
Parenteral Nutrition and Oxidant Load in Neonates


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
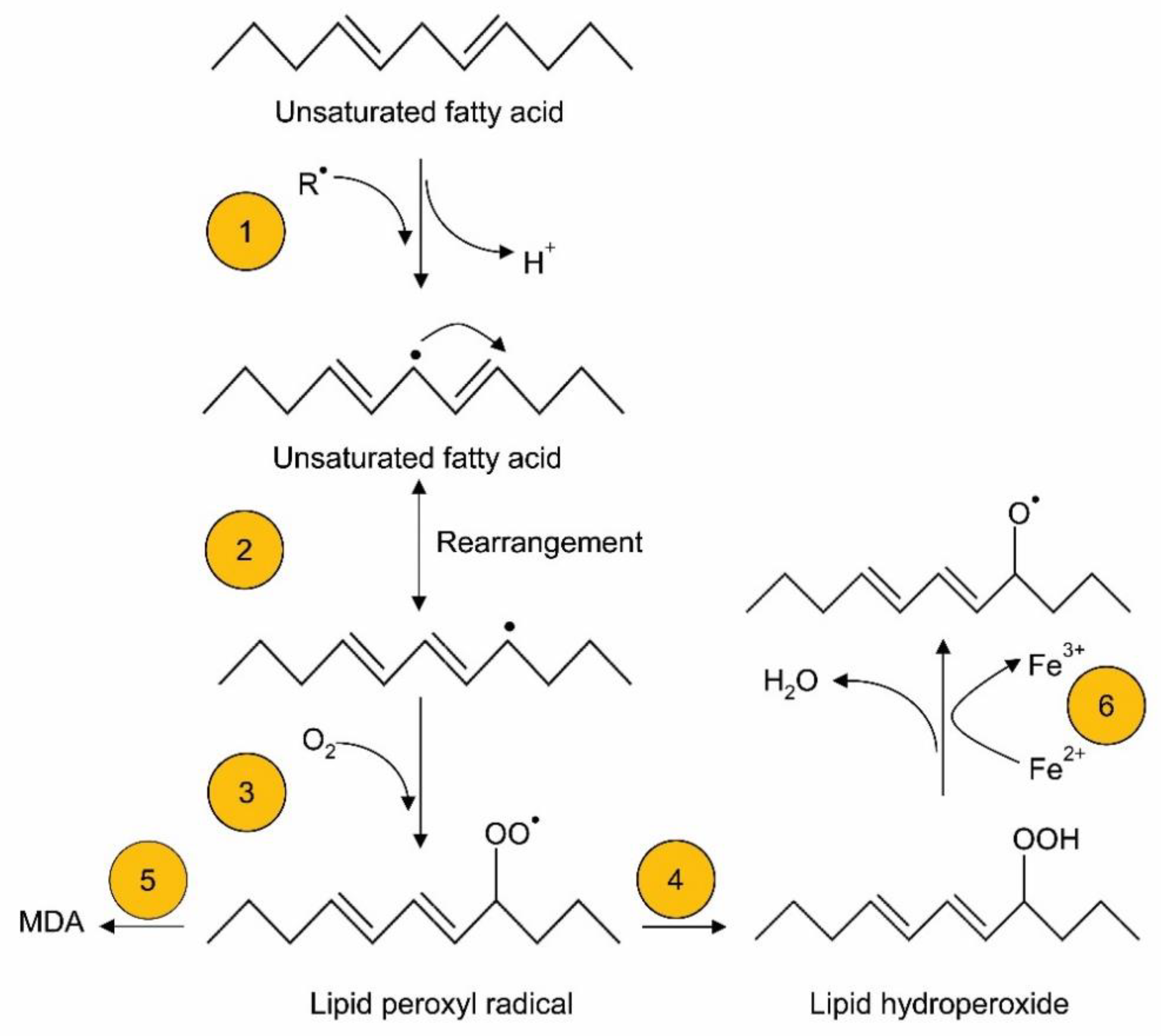
2. In Vitro Oxidation of TPN
2.1. Oxidation of Lipid Emulsions for TPN
2.2. Oxidation of Amino Acids within TPN
2.3. Role of Vitamins in the Oxidation of TPN Solutions
2.3.1. Polysorbate
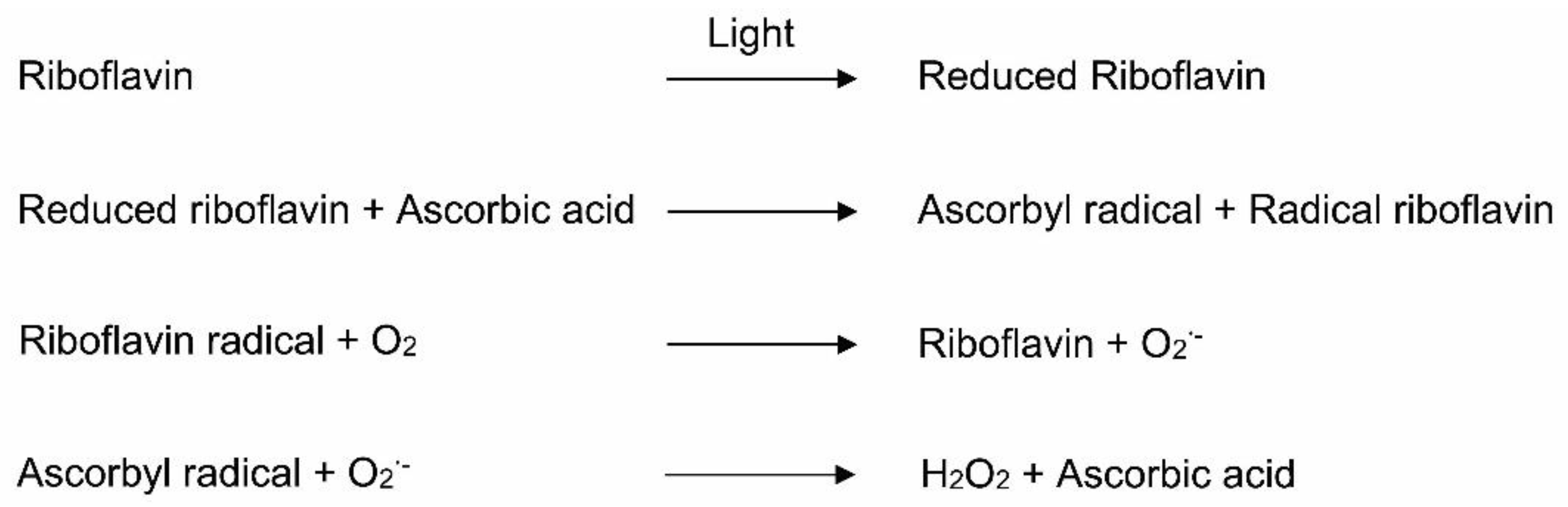
2.3.2. Riboflavin
2.3.3. Vitamin E
2.3.4. Vitamin C
2.4. Role of Trace Elements in Oxidation of TPN Solutions
3. Parenteral Nutrition and Environmental Conditions
3.1. TPN Exposure to Light
3.2. TPN Exposure to Oxygen
3.3. Effect of Storage on Oxidants in TPN Solutions
4. In Vivo Impact of Oxidized TPN after Infusion
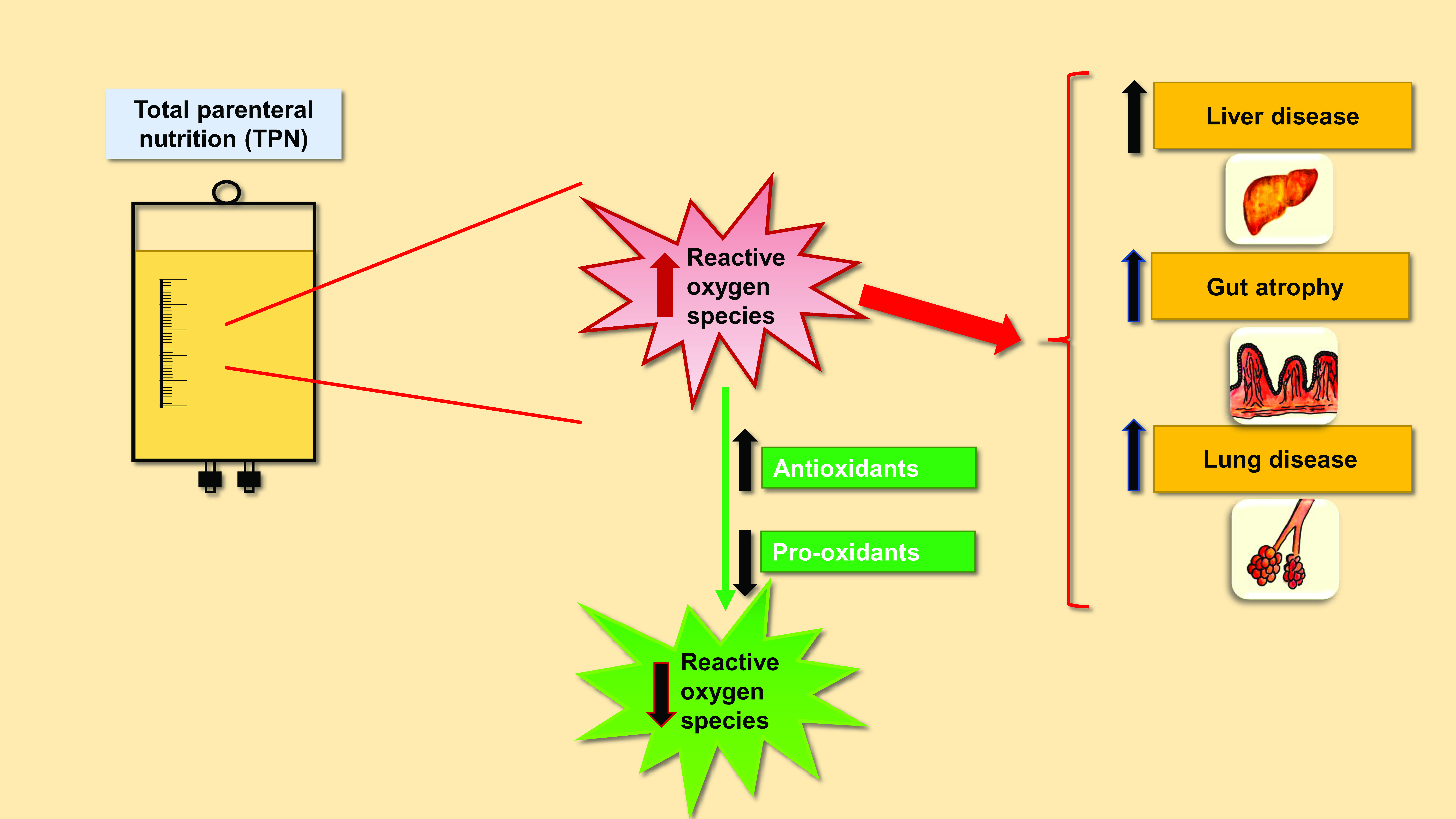
4.1. Parenteral Nutrition, Oxidant Load and Liver Diseases
4.2. Parenteral Nutrition and Gut Atrophy
4.3. Parenteral Nutrition and BPD
4.4. Biomolecules in the Body Affected by Oxidized TPN
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Johnston, K.M.; Gooch, K.; Korol, E.; Vo, P.; Eyawo, O.; Bradt, P.; Levy, A. The economic burden of prematurity in Canada. BMC Pediatr. 2014, 14, 93. [Google Scholar] [CrossRef] [Green Version]
- Shah, P.S.; McDonald, S.D.; Barrett, J.; Synnes, A.; Robson, K.; Foster, J.; Pasquier, J.; Joseph, K.S.; Piedboeuf, B.; Lacaze-Masmonteil, T.; et al. The Canadian Preterm Birth Network: A study protocol for improving outcomes for preterm infants and their families. CMAJ Open 2018, 18, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Statistics Canada. Table 13-10-0404-01 Low Birth Weight Babies, by Province and Territory. Available online: https://www150.statcan.gc.ca/t1/tbl1/en/tv.action?pid=1310040401 (accessed on 5 June 2021).
- Luu, T.M.; Katz, S.L.; Leeson, P.; Thébaud, B.; Nuyt, A.M. Preterm birth: Risk factor for early-onset chronic diseases. Can. Med. Assoc. J. 2015, 188, 736–746. [Google Scholar] [CrossRef] [Green Version]
- Lavoie, J.C.; Chessex, P. Parenteral nutrition and oxidant stress in the newborn: A narrative review. Free Radic. Biol. Med. 2019, 140, 155–167. [Google Scholar] [CrossRef]
- Saugstad, O.D. Bronchopulmonary dysplasia-oxidative stress and antioxidants. Semin. Neonatol. 2003, 8, 39–49. [Google Scholar] [CrossRef]
- Jankov, R.P.; Negus, A.; Tanswell, K. Antioxidants as therapy in the newborn: Some words of caution. Pediatr. Res. 2001, 50, 681–687. [Google Scholar] [CrossRef] [Green Version]
- Vento, M.; Moro, M.; Escrig, R.; Arruza, L.; Villar, G.; Izquierdo, I.; Roberts, L.J.; Arduini, A.; Escobar, J.J.; Sastre, J.; et al. Preterm resuscitation with low oxygen causes less oxidative stress, inflammation, and chronic lung disease. Pediatrics 2009, 124, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Saugstad, O.D. Room air resuscitation-two decades of neonatal research. Early Hum. Dev. 2005, 81, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Lucas, A.; Morley, R.; Cole, T.J.; Lister, G.; Leeson-Payne, C. Breast milk and subsequent intelligence quotient in children born preterm. Lancet 1992, 329, 261–264. [Google Scholar] [CrossRef]
- Kennedy, K.A.; Tyson, J.E.; Chamnanvanikij, S. Early versus delayed initiation of progressive enteral feedings for parenterally fed low birth weight or preterm infants. Cochrane Database Syst. Rev. 2000, 2, CD001970. [Google Scholar] [CrossRef]
- Calkins, K.L.; Venick, R.S.; Devaskar, S.U. Complications associated with parenteral nutrition in the neonate. Clin. Perinatol. 2014, 41, 331–345. [Google Scholar] [CrossRef] [Green Version]
- Ozsurekci, Y.; Aykac, K. Oxidative stress related diseases in newborns. Oxid. Med. Cell. Longev. 2016, 2016, 2768365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koseesirikul, P.; Chotinaruemol, S.; Ukarapol, N. Incidence and risk factors of parenteral nutrition-associated liver disease in newborn infants. Pediatr. Int. 2012, 54, 434–436. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wang, J.; Zhang, T.; Huang, L.; Yan, W.; Lu, L.; Jia, J.; Tao, Y.; Cai, W.; Wang, Y. Alterations of gut microbiota and serum bile acids are associated with parenteral nutrition-associated liver disease. J. Pediatr. Surg. 2021, 56, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.; Manithody, C.; Krebs, J.; Denton, C.; Besmer, S.; Rajalakshmi, P.; Jain, S.; Villalona, G.A.; Jain, A.K. Impaired gut-systemic signaling drives total parenteral nutrition-associated injury. Nutrients 2020, 12, 1493. [Google Scholar] [CrossRef]
- Morin, G.; Guiraut, C.; Marcogliese, M.P.; Mohamed, I.; Lavoie, J.C. Glutathione supplementation of parenteral nutrition prevents oxidative stress and sustains protein synthesis in guinea pig model. Nutrients 2019, 11, 2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elremaly, W.; Mohamed, I.; Mialet-Marty, T.; Rouleau, T.; Lavoie, J.C. Ascorbylperoxide from parenteral nutrition induces an increase of redox potential of glutathione and loss of alveoli in newborn guinea pig lungs. Redox Biol. 2014, 2, 725–731. [Google Scholar] [CrossRef] [Green Version]
- Hoff, D.S.; Michaelson, A.S. Effects of light exposure on total parenteral nutrition and its implications in the neonatal population. J. Pediatr. Pharmacol. Ther. 2009, 14, 132–143. [Google Scholar] [CrossRef]
- Zhang, Y.; Roh, Y.J.; Han, S.J.; Park, I.; Lee, H.M.; Ok, Y.S.; Lee, B.C.; Lee, S.R. Role of selenoproteins in redox regulation of signaling and the antioxidant System: A review. Antioxidants 2020, 9, 383. [Google Scholar] [CrossRef]
- Graham, E.M.; Mishra, O.P.; Delivoria-Papadopoulos, M. Antioxidants and oxidative stress injuries to the brain in the perinatal period. Semin. Neonatol. 1998, 3, 75–85. [Google Scholar] [CrossRef]
- Lunec, J. Free radicals: Their involvement in disease process. Ann. Clin. Biochem. 1990, 27, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, J.; Mims, L.C.; Roesel, R.A. The effect of phototherapy on amino acid solutions containing multivitamins. J. Pediatr. 1980, 96, 284–286. [Google Scholar] [CrossRef]
- Lavoie, J.C.; Bélanger, S.; Spalinger, M.; Chessex, P. Admixture of a multivitamin preparation to parenteral nutrition: The major contributor to in vitro generation of peroxides. Pediatrics 1997, 99, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindeman, J.H.N.; Van Zoeren-Grobben, D.; Schrijver, J.; Speek, A.J.; Poorthuis, B.J.H.M.; Berger, H.M. The total free radical trapping ability of cord blood plasma in preterm and term babies. Pediatr. Res. 1989, 26, 20–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saldanha, R.L.; Cepeda, E.E.; Poland, R.L. The effect of vitamin E prophylaxis on the incidence and severity of bronchopulmonary dysplasia. J. Pediatr. 1982, 101, 89–93. [Google Scholar] [PubMed]
- Autor, A.P.; Frank, L.; Roberts, R.J. Developmental characteristics of pulmonary superoxide dismutase: Relationship to idiopathic respiratory distress syndrome. Pediatr. Res. 1976, 10, 154–158. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, J.L. Iron, plasma antioxidants, and the ‘oxygen radical disease of prematurity’. Am. J. Dis. Child. 1988, 142, 1341–1344. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, J.L.; Newton, R.B. Serum antioxidant activity in neonates. Arch. Dis. Child. 1988, 63, 748–750. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.C. Glutathione synthesis. Biochim. Biophys. Acta 2013, 1830, 3143–3153. [Google Scholar] [CrossRef] [Green Version]
- Elremaly, W.; Mohamed, I.; Rouleau, T.; Lavoie, J.C. Impact of glutathione supplementation of parenteral nutrition on hepatic methionine adenosyltransferase activity. Redox Biol. 2016, 8, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Lubos, E.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 in health and disease: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2011, 15, 1957–1997. [Google Scholar] [CrossRef] [Green Version]
- Miloudi, K.; Comte, B.; Rouleau, T.; Montoudis, A.; Levy, E.; Lavoie, J.C. The mode of administration of total parenteral nutrition and nature of lipid content influence the generation of peroxides and aldehydes. Clin. Nutr. 2012, 14, 526–534. [Google Scholar] [CrossRef]
- Jalabert, A.; Grand, A.; Steghens, J.P.; Barbotte, E.; Pique, C.; Picaud, J.C. Lipid peroxidation in all-in-one admixtures for preterm neonates: Impact of amount of lipid, type of lipid emulsion and delivery condition. Acta. Paediatr. 2011, 100, 1200–1205. [Google Scholar] [CrossRef]
- Neuzil, J.; Darlow, B.A.; Inder, T.E.; Sluis, K.B.; Winterbourn, C.C.; Stocker, R. Oxidation of parenteral lipid emulsion by ambient and phototherapy lights: Potential toxicity of routine parenteral feeding. J. Pediatr. 1995, 126, 785–790. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360–438. [Google Scholar] [CrossRef]
- Milne, G.L.; Yin, H.; Hardy, K.D.; Davies, S.S.; Roberts, L.J. Isoprostane generation and function. Chem. Rev. 2011, 111, 5973–5996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niki, Y.E. Detection of lipid peroxidation in vivo: Total hydroxy octadecadienoic acid and 7-hydroxycholesterol as oxidative stress marker. Free Radic. Res. 2004, 38, 787–794. [Google Scholar]
- Fell, G.L.; Nandivada, G.P.; Gura, K.M.; Puder, M. Intravenous lipid emulsions in parenteral nutrition. Adv. Nutr. 2015, 6, 600–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helbock, H.J.; Motchnik, P.A.; Ames, B.N. Toxic hydroperoxides in intravenous lipid emulsions used in preterm infants. Pediatrics 1993, 91, 83–88. [Google Scholar]
- Padayatty, S.J.; Levine, M. Vitamin C: The known and the unknown and Goldilocks. Oral Dis. 2016, 22, 463–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janda, K.; Kasprzak, M.; Wolska, J. Vitamin C–structure, properties, occurrence and functions. Pomeranian J. Life Sci. 2015, 61, 419–425. [Google Scholar] [PubMed]
- Reis, A.; Spickett, C.M. Chemistry of phospholipid oxidation. Biochim. Biophys. Acta Biomembr. 2012, 1818, 2374–2387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, L.J.; Fessel, J.P.; Davies, S.S. The biochemistry of the isoprostane, neuroprostane, and isofuran pathways of lipid peroxidation. Brain Pathol. 2005, 15, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Fessel, J.P.; Porter, N.A.; Moore, K.P.; Sheller, J.R.; Roberts, L.J. Discovery of lipid peroxidation products formed in vivo with a substituted tetrahydrofuran ring (isofurans) that are favored by increased oxygen tension. Proc. Natl. Acad. Sci. USA 2002, 99, 16713–16718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esterbauer, H. Estimation of peroxidative damage. A critical review. Pathol. Biol. 1996, 44, 25–28. [Google Scholar] [PubMed]
- Dine, T.; Gressier, B.; Luyckx, M.; Gottrand, F.; Michaud, L.; Kambia, N. Plasma malondialdehyde levels in children on 12-hour cyclic parenteral nutrition: Are there health risks? Pediatr. Dev. Pathol. 2014, 17, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, B.; Watorek, K.; Strauss, R.; Witz, G.; Hiatt, M.; Hegyi, T. Association of lipid peroxidation with hepatocellular injury in preterm infants. Crit. Care 2002, 6, 521–525. [Google Scholar] [CrossRef]
- Halliwell, B. Lipid peroxidation, antioxidants and cardiovascular disease: How should we move forward? Cardiovasc. Res. 2000, 47, 410–418. [Google Scholar] [CrossRef] [Green Version]
- Esterbauer, H.; Schaur, R.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal malonedialdehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Giera, M.; Lingeman, H.; Niessen, W.M. Recent advancements in the LC- and GC-based analysis of malondialdehyde (MDA): A brief overview. Chromatographia 2012, 75, 433–440. [Google Scholar] [CrossRef] [Green Version]
- Lavoie, J.-C.; Mohamed, I.; Nuyt, A.-M.; Elremaly, W.; Rouleau, T. Impact of SMOFLipid on pulmonary alveolar development in newborn guinea pigs. J. Parenter. Enter. Nutr. 2018, 42, 1314–1321. [Google Scholar] [CrossRef]
- Leguina-Ruzzi, A.A.; Ortiz, R. Current evidence for the use of Smoflipid® emulsion in critical care patients for parenteral Nutrition. Crit. Care Res. Pract. 2018, 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Pastor-Clerigues, A.; Marti-Bonmati, E.; Milara, J.; Almudever, P.; Cortijo, J. Anti-inflammatory and anti-fibrotic profile of fish oil emulsions used in parenteral nutrition-associated liver disease. PLoS ONE 2014, 9, e115404. [Google Scholar] [CrossRef]
- Burns, D.L.; Gill, B.M. Reversal of parenteral nutrition-associated liver disease with a fish oil-based lipid emulsion (Omegaven) in an adult dependent on home parenteral nutrition. J. Parenter. Enter. Nutr. 2013, 37, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Yarandi, S.S.; Zhao, V.M.; Hebbar, G.; Ziegler, T.R. Amino acid composition in parenteral nutrition: What is the evidence? Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Brunton, J.A.; Shoveller, A.K.; Pencharz, P.B.; Ball, R.O. The indicator amino acid oxidation method identified limiting amino acids in two parenteral nutrition solutions in neonatal piglets. J. Nutr. 2007, 137, 1253–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raftery, M.J. Determination of oxidative protein modifications using mass spectrometry. Redox Rep. 2014, 19, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Verrastro, I.; Pasha, S.; Jensen, K.T.; Pitt, A.R.; Spickett, C.M. Mass spectrometry-based methods for identifying oxidized proteins in disease: Advances and challenges. Biomolecules 2015, 5, 378–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Xiao, S.; Ahn, D.U. Protein oxidation: Basic principles and implications for meat quality. Crit. Rev. Food. Sci. Nutr. 2013, 53, 1191–1201. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, C.E.; Carroll, K.S. Cysteine-mediated redox signaling: Chemistry, biology, and tools for discovery. Chem. Rev. 2013, 113, 4633–4679. [Google Scholar] [CrossRef]
- Te Braake, F.W.J.; Schierbeek, H.; Vermes, A.; Huijmans, J.G.M.; van Goudoever, J.B. High-dose cysteine administration does not increase synthesis of the antioxidant glutathione preterm infants. Pediatrics 2009, 124, 978–984. [Google Scholar] [CrossRef]
- Soghier, L.M.; Brion, L.P. Cysteine, cystine or N-acetylcysteine supplementation in parenterally fed neonates. Cochrane Database Syst. Rev. 2006, 18, CD004869. [Google Scholar] [CrossRef]
- Stadtman, E.R.; Moskovitz, J.; Levine, R.L. Oxidation of methionine residues of proteins: Biological consequences. Antioxid. Redox Signal. 2003, 5, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Boullata, J.I.; Gilbert, K.; Sacks, G.; Labossiere, R.J.; Crill, C.; Goday, P.; Kumpf, V.J.; Mattox, T.W.; Plogsted, S.; Holcombe, B.; et al. Clinical guidelines: Parenteral nutrition ordering, order review, compounding, labeling, and dispensing. J. Parenter. Enter. Nutr. 2014, 38, 334–377. [Google Scholar] [CrossRef] [Green Version]
- Oliver, C.; Watson, C.; Crowley, E.; Gilroy, M.; Page, D.; Weber, K.; Messina, D.; Cormack, B. Vitamin and mineral supplementation practices in preterm infants: A survey of Australian and New Zealand neonatal intensive and special care units. Nutrients 2019, 12, 51. [Google Scholar] [CrossRef] [Green Version]
- Abdul-Razzak, K.K.; Nusier, M.K.; Obediat, A.D.; Salim, A.M. Antioxidant vitamins and hyperbilirubinemia in neonates. Ger. Med. Sci. 2007, 5, Doc03. [Google Scholar]
- Kaz’mierczak-Baran´ska, J.; Boguszewska, K.; Adamus-Grabicka, A.; Karwowski, B.T. Two faces of vitamin C-antioxidative and pro-oxidative agent. Nutrients 2020, 12, 1501. [Google Scholar] [CrossRef]
- Bassiouny, M.R.; Almarsafawy, H.; Abdel-Hady, H.; Nasef, N.; Hammad, T.A.; Aly, H. A randomized controlled trial on parenteral nutrition, oxidative stress, and chronic lung diseases in preterm infants. J. Pediatr. Gastroenterol. Nutr. 2009, 48, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Ha, E.; Wang, W.; John Wang, Y. Peroxide formation in polysorbate 80 and protein stability. J. Pharm. Sci. 2002, 91, 2252–2264. [Google Scholar] [CrossRef]
- Laborie, S.; Lavoie, J.C.; Chessex, P. Paradoxical role of ascorbic acid and riboflavin in solutions of total parenteral nutrition: Implication in photoinduced peroxide generation. Pediatr. Res. 1998, 43, 601–606. [Google Scholar] [CrossRef] [Green Version]
- Pinto, J.T.; Zempleni, J. Riboflavin. Adv. Nutr. 2016, 7, 973–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Kirschenbaum, L.J.; Rosenthal, I.; Riesz, P. Photosensitized formation of ascorbate radicals by riboflavin: An ESR study. Photochem. Photobiol. 1993, 57, 777–784. [Google Scholar] [CrossRef]
- Khan, S.; Rayis, M.; Rizvi, A.; Alam, M.M.; Rizvi, M.; Naseem, I. ROS mediated antibacterial activity of photoilluminated riboflavin: A photodynamic mechanism against nosocomial infections. Toxicol. Rep. 2019, 6, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.C.; Zhu, B.; Frei, B. Potential antiatherogenic mechanisms of ascorbate (vitamin C) and α-tocopherol (vitamin E). Circ. Res. 2000, 87, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Bowry, V.W.; Mohr, D.; Cleary, J.; Stocker, R. Prevention of tocopherol-mediated peroxidation in ubiquinol-10-free human low-density lipoprotein. J. Biol. Chem. 1995, 270, 5756–5763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, K.; Stoll, B.; Chacko, S.; de Pipaon, M.S.; Lauridsen, C.; Gray, M.; Squires, E.J.; Marini, J.; Zamora, I.J.; Olutoye, O.O.; et al. Vitamin E in new-generation lipid emulsions protects against parenteral nutrition-associated liver disease in parenteral nutrition-fed preterm pigs. J. Parenter. Enter. Nutr. 2016, 40, 656–671. [Google Scholar] [CrossRef] [Green Version]
- Burrin, D.G.; Ng, K.; Stoll, B.; Sáenz De Pipaón, M. Impact of new-generation lipid emulsions on cellular mechanisms of parenteral nutrition-associated liver disease. Adv Nutr. 2014, 5, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Muto, M.; Lim, D.; Soukvilay, A.; Field, C.; Wizzard, P.R.; Goruk, S.; Ball, R.O.; Pencharz, P.B.; Si, M.; Curtis, J.; et al. Supplemental parenteral vitamin E into conventional soybean lipid emulsion does not prevent parenteral nutrition-associated liver disease in full-term neonatal piglets. J. Parenter. Enter. Nutr. 2017, 41, 575–582. [Google Scholar] [CrossRef]
- Niki, E. Tocopherylquinone and tocopherylhydroquinone. Redox Rep. 2007, 12, 204–210. [Google Scholar] [CrossRef]
- Esmaeilizadeh, M.; Hosseini, M.; Beheshti, F.; Alikhani, V.; Keshavarzi, Z.; Shoja, M.; Mansoorian, M.; Sadeghnia, H.R. Vitamin C improves liver and renal functions in hypothyroid rats by reducing tissue oxidative injury. Int. J. Vitam. Nutr. Res. 2020, 90, 84–94. [Google Scholar] [CrossRef]
- Dennis, J.M.; Witting, P.K. Protective role for antioxidants in acute kidney disease. Nutrients 2017, 9, 718. [Google Scholar] [CrossRef] [Green Version]
- Pohanka, M.; Pejchal, J.; Snopkova, S.; Havlickova, K.; Karasova, J.Z.; Bostik, P.; Pikula, J. Ascorbic acid: An old player with a broad impact on body physiology including oxidative stress suppression and immunomodulation: A review. Mini. Rev. Med. Chem. 2012, 12, 35–43. [Google Scholar] [CrossRef]
- Carr, A.; Frei, B. Does vitamin C act as a pro-oxidant under physiological conditions? FASEB J. 1999, 13, 1007–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buettner, G.R. The pecking order of free radicals and antioxidants: Lipid peroxidation, α-tocopherol, and ascorbate. Arch. Biochem. Biophys. 1993, 300, 535–543. [Google Scholar] [CrossRef]
- Buettner, G.R.; Jurkiewicz, B.A. Catalytic metals, ascorbate and free radicals: Combinations to avoid. Radiat. Res. 1996, 145, 532–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halliwell, B. Vitamin C: Antioxidant or pro-oxidant in vivo? Free Radic. Res. 1996, 25, 439–454. [Google Scholar] [CrossRef]
- Knafo, L.; Chessex, P.; Rouleau, T.; Lavoie, J.C. Association between hydrogen peroxide-dependent byproducts of ascorbic acid and increased hepatic acetyl-CoA carboxylase activity. Clin. Chem. 2005, 51, 1462–1471. [Google Scholar] [CrossRef]
- Deutsch, J.C. Ascorbic acid oxidation by hydrogen peroxide. Anal. Biochem. 1998, 255, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, I.; Elremaly, W.; Rouleau, T.; Lavoie, J.C. Ascorbylperoxide contaminating parenteral nutrition is associated with bronchopulmonary dysplasia or death in extremely preterm infants. J. Parenter. Enter. Nutr. 2016, 41, 1023–1029. [Google Scholar] [CrossRef]
- Burge, J.C.; Flancbaum, L.; Holcombe, B. Copper decreases ascorbic acid stability in total parenteral nutrition solutions. Res. Prof. Brief 1994, 94, 777–779. [Google Scholar] [CrossRef]
- Jin, J.; Mulesa, L.; Carrilero Rouillet, M. Trace elements in parenteral nutrition: Considerations for the prescribing clinician. Nutrients 2017, 9, 440. [Google Scholar] [CrossRef] [PubMed]
- Stehle, P.; Stoffel-Wagner, B.; Kuhn, K.S. Parenteral trace element provision: Recent clinical research and practical conclusions. Eur. J. Clin. Nutr. 2016, 70, 886–893. [Google Scholar] [CrossRef] [Green Version]
- Frankel, D.A. Supplementation of trace elements in parenteral nutrition: Rationale and recommendations. Nutr. Res. 1993, 13, 583–596. [Google Scholar] [CrossRef]
- Bertolo, R.F.; Pencharz, P.B.; Ball, R.O. Tissue mineral concentrations are profoundly altered in neonatal piglets fed identical diets via gastric, central venous, or portal venous routes. J. Parenter. Enter. Nutr. 2014, 38, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Steger, P.J.; Mühlebach, S.F. Lipid peroxidation of intravenous lipid emulsions and all-in-one admixtures in total parenteral nutrition bags: The influence of trace elements. J. Parenter. Enter. Nutr. 2000, 24, 37–41. [Google Scholar] [CrossRef]
- Pollak, A.; Hayde, M.; Hayn, M.; Herkner, K.; Lombard, K.A.; Lubec, G.; Weninger, M.; Widness, J.A. Effect of intravenous iron supplementation on erythropoiesis in erythropoietin-treated premature infants. Pediatrics 2001, 107, 78–85. [Google Scholar] [CrossRef]
- Meyer, M.P.; Haworth, C.; Meyer, J.H.; Commerford, A. A comparison of oral and intravenous iron supplementation in preterm infants receiving recombinant erythropoietin. J. Pediatr. 1996, 129, 258–263. [Google Scholar] [CrossRef]
- Grand, A.; Jalabert, A.; Mercier, G.; Florent, M.; Hansel-Esteller, S.G.; Cambonie, G.; Steghens, J.P.; Picaud, J.C. Influence of vitamins, trace elements, and iron on lipid peroxidation reactions in all-in-one admixtures for neonatal parenteral nutrition. J. Parenter. Enter. Nutr. 2011, 35, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Jansson, P.J.; Lindqvist, C.; Nordström, T. Iron prevents ascorbic acid (vitamin C) induce hydrogen peroxide accumulation in copper contaminated drinking water. Free Radic. Res. 2005, 39, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Laborie, S.; Lavoie, J.C.; Pineault, M.; Chessex, P. Contribution of multivitamins, air, and light in the generation of peroxides in adult and neonatal parenteral nutrition solutions. Ann. Pharmacother. 2000, 34, 440–445. [Google Scholar] [CrossRef] [PubMed]
- Picaud, J.C.; Steghens, J.P.; Auxenfans, C.; Barbieux, A.; Laborie, S.; Claris, O. Lipid peroxidation assessment by malondialdehyde measurement in parenteral nutrition solutions for newborn infants: A pilot study. Acta Paediatr. 2004, 93, 241–245. [Google Scholar] [CrossRef]
- Brawley, V.; Bathia, J.; Karp, B. Hydrogen peroxide generation in a model pediatric parenteral amino acid solution. Clin. Sci. 1993, 85, 709–712. [Google Scholar] [CrossRef] [PubMed]
- Shattuck, K.E.; Bhatia, J.; Grinnell, C.; Rassin, D.K. The effects of light exposure on the in vitro hepatic response to an amino acid-vitamin solution. J. Parenter. Enter. Nutr. 1995, 19, 398–402. [Google Scholar] [CrossRef]
- Chessex, P.; Laborie, S.; Nasef, N.; Masse, B.; Lavoie, J.C. Shielding parenteral nutrition from light improves survival rate in premature infants. J. Parenter. Enter. Nutr. 2017, 41, 378–383. [Google Scholar] [CrossRef]
- Vivekanandan-Giri, A.; Byun, J.; Pennathur, S. Quantitative analysis of amino acid oxidation markers by tandem mass spectrometry. Methods Enzymol. 2011, 491, 73–89. [Google Scholar]
- Yailian, A.L.; Serre, C.; Fayard, J.; Faucon, M.; Thomaré, P.; Filali, S.; Pivot, C.; Vételé, F.; Pirot, F.; Olivier, E. Production and stability study of a hospital parenteral nutrition solution for neonates. J. Pharm. Anal. 2019, 9, 83–90. [Google Scholar] [CrossRef]
- Pitkänen, O.M. Peroxidation of lipid emulsions: A hazard for the premature infant receiving parenteral nutrition? Free Radic. Biol. Med. 1992, 13, 239–245. [Google Scholar] [CrossRef]
- Pincemail, J.; Deby, C.; Lismonde, M.; Bertrand, Y.; Camus, G.; Lamy, M. Pentane measurement, an index of in vivo lipoperoxidation: Applications and limits of the method. In Free Radicals, Lipoproteins, and Membrane Lipids; De Paulet, A.C., Douste-Blazy, L., Paoletti, R., Eds.; NATO ASI Series (Series A: Life Sciences); Springer: New York, NY, USA, 1990; Volume 189, pp. 163–169. [Google Scholar]
- Kambia, K.; Dine, T.; Gressier, B.; Bah, S.; Germe, A.F.; Luyckx, M.; Brunet, C.; Michaud, L.; Gottrand, F. Evaluation of childhood exposure to di(2-ethylhexyl) phthalate from perfusion kits during long-term parenteral nutrition. Int. J. Pharm. 2003, 262, 83–91. [Google Scholar] [CrossRef]
- Mazur, H.I.; Stennett, D.J.; Egging, P.K. Extraction of di(2-ethylhexyl) phthalate from total nutrient solution containing polyvinyl chloride bags. J. Parent. Enter. Nutr. 1989, 13, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Faouzi, M.A.; Dine, T.; Luyckx, M.; Brunet, C.; Mallevais, M.L.; Goudaliez, F.; Gressier, B.; Cazin, M.; Kablan, J.; Cazin, J.C. Stability and plasticizer extraction of miconazole injection added to infusion solutions and stored in polyvinyl chloride containers. J. Pharm. Biomed. Anal. 1995, 13, 1363–1372. [Google Scholar] [CrossRef]
- Simmchen, J.; Ventura, R.; Segura, J. Progress in the removal of di-[2-ethylhexyl]-phthalate as plasticizer in blood bags. Transfus. Med. Rev. 2012, 26, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Balet, A.; Cardona, D.; Jane, S.; Molins-Pujol, A.; Sanchez Quesada, J.; Gich, I.; Mangues, M. Effects of multilayered bags vs ethylvinyl-acetate bags on oxidation of parenteral nutrition. J. Parenter. Enter. Nutr. 2004, 28, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Loff, S.; Hannmann, T.; Subotic, U.; Reinecke, F.M.; Wischmann, H.; Brade, J. Extraction of diethylhexylphthalate by home total parenteral nutrition from polyvinyl chloride infusion lines commonly used in the home. J. Pediatr. Gastroenterol. Nutr. 2008, 47, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Marie, C.; Hamlaoui, S.; Bernard, L.; Bourdeaux, D.; Sautou, V.; Lémery, D.; Vendittelli, F.; Sauvant-Rochat, M.P. Exposure of hospitalised pregnant women to plasticizers contained in medical devices. BMC Women’s Health 2017, 17, 45. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, F.; Shen, G.; Yang, Y.; Tang, Y. Comparison of the release behaviors of di (2-ethylhexyl) phthalate and tri(2-ethylhexyl) trimellitate from the polyvinyl-chloride infusion set into pharmaceutical solutions. Se Pu 2015, 33, 522–529. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, I.; Elremaly, W.; Rouleau, T.; Lavoie, J.C. Oxygen and parenteral nutrition two main oxidants for extremely preterm infants: “it all adds up”. J. Neonatal Perinat. Med. 2015, 8, 189–197. [Google Scholar] [CrossRef]
- Deshpande, G.C.; Simmer, K.; Mori, T.; Croft, K. Parenteral lipid emulsions based on olive oil compared with soybean oil in preterm (<28 weeks’ gestation) neonates: A randomised controlled trial. J. Pediatr. Gastroenterol. Nutr. 2009, 49, 619–625. [Google Scholar] [CrossRef]
- Unal, S.; Demirel, N.; Erol, S.; Isik, D.U.; Kulali, F.; Iyigun, F.; Bas, A.Y. Effects of two different lipid emulsions on morbidities and oxidant stress statuses in preterm infants: An observational study. J. Matern. Fetal Neonatal Med. 2017, 31, 850–856. [Google Scholar] [CrossRef]
- Chessex, P.; Friel, J.; Harrison, A. The mode of delivery of parenteral multivitamins influences nutrient handling in an animal model of total parenteral nutrition. Clin. Nutr. 2005, 24, 281–287. [Google Scholar] [CrossRef]
- Chessex, P.; Lavoie, J.C.; Rouleau, T.; Brochu, P.; St-Louis, P.; Lévy, E.; Alvarez, F. Photooxidation of parenteral multivitamins induces hepatic steatosis in a neonatal guinea pig model of intravenous nutrition. Pediatr. Res. 2002, 52, 958–963. [Google Scholar] [CrossRef]
- Żalikowska-Gardocka, M.; Przybyłkowski, A. Review of parenteral nutrition-associated liver disease. Clin. Exp. Hepatol. 2020, 6, 65–73. [Google Scholar] [CrossRef]
- Bhatia, J.; Moslen, M.T.; Haque, A.K.; McCleery, R.; Rassin, D.K. Total parenteral nutrition-associated alterations in hebatobiliary function and histology in rats: Is light exposure a clue? Pediatr. Res. 1993, 33, 487–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nandivada, P.; Carlson, S.J.; Chang, M.I.; Cowan, E.; Gura, K.M.; Puder, M. Treatment of parenteral nutrition-associated liver disease: The role of lipid emulsions. Adv. Nutr. 2013, 4, 711–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waitzberg, D.L.; Torrinhas, R.S. Fish oil lipid emulsions and immune response: What clinicians need to know. Nutr. Clin. Pract. 2009, 24, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Kurvinen, A.; Nissinen, M.J.; Andersson, S.; Korhonen, P.; Ruuska, T.; Taimisto, M.; Kalliomäki, M.; Lehtonen, L.; Sankilampi, U.; Arikoski, P.; et al. Parenteral plant sterols and intestinal failure–associated liver disease in neonates. J. Pediatr. Gastroenterol. Nutr. 2012, 54, 803–811. [Google Scholar] [CrossRef]
- Biesalski, H.K. Vitamin E requirements in parenteral nutrition. Gastroenterology 2009, 137, 92–104. [Google Scholar] [CrossRef]
- Diamond, I.R.; de Silva, N.T.; Tomlinson, G.A.; Pencharz, P.B.; Feldman, B.M.; Moore, A.M.; Ling, S.C.; Wales, P.W. The role of parenteral lipids in the development of advanced intestinal failure-associated liver disease in infants: A multiple-variable analysis. J. Parenter. Enter. Nutr. 2011, 35, 596–602. [Google Scholar] [CrossRef]
- Le, H.D.; de Meijer, V.E.; Robinson, E.M.; Zurakowski, D.; Potemkin, A.K.; Arsenault, D.A.; Fallon, E.M.; Malkan, A.; Bistrian, B.R.; Gura, K.M.; et al. Parenteral fish-oil-based lipid emulsion improves fatty acid profiles and lipids in parenteral nutrition dependent children. Am. J. Clin. Nutr. 2011, 94, 749–758. [Google Scholar] [CrossRef] [Green Version]
- Premkumar, M.H.; Carter, B.A.; Hawthorne, K.M.; King, K.; Abrams, S.A. Fish oil-based lipid emulsions in the treatment of parenteral nutrition-associated liver disease: An ongoing positive experience. Adv. Nutr. 2014, 5, 65–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, A.K.; Stoll, B.; Burrin, D.G.; Holst, J.J.; Moore, D.D. Enteral bile acid treatment improves parenteral nutrition-related liver disease and intestinal mucosal atrophy in neonatal pigs. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, 218–224. [Google Scholar] [CrossRef] [Green Version]
- Pironi, L.; Sasdelli, A.S. Intestinal failure-associated liver disease. Clin. Liver Dis. 2019, 23, 279–291. [Google Scholar] [CrossRef]
- Rochling, F.A.; Catron, H.A. Intestinal failure-associated liver disease: Causes, manifestations and therapies. Curr. Opin. Gastroenterol. 2019, 35, 126–133. [Google Scholar] [CrossRef]
- Lacaille, F.; Gupte, G.; Colomb, V.; d’Antiga, L.; Hartman, C.; Hojsak, I.; Kolacek, S.; Puntis, J.; Shamir, R. Intestinal failure associated liver disease: A position paper of the ESPGHAN working group of intestinal failure and intestinal transplantation. J. Pediatr. Gastroenterol. Nutr. 2015, 60, 272–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madnawat, H.; Welu, A.L.; Gilbert, E.J.; Taylor, D.B.; Jain, S.; Manithody, C.; Blomenkamp, K.; Jain, A.K. Mechanisms of parenteral nutrition-associated liver and gut injury. Nutr. Clin. Pract. 2020, 35, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Kir, S.; Beddow, S.A.; Samuel, V.T.; Miller, P.; Previs, S.F.; Suino-Powell, K.; Xu, H.E.; Shulman, G.I.; Kliewer, S.A.; Mangelsdorf, D.J. FGF19 as a postprandial, insulin-independent activator of hepatic protein and glycogen synthesis. Science 2011, 331, 1621–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagaki, T.; Choi, M.; Moschetta, A.; Peng, L.; Cummins, C.L.; McDonald, J.G.; Luo, G.; Jones, S.A.; Goodwin, B.; Richardson, J.A.; et al. Fibroblast growth factor 15 functions as an enterohepatic signal to regulate bile acid homeostasis. Cell Metab. 2005, 2, 217–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertolo, R.F.; Brunton, J.A.; Pencharz, P.B.; Ball, R.O. Arginine, ornithine, and proline interconversion is dependent on small intestinal metabolism in neonatal pigs. Am. J. Physiol. Endocrinol. Metab. 2003, 284, 915–922. [Google Scholar] [CrossRef] [Green Version]
- Niinikoski, H.; Stoll, B.; Guan, X.; Kansagra, K.; Lambert, B.D.; Stephens, J.; Hartmann, B.; Holst, J.J.; Burrin, D.G. Onset of small intestinal atrophy is associated with reduced intestinal blood flow in TPN-fed neonatal piglets. J. Nutr. 2004, 134, 1467–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezzati, M.; Dani, C.; Tronchin, M.; Filippi, L.; Rossi, S.; Rubaltelli, F.F. Prediction of early tolerance to enteral feeding by measurement of superior mesenteric artery blood flow velocity: Appropriate- versus small-for-gestational-age preterm infants. Acta Paediatr. 2004, 93, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Kempley, S.T.; Gamsu, H.R. Prediction of early tolerance to enteral feeding in preterm infants by measurement of superior mesenteric artery blood flow velocity. Arch. Dis. Child Fetal Neonatal Ed. 2001, 85, 42–45. [Google Scholar] [CrossRef] [Green Version]
- Dinesh, O.C.; Dodge, M.E.; Baldwin, M.P.; Bertolo, R.F.; Brunton, J.A. Enteral arginine partially ameliorates parenteral nutrition–induced small intestinal atrophy and stimulates hepatic protein synthesis in neonatal piglets. J. Parenter. Enter. Nutr. 2013, 38, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Huber, L.A.; Lee, T.; LeDrew, R.L.; Dodge, M.E.; Brunton, J.A.; Bertolo, R.F. Photoprotection but not N-acetylcysteine improves intestinal blood flow and oxidation status in parenterally fed piglets. J. Pediatr. Gastroenterol. Nutr. 2019, 69, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Ticho, A.L.; Malhotra, P.; Dudeja, P.K.; Gill, R.K.; Alrefai, W.A. Bile acid receptors and gastrointestinal functions. Liver Res. 2019, 3, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Pathak, P.; Xie, C.; Nichols, R.G.; Ferrell, J.M.; Boehme, S.; Krausz, K.W.; Patterson, A.D.; Gonzalez, F.J.; Chiang, J.Y.L. Intestine farnesoid X receptor agonist and the gut microbiota activate G-protein bile acid receptor-1 signaling to improve metabolism. Hepatology 2018, 68, 1574–1588. [Google Scholar] [CrossRef]
- Burrin, D.; Stoll, B.; Moore, D. Digestive physiology of the pig symposium: Intestinal bile acid sensing is linked to key endocrine and metabolic signaling pathways. J. Anim. Sci. 2013, 91, 1991–2000. [Google Scholar] [CrossRef]
- Keitel, V.; Stindt, J.; Häussinger, D. Bile acid-activated receptors: GPBAR1 (TGR5) and other G protein-coupled receptors. Handb. Exp. Pharmacol. 2019, 256, 19–49. [Google Scholar]
- Jain, A.K.; Wen, J.X.; Blomenkamp, K.S.; Arora, S.; Blaufuss, T.A.; Rodrigues, J.; Long, J.P.; Neuschwander-Tetri, B.A.; Teckman, J.H. Oleanolic acid improves gut atrophy induced by parenteral nutrition. J. Parenter. Enter. Nutr. 2015, 40, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Lavoie, J.C.; Rouleau, T.; Gagnon, C.; Chessex, P. Photoprotection prevents TPN induced lung procollagen mRNA in newborn guinea pigs. Free Radic. Biol. Med. 2002, 33, 512–520. [Google Scholar] [CrossRef]
- Lavoie, J.C.; Rouleau, T.; Chessex, P. Interaction between ascorbate and light-exposed riboflavin induces lung remodeling. J. Pharmacol. Exp. Ther. 2004, 31, 634–639. [Google Scholar] [CrossRef] [Green Version]
- Elremaly, W.; Mohamed, I.; Rouleau, T.; Lavoie, J.C. Adding glutathione to parenteral nutrition prevents alveolar loss in newborn guinea pig. Free Radic. Biol. Med. 2015, 87, 274–281. [Google Scholar] [CrossRef]
- Shichiri, M. The role of lipid peroxidation in neurological disorders. J. Clin. Biochem. Nutr. 2014, 54, 151–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrone, S.; Laschi, E.; Buonocore, G. Biomarkers of oxidative stress in the fetus and in the newborn. Free Radic. Biol. Med. 2019, 142, 23–31. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karthigesu, K.; Bertolo, R.F.; Brown, R.J. Parenteral Nutrition and Oxidant Load in Neonates. Nutrients 2021, 13, 2631. https://doi.org/10.3390/nu13082631
Karthigesu K, Bertolo RF, Brown RJ. Parenteral Nutrition and Oxidant Load in Neonates. Nutrients. 2021; 13(8):2631. https://doi.org/10.3390/nu13082631
Chicago/Turabian StyleKarthigesu, Kandeepan, Robert F. Bertolo, and Robert J. Brown. 2021. "Parenteral Nutrition and Oxidant Load in Neonates" Nutrients 13, no. 8: 2631. https://doi.org/10.3390/nu13082631
APA StyleKarthigesu, K., Bertolo, R. F., & Brown, R. J. (2021). Parenteral Nutrition and Oxidant Load in Neonates. Nutrients, 13(8), 2631. https://doi.org/10.3390/nu13082631