
Myoinositol Reduces Inflammation and Oxidative Stress in Human Endothelial Cells Exposed In Vivo to Chronic Hyperglycemia

 ,
, 

 ,
,
Abstract
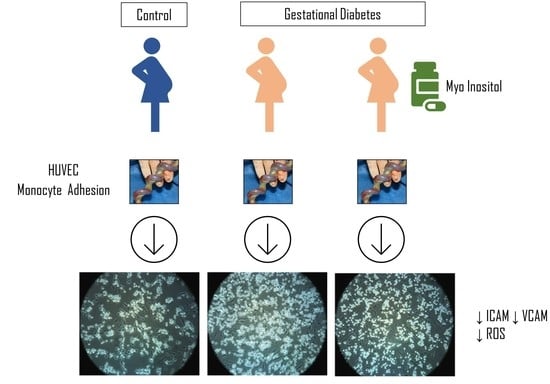
:
1. Introduction
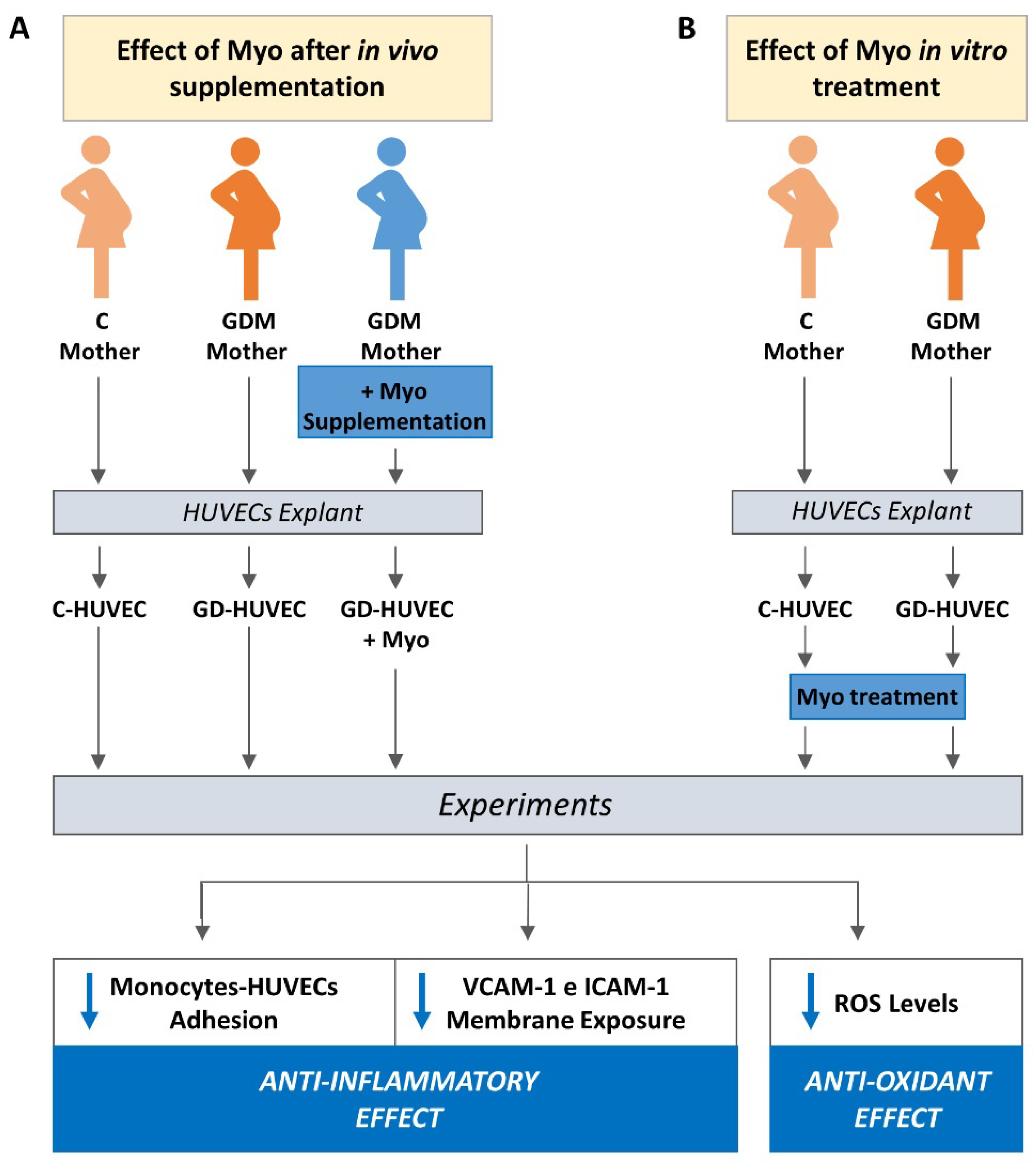
2. Materials and Methods
2.1. Study Design and Clinical Characteristic of Cords Donors
- carbohydrates 50% kcal/day (<10% simple sugars);
- lipids 30% kcal/day (<10% saturated fatty acids);
- protein 20% kcal/day (about 0.9 gr/kg/day);
- fibers at least 28 gr/day.
2.2. Cell Cultures and Experimental Protocols
2.3. Cell Growth and Viability
2.4. U937 Culture and Adhesion Assays
2.5. VCAM-1 and ICAM-1 Membrane Exposure
2.6. ROS Levels
2.7. Statistical Analysis
3. Results
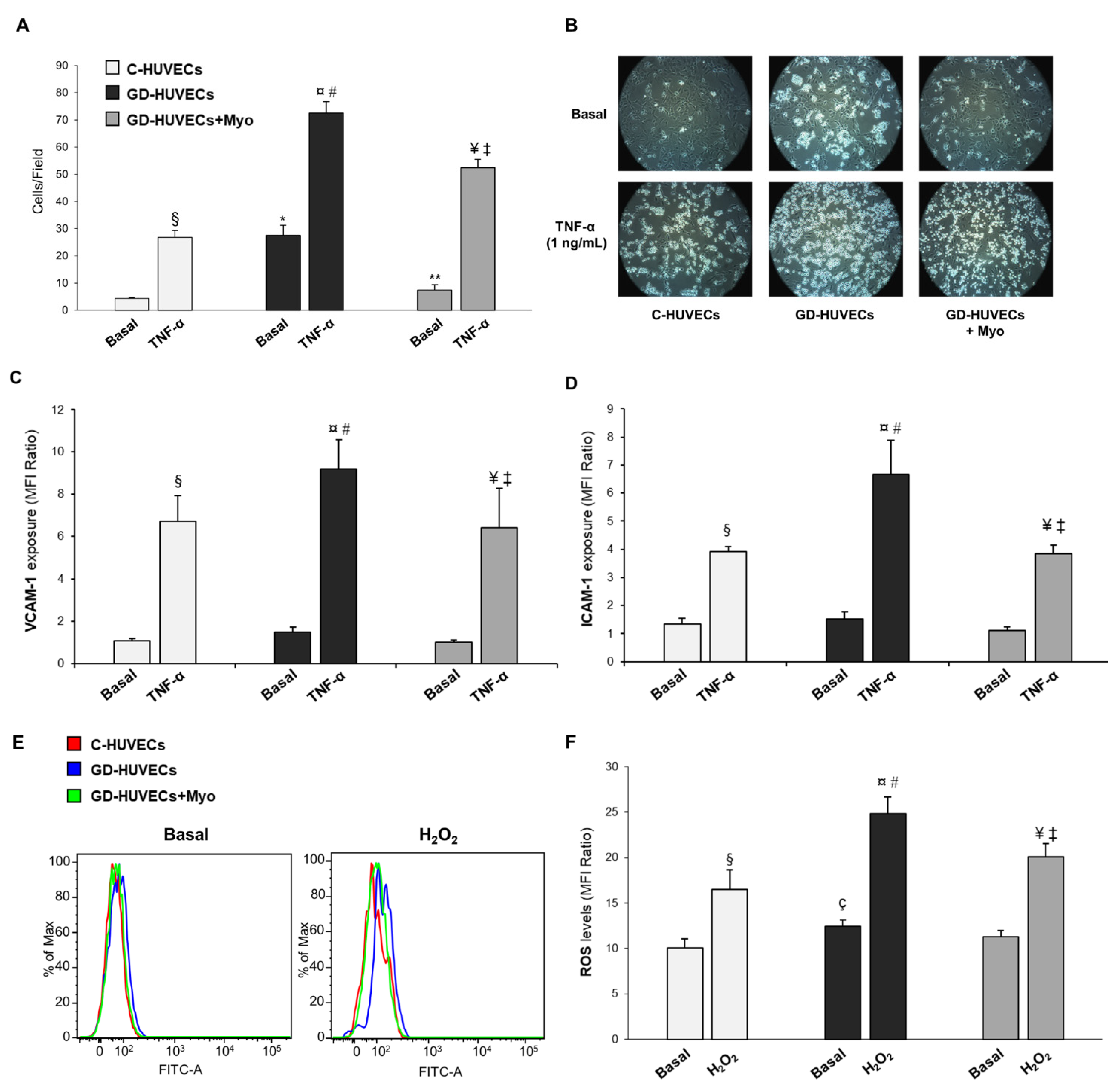
3.1. Effect of Supplementing GD Mothers with Myo during Pregnancy on GD-HUVECs In Vitro Features
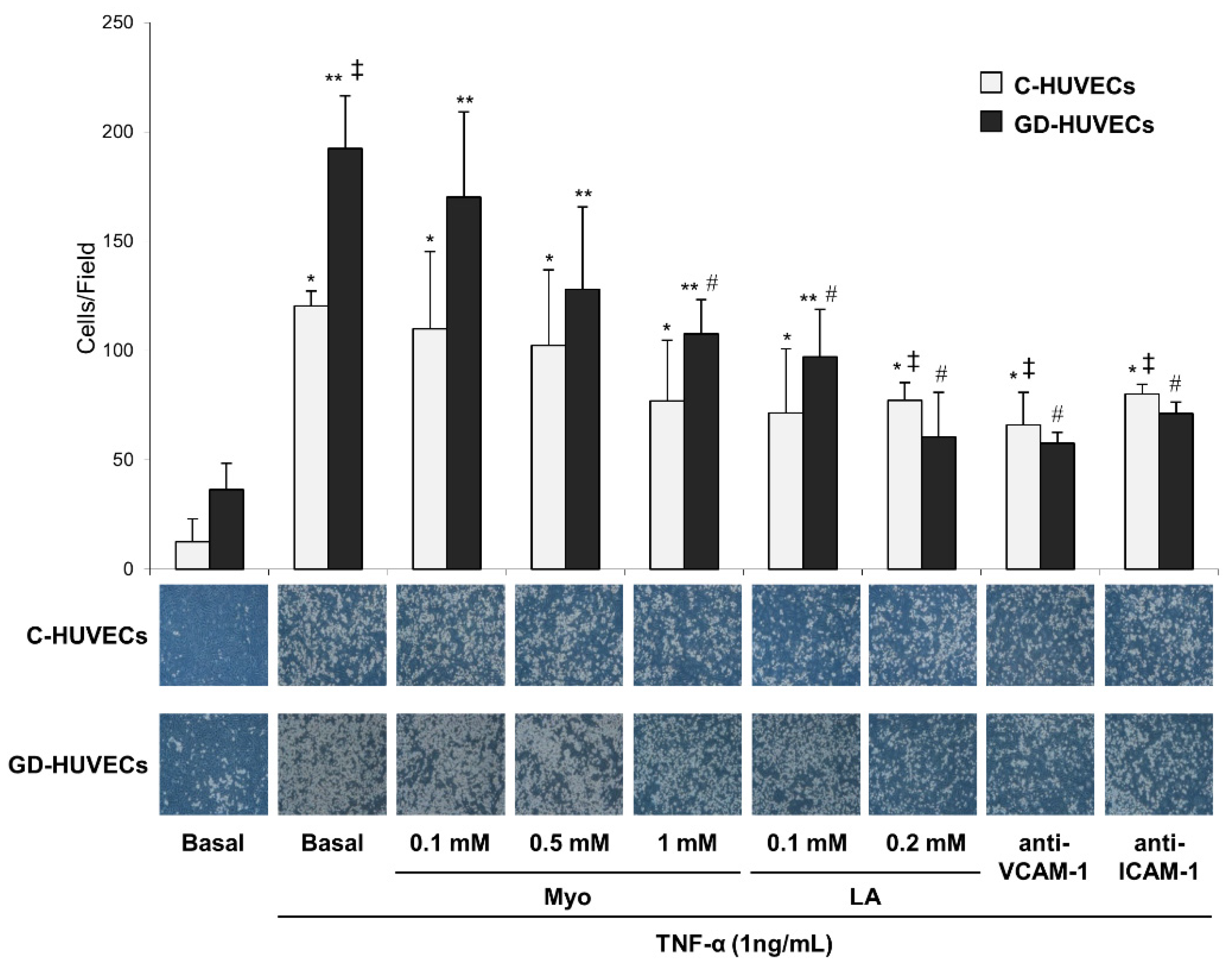
3.2. Effect of Myo on TNF-α Stimulated U937 Monocyte Adhesion to C- and GD-HUVECs
3.3. Effect of Myo on TNF-α Stimulated Adhesion Molecule Membrane Exposure in C- and GD-HUVECs
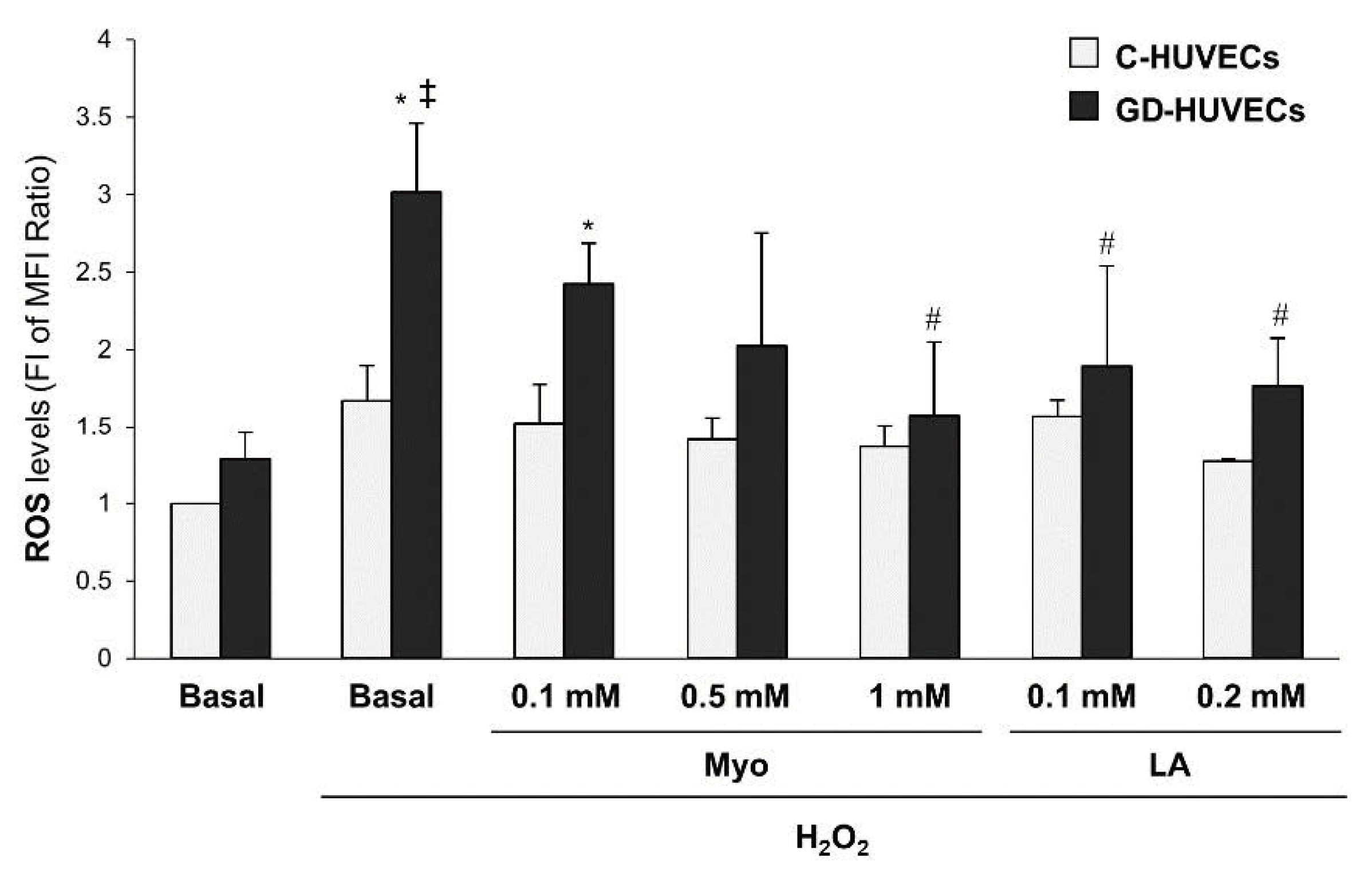
3.4. Effect of Myo on Intracellular ROS Levels in GD HUVECs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Pitocco, D.; Tesauro, M.; Alessandro, R.; Ghirlanda, G.; Cardillo, C. Oxidative Stress in Diabetes: Implications for Vascular and Other Complications. Int. J. Mol. Sci. 2013, 14, 21525–21550. [Google Scholar] [CrossRef] [Green Version]
- Sobrevia, L.; Yudilevich, D.L.; Mann, G.E. Elevated D-glucose induces insulin insensitivity in human umbilical endothelial cells isolated from gestational diabetic pregnancies. J. Physiol. 1998, 506, 219–230. [Google Scholar] [CrossRef]
- Ceriello, A. New Insights on Oxidative Stress and Diabetic Complications May Lead to a “Causal” Antioxidant Therapy. Diabetes Care 2003, 26, 1589–1596. [Google Scholar] [CrossRef] [Green Version]
- Giri, H.; Chandel, S.; Dwarakanath, L.S.; Sreekumar, S.; Dixit, M. Increased Endothelial Inflammation, sTie-2 and Arginase Activity in Umbilical Cords Obtained from Gestational Diabetic Mothers. PLoS ONE 2013, 8, e84546. [Google Scholar] [CrossRef] [Green Version]
- Di Fulvio, P.; Pandolfi, A.; Formoso, G.; Di Silvestre, S.; Di Tomo, P.; Giardinelli, A.; De Marco, A.; Di Pietro, N.; Taraborrelli, M.; Sancilio, S.; et al. Features of endothelial dysfunction in umbilical cord vessels of women with gestational diabetes. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 1337–1345. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, A. Translating Research on Diabetes and Obesity in Pregnancy into Prevention: The 2019 Norbert Freinkel Award Lecture. Diabetes Care 2020, 43, 2635–2642. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association. Management of diabetes in pregnancy: Standards of medical care in diabetes—2020. Diabetes Care 2020, 43, S183–S192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Formoso, G.; Baldassarre, M.P.; Ginestra, F.; Carlucci, M.A.; Bucci, I.; Consoli, A. Inositol and antioxidant supplementation: Safety and efficacy in pregnancy. Diabetes Metab. Res. Rev. 2019, 35, e3154. [Google Scholar] [CrossRef] [PubMed]
- Fraticelli, F.; Celentano, C.; Al Zecca, I.; Di Vieste, G.; Pintaudi, B.; Liberati, M.; Franzago, M.; Di Nicola, M.; Vitacolonna, E. Effect of inositol stereoisomers at different dosages in gestational diabetes: An open-label, parallel, randomized controlled trial. Acta Diabetol. 2018, 55, 805–812. [Google Scholar] [CrossRef]
- D’Anna, R.; Scilipoti, A.; Giordano, D.; Caruso, C.; Cannata, M.L.; Interdonato, M.L.; Corrado, F.; Di Benedetto, A. Myo-Inositol Supplementation and Onset of Gestational Diabetes Mellitus in Pregnant Women With a Family History of Type 2 Diabetes: A prospective, randomized, placebo-controlled study. Diabetes Care 2013, 36, 854–857. [Google Scholar] [CrossRef] [Green Version]
- D’Anna, R.; Di Benedetto, A.; Scilipoti, A.; Santamaria, A.; Interdonato, M.L.; Petrella, E.; Neri, I.; Pintaudi, B.; Corrado, F.; Facchinetti, F. Myo-inositol Supplementation for Prevention of Gestational Diabetes in Obese Pregnant Women: A Randomized Controlled Trial. Obstet. Gynecol. 2015, 126, 310–315. [Google Scholar] [CrossRef] [Green Version]
- Santamaria, A.; Di Benedetto, A.; Petrella, E.; Pintaudi, B.; Corrado, F.; D’Anna, R.; Neri, I.; Facchinetti, F. Myo-inositol may prevent gestational diabetes onset in overweight women: A randomized, controlled trial. J. Matern. Fetal Neonatal Med. 2015, 29, 3234–3237. [Google Scholar] [CrossRef]
- Matarrelli, B.; Vitacolonna, E.; D’Angelo, M.; Pavone, G.; Mattei, P.A.; Liberati, M.; Celentano, C. Effect of dietary myo-inositol supplementation in pregnancy on the incidence of maternal gestational diabetes mellitus and fetal outcomes: A randomized controlled trial. J. Matern. Fetal Neonatal Med. 2013, 26, 967–972. [Google Scholar] [CrossRef] [PubMed]
- Celentano, C.; Matarrelli, B.; Pavone, G.; Vitacolonna, E.; Mattei, P.A.; Berghella, V.; Liberati, M. The influence of different inositol stereoisomers supplementation in pregnancy on maternal gestational diabetes mellitus and fetal outcomes in high-risk patients: A randomized controlled trial. J. Matern. Fetal Neonatal Med. 2018, 33, 743–751. [Google Scholar] [CrossRef]
- Benvenga, S.; Feldt-Rasmussen, U.; Bonofiglio, D.; Asamoah, E. Nutraceutical Supplements in the Thyroid Setting: Health Benefits beyond Basic Nutrition. Nutrients 2019, 11, 2214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, K.Y.; Wong, M.M.H.; Pang, S.S.H.; Lo, K.K.H. Dietary supplementation for gestational diabetes prevention and management: A meta-analysis of randomized controlled trials. Arch. Gynecol. Obstet. 2021, 303, 1381–1391. [Google Scholar] [CrossRef] [PubMed]
- D’Anna, R.; Corrado, F.; Loddo, S.; Gullo, G.; Giunta, L.; Di Benedetto, A. Myoinositol plus α-lactalbumin supplementation, insulin resistance and birth outcomes in women with gestational diabetes mellitus: A randomized, controlled study. Sci. Rep. 2021, 11, 8866. [Google Scholar] [CrossRef] [PubMed]
- Dell’Edera, D.; Sarlo, F.; Allegretti, A.; Simone, F.; Lupo, M.G.; Epifania, A.A. The influence of D-chiro-inositol and D-myo-inositol in pregnant women with glucose intolerance. Biomed. Rep. 2017, 7, 169–172. [Google Scholar] [CrossRef]
- Gambioli, R.; Forte, G.; Buzzaccarini, G.; Unfer, V.; Laganà, A. Myo-Inositol as a Key Supporter of Fertility and Physiological Gestation. Pharmaceutical 2021, 14, 504. [Google Scholar] [CrossRef]
- 17/10/2018—POSITION STATEMENT AMD—SID “Integratori Vitaminici, Inositolo e Probiotici Nelle Donne Con Iperglicemia in Gravidanza”. Available online: http://Www.Siditalia.It/News/2103-17-10-2018-Position-Statement-Amd-Sid-Integratori-Vitaminici-Inositolo-e-Probiotici-N (accessed on 21 May 2021).
- Abdali, D.; Samson, S.E.; Grover, A.K. How Effective Are Antioxidant Supplements in Obesity and Diabetes? Med. Princ. Pract. 2015, 24, 201–215. [Google Scholar] [CrossRef]
- Nascimento, N.R.F.; Lessa, L.M.A.; Kerntopf, M.R.; Sousa, C.M.; Alves, R.S.; Queiroz, M.G.; Price, J.; Heimark, D.B.; Larner, J.; Du, X.; et al. Inositols prevent and reverse endothelial dysfunction in diabetic rat and rabbit vasculature metabolically and by scavenging superoxide. Proc. Natl. Acad. Sci. USA 2006, 103, 218–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Incalza, M.A.; D’Oria, R.; Natalicchio, A.; Perrini, S.; Laviola, L.; Giorgino, F. Oxidative stress and reactive oxygen species in endothelial dysfunction associated with cardiovascular and metabolic diseases. Vasc. Pharmacol. 2018, 100, 1–19. [Google Scholar] [CrossRef]
- Ormazabal, V.; Nair, S.; Elfeky, O.; Aguayo, C.; Salomon, C.; Zuñiga, F.A. Association between insulin resistance and the development of cardiovascular disease. Cardiovasc. Diabetol. 2018, 17, 122. [Google Scholar] [CrossRef]
- SID-AMD. Standard Italiani per La Cura Del Diabete Mellito 2018; Società Italiana di Diabetologia, Associazione Medici Diabetologi: Roma, Italy, 2018. [Google Scholar]
- Di Tomo, P.; Lanuti, P.; Di Pietro, N.; Baldassarre, M.P.A.; Marchisio, M.; Pandolfi, A.; Consoli, A.; Formoso, G. Liraglutide mitigates TNF-α induced pro-atherogenic changes and microvesicle release in HUVEC from diabetic women. Diabetes Metab. Res. Rev. 2017, 33, e2925. [Google Scholar] [CrossRef] [PubMed]
- Di Tomo, P.; Di Silvestre, S.; Cordone, V.; Giardinelli, A.; Faricelli, B.; Pipino, C.; Lanuti, P.; Peng, T.; Formoso, G.; Yang, D.; et al. Centella Asiatica and Lipoic Acid, or a combination thereof, inhibit monocyte adhesion to endothelial cells from umbilical cords of gestational diabetic women. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Di Tomo, P.; Canali, R.; Ciavardelli, D.; Di Silvestre, S.; De Marco, A.; Giardinelli, A.; Pipino, C.; Di Pietro, N.; Virgili, F.; Pandolfi, A. β-Carotene and lycopene affect endothelial response to TNF-α reducing nitro-oxidative stress and interaction with monocytes. Mol. Nutr. Food Res. 2012, 56, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Ucci, M.; Di Tomo, P.; Tritschler, F.; Cordone, V.G.P.; Lanuti, P.; Bologna, G.; Di Silvestre, S.; Di Pietro, N.; Pipino, C.; Mandatori, D.; et al. Anti-inflammatory Role of Carotenoids in Endothelial Cells Derived from Umbilical Cord of Women Affected by Gestational Diabetes Mellitus. Oxidative Med. Cell. Longev. 2019, 2019, 8184656. [Google Scholar] [CrossRef]
- Aslfalah, H.; Jamilian, M.; Rafiei, F.; Khosrowbeygi, A. Reduction in maternal serum values of glucose and gamma-glutamyltransferase after supplementation with alpha-lipoic acid in women with gestational diabetes mellitus. J. Obstet. Gynaecol. Res. 2019, 45, 313–317. [Google Scholar] [CrossRef]
- Aslfalah, H.; Jamilian, M.; Khosrowbeygi, A. Elevation of the adiponectin/leptin ratio in women with gestational diabetes mellitus after supplementation with alpha-lipoic acid. Gynecol. Endocrinol. 2019, 35, 271–275. [Google Scholar] [CrossRef]
- Aslfalah, H.; Jamilian, M.; Ansarihadipour, H.; Abdollahi, M.; Khosrowbeygi, A. Effect of alpha-lipoic acid supplementation on the lipid profile and lipid ratios in women with gestational diabetes mellitus: A clinical trial study. Int. J. Reprod. Biomed. 2020, 18, 1029–1038. [Google Scholar] [CrossRef]
- Fogacci, F.; Rizzo, M.; Krogager, C.; Kennedy, C.; Georges, C.M.; Knežević, T.; Liberopoulos, E.; Vallée, A.; Pérez-Martínez, P.; Wenstedt, E.F.; et al. Safety Evaluation of α-Lipoic Acid Supplementation: A Systematic Review and Meta-Analysis of Randomized Placebo-Controlled Clinical Studies. Antioxidants 2020, 9, 1011. [Google Scholar] [CrossRef] [PubMed]
- Kramer, C.K.; Campbell, S.; Retnakaran, R. Gestational diabetes and the risk of cardiovascular disease in women: A systematic review and meta-analysis. Diabetologia 2019, 62, 905–914. [Google Scholar] [CrossRef] [Green Version]
- Carlomagno, G.; De Grazia, S.; Unfer, V.; Manna, F. Myo-inositol in a new pharmaceutical form: A step forward to a broader clinical use. Expert Opin. Drug Deliv. 2012, 9, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Pillai, R.A.; Islam, M.O.; Selvam, P.; Sharma, N.; Chu, A.H.Y.; Watkins, O.C.; Godfrey, K.M.; Lewis, R.M.; Chan, S.Y. Placental Inositol Reduced in Gestational Diabetes as Glucose Alters Inositol Transporters and IMPA1 Enzyme Expression. J. Clin. Endocrinol. Metab. 2021, 106, e875–e890. [Google Scholar] [CrossRef]
- Chu, A.H.Y.; Tint, M.T.; Chang, H.F.; Wong, G.; Yuan, W.L.; Tull, D.; Nijagal, B.; Narayana, V.K.; Meikle, P.J.; Chang, K.T.E.; et al. High placental inositol content associated with suppressed pro-adipogenic effects of maternal glycaemia in offspring: The GUSTO cohort. Int. J. Obes. 2021, 45, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Kim, J.H.; An, H.S.; Park, K.K.; Kim, B.K.; Park, T. Myo-Inositol restores the inflammation-induced down-regulation of taurine transport by the murine macrophage cell line, RAW 264.7. Life Sci. 2003, 73, 2477–2489. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, C.; Zhang, W.; Zheng, X.; Chen, X. Decreased Insulin Resistance by Myo-Inositol Is Associated with Suppressed Interleukin 6/Phospho-STAT3 Signaling in a Rat Polycystic Ovary Syndrome Model. J. Med. Food 2020, 23, 375–387. [Google Scholar] [CrossRef]
- Laganà, A.S.; Unfer, V.; Garzon, S.; Bizzarri, M. Role of inositol to improve surfactant functions and reduce IL-6 levels: A potential adjuvant strategy for SARS-CoV-2 pneumonia? Med. Hypotheses 2020, 144, 110262. [Google Scholar] [CrossRef]
- Tilton, R.G.; Faller, A.M.; LaRose, L.S.; Burgan, J.; Williamson, J.R. Dietary myo-inositol supplementation does not prevent retinal and glomerular vascular structural changes in chronically diabetic rats. J. Diabetes Complicat. 1993, 7, 188–198. [Google Scholar] [CrossRef]
- D’Oria, R.; Laviola, L.; Giorgino, F.; Unfer, V.; Bettocchi, S.; Scioscia, M. PKB/Akt and MAPK/ERK phosphorylation is highly induced by inositols: Novel potential insights in endothelial dysfunction in preeclampsia. Pregnancy Hypertens. 2017, 10, 107–112. [Google Scholar] [CrossRef]
- Franks, P.W.; Estampador, A.C. Genetic and epigenetic catalysts in early-life programming of adult cardiometabolic disorders. Diabetes Metab. Syndr. Obes. Targets Ther. 2014, 7, 575–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Control (n = 10) | GD (n = 10) | GD+Myo (n = 6) |
|---|---|---|---|
| Age (years) | 34.6 ± 4.93 | 39 ± 8.74 * | 41 ± 5.45 * |
| Pre-gestational weight (kg) | 63.05 ± 14.47 | 67.1 ± 10.73 | 64.75 ± 13.15 |
| Pre-gestational BMI (Kg/m2) | 25.12 ± 4.60 | 26.8 ± 3.10 | 26.2 ± 3.70 |
| Weight at delivery (kg) | 73.05 ± 14.47 | 79.23 ± 16.32 | 77.75 ± 13.18 # |
| Gestational week OGTT | 28.1 ± 2.85 | 25 ± 2.64 * | 25 ± 8.08 * |
| Glycemia T0 (mmol/L) | 4.5 ± 0.24 | 5.1 ± 0.24 * | 5.2 ± 1.26 * |
| Glycemia T60 (mmol/L) | 8.1 ± 0.99 | 10.2 ± 1.16 * | 11.3 ± 1.59 * |
| Glycemia T120 (mmol/L) | 6.54 ± 1.14 | 8.04 ± 1.71 * | 9.8 ± 1.51 * |
| Systolic BP (mmHg) | 110 ± 20 | 100 ± 28.28 | 110 ± 8.54 |
| Diastolic BP (mmHg) | 75 ± 15 | 75 ± 21.21 | 70 ± 7.07 |
| Treatment | Diet | Diet | Diet + Myo (1500 ± 500 mg) |
| Gestational week at delivery | 38.7 ± 1.40 | 38.5 ± 1.52 | 38.5 ± 1.60 |
| Birth Weight (g) | 3408.50 ± 589.50 | 3490.30 ± 501.50 | 3475.35 ± 473.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldassarre, M.P.A.; Di Tomo, P.; Centorame, G.; Pandolfi, A.; Di Pietro, N.; Consoli, A.; Formoso, G. Myoinositol Reduces Inflammation and Oxidative Stress in Human Endothelial Cells Exposed In Vivo to Chronic Hyperglycemia. Nutrients 2021, 13, 2210. https://doi.org/10.3390/nu13072210
Baldassarre MPA, Di Tomo P, Centorame G, Pandolfi A, Di Pietro N, Consoli A, Formoso G. Myoinositol Reduces Inflammation and Oxidative Stress in Human Endothelial Cells Exposed In Vivo to Chronic Hyperglycemia. Nutrients. 2021; 13(7):2210. https://doi.org/10.3390/nu13072210
Chicago/Turabian StyleBaldassarre, Maria Pompea Antonia, Pamela Di Tomo, Giorgia Centorame, Assunta Pandolfi, Natalia Di Pietro, Agostino Consoli, and Gloria Formoso. 2021. "Myoinositol Reduces Inflammation and Oxidative Stress in Human Endothelial Cells Exposed In Vivo to Chronic Hyperglycemia" Nutrients 13, no. 7: 2210. https://doi.org/10.3390/nu13072210
APA StyleBaldassarre, M. P. A., Di Tomo, P., Centorame, G., Pandolfi, A., Di Pietro, N., Consoli, A., & Formoso, G. (2021). Myoinositol Reduces Inflammation and Oxidative Stress in Human Endothelial Cells Exposed In Vivo to Chronic Hyperglycemia. Nutrients, 13(7), 2210. https://doi.org/10.3390/nu13072210