
Promising Anticancer Activities of Alismatis rhizome and Its Triterpenes via p38 and PI3K/Akt/mTOR Signaling Pathways


Abstract
:
1. Introduction
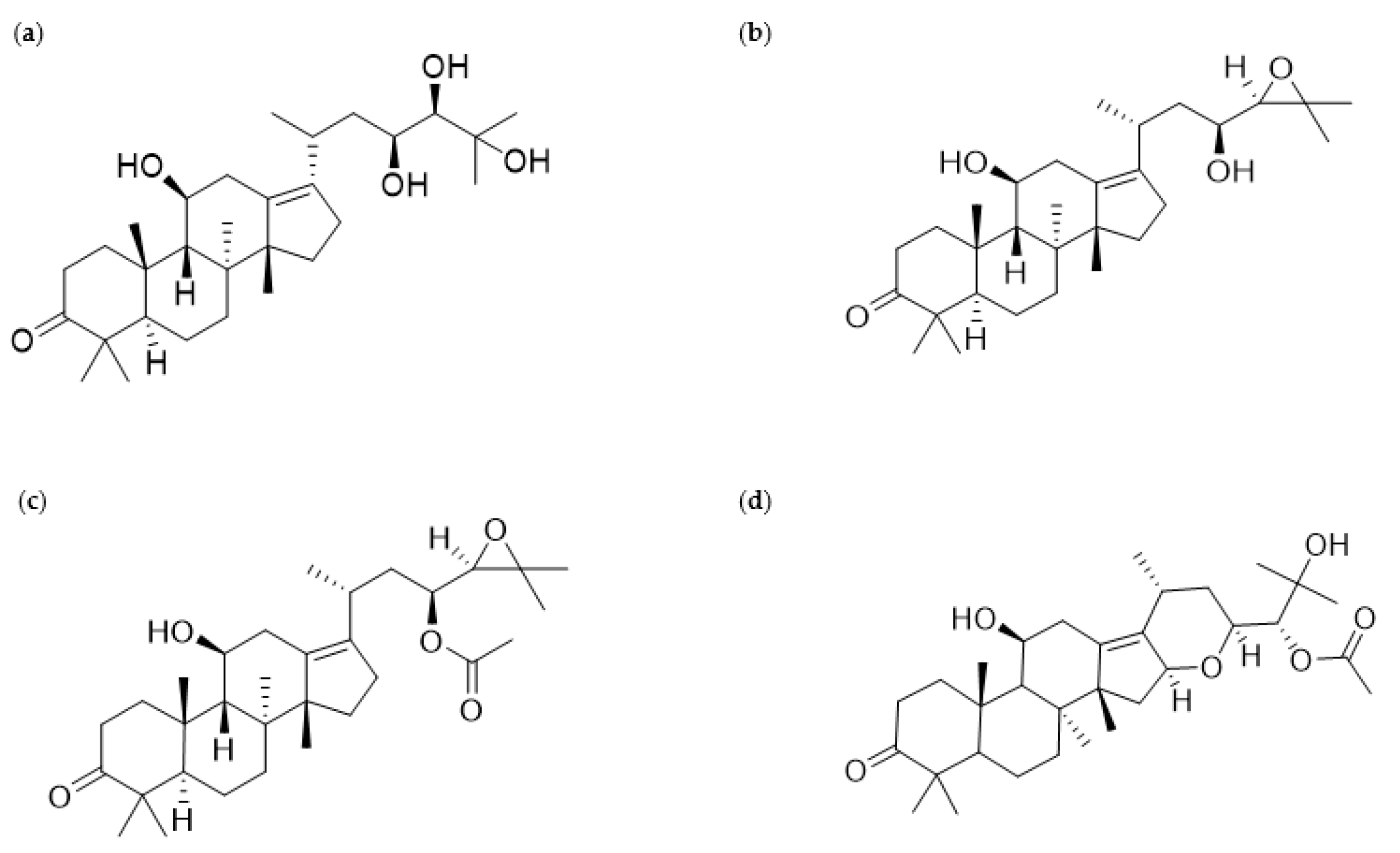
2. Protostane-Type Triterpenes from AR
3. Methodology
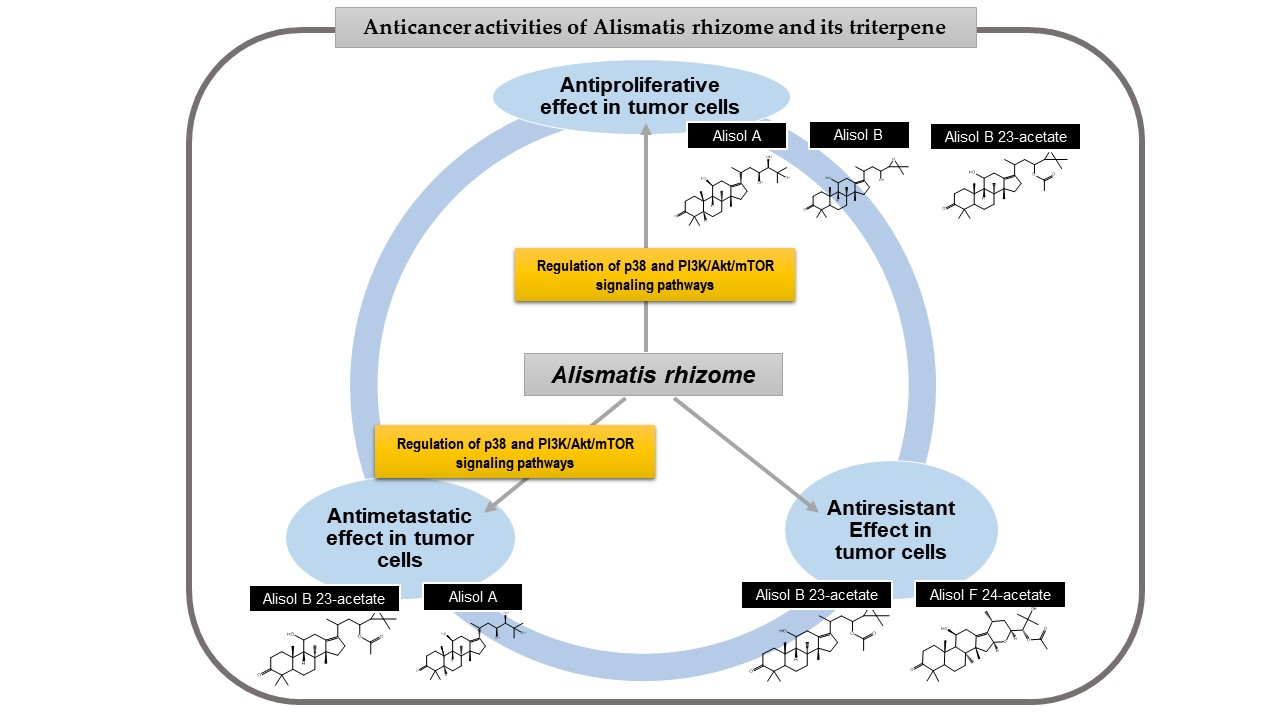
4. Anticancer Properties of AR and Its Triterpenes
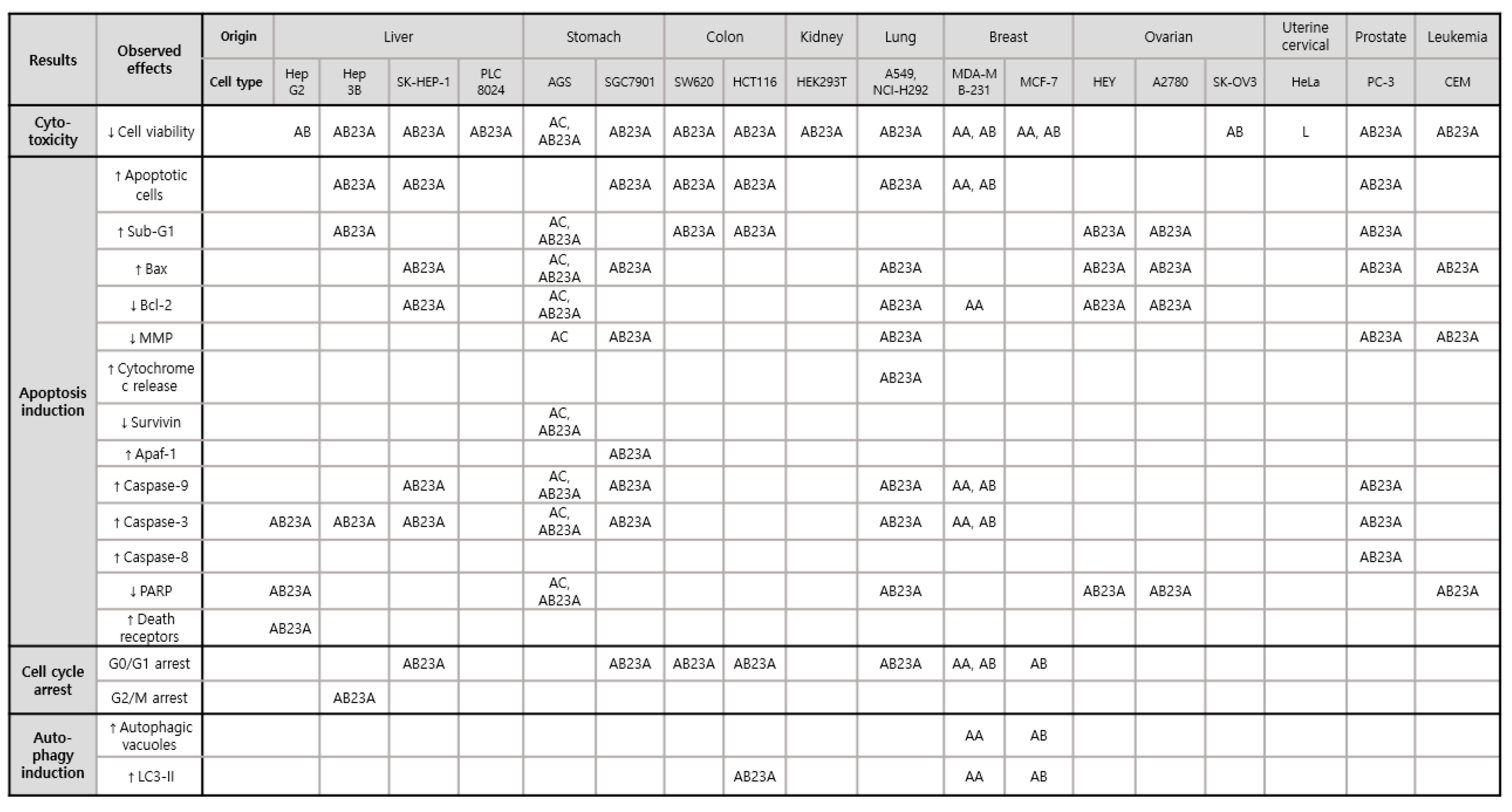
4.1. Antiproliferative Effects
4.2. Antimigratory and Anti-Invasive Effefcts
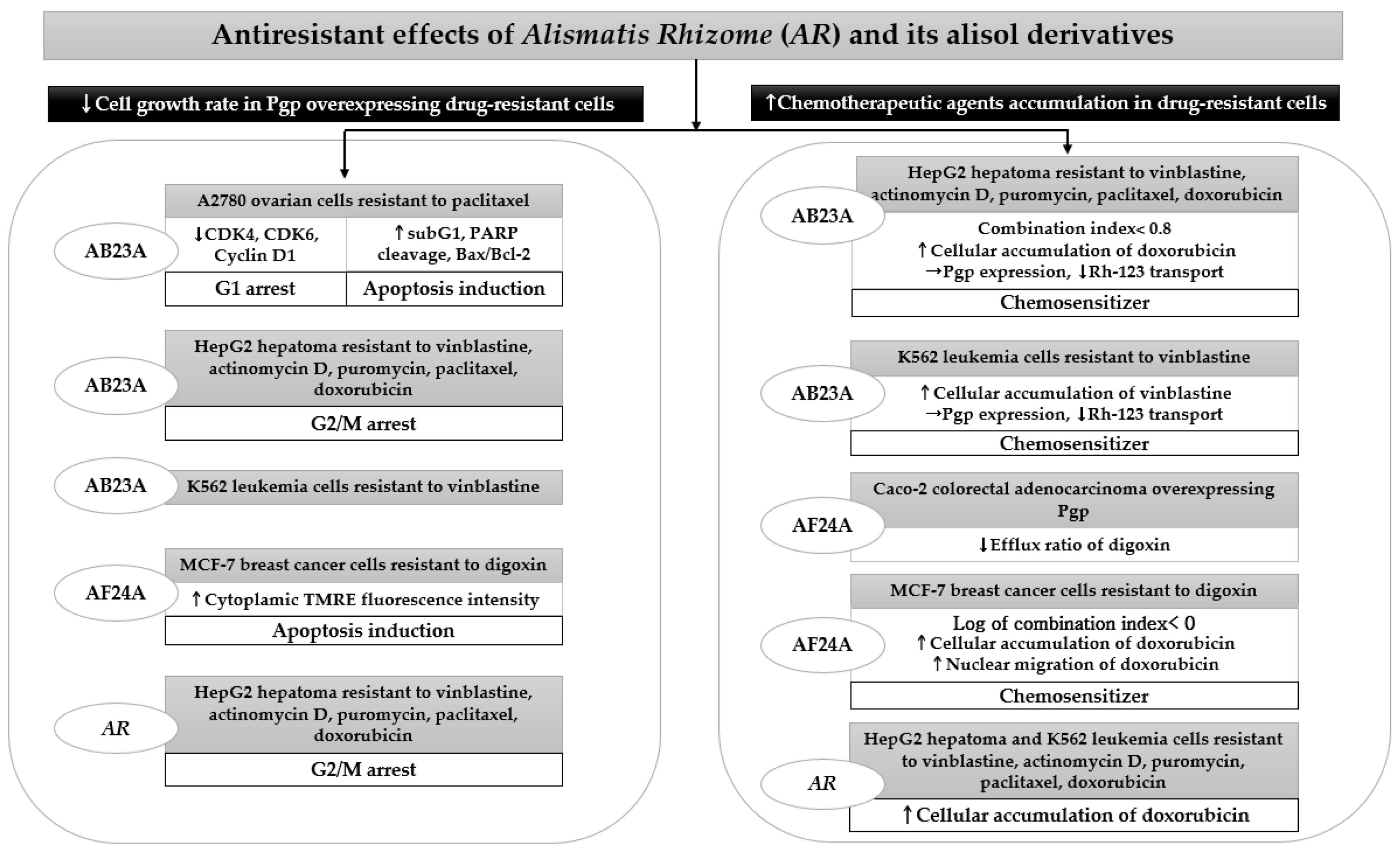
4.3. Antiresistant Effects
5. The p38 and the Phosphoinositide 3-Kinase-AKT-Mammalian Target of Rapamycin (PI3K/AKT/mTOR) Pathway
6. Discussion
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, N.J.; Cellurale, C.; Davis, R.J. A radical role for p38 MAPK in tumor initiation. Cancer Cell 2007, 11, 101–103. [Google Scholar] [CrossRef] [Green Version]
- LoRusso, P.M. Inhibition of the PI3K/AKT/mTOR pathway in solid tumors. J. Clin. Oncol. 2016, 34, 3803–3815. [Google Scholar] [CrossRef]
- Patnaik, A.; Haluska, P.; Tolcher, A.W.; Erlichman, C.; Papadopoulos, K.P.; Lensing, J.L.; Beeram, M.; Molina, J.R.; Rasco, D.W.; Arcos, R.R. A first-in-human phase I study of the oral p38 MAPK inhibitor, ralimetinib (LY2228820 Dimesylate), in patients with advanced cancer. Clin. Cancer Res. 2016, 22, 1095–1102. [Google Scholar] [CrossRef] [Green Version]
- Polivka, J., Jr.; Janku, F. Molecular targets for cancer therapy in the PI3K/AKT/mTOR pathway. Pharmacol. Ther. 2014, 142, 164–175. [Google Scholar] [CrossRef]
- Martínez-Limón, A.; Joaquin, M.; Caballero, M.; Posas, F.; de Nadal, E. The p38 pathway: From biology to cancer therapy. Int. J. Mol. Sci. 2020, 21, 1913. [Google Scholar] [CrossRef] [Green Version]
- Yin, S.-Y.; Wei, W.-C.; Jian, F.-Y.; Yang, N.-S. Therapeutic applications of herbal medicines for cancer patients. Evid. Based Complement. Altern. Med. 2013, 2013, 302426. [Google Scholar] [CrossRef]
- Huang, M.; Lu, J.-J.; Huang, M.-Q.; Bao, J.-L.; Chen, X.-P.; Wang, Y.-T. Terpenoids: Natural products for cancer therapy. Expert Opin. Investig. Drugs 2012, 21, 1801–1818. [Google Scholar] [CrossRef]
- Feng, L.; Liu, T.T.; Huo, X.K.; Tian, X.G.; Wang, C.; Lv, X.; Ning, J.; Zhao, W.Y.; Zhang, B.J.; Sun, C.P. Alisma genus: Phytochemical constituents, biosynthesis, and biological activities. Phytother. Res. 2021, 35, 1872–1886. [Google Scholar] [CrossRef]
- Park, Y.-J.; Kim, M.-S.; Kim, H.-R.; Kim, J.-M.; Hwang, J.-K.; Yang, S.-H.; Kim, H.-J.; Lee, D.-S.; Oh, H.; Kim, Y.-C. Ethanol extract of Alismatis rhizome inhibits adipocyte differentiation of OP9 cells. Evid. Based Complement. Altern. Med. 2014, 2014, 415097. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Song, T.; Shi, R.; He, M.; Wang, R.; Lv, J.; Jiang, M. Triterpenoids from Alisma species: Phytochemistry, structure modification, and bioactivities. Front. Chem. 2020, 8, 363. [Google Scholar] [CrossRef] [PubMed]
- Petronelli, A.; Pannitteri, G.; Testa, U. Triterpenoids as new promising anticancer drugs. Anti Cancer Drugs 2009, 20, 880–892. [Google Scholar] [CrossRef]
- Xu, W.; Li, T.; Huang, M.; Chen, X.P.; Lu, J.J. Anti-cancer effects of triterpenoids isolated form Alismatis rhizoma on HepG2 cells. Acta Pharmacol. Sin. 2013, 34, 16–17. [Google Scholar]
- Ma, Q.; Han, L.; Bi, X.; Wang, X.; Mu, Y.; Guan, P.; Li, L.; Huang, X. Structures and biological activities of the triterpenoids and sesquiterpenoids from Alisma orientale. Phytochemistry 2016, 131, 150–157. [Google Scholar] [CrossRef]
- Jin, S. Autophagy, mitochondrial quality control, and oncogenesis. Autophagy 2006, 2, 80–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, M.J.; Kim, J.N.; Park, J.; Kim, Y.T.; Lee, M.J.; Kim, B.J. Alisma canaliculatum Extract Affects AGS Gastric Cancer Cells by Inducing Apoptosis. Int. J. Med. Sci. 2021, 18, 2155–2161. [Google Scholar] [CrossRef]
- Choi, J.; Ahn, S.S.; Lim, Y.; Lee, Y.H.; Shin, S.Y. Inhibitory effect of Alisma canaliculatum ethanolic extract on NF-κB-dependent CXCR3 and CXCL10 expression in TNFα-exposed MDA-MB-231 breast cancer cells. Int. J. Mol. Sci. 2018, 19, 2607. [Google Scholar] [CrossRef] [Green Version]
- Lou, C.; Xu, X.; Chen, Y.; Zhao, H. Alisol A suppresses proliferation, migration, and invasion in human breast cancer MDA-MB-231 cells. Molecules 2019, 24, 3651. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Wang, M.; Wang, P.; Zhang, T.; Yu, J.; Shi, L.; Li, M.; Wang, H.; Zhang, Q.; Zhao, H. Alisol A is potentially therapeutic in human breast cancer cells. Oncol. Rep. 2020, 44, 1266–1274. [Google Scholar] [CrossRef]
- Xu, F.; Lu, C.; Wu, Q.; Chen, J.; Gu, W.; Du, W.; You, M. Study on antitumor molecular mechanism of Alisols based on p53DNA. Int. J. Biol. Macromol. 2018, 116, 1163–1174. [Google Scholar] [CrossRef]
- Law, B.Y.; Wang, M.; Ma, D.-L.; Al-Mousa, F.; Michelangeli, F.; Cheng, S.-H.; Ng, M.H.; To, K.-F.; Mok, A.Y.; Ko, R.Y. Alisol B, a novel inhibitor of the sarcoplasmic/endoplasmic reticulum Ca2+ ATPase pump, induces autophagy, endoplasmic reticulum stress, and apoptosis. Mol. Cancer Ther. 2010, 9, 718–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, A.; Sheng, Y.; Zou, M. Antiproliferative activity of Alisol B in MDA-MB-231 cells is mediated by apoptosis, dysregulation of mitochondrial functions, cell cycle arrest and generation of reactive oxygen species. Biomed. Pharmacother. 2017, 87, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.; Pan, S.; Teng, C.; Guh, J. Pharmacological evaluation of several major ingredients of Chinese herbal medicines in human hepatoma Hep3B cells. Eur. J. Pharm. Sci. 2003, 19, 403–412. [Google Scholar] [CrossRef]
- Xu, Y.-H.; Zhao, L.-J.; Li, Y. Alisol B acetate induces apoptosis of SGC7901 cells via mitochondrial and phosphatidylinositol 3-kinases/Akt signaling pathways. World J. Gastroenterol. WJG 2009, 15, 2870–2877. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-T.; Huang, D.-M.; Chueh, S.-C.; Teng, C.-M.; Guh, J.-H. Alisol B acetate, a triterpene from Alismatis rhizoma, induces Bax nuclear translocation and apoptosis in human hormone-resistant prostate cancer PC-3 cells. Cancer Lett. 2006, 231, 270–278. [Google Scholar] [CrossRef]
- Kwon, M.J.; Kim, J.N.; Lee, M.J.; Kim, W.K.; Nam, J.H.; Kim, B.J. Apoptotic effects of alisol B 23-acetate on gastric cancer cells. Mol. Med. Rep. 2021, 23, 248. [Google Scholar] [CrossRef]
- Wang, J.; Li, H.; Wang, X.; Shen, T.; Wang, S.; Ren, D. Alisol B-23-acetate, a tetracyclic triterpenoid isolated from Alisma orientale, induces apoptosis in human lung cancer cells via the mitochondrial pathway. Biochem. Biophys. Res. Commun. 2018, 505, 1015–1021. [Google Scholar] [CrossRef]
- Liu, Y.; Xia, X.C.; Meng, L.Y.; Wang, Y.; Li, Y.M. Alisol B 23-acetate inhibits the viability and induces apoptosis of non-small cell lung cancer cells via PI3K/AKT/mTOR signal pathway. Mol. Med. Rep. 2019, 20, 1187–1195. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Luo, Q.; Huang, S.; Jiang, F.; Wang, L.; Wang, G.; Xie, J.; Liu, J.; Xu, Y. Alisol B 23-acetate-induced HepG2 hepatoma cell death through mTOR signaling-initiated G1 cell cycle arrest and apoptosis: A quantitative proteomic study. Chin. J. Cancer Res. 2019, 31, 375–388. [Google Scholar] [CrossRef]
- Li, L.; Cheng, J.; Zhu, D.; Shi, X.; Wei, Y.; Chen, S.; Wang, Z.; Yuan, D. The effects of Alisol B 23-acetate in hepatocellular carcinoma via inducing cell apoptosis and inhibiting cell migration and invasion. Gen. Physiol. Biophys. 2020, 39, 219–228. [Google Scholar] [CrossRef]
- Xu, W.; Li, T.; Qiu, J.F.; Wu, S.S.; Huang, M.Q.; Lin, L.G.; Zhang, Q.W.; Chen, X.P.; Lu, J.J. Anti-proliferative activities of terpenoids isolated from Alisma orientalis and their structure-activity relationships. Anti Cancer Agents Med. Chem. 2015, 15, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, K.L.; Goolsby, G.; Kiener, P.A.; Wahl, A.F. Activation of p34cdc2 coincident with taxol-induced apoptosis. Cell Growth Differ. 1994, 5, 1041–1050. [Google Scholar]
- Zhang, L.-L.; Xu, Y.-L.; Tang, Z.-H.; Xu, X.-H.; Chen, X.; Li, T.; Ding, C.-Y.; Huang, M.-Q.; Chen, X.-P.; Wang, Y.-T. Effects of alisol B 23-acetate on ovarian cancer cells: G1 phase cell cycle arrest, apoptosis, migration and invasion inhibition. Phytomedicine 2016, 23, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Li, X.-Y.; Wang, W.-H.; Cai, F.-F.; Chen, X.-L.; Yang, M.-D.; Pan, Q.-S.; Chen, Q.-L.; Zhou, R.-Y.; Su, S.-B. Network pharmacology-based study on the mechanism of Bushen-Jianpi decoction in liver cancer treatment. Evid. Based Complement. Altern. Med. 2019, 2019, 3242989. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Kho, Y.; Min, B.; Kim, J.; Na, M.; Kang, S.; Maeng, H.; Bae, K. Cytotoxic triterpenoides from Alismatis rhizoma. Arch. Pharmacal Res. 2001, 24, 524–526. [Google Scholar] [CrossRef] [PubMed]
- Shao, B.; Wang, S.; Zhou, J.; Ke, L.; Rao, P. A novel lectin from fresh rhizome of Alisma orientale (Sam.) Juzep. Process Biochem. 2011, 46, 1554–1559. [Google Scholar] [CrossRef]
- Chen, H.-W.; Hsu, M.-J.; Chien, C.-T.; Huang, H.-C. Effect of alisol B acetate, a plant triterpene, on apoptosis in vascular smooth muscle cells and lymphocytes. Eur. J. Pharmacol. 2001, 419, 127–138. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, E.T.; Wang, M. Alisol B 23-acetate induces autophagic-dependent apoptosis in human colon cancer cells via ROS generation and JNK activation. Oncotarget 2017, 8, 70239–70249. [Google Scholar] [CrossRef] [Green Version]
- Valastyan, S.; Weinberg, R.A. Tumor metastasis: Molecular insights and evolving paradigms. Cell 2011, 147, 275–292. [Google Scholar] [CrossRef] [Green Version]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular principles of metastasis: A hallmark of cancer revisited. Signal Transduct. Target. Ther. 2020, 5, 1–17. [Google Scholar] [CrossRef]
- Ma, B.; Xiang, Y.; Li, T. Inhibitory effect of Alisma orientalis on spontaneous metastasis of Lewis lung carcinoma and its mechanism. Chin. Tradit. Drug 2003, 34, 743–746. [Google Scholar]
- Pires, B.R.; Mencalha, A.L.; Ferreira, G.M.; de Souza, W.F.; Morgado-Díaz, J.A.; Maia, A.M.; Corrêa, S.; Abdelhay, E.S. NF-kappaB is involved in the regulation of EMT genes in breast cancer cells. PLoS ONE 2017, 12, e0169622. [Google Scholar] [CrossRef] [Green Version]
- D’Alterio, C.; Scala, S.; Sozzi, G.; Roz, L.; Bertolini, G. Paradoxical effects of chemotherapy on tumor relapse and metastasis promotion. Semin. Cancer Biol. 2019, 60, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Thomas, H.; Coley, H.M. Overcoming multidrug resistance in cancer: An update on the clinical strategy of inhibiting p-glycoprotein. Cancer Control 2003, 10, 159–165. [Google Scholar] [CrossRef]
- Katayama, K.; Noguchi, K.; Sugimoto, Y. Regulations of P-glycoprotein/ABCB1/MDR1 in human cancer cells. New J. Sci. 2014, 2014, 476974. [Google Scholar]
- Uramoto, H.; Tanaka, F. Recurrence after surgery in patients with NSCLC. Transl. Lung Cancer Res. 2014, 3, 242–249. [Google Scholar]
- Castells, M.; Thibault, B.; Delord, J.-P.; Couderc, B. Implication of tumor microenvironment in chemoresistance: Tumor-associated stromal cells protect tumor cells from cell death. Int. J. Mol. Sci. 2012, 13, 9545–9571. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, D.; Sibson, K.; Caswell, M.; Connor, P.; Cummins, M.; Mitchell, C.; Motwani, J.; Taj, M.; Vora, A.; Wynn, R. Early UK experience in the use of clofarabine in the treatment of relapsed and refractory paediatric acute lymphoblastic leukaemia. Br. J. Haematol. 2011, 154, 482–485. [Google Scholar] [CrossRef]
- Fong, W.-F.; Wang, C.; Zhu, G.-Y.; Leung, C.-H.; Yang, M.-S.; Cheung, H.-Y. Reversal of multidrug resistance in cancer cells by Rhizoma Alismatis extract. Phytomedicine 2007, 14, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, J.-X.; Shen, X.-L.; Wan, C.-K.; Tse, A.K.-W.; Fong, W.-F. Reversal of P-glycoprotein-mediated multidrug resistance by Alisol B 23-acetate. Biochem. Pharmacol. 2004, 68, 843–855. [Google Scholar] [CrossRef] [PubMed]
- Pan, G.; Li, T.; Zeng, Q.; Wang, X.; Zhu, Y. Alisol F 24 acetate enhances chemosensitivity and apoptosis of MCF-7/DOX Cells by inhibiting P-glycoprotein-mediated drug efflux. Molecules 2016, 21, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuenda, A.; Rousseau, S. p38 MAP-kinases pathway regulation, function and role in human diseases. Biochim. Biophys. Acta Mol. Cell Res. 2007, 1773, 1358–1375. [Google Scholar] [CrossRef] [Green Version]
- Pan, S.-T.; Qin, Y.; Zhou, Z.-W.; He, Z.-X.; Zhang, X.; Yang, T.; Yang, Y.-X.; Wang, D.; Qiu, J.-X.; Zhou, S.-F. Plumbagin induces G2/M arrest, apoptosis, and autophagy via p38 MAPK-and PI3K/Akt/mTOR-mediated pathways in human tongue squamous cell carcinoma cells. Drug Des. Dev. Ther. 2015, 9, 1601–1626. [Google Scholar]
- Xu, Y.; Li, N.; Xiang, R.; Sun, P. Emerging roles of the p38 MAPK and PI3K/AKT/mTOR pathways in oncogene-induced senescence. Trends Biochem. Sci. 2014, 39, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Bode, A.M.; Dong, Z.; Lee, M.-H. AKT as a therapeutic target for cancer. Cancer Res. 2019, 79, 1019–1031. [Google Scholar] [CrossRef] [Green Version]
- DiDonato, J.A.; Mercurio, F.; Karin, M. NF-κB and the link between inflammation and cancer. Immunol. Rev. 2012, 246, 379–400. [Google Scholar] [CrossRef]
- Fulda, S.; Debatin, K.M. Extrinsic versus intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene 2006, 25, 4798–4811. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.; Wu, J.; Chen, Y.; Nie, J.; Chen, C. Activation of PI3K/AKT/mTOR Pathway Causes Drug Resistance in Breast Cancer. Front. Pharmacol. 2021, 12, 143. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Viatour, P. Hepatocellular carcinoma: Old friends and new tricks. Exp. Mol. Med. 2020, 52, 1898–1907. [Google Scholar] [CrossRef] [PubMed]
- Suresh, D.; Srinivas, A.N.; Kumar, D.P. Etiology of Hepatocellular Carcinoma: Special Focus on Fatty Liver Disease. Front. Oncol. 2020, 10, 601710. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.; Jang, E.; Lee, J.-H. Pharmacological activities of Alisma orientale against nonalcoholic fatty liver disease and metabolic syndrome: Literature review. Evid. Based Complement. Altern. Med. 2019, 2019, 2943162. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
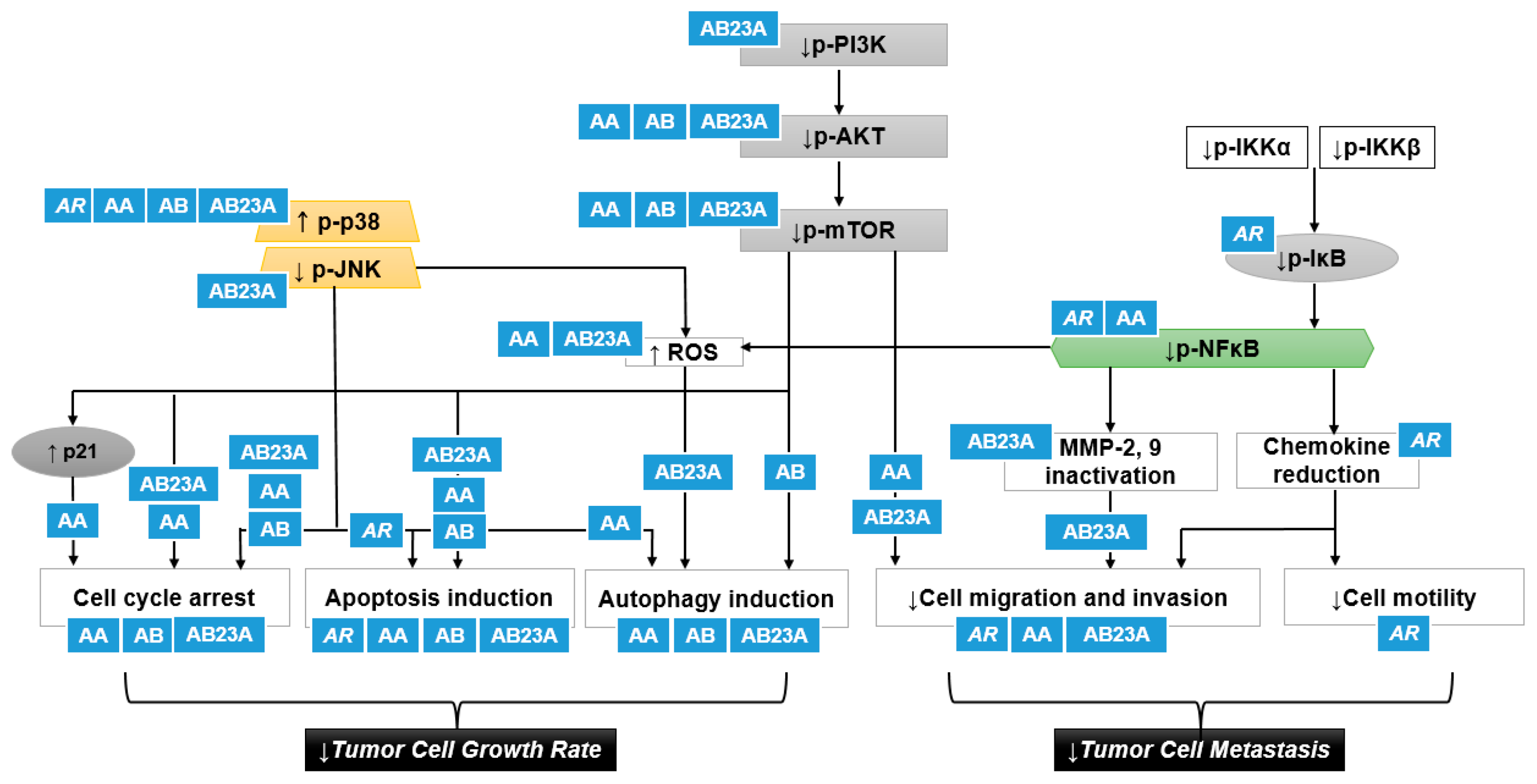{kind=link}
| Sources | Country | Models | Doses | Results and Mechanisms | Ref. |
|---|---|---|---|---|---|
| A. canaliculatum extract | Korea | In vitro, human gastric cancer cells AGS | 300–500 μg/mL | ↓Cell Growth Rate | [16] |
| Apoptosis induction ↑Sub-G1 ↑Mitochondrial membrane depolarization ↑Bax protein ↓Bcl-2 protein ↓Survivin protein ↑PARP cleavage | |||||
| ↑p-p38 protein | |||||
| AB23A | Korea | In vitro, human gastric cancer cells AGS | 50 μm | ↓Cell growth rate | [26] |
| Apoptosis induction ↑Sub-G1 ↑Bax protein ↓Bcl-2 protein ↓Survivin protein ↑PARP cleavage ↑Caspase-3,-9 protein cleavage | |||||
| ↑p-p38, p-ERK, p-JNK protein | |||||
| ROS generation ↑Intracellular ROS levels | |||||
| AB23A obtained from National Institutes for Food and Drug Control (Beijing, China) | China | In vitro, human ovarian cancer cells HEY | 2.5–20 μm | ↓Cell growth rate (IC50: 10.73 μm) | [33] |
| Cell cycle arrest ↑G1 arrest ↓Cyclin D1, CDK4, CDK6 protein | |||||
| Apoptosis induction ↑Sub-G1 ↑PARP cleavage ↑Bax/Bcl-2 | |||||
| AB23A obtained from National Institutes for Food and Drug Control (Beijing, China) | China | In vitro, human ovarian cancer cells A2780 | 9–18 μm | ↓Cell growth rate (IC50: 11.21 μm) | [33] |
| Cell cycle arrest ↑G1 arrest ↓Cyclin D1, CDK4, CDK6 protein | |||||
| Apoptosis induction ↑Sub-G1 ↑PARP cleavage ↑Bax/Bcl-2 | |||||
| AB23A | China | In vitro, human hepatoma HepG2 | 10, 15, 20 μm | ↓Cell growth rate (IC50: 17.82 F) | [29] |
| Cell cycle arrest ↑G1 arrest ↓Cyclin D1, CDK4, Rb, p-Rb protein | |||||
| Apoptosis induction ↑Cells displaying nuclear condensation and fragmentation ↑The percentage of apoptotic cells ↑PARP cleavage ↑Caspase-3 cleavage Through DR3 and DR4/5 death receptors | |||||
| ↓p-mTOR | |||||
| AB purchased from the Standardization Research Center of TCM (Shanghai, China) | Shanghai, China | In vitro, human hepatoma HepG2 | 6.25–200 μmol/L | ↓Cell growth rate (IC50: 32.57 μm) | [34] |
| AB23A from A. plantago-aquatica | Kyoto, Japan | In vitro, human hepatoma containing hepatitis B virus Hep3B | 50 μm | ↓Cell growth rate (IC50: 42.4 μm) | [23] |
| Cell cycle arrest ↑G2/M arrest | |||||
| Apoptosis induction ↑The percentage of apoptotic cells↑DNA fragmentation ↑Sub-G1 ↑Caspase-3 protein | |||||
| AB23A | Shanghai, China | In vitro, human non-small cell lung cancer cells A549 | 6, 9 mM | ↓Cell growth rate | [28] |
| Cell cycle arrest ↑G0/G1 arrest | |||||
| Apoptosis induction ↑The percentage of apoptotic cells↑Bax protein ↓Bcl-2 protein | |||||
| ↓p-PI3K/AKT/mTOR protein | |||||
| AB23A isolated from the 90% ethanol extract of A. orientale | Shandong, China | In vitro, human lung cancer cells A549 (adenocarcinoma) and NCI-H292 (mucoepidermoid cancer) | 10, 20 μm | ↓Cell growth rate | [27] |
| Apoptosis induction ↑The percentage of apoptotic cells ↑Bax, Caspase-3,-9 protein ↓Bcl-2, Bcl-xL protein ↑PARP cleavage ↑Mitochondrial membrane depolarization ↑Cytochrome c release | |||||
| ROS generation ↑Intracellular ROS levels | |||||
| AB23A | Beijing, China | In vitro, human Hepatoma SK-HEP-1 | 30 μmol/L | ↓Cell growth rate | [30] |
| Cell cycle arrest ↑G0/G1 arrest | |||||
| Apoptosis induction ↑The percentage of apoptotic cells ↑Bax, Caspase-3,-9 protein and mRNA ↓Bcl-2 protein and mRNA | |||||
| ↓p-PI3K/AKT protein and mRNA | |||||
| AB23A purchased from Wako Pure Chemical Industries (Osaka, Japan) | Osaka, Japan | In vitro, human gastric cancer cells SGC7901 | 30 μmol/L | ↓Cell growth rate | [24] |
| Cell cycle arrest ↑G0/G1 arrest | |||||
| Apoptosis induction ↑The percentage of apoptotic cells ↑Bax, Caspase-3,-9 protein ↑Apaf-1 protein ↑Mitochondrial membrane depolarization | |||||
| ↓p-PI3K/AKT protein | |||||
| AB23A | Beijing, China | In vivo, BALB/c mice inoculated with SK-HEP-1 | 30 μmol/L | ↓Tumor volume and weight | [30] |
| Apoptosis induction ↑Bax, Caspase-3,-9 protein and mRNA in tumor xenograft ↓Bcl-2 protein and mRNA in tumor xenograft | |||||
| AB | Daejeon, Korea | In vitro, human ovarian cancer cells SK-OV3 | ↓Cell growth rate (IC50: 7.5 μg/mL) | [35] | |
| Lectin from A. orientale | Jianou, China | In vitro, human uterine cervical cancer cells HeLa | 5, 10, 20 μm | ↓Cell growth rate (IC50: 7.3 μm) | [36] |
| AA purchased from MedChemExpress (Monmouth Junction, NJ, USA) | Hangzhou, China | In vitro, human breast cancer cells MDA-MB-231 | 20, 40 μm | ↓Cell growth rate | [18] |
| Cell cycle arrest ↑G0/G1 arrest ↓Cyclin D1 ↑p21 | |||||
| Apoptosis induction ↑The percentage of apoptotic cells | |||||
| Autophagy induction ↑Autophagic vacuoles ↑LC3-II protein | |||||
| ↓p-AKT protein ↓p-mTOR protein ↓p-70S6K protein ↓p-NFκB protein | |||||
| AA | Chengdu, China | In vitro, human breast cancer cells MCF-7 | 30, 100 μm | ↓Cell growth rate | [20] |
| AA | Beijing, China | In vitro, human breast cancer cells MDA-MB-231 | 5, 10, 20 μm | ↓Cell growth rate (IC50: 8.112 μm) | [19] |
| Cell cycle arrest ↑G1 arrest ↓Cyclin A/D1 | |||||
| Apoptosis induction ↑The percentage of apoptotic cells ↑Caspase-3,-9 protein ↓Bcl-2 protein | |||||
| Autophagy induction ↑LC3-II protein | |||||
| ↑p-p38 protein ↑ROS positive cells ↑DNA damage markers | |||||
| AB | Chengdu, China | In vitro, human breast cancer cells MCF-7 | 10, 30, 100 μm | ↓Cell growth rate | [20] |
| AA-AB (1:1) | Chengdu, China | In vitro, human breast cancer cells MCF-7 | 100 μm | ↓Cell growth rate | [20] |
| AB purchased from Wako Pure Chemical Industries (Osaka, Japan) | Osaka, Japan | In vitro, human breast cancer cells MCF-7 | 30 μmol/L | ↓Cell growth rate (IC50: 29.9 μmol/L) | [21] |
| Cell cycle arrest ↑G1 arrest ↑p27 | |||||
| Autophagy induction ↑Autophagic vacuoles ↑LC3-II protein | |||||
| ↑p-AMPK ↓p-70S6 | |||||
| AB purchased from Sigma Chemical Co. (St. Louis, MO, USA) | Zhenjiang, China | In vitro, human breast cancer cells MDA-MB-231 | 10, 20 μm | ↓Cell growth rate (IC50: 13.96 μm) | [22] |
| Cell cycle arrest ↑G0/G1 arrest | |||||
| Apoptosis induction ↑The percentage of apoptotic cells ↑Caspase-3,-9 protein | |||||
| ↓p-AKT/mTOR protein ↓p-p65 protein ↑p-p38 protein | |||||
| AB23A purchased from Wako Pure Chemical Industries (Osaka, Japan) | Osaka, Japan | In vitro, human prostate cancer PC-3 cells | 30 μm | ↓Cell growth rate (IC50: 13.5 μm) | [25] |
| Apoptosis induction ↑The percentage of apoptotic cells ↑Mitochondrial membrane depolarization ↑Bax protein ↑Cleaved caspase-3,-9 protein ↑Cleaved caspase-8 protein | |||||
| AB23 A from A. plantago-aquatica purchased from Nacalai Tesque (Kyoto, Japan) | Taipei, Taiwan | In vitro, human acute lymphoblastic leukemia CEM cells | 10−6–10−4 M | ↓Cell growth rate (IC50: 10−4 M) | [37] |
| Apoptosis induction ↑Mitochondrial membrane depolarization ↑c-myc and Bax mRNA and proteins | |||||
| AB23A | China | In vitro, human colon cancer cells SW620 and HCT116 | 5, 10, 20 μm | ↓Cell growth rate (IC50: 20 μm) | [38] |
| Cell cycle arrest ↑G1 arrest | |||||
| Apoptosis induction ↑The percentage of apoptotic cells ↑Sub-G1 ↑PARP cleavage | |||||
| Autophagy induction in HCT116 ↑LC3-II protein ↓p62 protein (substrate) | |||||
| ROS generation ↑Intracellular ROS levels ↓p-JNK protein | |||||
| AB23A | China | In vitro, human kidney, gastric, and liver cancer cells HEK293T, AGS, PLC8024 | 20 μm | ↓Cell growth rate | [38] |
| Sources | Country | Models | Doses | Results and Mechanisms | Ref. |
|---|---|---|---|---|---|
| AB23A | Shanghai, China | In vitro, human non-small cell lung cancer cells A549 | 6, 9 mM | ↓Cell migration and invasion | [28] |
| AB23A | China | In vitro, human hepatoma cells SK-HEP-1 | 30 μmol/L | ↓Cell migration and invasion ↓Migration and invasion rate using wound healing assay and transwell assay | [30] |
| ↓MMP-2, 9 protein and mRNA ↓p-PI3K/AKT protein and mRNA | |||||
| AB23A | Beijing, China | In vitro, human ovarian cancer cells HEY | 5, 10 μm | ↓Cell migration and invasion ↓Migration and invasion rate using wound healing assay and transwell assay | [33] |
| ↓MMP-2, 9 protein | |||||
| AA Purchased from MedChem Express (Monmouth Junction, NJ, USA) | Hanzhou, China | In vitro, human breast cancer cells MDA-MB-231 | 5 μm | ↓Cell migration and invasion ↓Migration and invasion rate using wound healing assay and transwell assay | [18] |
| ↓MMP-2, 9 protein ↓p-AKT protein ↓p-mTOR protein ↓p-70S6K protein ↓p-NFκB protein | |||||
| A. canaliculatum Purchased from the Kyungdong traditional medicine market (Seoul, Korea) | Seoul, Korea | In vitro, human breast cancer cells MDA-MB-231 | 20 μg/mL | ↓Cell migration and invasion ↓TNFα-induced migration rate using wound healing assay ↓TNFα-induced motility causing morphological changes to spindle-like cells | [17] |
| ↓TNFα-induced CXCR3 and CXCL10 mRNA ↓p-IKKα/β protein ↓p-IκB, p-p65/RelA protein | |||||
| A. orientalis | China | In vivo, C57BL/6 transplanted with Lewis lung carcinoma | 10, 20 g/kg/d | ↓Spontaneous metastasis | [41] |
| Sources | Country | Models | Doses | Positive control | Results and Mechanisms | Ref. |
|---|---|---|---|---|---|---|
| AB23A | Beijing, China | In vitro, human ovarian cancer A2780 cells resistant to paclitaxel drug | 12–18 μm | paclitaxel 10 μm | ↓Cell growth rate (IC50: 15.18 μm) | [33] |
| Cell cycle arrest ↑G1 arrest ↓Cyclin D1, CDK4, CDK6 protein | ||||||
| Apoptosis induction ↑Sub-G1 ↑PARP cleavage ↑Bax/Bcl-2 | ||||||
| AB23A from 95% ethanol extract of AR | Hong Kong | In vitro, human hepatoma HepG cells resistant to vinblastine, actinomycin D, puromycin, paclitaxel, doxorubicin drugs | 10 μm | vinblastine 300 nM | ↓Cell growth rate | [50] |
| Cell cycle arrest ↑G2/M arrest | ||||||
| Chemosensitizer Combination index < 0.8 ↑Cellular accumulation of doxorubicin ↓Pgp activity ↓Rh-123 transport | ||||||
| AB23A from 95% ethanol extract of AR | Hong Kong | In vitro, human leukemia K562 cells resistant to vinblastine drug | 10 μm | vinblastine 300 nM | ↓Cell growth rate | [50] |
| Chemosensitizer ↑Cellular accumulation of vinblastine →Pgp expression ↓Rh-123 transport | ||||||
| AF24A purchased from Science and Technology Co., Ltd. (Tianjin, China) | Tianjin, China | In vitro, human colorectal adenocarcinoma Caco-2 monolayers overexpressing Pgp | 10 μm | verapamil 10 μm | ↓Efflux ratio of digoxin | [51] |
| AF24A purchased from Science and Technology Co., Ltd. (Tianjin, China) | Tianjin, China | In vitro, human breast cancer cells MCF-7-digoxin resistant cells | 5, 10, 20 μm | verapamil 10 μm | ↓IC50 of doxorubicin | [51] |
| Chemosensitizer Log of combination index < 0 ↑Cellular accumulation of doxorubicin ↑Nuclear migration of doxorubicin | ||||||
| Apoptosis induction ↑Cytoplasmic TMRE fluorescence intensity ↑Mitochondrial membrane depolarization | ||||||
| 95% ethanol extract of AR | Hong Kong | In vitro, human hepatoma HepG2 cells resistant to vinblastine, actinomycin D, puromycin, paclitaxel, doxorubicin drugs | 25 μg/ml | verapamil 10 μm | ↓Cell growth rate | [49] |
| Cell cycle arrest ↑G2/M arrest | ||||||
| Human leukemia K562 cells resistant to vinblastine, actinomycin D, puromycin, paclitaxel, doxorubicin drugs | ↑Cellular accumulation of doxorubicin ↓Rh-123 transport →Pgp expression |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, E.; Lee, J.-H. Promising Anticancer Activities of Alismatis rhizome and Its Triterpenes via p38 and PI3K/Akt/mTOR Signaling Pathways. Nutrients 2021, 13, 2455. https://doi.org/10.3390/nu13072455
Jang E, Lee J-H. Promising Anticancer Activities of Alismatis rhizome and Its Triterpenes via p38 and PI3K/Akt/mTOR Signaling Pathways. Nutrients. 2021; 13(7):2455. https://doi.org/10.3390/nu13072455
Chicago/Turabian StyleJang, Eungyeong, and Jang-Hoon Lee. 2021. "Promising Anticancer Activities of Alismatis rhizome and Its Triterpenes via p38 and PI3K/Akt/mTOR Signaling Pathways" Nutrients 13, no. 7: 2455. https://doi.org/10.3390/nu13072455
APA StyleJang, E., & Lee, J. -H. (2021). Promising Anticancer Activities of Alismatis rhizome and Its Triterpenes via p38 and PI3K/Akt/mTOR Signaling Pathways. Nutrients, 13(7), 2455. https://doi.org/10.3390/nu13072455