
Food Enrichment with Glycyrrhiza glabra Extract Suppresses ACE2 mRNA and Protein Expression in Rats—Possible Implications for COVID-19



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Study Design
2.3. Treatment
2.4. Organ Collection
2.5. ACE2 mRNA and Protein Quantification
2.6. Hormone Measurements
2.7. Statistical Analyses
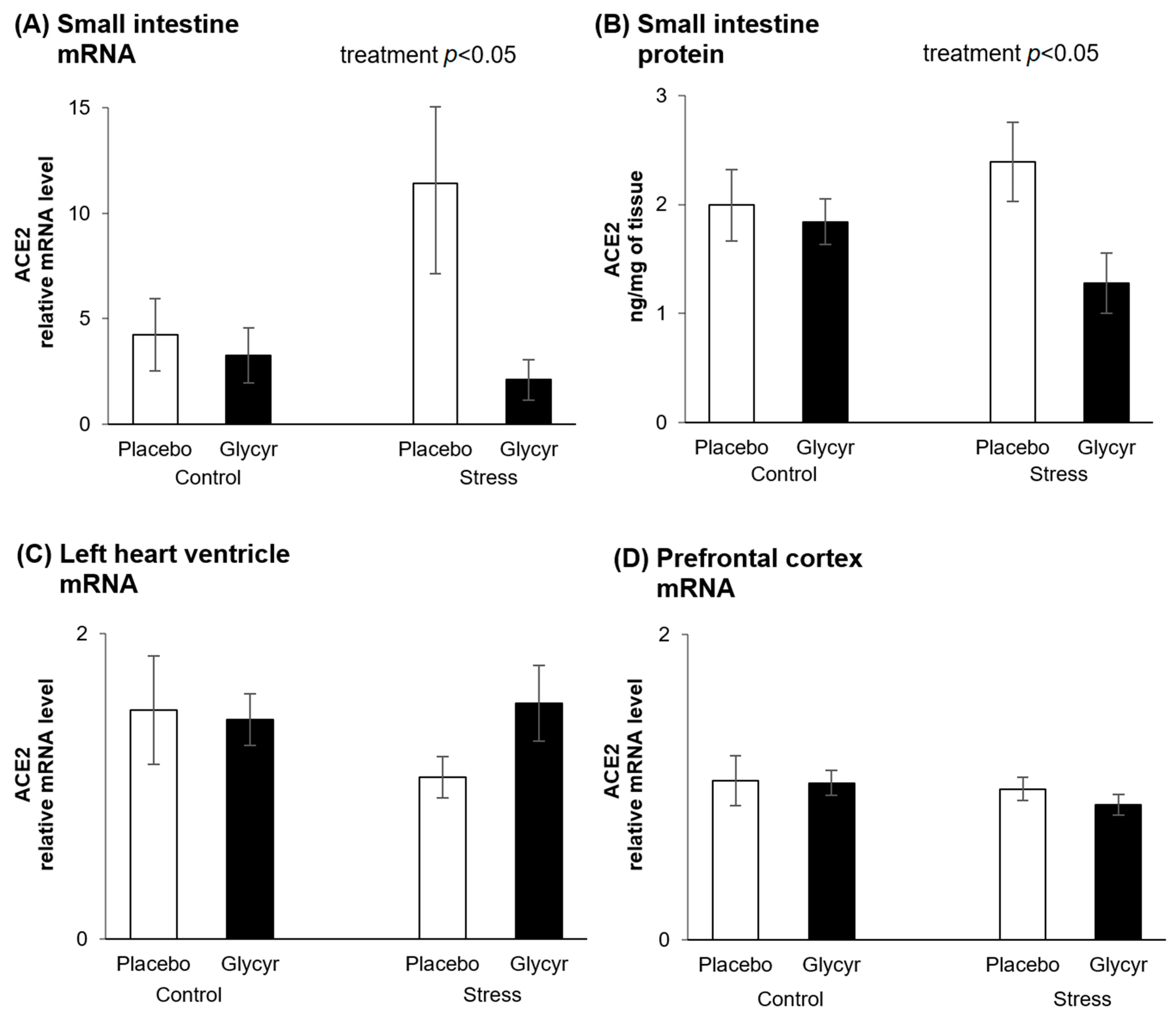
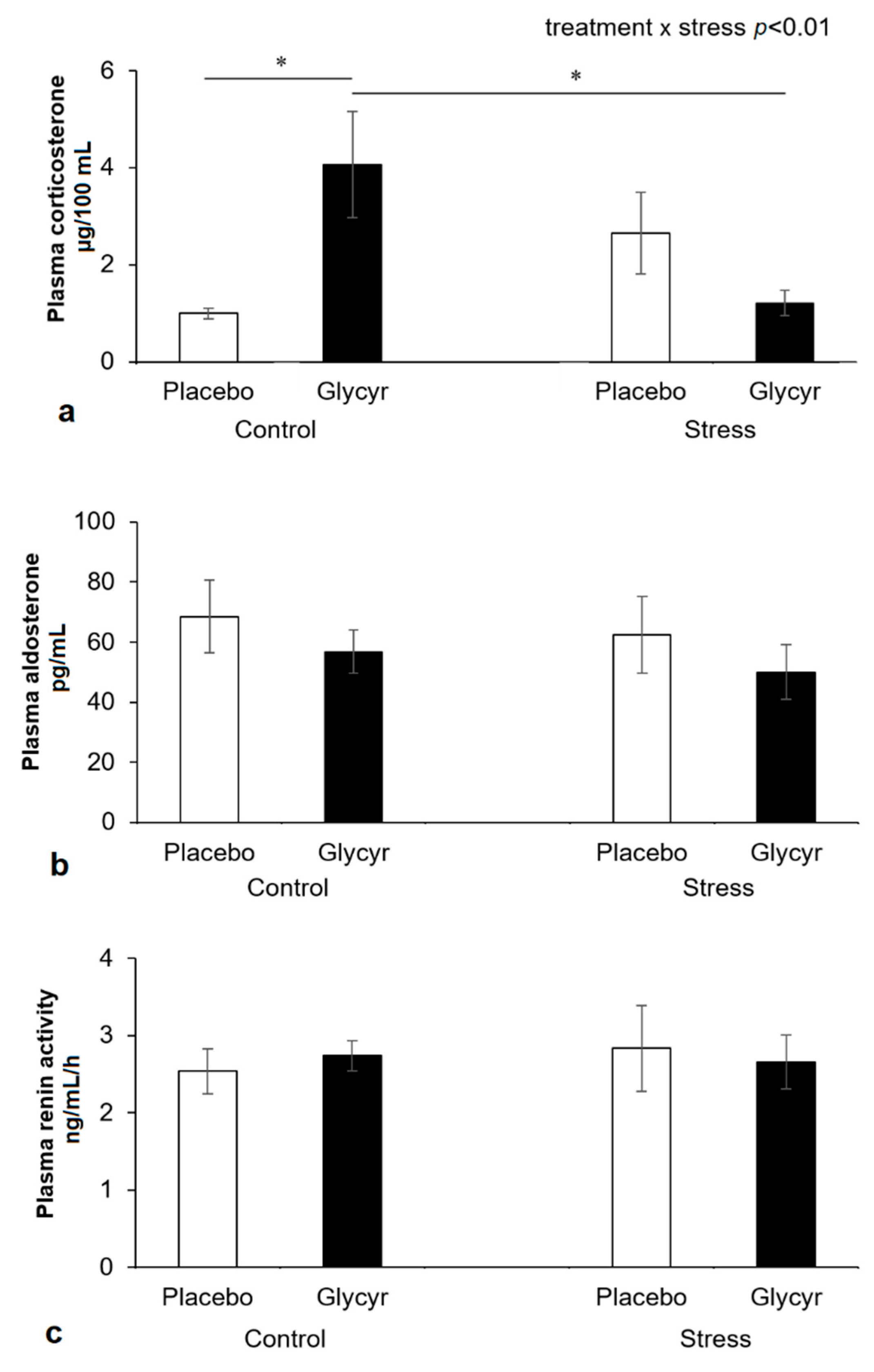
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cawood, A.L.; Walters, E.R.; Smith, T.R.; Sipaul, R.H.; Stratton, R.J. A Review of Nutrition Support Guidelines for Individuals with or Recovering from COVID-19 in the Community. Nutrients 2020, 12, 3230. [Google Scholar] [CrossRef]
- Hensel, A.; Bauer, R.; Heinrich, M.; Spiegler, V.; Kayser, O.; Hempel, G.; Kraft, K. Challenges at the Time of COVID-19: Opportunities and Innovations in Antivirals from Nature. Planta Medica 2020, 86, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, C.G.K.; Allon, S.J.; Nyquist, S.K.; Mbano, I.M.; Miao, V.N.; Tzouanas, C.N.; Cao, Y.; Yousif, A.S.; Bals, J.; Hauser, B.M.; et al. SARS-CoV-2 Receptor ACE2 Is an Interferon-Stimulated Gene in Human Airway Epithelial Cells and Is Detected in Specific Cell Subsets across Tissues. Cell 2020, 181, 1016–1035.e19. [Google Scholar] [CrossRef]
- Murck, H. Symptomatic Protective Action of Glycyrrhizin (Licorice) in COVID-19 Infection? Front. Immunol. 2020, 11, 1239. [Google Scholar] [CrossRef]
- Huan, C.; Chen, C.; Xu, W.; Guo, T.; Pan, H.; Gao, S. Study on Antiviral Activities of Glycyrrhizin. Int. J. Biomed. Eng. Clin. Sci. 2020, 6, 68–70. [Google Scholar] [CrossRef]
- Gomaa, A.; Abdel-Wadood, Y. The potential of glycyrrhizin and licorice extract in combating COVID-19 and associated conditions. Phytomedicine Plus 2021, 1, 100043. [Google Scholar] [CrossRef]
- Li, F.; Liu, B.; Li, T.; Wu, Q.; Xu, Z.; Gu, Y.; Li, W.; Wang, P.; Ma, T.; Lei, H. Review of Constituents and Biological Activities of Triterpene Saponins from Glycyrrhizae Radix et Rhizoma and Its Solubilization Characteristics. Molecules 2020, 25, 3904. [Google Scholar] [CrossRef]
- Cosmetic Ingredient Review Expert Panel. Final report on the safety assessment of Glycyrrhetinic Acid, Potassium Glycyrrhetinate, Disodium Succinoyl Glycyrrhetinate, Glyceryl Glycyrrhetinate, Glycyrrhetinyl Stearate, Stearyl Glycyrrhetinate, Glycyrrhizic Acid, Ammonium Glycyrrhizate, Dipotassium Glycyrrhizate, Disodium Glycyrrhizate, Trisodium Glycyrrhizate, Methyl Glycyrrhizate, and Potassium Glycyrrhizinate. Int. J. Toxicol. 2007, 26, 79–112. [Google Scholar] [CrossRef]
- Esler, M.; Esler, D. Can angiotensin receptor-blocking drugs perhaps be harmful in the COVID-19 pandemic? J. Hypertens. 2020. [Google Scholar] [CrossRef]
- Fukuda, S.; Horimai, C.; Harada, K.; Wakamatsu, T.; Fukasawa, H.; Muto, S.; Itai, A.; Hayashi, M. Aldosterone-induced kidney injury is mediated by NFkappaB activation. Clin. Exp. Nephrol. 2011, 15, 41–49. [Google Scholar] [CrossRef]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef] [PubMed]
- Gheblawi, M.; Wang, K.; Viveiros, A.; Nguyen, Q.; Zhong, J.C.; Turner, A.J.; Raizada, M.K.; Grant, M.B.; Oudit, G.Y. Angiotensin-Converting Enzyme 2: SARS-CoV-2 Receptor and Regulator of the Renin-Angiotensin System: Celebrating the 20th Anniversary of the Discovery of ACE2. Circ. Res. 2020, 126, 1456–1474. [Google Scholar] [CrossRef]
- Kaparianos, A.; Argyropoulou, E. Local renin-angiotensin II systems, angiotensin-converting enzyme and its homologue ACE2: Their potential role in the pathogenesis of chronic obstructive pulmonary diseases, pulmonary hypertension and acute respiratory distress syndrome. Curr. Med. Chem. 2011, 18, 3506–3515. [Google Scholar] [CrossRef]
- Tikellis, C.; Thomas, M.C. Angiotensin-Converting Enzyme 2 (ACE2) Is a Key Modulator of the Renin Angiotensin System in Health and Disease. Int. J. Pept. 2012, 2012, 256294. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.; Liu, Z. ACE2 exhibits protective effects against LPS-induced acute lung injury in mice by inhibiting the LPS-TLR4 pathway. Exp. Mol. Pathol. 2020, 113, 104350. [Google Scholar] [CrossRef]
- Ingraham, N.E.; Lotfi-Emran, S.; Thielen, B.K.; Techar, K.; Morris, R.S.; Holtan, S.G.; Dudley, R.A.; Tignanelli, C.J. Immunomodulation in COVID-19. Lancet Respir. Med. 2020. [Google Scholar] [CrossRef]
- Armanini, D.; Lewicka, S.; Pratesi, C.; Scali, M.; Zennaro, M.C.; Zovato, S.; Gottardo, C.; Simoncini, M.; Spigariol, A.; Zampollo, V. Further studies on the mechanism of the mineralocorticoid action of licorice in humans. J. Endocrinol. Investig. 1996, 19, 624–629. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, M.A.; Hoefnagels, W.H.; Jansen, R.W.; Benraad, T.J.; Kloppenborg, P.W. The influence of glycyrrhetinic acid on plasma cortisol and cortisone in healthy young volunteers. J. Clin. Endocrinol. Metab. 1990, 70, 1637–1643. [Google Scholar] [CrossRef]
- Benicky, J.; Sanchez-Lemus, E.; Honda, M.; Pang, T.; Orecna, M.; Wang, J.; Leng, Y.; Chuang, D.M.; Saavedra, J.M. Angiotensin II AT1 receptor blockade ameliorates brain inflammation. Neuropsychopharmacology 2011, 36, 857–870. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Ohtaki, Y.; Kai, K.; Sasano, T.; Shimauchi, H.; Yokochi, T.; Takada, H.; Sugawara, S.; Kumagai, K.; Endo, Y. Critical roles of platelets in lipopolysaccharide-induced lethality: Effects of glycyrrhizin and possible strategy for acute respiratory distress syndrome. Int. Immunopharmacol. 2005, 5, 571–580. [Google Scholar] [CrossRef]
- Seo, E.H.; Song, G.Y.; Kwak, B.O.; Oh, C.S.; Lee, S.H.; Kim, S.H. Effects of Glycyrrhizin on the Differentiation of Myeloid Cells of the Heart and Lungs in Lipopolysaccharide-Induced Septic Mice. Shock 2017, 48, 371–376. [Google Scholar] [CrossRef]
- Duncko, R.; Kiss, A.; Skultetyova, I.; Rusnak, M.; Jezova, D. Corticotropin-releasing hormone mRNA levels in response to chronic mild stress rise in male but not in female rats while tyrosine hydroxylase mRNA levels decrease in both sexes. Psychoneuroendocrinology 2001, 26, 77–89. [Google Scholar] [CrossRef]
- Chakravarthi, K.K.; Avadhani, R. Beneficial effect of aqueous root extract of Glycyrrhiza glabra on learning and memory using different behavioral models: An experimental study. J. Nat. Sci. Biol. Med. 2013, 4, 420–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buzgoova, K.; Graban, J.; Balagova, L.; Hlavacova, N.; Jezova, D. Brain derived neurotrophic factor expression and DNA methylation in response to subchronic valproic acid and/or aldosterone treatment. Croat. Med. J. 2019, 60, 71–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, R.M.; Vinetz, J.M. Dexamethasone in the management of covid -19. BMJ 2020, 370, m2648. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Gong, L.; Wang, B.; Wu, Y.; Wang, Y.; Mei, X.; Xu, H.; Tang, L.; Liu, R.; Zeng, Z.; et al. Glycyrrhizin Attenuates Salmonella enterica Serovar Typhimurium Infection: New Insights into Its Protective Mechanism. Front. Immunol. 2018, 9, 2321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, D.J.; Ridlon, J.M. Glucocorticoids and gut bacteria: “The GALF Hypothesis” in the metagenomic era. Steroids 2017, 125, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Richards, E.M.; Pepine, C.J.; Raizada, M.K.; Kim, S. The Gut, Its Microbiome, and Hypertension. Curr. Hypertens. Rep. 2017, 19, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, G.A.; White, C.I.; Castellan, R.F.; McSweeney, S.J.; Chapman, K.E. Getting to the heart of intracellular glucocorticoid regeneration: 11beta-HSD1 in the myocardium. J. Mol. Endocrinol. 2017, 58, R1–R13. [Google Scholar] [CrossRef] [Green Version]
- Wyrwoll, C.S.; Holmes, M.C.; Seckl, J.R. 11beta-hydroxysteroid dehydrogenases and the brain: From zero to hero, a decade of progress. Front. Neuroendocrinol. 2011, 32, 265–286. [Google Scholar] [CrossRef] [Green Version]
- Shahzad, F.; Anderson, D.; Najafzadeh, M. The Antiviral, Anti-Inflammatory Effects of Natural Medicinal Herbs and Mushrooms and SARS-CoV-2 Infection. Nutrients 2020, 12, 2573. [Google Scholar] [CrossRef]
- Silveira, D.; Prieto-Garcia, J.M.; Boylan, F.; Estrada, O.; Fonseca-Bazzo, Y.M.; Jamal, C.M.; Magalhaes, P.O.; Pereira, E.O.; Tomczyk, M.; Heinrich, M. COVID-19: Is There Evidence for the Use of Herbal Medicines as Adjuvant Symptomatic Therapy? Front. Pharmacol. 2020, 11, 581840. [Google Scholar] [CrossRef]
- Zhong, L.L.D.; Lam, W.C.; Yang, W.; Chan, K.W.; Sze, S.C.W.; Miao, J.; Yung, K.K.L.; Bian, Z.; Wong, V.T. Potential Targets for Treatment of Coronavirus Disease 2019 (COVID-19): A Review of Qing-Fei-Pai-Du-Tang and Its Major Herbs. Am. J. Chin. Med. 2020, 48, 1051–1071. [Google Scholar] [CrossRef]
- Jalali, A.; Dabaghian, F.; Akbrialiabad, H.; Foroughinia, F.; Zarshenas, M.M. A pharmacology-based comprehensive review on medicinal plants and phytoactive constituents possibly effective in the management of COVID-19. Phytother. Res. 2021, 35, 1925–1938. [Google Scholar] [CrossRef] [PubMed]
- Pastor, N.; Collado, M.C.; Manzoni, P. Phytonutrient and Nutraceutical Action against COVID-19: Current Review of Characteristics and Benefits. Nutrients 2021, 13, 464. [Google Scholar] [CrossRef] [PubMed]
- Merarchi, M.; Dudha, N.; Das, B.; Garg, M. Natural products and phytochemicals as potential anti-SARS-CoV-2 drugs. Phytother. Res. 2021. [Google Scholar] [CrossRef]
- Wu, C.Y.; Lin, Y.S.; Yang, Y.H.; Shu, L.H.; Cheng, Y.C.; Liu, H.T. Potential Simultaneous Inhibitors of Angiotensin-Converting Enzyme 2 and Transmembrane Protease, Serine 2. Front. Pharmacol. 2020, 11, 584158. [Google Scholar] [CrossRef]
- Li, Y.; Chu, F.; Li, P.; Johnson, N.; Li, T.; Wang, Y.; An, R.; Wu, D.; Chen, J.; Su, Z.; et al. Potential effect of Maxing Shigan decoction against coronavirus disease 2019 (COVID-19) revealed by network pharmacology and experimental verification. J. Ethnopharmacol. 2021, 271, 113854. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Qiu, Q.; Li, M.; Lin, H.; Cao, S.; Wang, Q.; Chen, Z.; Jiang, W.; Zhang, W.; Huang, Y.; et al. Chemical composition and pharmacological mechanism of ephedra-glycyrrhiza drug pair against coronavirus disease 2019 (COVID-19). Aging 2021, 13, 4811–4830. [Google Scholar] [CrossRef]
- Van de Sand, L.; Bormann, M.; Alt, M.; Schipper, L.; Heilingloh, C.S.; Steinmann, E.; Todt, D.; Dittmer, U.; Elsner, C.; Witzke, O.; et al. Glycyrrhizin Effectively Inhibits SARS-CoV-2 Replication by Inhibiting the Viral Main Protease. Viruses 2021, 13, 609. [Google Scholar] [CrossRef]
- Klempa, B.; Slavikova, M.; Murck, H.; Jezova, D. Direct inhibitory effects of glycyrrhizin on isolated human SARS-CoV-2. Psychiatrie 2021, 25, 6. [Google Scholar]
- Gowda, P.; Patrick, S.; Joshi, S.D.; Kumawat, R.K.; Sen, E. Glycyrrhizin prevents SARS-CoV-2 S1 and Orf3a induced high mobility group box 1 (HMGB1) release and inhibits viral replication. Cytokine 2021, 142, 155496. [Google Scholar] [CrossRef]
- Ding, H.; Deng, W.; Ding, L.; Ye, X.; Yin, S.; Huang, W. Glycyrrhetinic acid and its derivatives as potential alternative medicine to relieve symptoms in nonhospitalized COVID-19 patients. J. Med. Virol. 2020, 92, 2200–2204. [Google Scholar] [CrossRef]
- Luo, H.; Tang, Q.L.; Shang, Y.X.; Liang, S.B.; Yang, M.; Robinson, N.; Liu, J.P. Can Chinese Medicine Be Used for Prevention of Corona Virus Disease 2019 (COVID-19)? A Review of Historical Classics, Research Evidence and Current Prevention Programs. Chin. J. Integr. Med. 2020, 26, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Tsubochi, H.; Darnel, A.; Suzuki, T.; Sasano, H.; Krozowski, Z.S.; Kondo, T. Expression of 11 beta-hydroxysteroid dehydrogenase type 1 in alveolar epithelial cells in rats. Endocr. J. 2003, 50, 445–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Sasano, H.; Suzuki, S.; Hirasawa, G.; Takeyama, J.; Muramatsu, Y.; Date, F.; Nagura, H.; Krozowski, Z.S. 11Beta-hydroxysteroid dehydrogenase type 2 in human lung: Possible regulator of mineralocorticoid action. J. Clin. Endocrinol. Metab. 1998, 83, 4022–4025. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Tsubochi, H.; Ishibashi, H.; Suzuki, T.; Kondo, T.; Sasano, H. Increased expression of 11 beta-hydroxysteroid dehydrogenase type 2 in the lungs of patients with acute respiratory distress syndrome. Pathol. Int. 2003, 53, 751–756. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Tsubochi, H.; Suzuki, T.; Darnel, A.D.; Krozowski, Z.S.; Sasano, H.; Kondo, T. Modulation of transalveolar fluid absorption by endogenous aldosterone in adult rats. Exp. Lung. Res. 2001, 27, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Archontakis Barakakis, P.; Palaiodimos, L.; Fleitas Sosa, D.; Benes, L.; Gulani, P.; Fein, D. Combination of low-dose glucocorticosteroids and mineralocorticoids as adjunct therapy for adult patients with septic shock: A systematic review and meta-analysis of randomized trials and observational studies. Avicenna. J. Med. 2019, 9, 134–142. [Google Scholar] [CrossRef]
- Cheung, K.S.; Hung, I.F.N.; Chan, P.P.Y.; Lung, K.C.; Tso, E.; Liu, R.; Ng, Y.Y.; Chu, M.Y.; Chung, T.W.H.; Tam, A.R.; et al. Gastrointestinal Manifestations of SARS-CoV-2 Infection and Virus Load in Fecal Samples From a Hong Kong Cohort: Systematic Review and Meta-analysis. Gastroenterology 2020, 159, 81–95. [Google Scholar] [CrossRef]
- Guo, M.; Tao, W.; Flavell, R.A.; Zhu, S. Potential intestinal infection and faecal-oral transmission of SARS-CoV-2. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Cuicchi, D.; Lazzarotto, T.; Poggioli, G. Fecal-oral transmission of SARS-CoV-2: Review of laboratory-confirmed virus in gastrointestinal system. Int. J. Colorectal. Dis. 2021, 36, 437–444. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene | Sense | Sequence 5′→3′ |
|---|---|---|
| ACE2 | Forward | ACCCTTCTTACATCAGCCCTACTG |
| Reverse | TGTCCAAAACCTACCCCACATAT | |
| UQCRFS1—reference gene | Forward | ACAGTGGGCCTGAATGTTCC |
| Reverse | CACGGCGATAGTCAGAGAAGTC | |
| TfR1—reference gene | Forward | ATACGTTCCCCGTTGTTGAGG |
| Reverse | GGCGGAAACTGAGTATGGTTGA | |
| HPRT1—reference gene | Forward | CGTCGTGATTAGTGATGATGAAC |
| Reverse | CAAGTCTTTCAGTCCTGTCCATAA |
| Food Intake (g) | Group | Statistical Significance | |||
|---|---|---|---|---|---|
| Control | Stress | ||||
| Treatment Day | Placebo | Glycyr | Placebo | Glycyr | |
| 1–2 | 62.6 ± 1.2 | 65.2 ± 0.9 | 56.3 ± 0.6 | 59.8 ± 1.1 | Treatment N.S. Stress p < 0.001 time p < 0.001 |
| 8–9 | 62.7 ± 0.9 | 61.7 ± 1.1 | 57.3 ± 0.8 | 53.8 ± 1.2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jezova, D.; Karailiev, P.; Karailievova, L.; Puhova, A.; Murck, H. Food Enrichment with Glycyrrhiza glabra Extract Suppresses ACE2 mRNA and Protein Expression in Rats—Possible Implications for COVID-19. Nutrients 2021, 13, 2321. https://doi.org/10.3390/nu13072321
Jezova D, Karailiev P, Karailievova L, Puhova A, Murck H. Food Enrichment with Glycyrrhiza glabra Extract Suppresses ACE2 mRNA and Protein Expression in Rats—Possible Implications for COVID-19. Nutrients. 2021; 13(7):2321. https://doi.org/10.3390/nu13072321
Chicago/Turabian StyleJezova, Daniela, Peter Karailiev, Lucia Karailievova, Agnesa Puhova, and Harald Murck. 2021. "Food Enrichment with Glycyrrhiza glabra Extract Suppresses ACE2 mRNA and Protein Expression in Rats—Possible Implications for COVID-19" Nutrients 13, no. 7: 2321. https://doi.org/10.3390/nu13072321
APA StyleJezova, D., Karailiev, P., Karailievova, L., Puhova, A., & Murck, H. (2021). Food Enrichment with Glycyrrhiza glabra Extract Suppresses ACE2 mRNA and Protein Expression in Rats—Possible Implications for COVID-19. Nutrients, 13(7), 2321. https://doi.org/10.3390/nu13072321