]
- Goyal, R.K.; Hirano, I. The enteric nervous system. N. Engl. J. Med. 1996, 334, 1106–1115. [Google Scholar] [CrossRef]
- Yoo, B.B.; Mazmanian, S.K. The Enteric Network: Interactions between the Immune and Nervous Systems of the Gut. Immunity 2017, 46, 910–926. [Google Scholar] [CrossRef] [Green Version]
- Vergnolle, N.; Cirillo, C. Neurons and Glia in the Enteric Nervous System and Epithelial Barrier Function. Physiology 2018, 33, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Wedel, T.; Krammer, H.J.; Kuhnel, W.; Sigge, W. Alterations of the enteric nervous system in neonatal necrotizing enterocolitis revealed by whole-mount immunohistochemistry. Pediatr. Pathol. Lab. Med. 1998, 18, 57–70. [Google Scholar] [CrossRef]
- Sigge, W.; Wedel, T.; Kuhnel, W.; Krammer, H.J. Morphologic alterations of the enteric nervous system and deficiency of non-adrenergic non-cholinergic inhibitory innervation in neonatal necrotizing enterocolitis. Eur. J. Pediatr. Surg. 1998, 8, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yang, J.; Watkins, D.J.; Boomer, L.A.; Matthews, M.A.; Su, Y.; Besner, G.E. Enteric nervous system abnormalities are present in human necrotizing enterocolitis: Potential neurotransplantation therapy. Stem Cell Res. 2013, 4, 157. [Google Scholar] [CrossRef] [Green Version]
- Fagbemi, A.O.; Torrente, F.; Puleston, J.; Lakhoo, K.; James, S.; Murch, S.H. Enteric neural disruption in necrotizing enterocolitis occurs in association with myenteric glial cell CCL20 expression. J. Pediatr. Gastroenterol. Nutr. 2013, 57, 788–793. [Google Scholar] [CrossRef] [PubMed]
- Bush, T.G. Enteric glial cells. An upstream target for induction of necrotizing enterocolitis and Crohn’s disease? Bioessays 2002, 24, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Sun, J.; Kappel, S.S.; Gormsen, M.; Sangild, P.T.; Aunsholt, L. Gut transit time, using radiological contrast imaging, to predict early signs of necrotizing enterocolitis. Pediatr. Res. 2020. [Google Scholar] [CrossRef]
- Latorre, R.; Sternini, C.; De Giorgio, R.; Greenwood-Van Meerveld, B. Enteroendocrine cells: A review of their role in brain-gut communication. Neurogastroenterol. Motil. 2016, 28, 620–630. [Google Scholar] [CrossRef] [Green Version]
- Hodzic, Z.; Bolock, A.M.; Good, M. The Role of Mucosal Immunity in the Pathogenesis of Necrotizing Enterocolitis. Front. Pediatr. 2017, 5, 40. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Wang, Y.; Olson, J.; Yang, J.; Besner, G.E. Heparin-binding EGF-like growth factor promotes neuronal nitric oxide synthase expression and protects the enteric nervous system after necrotizing enterocolitis. Pediatr. Res. 2017, 82, 490–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Lavoie, P.M.; Lacaze-Masmonteil, T.; Rhainds, M.; Marc, I. Omega-3 long-chain polyunsaturated fatty acids for extremely preterm infants: A systematic review. Pediatrics 2014, 134, 120–134. [Google Scholar] [CrossRef] [Green Version]
- Collins, C.T.; Makrides, M.; McPhee, A.J.; Sullivan, T.R.; Davis, P.G.; Thio, M.; Simmer, K.; Rajadurai, V.S.; Travadi, J.; Berry, M.J.; et al. Docosahexaenoic Acid and Bronchopulmonary Dysplasia in Preterm Infants. N. Engl. J. Med. 2017, 376, 1245–1255. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.; Buys, N.; Li, C.; Sun, J.; Yin, C. Effects of prebiotics on sepsis, necrotizing enterocolitis, mortality, feeding intolerance, time to full enteral feeding, length of hospital stay, and stool frequency in preterm infants: A meta-analysis. Eur. J. Clin. Nutr. 2019, 73, 657–670. [Google Scholar] [CrossRef] [Green Version]
- Pammi, M.; Suresh, G. Enteral lactoferrin supplementation for prevention of sepsis and necrotizing enterocolitis in preterm infants. Cochrane Database Syst. Rev. 2020, 3, Cd007137. [Google Scholar] [CrossRef]
- Shah, P.S.; Shah, V.S.; Kelly, L.E. Arginine supplementation for prevention of necrotising enterocolitis in preterm infants. Cochrane Database Syst. Rev. 2017, 4, Cd004339. [Google Scholar] [CrossRef] [PubMed]
- Moe-Byrne, T.; Brown, J.V.; McGuire, W. Glutamine supplementation to prevent morbidity and mortality in preterm infants. Cochrane Database Syst. Rev. 2016, 4, Cd001457. [Google Scholar] [CrossRef]
- Foster, J.P.; Seth, R.; Cole, M.J. Oral immunoglobulin for preventing necrotizing enterocolitis in preterm and low birth weight neonates. Cochrane Database Syst. Rev. 2016, 4, Cd001816. [Google Scholar] [CrossRef] [Green Version]
- Ananthan, A.; Balasubramanian, H.; Rao, S.; Patole, S. Clinical Outcomes Related to the Gastrointestinal Trophic Effects of Erythropoietin in Preterm Neonates: A Systematic Review and Meta-Analysis. Adv. Nutr. 2018, 9, 238–246. [Google Scholar] [CrossRef]
- Hosseini, M.; Azampour, H.; Raeisi, S.; Behtari, M.; Valizadeh, H.; Saboohi, R. The effects of enteral artificial amniotic fluid-containing erythropoietin on short term outcomes of preterm infants. Turk. J. Pediatr. 2019, 61, 392–398. [Google Scholar] [CrossRef]
- Omar, O.M.; Massoud, M.N.; Ghazal, H.; Hassouna, H.; Somaa, M.F. Effect of enteral erythropoietin on feeding-related complications in preterm newborns: A pilot randomized controlled study. Arab J. Gastroenterol. 2020, 21, 37–42. [Google Scholar] [CrossRef] [PubMed]
- El-Ganzoury, M.M.; Awad, H.A.; El-Farrash, R.A.; El-Gammasy, T.M.; Ismail, E.A.; Mohamed, H.E.; Suliman, S.M. Enteral granulocyte-colony stimulating factor and erythropoietin early in life improves feeding tolerance in preterm infants: A randomized controlled trial. J. Pediatr. 2014, 165, 1140–1145.e1. [Google Scholar] [CrossRef] [PubMed]
- Wardle, S.P.; Hughes, A.; Chen, S.; Shaw, N.J. Randomised controlled trial of oral vitamin A supplementation in preterm infants to prevent chronic lung disease. Arch. Dis. Child. Fetal Neonatal Ed. 2001, 84, F9–F13. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Cheng, R.; Wang, Z. Early Vitamin A supplementation improves the outcome of retinopathy of prematurity in extremely preterm infants. Retina 2020, 40, 1176–1184. [Google Scholar] [CrossRef] [PubMed]
- Gray, K.D.; Messina, J.A.; Cortina, C.; Owens, T.; Fowler, M.; Foster, M.; Gbadegesin, S.; Clark, R.H.; Benjamin, D.K., Jr.; Zimmerman, K.O.; et al. Probiotic Use and Safety in the Neonatal Intensive Care Unit: A Matched Cohort Study. J. Pediatr. 2020. [Google Scholar] [CrossRef] [PubMed]
- Xiong, T.; Maheshwari, A.; Neu, J.; Ei-Saie, A.; Pammi, M. An Overview of Systematic Reviews of Randomized-Controlled Trials for Preventing Necrotizing Enterocolitis in Preterm Infants. Neonatology 2019, 1–11. [Google Scholar] [CrossRef]
- Morgan, R.L.; Preidis, G.A.; Kashyap, P.C.; Weizman, A.V.; Sadeghirad, B.; Chang, Y.; Florez, I.D.; Foroutan, F.; Shahid, S.; Zeraatkar, D. Probiotics Reduce Mortality and Morbidity in Preterm, Low Birth Weight Infants: A Systematic Review and Network Meta-analysis of Randomized Trials. Gastroenterology 2020. [Google Scholar] [CrossRef]
- Van den Akker, C.H.P.; van Goudoever, J.B.; Szajewska, H.; Embleton, N.D.; Hojsak, I.; Reid, D.; Shamir, R. Probiotics for Preterm Infants: A Strain-Specific Systematic Review and Network Meta-analysis. J. Pediatr. Gastroenterol. Nutr. 2018, 67, 103–122. [Google Scholar] [CrossRef]
- Van den Akker, C.H.P.; van Goudoever, J.B.; Shamir, R.; Domellöf, M.; Embleton, N.D.; Hojsak, I.; Lapillonne, A.; Mihatsch, W.A.; Canani, R.B.; Bronsky, J.; et al. Probiotics and Preterm Infants: A Position Paper by the ESPGHAN Committee on Nutrition and the ESPGHAN Working Group for Probiotics and Prebiotics. J. Pediatr. Gastroenterol. Nutr. 2020. [Google Scholar] [CrossRef]
- Manzoni, P.; Guardione, R.; Bonetti, P.; Priolo, C.; Maestri, A.; Mansoldo, C.; Mostert, M.; Anselmetti, G.; Sardei, D.; Bellettato, M.; et al. Lutein and zeaxanthin supplementation in preterm very low-birth-weight neonates in neonatal intensive care units: A multicenter randomized controlled trial. Am. J. Perinatol. 2013, 30, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Serce Pehlevan, O.; Benzer, D.; Gursoy, T.; Karatekin, G.; Ovali, F. Synbiotics use for preventing sepsis and necrotizing enterocolitis in very low birth weight neonates: A randomized controlled trial. Clin. Exp. Pediatr. 2020, 63, 226–231. [Google Scholar] [CrossRef]
- Indrio, F.; Riezzo, G.; Raimondi, F.; Bisceglia, M.; Cavallo, L.; Francavilla, R. Effects of probiotic and prebiotic on gastrointestinal motility in newborns. J. Physiol. Pharm. 2009, 60, 27–31. [Google Scholar]
- Mihatsch, W.A.; Hoegel, J.; Pohlandt, F. Prebiotic oligosaccharides reduce stool viscosity and accelerate gastrointestinal transport in preterm infants. Acta Paediatr. 2006, 95, 843–848. [Google Scholar] [CrossRef]
- Westerbeek, E.A.M.; MØrch, E.; Lafeber, H.N.; Fetter, W.P.F.; Twisk, J.W.R.; Van Elburg, R.M. Effect of neutral and acidic oligosaccharides on fecal IL-8 and fecal calprotectin in preterm infants. Pediatr. Res. 2011, 69, 255–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akin, I.M.; Atasay, B.; Dogu, F.; Okulu, E.; Arsan, S.; Karatas, H.D.; Ikinciogullari, A.; Turmen, T. Oral lactoferrin to prevent nosocomial sepsis and necrotizing enterocolitis of premature neonates and effect on T-regulatory cells. Am. J. Perinatol. 2014, 31, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Sevastiadou, S.; Malamitsi-Puchner, A.; Costalos, C.; Skouroliakou, M.; Briana, D.D.; Antsaklis, A.; Roma-Giannikou, E. The impact of oral glutamine supplementation on the intestinal permeability and incidence of necrotizing enterocolitis/septicemia in premature neonates. J. Matern. Fetal Neonatal Med. 2011, 24, 1294–1300. [Google Scholar] [CrossRef]
- Stratiki, Z.; Costalos, C.; Sevastiadou, S.; Kastanidou, O.; Skouroliakou, M.; Giakoumatou, A.; Petrohilou, V. The effect of a bifidobacter supplemented bovine milk on intestinal permeability of preterm infants. Early Hum. Dev. 2007, 83, 575–579. [Google Scholar] [CrossRef]
- Serce Pehlevan, O.; Benzer, D.; Gursoy, T.; Aktas Cetin, E.; Karatekin, G.; Ovali, M.F. Cytokine responses to symbiotic and lactoferrin combination in very low birth weight neonates: A randomized control trial. Arch. Argent. Pediatr. 2020, 118, e8–e15. [Google Scholar] [CrossRef] [PubMed]
- Nolan, L.S.; Parks, O.B.; Good, M. A Review of the Immunomodulating Components of Maternal Breast Milk and Protection Against Necrotizing Enterocolitis. Nutrients 2019, 12, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzoni, P.; Meyer, M.; Stolfi, I.; Rinaldi, M.; Cattani, S.; Pugni, L.; Romeo, M.G.; Messner, H.; Decembrino, L.; Laforgia, N.; et al. Bovine lactoferrin supplementation for prevention of necrotizing enterocolitis in very-low-birth-weight neonates: A randomized clinical trial. Early Hum. Dev. 2014, 90, S60–S65. [Google Scholar] [CrossRef]
- Gephart, S.M.; Underwood, M.A.; Rosito, S.; Kim, J.H.; Caplan, M.S. Grading the evidence to identify strategies to modify risk for necrotizing enterocolitis. Pediatr. Res. 2020, 88, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Ellis, Z.-M.; Tan, H.S.G.; Embleton, N.D.; Sangild, P.T.; van Elburg, R.M. Milk feed osmolality and adverse events in newborn infants and animals: A systematic review. Arch. Dis. Child. Fetal Neonatal Ed. 2019, 104, F333. [Google Scholar] [CrossRef] [Green Version]
- Ter Riet, G.; Korevaar, D.A.; Leenaars, M.; Sterk, P.J.; Van Noorden, C.J.; Bouter, L.M.; Lutter, R.; Elferink, R.P.; Hooft, L. Publication bias in laboratory animal research: A survey on magnitude, drivers, consequences and potential solutions. PLoS ONE 2012, 7, e43404. [Google Scholar] [CrossRef]
- Van der Naald, M.; Wenker, S.; Doevendans, P.A.; Wever, K.E.; Chamuleau, S.A.J. Publication rate in preclinical research: A plea for preregistration. BMJ Open Sci. 2020, 4, e100051. [Google Scholar] [CrossRef]
- Kilkenny, C.; Parsons, N.; Kadyszewski, E.; Festing, M.F.W.; Cuthill, I.C.; Fry, D.; Hutton, J.; Altman, D.G. Survey of the quality of experimental design, statistical analysis and reporting of research using animals. PLoS ONE 2009, 4, e7824. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, C.; Richardson, W.; Gribar, S.; Hackam, D.J. The development of animal models for the study of necrotizing enterocolitis. Dis. Model. Mech. 2008, 1, 94–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klinke, M.; Vincent, D.; Trochimiuk, M.; Appl, B.; Tiemann, B.; Reinshagen, K.; Pagerols Raluy, L.; Boettcher, M. Development of an improved murine model of necrotizing enterocolitis shows the importance of neutrophils in NEC pathogenesis. Sci. Rep. 2020, 10, 8049. [Google Scholar] [CrossRef]
- Lu, P.; Sodhi, C.P.; Jia, H.; Shaffiey, S.; Good, M.; Branca, M.F.; Hackam, D.J. Animal models of gastrointestinal and liver diseases. Animal models of necrotizing enterocolitis: Pathophysiology, translational relevance, and challenges. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G917–G928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovler, M.L.; Sodhi, C.P.; Hackam, D.J. Precision-based modeling approaches for necrotizing enterocolitis. Dis. Model. Mech. 2020, 13. [Google Scholar] [CrossRef]
- Neu, J. Necrotizing Enterocolitis: The Future. Neonatology 2020. [Google Scholar] [CrossRef] [PubMed]
- Neu, J.; Modi, N.; Caplan, M. Necrotizing enterocolitis comes in different forms: Historical perspectives and defining the disease. Semin. Fetal Neonatal Med. 2018, 23, 370–373. [Google Scholar] [CrossRef] [PubMed]
- Senger, S.; Ingano, L.; Freire, R.; Anselmo, A.; Zhu, W.; Sadreyev, R.; Walker, W.A.; Fasano, A. Human Fetal-Derived Enterospheres Provide Insights on Intestinal Development and a Novel Model to Study Necrotizing Enterocolitis (NEC). Cell. Mol. Gastroenterol. Hepatol. 2018, 5, 549–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashammakhi, N.; Nasiri, R.; Barros, N.R.d.; Tebon, P.; Thakor, J.; Goudie, M.; Shamloo, A.; Martin, M.G.; Khademhosseini, A. Gut-on-a-chip: Current progress and future opportunities. Biomaterials 2020, 255, 120196. [Google Scholar] [CrossRef] [PubMed]


| Fat-based interventions | AA and DHA [44] Egg phospholipids [44] PUFA [45] BCFA [46] Pomegranate seed oil [47] MFGM [48] DHA or EPA maternal intervention during pregnancy [43] |
| Carbohydrate/sugar-based interventions | HMO [49] GD3 [50] |
| Protein/amino acid-based interventions | Lactadherin [51] rPAF-AH [52] |
| Hormone/growth factor/vitamin-based interventions | EGF [37,41,53,54,55,56] HB-EGF [37,41,57,58,59,60,61,62] HGF [63] TGF-β1 [64] IGF1 [65] EPO [66] |
| Probiotic interventions | Lactobacillus reuteri DSM 17938 [67,68,69,70] Lactobacillus reuteri ATCC PTA 4659 [68] Lactobacillus reuteri biofilm on unloaded microspheres [71,72] Lactobacillus reuteri biofilm on MRS loaded microspheres [71] Lactobacillus reuteri biofilm on sucrose loaded microspheres [72] Lactobacillus reuteri biofilm on maltose loaded microspheres [72] Bifidobacterium bifidum OLB6378 [73,74] Bifidobacterium infantis [75] Bifidobacterium adolescentis [76] probiotic mixture (Bifidobacterium animalis DSM15954, Lactobacillus acidophilus DSM13241, Lactobacillus casei ATCC55544, Lactobacillus pentosus DSM14025 and Lactobacillus plantarum DSM13367) [77] |
| Other interventions | Amniotic fluid [63] Human breast milk extracellular vesicles [78] Berberine [79] Surfactant protein a [80] Human β-defensin-3 [81] |
| Fat-based interventions | Fish oil (rich in n-3 PUFA) [82,83] MPL [84] MFGM [48] Very low fat diet [85] Reduced long chain triacylglycerol diet (considered pre-digested) [85] Pomegranate seed oil [47] |
| Carbohydrate/sugar-based interventions | HMO [38,42,49,86,87,88,89,90] Neutral HMO (no sialic acids) [42] −2 HMO (two sialic acids) [42] DSLNT (HMO) [42] DSLNnT (synthetic disialyl glycan) [89,90] DS’LNnT (synthetic disialyl glycan) [89,90] 2′-FL [87,91,92] 6′-SL [92] 2′-FL and 6′-SL [42,92] Sialylated HMO [93] Sialylated GOS [87] GD3 [50] Hyaluronan 35 kD [94] |
| Protein/amino acid-based interventions | L-Glutamine/glutamine [84,95,96,97] Arginine [98,99] L-Carnitine [99] N-Acetylcysteine [85] Lactadherin [51] OPN [100] Lactoferrin [101] IAP [102,103] |
| Hormone/growth factor/vitamin-based interventions | EGF [53,54,56] Recombinant EGF from soybean extract [104] HB-EGF [41,58,59,60,62,105,106] HGF [63] relaxin [107] TGF-β1 [64] TGF-β2 [108] ATRA [109,110] Vitamin A [111] Vitamin D [112] |
| Probiotic interventions | Bacteroides fragilis ZY-312 [113] Lactobacillus reuteri DSM 17938 [68,69] Lactobacillus reuteri ATCC PTA 4659 [68] Lactobacillus reuteri biofilm on unloaded microspheres [71,72] Lactobacillus reuteri biofilm on MRS loaded microspheres [71] Lactobacillus reuteri biofilm on sucrose loaded microspheres [72] Lactobacillus reuteri biofilm on maltose loaded microspheres [72] Bifidobacterium microcapsules [114] Bifidobacterium mixture [115] Bifidobacterium adolescentis [76] Bifidobacterium infantis [116] Bifidobacterium bifidum OLB6378 [74] Bifidobacterium breve M-16V [117] Lactobacillus rhamnosus HN001 (live) [39] Lactobacillus rhamnosus HN001 (dead) [39] Lactobacillus rhamnosus isolated DNA [39] Probiotic mixture (Bifidobacterium animalis DSM15954, Lactobacillus acidophilus DSM13241, Lactobacillus casei ATCC55544, Lactobacillus pentosus DSM14025 and Lactobacillus plantarum DSM13367) [77] CpG-DNA [39] |
| Other interventions | Bovine milk exosomes [118] Native human breast milk exosomes [78,119] Pasteurized human breast milk exosomes [119] Preterm human breast milk exosomes [120] Ginger [121] Fennel seed extracts [122] Amniotic fluid [40,63,123] Curcumin [124] Sesamol [125] Astragaloside iv [126] Resveratrol [127] Berberine [79] Surfactant protein a [80] Human β-defensin-3 [81] |
| Fat-based interventions | DHA and EPA [83] MFGM [48] Very low fat diet [85] Reduced long chain triacylglycerol diet (considered pre-digested) [85] MPL [84] |
| carbohydrate/sugar based interventions | 2′-FL [91,92] 6′-SL [92] 2′-FL and 6′-SL [92] FOS [128] GD3 [50] |
| Protein/amino acid based interventions | Lactadherin [51] CGMP [129] OPN [129] |
| Hormone/growth factor/vitamin based interventions | EGF [54] HB-EGF [58] IGF1 [65] Vitamin D [112] Relaxin [107] |
| Probiotic interventions | Lactobacillus reuteri DSM17938 [70] Bifidobacterium infantis-longum strain CUETM 89-215 [130] Bifidobacterium adolescentis [76] Bacteroides fragilis ZY-312 [113] Lactobacillus rhamnosus HN001 (live) [39] Lactobacillus rhamnosus HN001 (dead) [39] Lactobacillus rhamnosus isolated DNA [39] |
| Other interventions | Ginger [121] Fennel seed extracts [122] Amniotic fluid [123] Sesamol [125] Human β-defensin-3 [81] |
| Fat-based interventions | PUFA [45] MFGM [48] |
| Carbohydrate/sugar-based interventions | HMO [42,49,88] Hyaluronan 35 kD [94] |
| Protein/amino acid-based interventions | Lactadherin [51] Lysozyme [131] rPAF-AH [52] |
| Hormone/growth factor/vitamin-based interventions | HB-EGF [41,58,59,62,105] |
| Probiotic interventions | Bacteroides fragilis ZY-312 [113] Lactobacillus reuteri DSM 17938 [68,69,132] Lactobacillus reuteri ATCC PTA 4659 [68] Lactobacillus reuteri biofilm on sucrose loaded microspheres [72] Lactobacillus reuteri biofilm on maltose loaded microspheres [72] Bifidobacterium adolescentis [76] Bifidobacterium infantis [75] Bifidobacterium breve M-16V [117] |
| Other interventions | Surfactant protein A [80] Human β-defensin-3 [81] |
| NEC Incidence | |
|---|---|
| Carbohydrate/sugar-based interventions | 2′-FL [133] Gangliosides [100] SL [100] Lactose [92] Mixture of 4 HMO [134] Mixture of 25 HMO [134] IFOS [49] |
| Protein/amino acid-based interventions | OPN [100,129] CGMP [129] Bovine lactoferrin [35,36] |
| Probiotic interventions | Lactobacillus reuteri DSM 20016 [71,72] |
| Other interventions | amniotic fluid [123] |
| NEC histological injury scores | |
| Fat-based interventions | |
| Carbohydrate/sugar-based interventions | 2′-FL [133] GOS [42,89] Lactose [38] 0 HMO (no sialic acids) [42] −1 HMO (one sialic acid) [42] −3 HMO (three sialic acids) [42] −4 HMO (four sialic acids) [42] Mixture of 4 HMO [134] Mixture of 25 HMO [134] 3‴-sLNnT [89] GD3 [89] DSLac [89] Neu5GC-DS’LNT [90] DS’LNnT [90] DSTa [90] DSGalB [90] Gangliosides [100] SL [100] |
| Protein/amino acid-based interventions | Bovine lactoferrin [35,36] (even higher score for [36]) OPN [129] CGMP [129] |
| Probiotic interventions | Lactobacillus reuteri DSM 20016 [71] |
| NEC clinical disease score or signs and symptoms | |
| Fat-based interventions | BCFA [46] |
| carbohydrate/sugar based interventions | Lactose [92] HMO [38,42] Mixture of four HMO [134] Mixture of 25 HMO [134] 2′-FL [133] GOS/FOS [135,136] GOS [42] |
| Protein/amino acid-based interventions | Glutamine [96] OPN [129] CGMP [129] |
| Probiotic interventions | Saccharomyces Boulardii [135,136] |
| Other interventions | Resveratrol [127] |
| NEC survival | |
| Fat-based interventions | Pomegranate seed oil [47] DHA [131] DHA or EPA maternal intervention during pregnancy [43] |
| Carbohydrate/sugar-based interventions | GOS/FOS [135] GOS [42] IFOS [49] |
| Hormone/growth factor/vitamin-based interventions | EGF [104,137] |
| Probiotic interventions | Saccharomyces boulardii [135] Lactobacillus reuteri DSM 20016 [72] Lactobacillus reuteri biofilm on unloaded microspheres [72] |
| Enteral Feeding Intervention | Effect on Systemic Inflammation (Compared to NEC Protocol Exposure without Feeding Intervention) |
|---|---|
| Carbohydrate/sugar-based interventions | |
| HMO | Serum IL8 (protein) ↓ [49,88] |
| Hyaluronan 35 kD | Plasma TNFα (protein) ↓ [94] Serum CXCL1 (protein) ↓ [94] Serum IL12p70 (protein) ↓ [94] Serum IL6 (protein) ↓ [94] Serum IFNϒ (protein) ↓ [94] |
| Protein/amino acid-based interventions | |
| IAP | Serum TNFα (protein) ↓ (dose dependent) [162] Serum IL1β (protein) ↓ (dose dependent) [162] Serum IL6 (protein) ↓ (dose dependent) [162] |
| Hormone/growth factor/Vitamin-based interventions | |
| TGF-β | Serum IL6 (protein) ↓ [64] Serum IFNϒ (protein) ↓ [64] |
| Probiotic interventions | |
| Bacteroides fragilis ZY-312 | Serum TNFα (protein) ↓ [113] Serum IFNϒ (protein) ↓ [113] Serum IL10 (protein) ↑ [113] |
| Other interventions | |
| Berberine | Serum IL6 (protein) ↓ [79] serum IL10 (protein) ↓ [79] |
| Human β-defensin-3 | Serum TNFα (protein) ↓ [81] |
| Astragaloside IV | Serum TNFα (protein) ↓ [126] Serum IL6 (protein) ↓ [126] serum IL1β (protein) ↓ [126] |
| Enteral Feeding Intervention | Effect on Enteric Nervous System (Compared to NEC Protocol Exposure without Feeding Intervention) |
|---|---|
| Hormone/growth factor/vitamin-based interventions | |
| HB-EGF | Intestinal motility (dye migration assay) ↑ [175] Intestinal neuronal integrity Hu/D (protein) ↑ [222] Intestinal total neuronal count Hu/D = [175] Intestinal glial cell integrity GFAP (protein) ↑ [222] Intestinal nNOS expression (protein) ↑ [222] Intestinal neuronal apoptosis HuC/D TUNEL (protein) ↑ [222] Intestinal neuronal degeneration HuC/D FluoJade C (protein) ↑ [222] Jejunal entero-endocrine cells Chromogranin A (protein) ↑ [176] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Lange, I.H.; van Gorp, C.; Eeftinck Schattenkerk, L.D.; van Gemert, W.G.; Derikx, J.P.M.; Wolfs, T.G.A.M. Enteral Feeding Interventions in the Prevention of Necrotizing Enterocolitis: A Systematic Review of Experimental and Clinical Studies. Nutrients 2021, 13, 1726. https://doi.org/10.3390/nu13051726
de Lange IH, van Gorp C, Eeftinck Schattenkerk LD, van Gemert WG, Derikx JPM, Wolfs TGAM. Enteral Feeding Interventions in the Prevention of Necrotizing Enterocolitis: A Systematic Review of Experimental and Clinical Studies. Nutrients. 2021; 13(5):1726. https://doi.org/10.3390/nu13051726
Chicago/Turabian Stylede Lange, Ilse H., Charlotte van Gorp, Laurens D. Eeftinck Schattenkerk, Wim G. van Gemert, Joep P. M. Derikx, and Tim G. A. M. Wolfs. 2021. "Enteral Feeding Interventions in the Prevention of Necrotizing Enterocolitis: A Systematic Review of Experimental and Clinical Studies" Nutrients 13, no. 5: 1726. https://doi.org/10.3390/nu13051726
APA Stylede Lange, I. H., van Gorp, C., Eeftinck Schattenkerk, L. D., van Gemert, W. G., Derikx, J. P. M., & Wolfs, T. G. A. M. (2021). Enteral Feeding Interventions in the Prevention of Necrotizing Enterocolitis: A Systematic Review of Experimental and Clinical Studies. Nutrients, 13(5), 1726. https://doi.org/10.3390/nu13051726