
Acute Low-Intensity Treadmill Running Upregulates the Expression of Intestinal Glucose Transporters via GLP-2 in Mice

 ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Treatment and Acute Exercise
2.2. Tissue Sampling
2.3. Histochemical Staining and Immunohistochemistry
2.4. Measurements of Blood Lactate and Plasma GLP-2, Glucose, Triglyceride (TG), and Non-Esterified Fatty Acid (NEFA) Levels
2.5. Immunoblotting
2.6. Statistical Analysis
3. Results
3.1. Acute Exercise Did Not Affect the Blood Lactate Levels
3.2. Acute Exercise Increased Plasma GLP-2 Levels and Upregulated GLP-2R Expression in the Jejunum
3.3. Acute Exercise Did Not Affect the Plasma Concentrations of Glucose, TG, and NEFA
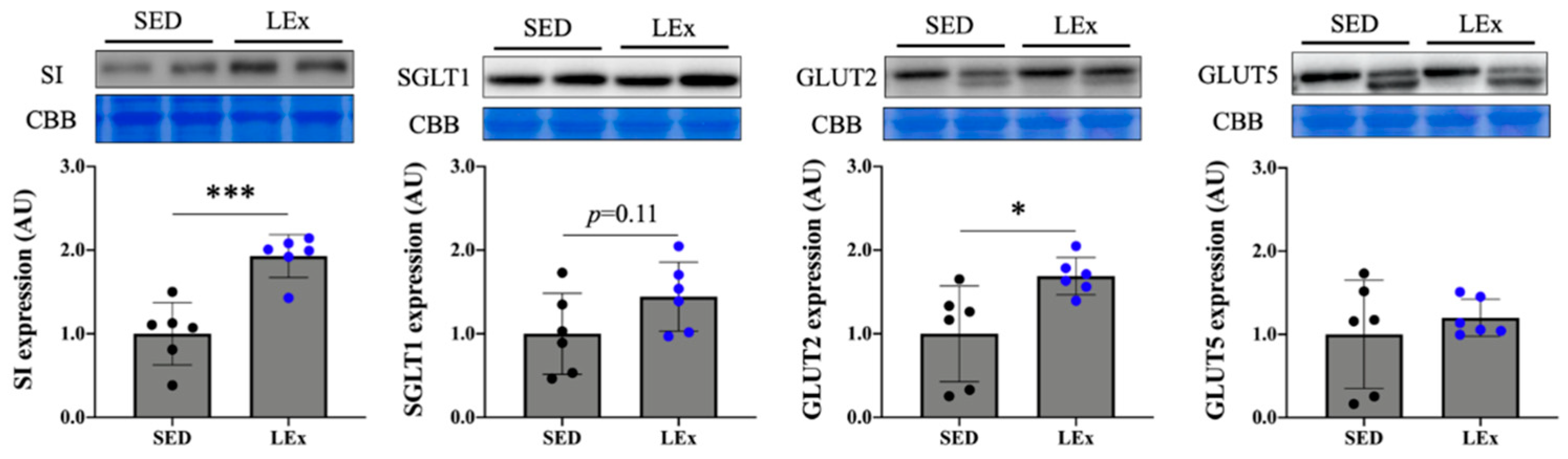
3.4. Acute Exercise Upregulated the Expression of SI and GLUT2 Proteins in the Jejunum
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lourenco, M.V.; Frozza, R.L.; de Freitas, G.B.; Zhang, H.; Kincheski, G.C.; Ribeiro, F.C.; Gonçalves, R.A.; Clarke, J.R.; Beckman, D.; Staniszewski, A.; et al. Exercise-linked FNDC5/irisin rescues synaptic plasticity and memory defects in Alzheimer’s models. Nat. Med. 2019, 25, 165–175. [Google Scholar] [CrossRef]
- Maesako, M.; Uemura, K.; Kubota, M.; Kuzuya, A.; Sasaki, K.; Hayashida, N.; Asada-Utsugi, M.; Watanabe, K.; Uemura, M.; Kihara, T.; et al. Exercise is more effective than diet control in preventing high fat diet-induced β-amyloid deposition and memory deficit in amyloid precursor protein transgenic mice. J. Biol. Chem. 2012, 287, 23024–23033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, K.; Konno, M.; Honda, K.; Abe, T.; Nagata, T.; Takehara, M.; Sugasawa, T.; Takekoshi, K.; Ohmori, H. Habitual aerobic exercise diminishes the effects of Prone8 model. Geriatrics 2020, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Ziaaldini, M.M.; Koltai, E.; Csende, Z.; Goto, S.; Boldogh, I.; Taylor, A.W.; Radak, Z. Exercise training increases anabolic and attenuates catabolic and apoptotic processes in aged skeletal muscle of male rats. Exp. Gerontol. 2015, 67, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eshima, H.; Tamura, Y.; Kakehi, S.; Nakamura, K.; Kurebayashi, N.; Murayama, T.; Kakigi, R.; Sakurai, T.; Kawamori, R.; Watada, H. Dysfunction of muscle contraction with impaired intracellular Ca2 handling in skeletal muscle and the effect of exercise training in male db/db mice. J. Appl. Physiol. 2019, 126, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Polaki, V.; Chen, S.; Bihl, J.C. Exercise improves endothelial function associated with alleviated inflammation and oxidative stress of perivascular adipose tissue in Type 2 diabetic mice. Oxid. Med. Cell. Longev. 2020, 2020, 8830537. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, K. Effect of resistance exercise training on Crohn’s disease patients. Intest. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Scheiman, J.; Luber, J.M.; Chavkin, T.A.; MacDonald, T.; Tung, A.; Pham, L.D.; Wibowo, M.C.; Wurth, R.C.; Punthambaker, S.; Tierney, B.T.; et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nat. Med. 2019, 25, 1104–1109. [Google Scholar] [CrossRef]
- Pasini, E.; Corsetti, G.; Assanelli, D.; Testa, C.; Romano, C.; Dioguardi, F.S.; Aquilani, R. Effects of chronic exercise on gut microbiota and intestinal barrier in human with type 2 diabetes. Minerva Med. 2019, 110, 3–11. [Google Scholar] [CrossRef]
- Holland, A.M.; Hyatt, H.W.; Smuder, A.J.; Sollanek, K.J.; Morton, A.B.; Roberts, M.D.; Kavazis, A.N. Influence of endurance exercise training on antioxidant enzymes, tight junction proteins, and inflammatory markers in the rat ileum. BMC Res. Notes 2015, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Maillard, F.; Vazeille, E.; Sauvanet, P.; Sirvent, P.; Combaret, L.; Sourdrille, A.; Chavanelle, V.; Bonnet, R.; Otero, Y.F.; Delcros, G.; et al. High intensity interval training promotes total and visceral fat mass loss in obese Zucker rats without modulating gut microbiota. PLoS ONE 2019, 14, e0214660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, K.; Yanazawa, K.; Tokinoya, K.; Sugasawa, T.; Suzuki, T.; Yoshida, Y.; Nakano, T.; Omi, N.; Kawakami, Y.; Takekoshi, K. Renalase is localized to the small intestine crypt and expressed upon the activation of NF- κ B p65 in mice model of fasting-induced oxidative stress. Life Sci. 2021, 267, 118904. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Mayanagi, Y.; Keta, A.; Kasahara, A.; Sato, A.; Takahashi, T. Oral administration of fructose improves jejunal villous morphology and nutrient digestion and absorption capabilities in a rat model of total parenteral nutrition. Biomed. Res. Clin. Pract. 2019, 4, 1–8. [Google Scholar] [CrossRef]
- Motiani, K.K.; Savolainen, A.M.; Eskelinen, J.J.; Toivanen, J.; Ishizu, T.; Yli-Karjanmaa, M.; Virtanen, K.A.; Parkkola, R.; Kapanen, J.; Grönroos, T.J.; et al. Two weeks of moderate-intensity continuous training, but not high-intensity interval training, increases insulin-stimulated intestinal glucose uptake. J. Appl. Physiol. 2017, 122, 1188–1197. [Google Scholar] [CrossRef] [Green Version]
- Kondo, S.; Fukazawa, A.; Karasawa, T.; Terada, S. Effects of long-term exercise training for different durations on pancreatic amylase activity and intestinal glucose transporter content in rats. Physiol. Rep. 2019, 7, e14255. [Google Scholar] [CrossRef] [PubMed]
- Au, A.; Gupta, A.; Schembri, P.; Cheeseman, C.I. Rapid insertion of GLUT2 into the rat jejunal brush-border membrane promoted by glucagon-like peptide 2. Biochem. J. 2002, 367, 247–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, R.B.; Kirk, D.; Macnaughton, W.K.; Meddings, J.B. GLP-2 augments the adaptive response to massive intestinal resection in rat. Am. J. Physiol 1998, 275, 911–921. [Google Scholar] [CrossRef]
- Kitchen, P.A.; Fitzgerald, A.J.; Goodlad, R.A.; Barley, N.F.; Ghatei, M.A.; Legon, S.; Bloom, S.R.; Price, A.; Walters, J.R.; Forbes, A. Glucagon-like peptide-2 increases sucrase-isomaltase but not caudal-related homeobox protein-2 gene expression. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 278, 425–428. [Google Scholar] [CrossRef]
- Cheeseman, C.I. Upregulation of SGLT-1 transport activity in rat jejunum induced by GLP-2 infusion in vivo. Am. J. Physiol. 1997, 273, 1965–1971. [Google Scholar] [CrossRef] [Green Version]
- Topstad, D.; Martin, G.; Sigalet, D. Systemic GLP-2 levels do not limit adaptation after distal intestinal resection. J. Pediatr. Surg. 2001, 36, 750–754. [Google Scholar] [CrossRef]
- Jeppesen, P.B.; Hartmann, B.; Thulesen, J.; Hansen, B.S.; Holst, J.J.; Poulsen, S.S.; Mortensen, P.B. Elevated plasma glucagon-like peptide 1 and 2 concentrations in ileum resected short bowel patients with a preserved colon. Gut 2000, 47, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J.; Shi, Q.; Crivici, A.; Sumner-Smith, M.; Tavares, W.; Hill, M.; DeForest, L.; Cooper, S.; Brubaker, P.L. Regulation of the biological activity of glucagon-like peptide 2 in vivo by dipeptidyl peptidase IV. Nat. Biotechnol. 1997, 15, 673–677. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J.; DeForest, L.; Brubaker, P.L. Intestinal response to growth factors administered alone or in combination with human [Gly2] glucagon-like peptide 2. Am. J. Physiol. 1997, 273, G1252–G1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chance, W.T.; Foley-Nelson, T.; Thomas, I.; Balasubramaniam, A. Prevention of parenteral nutrition-induced gut hypoplasia by coinfusion of glucagon-like peptide-2. Am. J. Physiol. 1997, 273, G559–G563. [Google Scholar] [CrossRef] [PubMed]
- L’Heureux, M.C.; Brubaker, P.L. Glucagon-like peptide-2 and common therapeutics in a murine model of ulcerative colitis. J. Pharmacol. Exp. Ther. 2003, 306, 347–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alavi, K.; Schwartz, M.Z.; Palazzo, J.P.; Prasad, R. Treatment of inflammatory bowel disease in a rodent model with the intestinal growth factor glucagon-like peptide-2. J. Pediatr. Surg. 2000, 35, 847–851. [Google Scholar] [CrossRef]
- Buchman, A.L.; Katz, S.; Fang, J.C.; Bernstein, C.N.; Abou-Assi, S.G.; Teduglutide Study Group. Teduglutide, a novel mucosally active analog of glucagon-like peptide-2 (GLP-2) for the treatment of moderate to severe Crohn’s disease. Inflamm. Bowel Dis. 2010, 16, 962–973. [Google Scholar] [CrossRef]
- Martins, C.; Morgan, L.M.; Bloom, S.R.; Robertson, M.D. Effects of exercise on gut peptides, energy intake and appetite. J. Endocrinol. 2007, 193, 251–258. [Google Scholar] [CrossRef]
- JanssenDuijghuijsen, L.M.; Mensink, M.; Lenaerts, K.; Fiedorowicz, E.; van Dartel, D.A.M.; Mes, J.J.; Luiking, Y.C.; Keijer, J.; Wichers, H.J. The effect of endurance exercise on intestinal integrity in well-trained healthy men. Physiol. Rep. 2016, 4, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Brubaker, P.L. Glucagon-like peptide-2 and the regulation of intestinal growth and function. Compr. Physiol. 2018, 18, 1185–1210. [Google Scholar] [CrossRef]
- Hiramoto, K.; Kobayashi, H.; Sekiyama, A.F.; Sato, E.; Tsuruta, D.; Ishii, M. Mild exercise suppresses exacerbation of dermatitis by increasing cleavage of the β-endorphin from proopiomelanocortin in NC/Nga mice. J. Clin. Biochem. Nutr. 2013, 52, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Rocca, A.S.; Brubaker, P.L. Role of the vagus nerve in mediating proximal nutrient-induced glucagon-like peptide-1 secretion. Endocrinology 1999, 140, 1687–1694. [Google Scholar] [CrossRef] [PubMed]
- Brenner, I.K.M.; Brown, C.A.; Hains, S.J.M.; Tranmer, J.; Zelt, D.T.; Brown, P.M. Low-intensity exercise training increases heart rate variability in patients with peripheral artery disease. Biol. Res. Nurs. 2020, 22, 24–33. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aoki, K.; Suzuki, T.; Hui, F.; Nakano, T.; Yanazawa, K.; Yonamine, M.; Fujita, S.; Sugasawa, T.; Yoshida, Y.; Omi, N.; et al. Acute Low-Intensity Treadmill Running Upregulates the Expression of Intestinal Glucose Transporters via GLP-2 in Mice. Nutrients 2021, 13, 1735. https://doi.org/10.3390/nu13051735
Aoki K, Suzuki T, Hui F, Nakano T, Yanazawa K, Yonamine M, Fujita S, Sugasawa T, Yoshida Y, Omi N, et al. Acute Low-Intensity Treadmill Running Upregulates the Expression of Intestinal Glucose Transporters via GLP-2 in Mice. Nutrients. 2021; 13(5):1735. https://doi.org/10.3390/nu13051735
Chicago/Turabian StyleAoki, Kai, Takuji Suzuki, Fang Hui, Takuro Nakano, Koki Yanazawa, Masato Yonamine, Shinichiro Fujita, Takehito Sugasawa, Yasuko Yoshida, Naomi Omi, and et al. 2021. "Acute Low-Intensity Treadmill Running Upregulates the Expression of Intestinal Glucose Transporters via GLP-2 in Mice" Nutrients 13, no. 5: 1735. https://doi.org/10.3390/nu13051735
APA StyleAoki, K., Suzuki, T., Hui, F., Nakano, T., Yanazawa, K., Yonamine, M., Fujita, S., Sugasawa, T., Yoshida, Y., Omi, N., Kawakami, Y., & Takekoshi, K. (2021). Acute Low-Intensity Treadmill Running Upregulates the Expression of Intestinal Glucose Transporters via GLP-2 in Mice. Nutrients, 13(5), 1735. https://doi.org/10.3390/nu13051735