
Fructose plus High-Salt Diet in Early Life Results in Salt-Sensitive Cardiovascular Changes in Mature Male Sprague Dawley Rats

Abstract
:1. Introduction
2. Materials and Methods
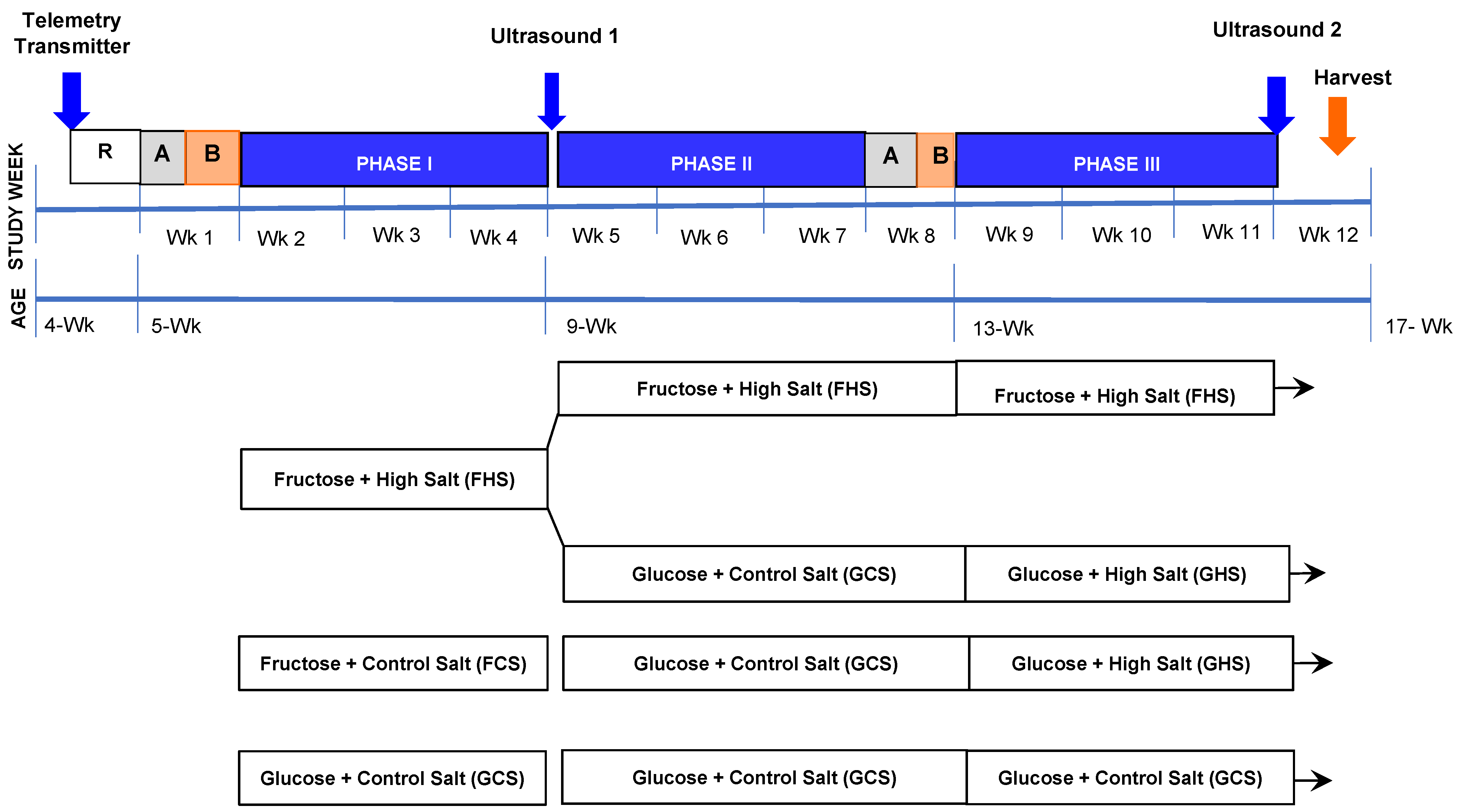
2.1. Dietary Regimen
2.2. Ultrasonography
2.3. Surgical Procedures
2.4. Analytical Measurements and Calculations
3. Results
3.1. Metabolic and Humoral Profiles
3.2. Hemodynamic Effects
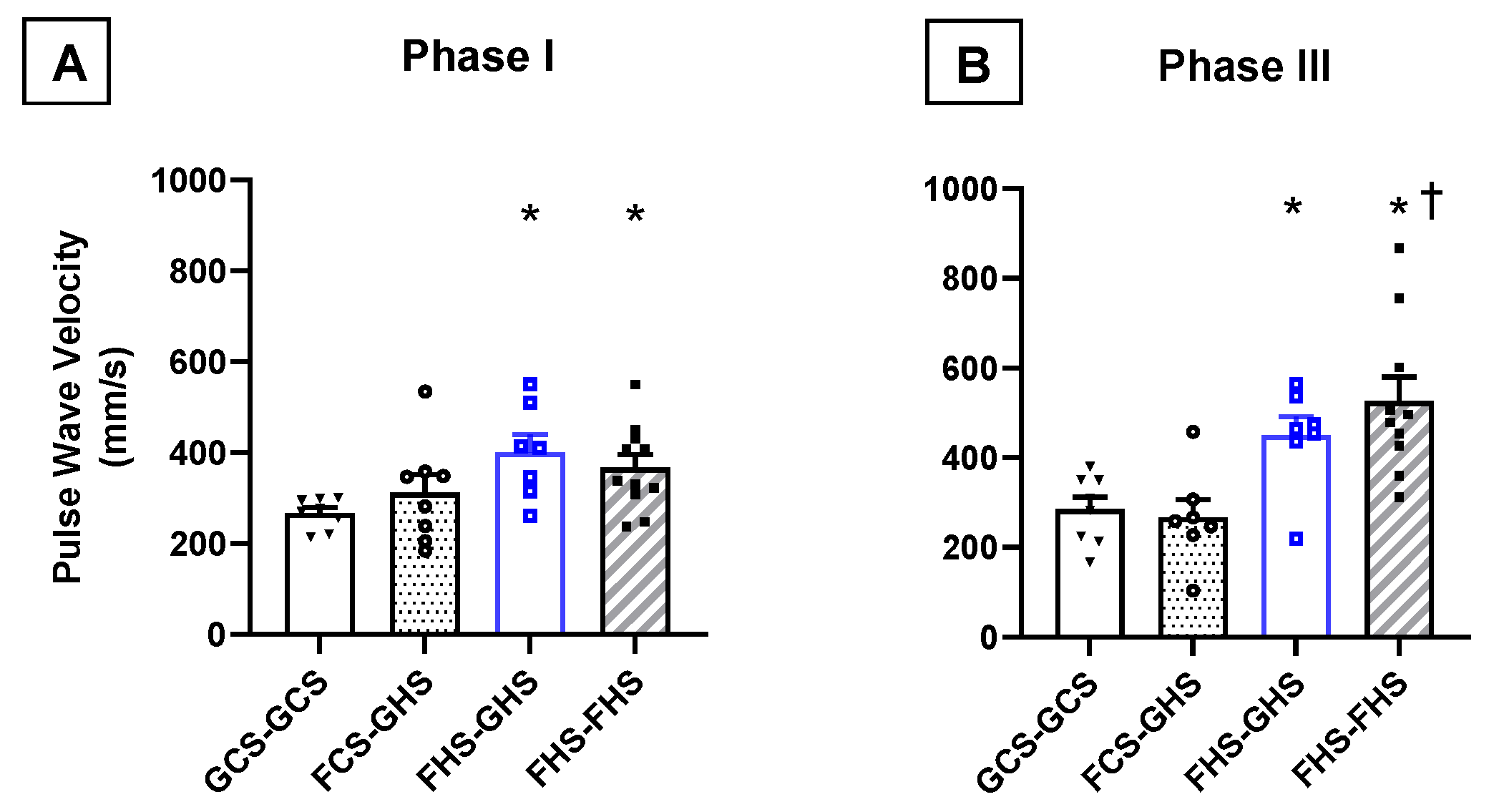
3.3. Measurements of Vascular Compliance
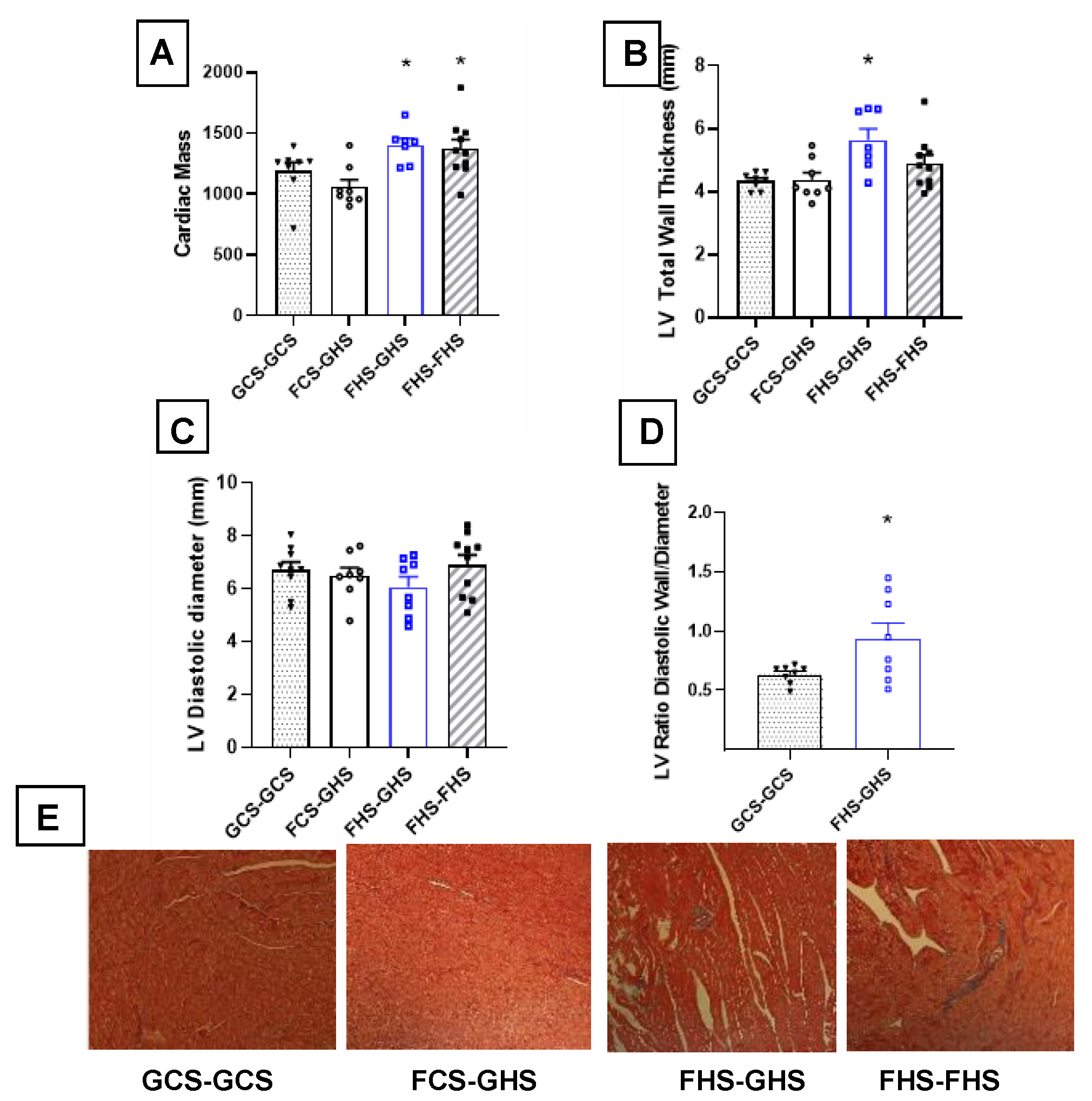
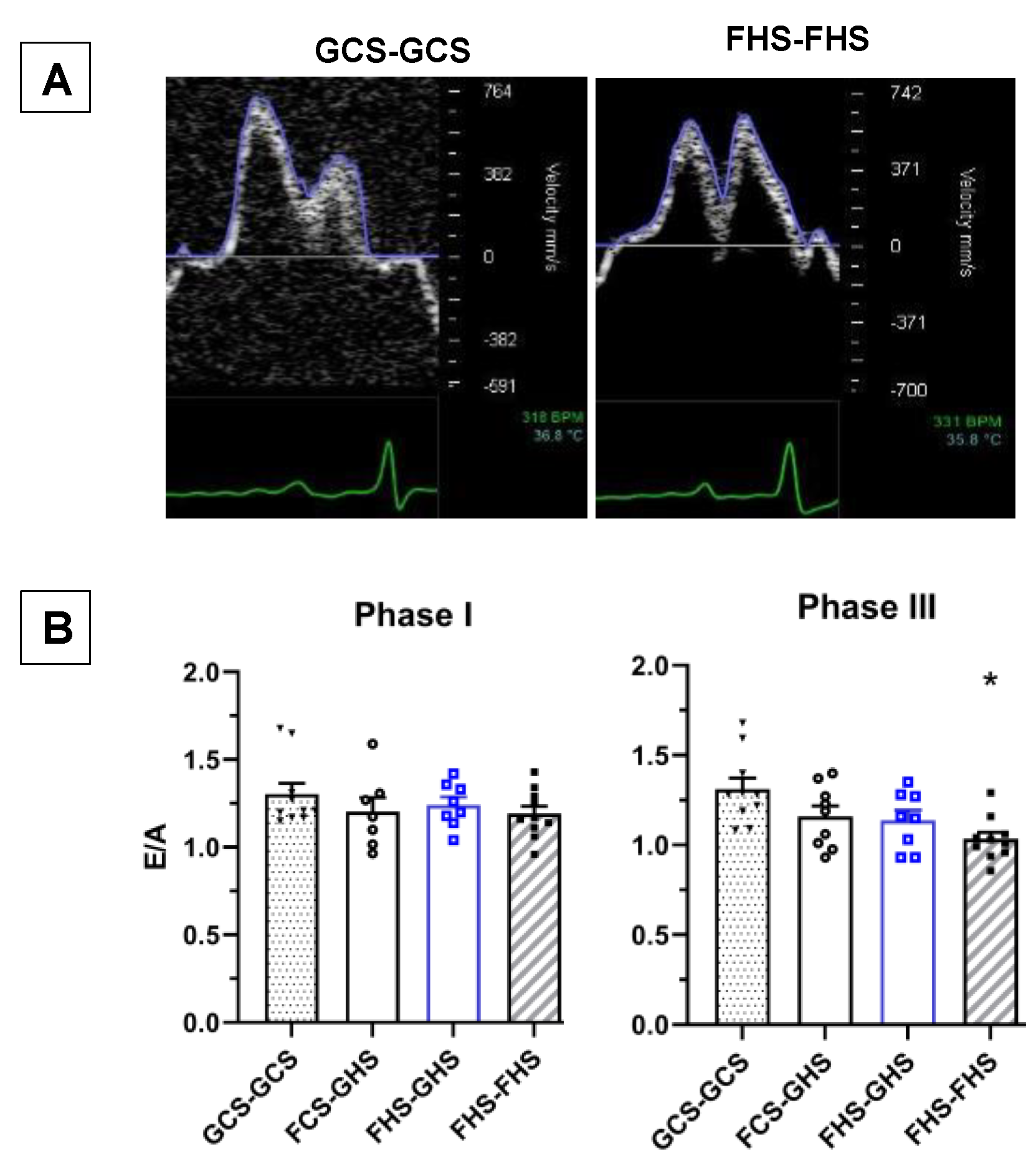
3.4. Echocardiographic Assessments
4. Discussion
4.1. Limitations
4.2. Perspectives
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ostchega, Y.; Fryar, C.D.; Nwankwo, T.; Nguyen, D.T. Hypertension Prevalence Among Adults Aged 18 and Over: United States, 2017–2018. NCHS Data Brief. 2020; pp. 1–8. Available online: https://www.cdc.gov/nchs/products/databriefs/db364.htm (accessed on 1 September 2021).
- Bray, G.A.; Nielsen, S.J.; Popkin, B.M. Consumption of high-fructose corn syrup in beverages may play a role in the epidemic of obesity. Am. J. Clin. Nutr. 2004, 79, 537–543. [Google Scholar] [CrossRef]
- Tappy, L.; Le, K.A. Metabolic effects of fructose and the worldwide increase in obesity. Physiol. Rev. 2010, 90, 23–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, L.T.; Yuen, V.G.; McNeill, J.H. The fructose-fed rat: A review on the mechanisms of fructose-induced insulin resistance and hypertension. Mol. Cell Biochem. 2009, 332, 145–159. [Google Scholar] [CrossRef]
- Martinez, F.J.; Rizza, R.A.; Romero, J.C. High-fructose feeding elicits insulin resistance, hyperinsulinism, and hypertension in normal mongrel dogs. Hypertension 1994, 23, 456–463. [Google Scholar] [CrossRef] [Green Version]
- Gordish, K.L.; Kassem, K.M.; Ortiz, P.A.; Beierwaltes, W.H. Moderate (20%) fructose-enriched diet stimulates salt-sensitive hypertension with increased salt retention and decreased renal nitric oxide. Physiol. Rep. 2017, 5. [Google Scholar] [CrossRef]
- Soncrant, T.; Komnenov, D.; Beierwaltes, W.H.; Chen, H.; Wu, M.; Rossi, N.F. Bilateral renal cryodenervation decreases arterial pressure and improves insulin sensitivity in fructose-fed Sprague-Dawley rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R529–R538. [Google Scholar] [CrossRef]
- Narain, A.; Kwok, C.S.; Mamas, M.A. Soft drinks and sweetened beverages and the risk of cardiovascular disease and mortality: A systematic review and meta-analysis. Int J. Clin. Pract. 2016, 70, 791–805. [Google Scholar] [CrossRef]
- Lelis, D.F.; Andrade, J.M.O.; Almenara, C.C.P.; Broseguini-Filho, G.B.; Mill, J.G.; Baldo, M.P. High fructose intake and the route towards cardiometabolic diseases. Life Sci. 2020, 259, 118235. [Google Scholar] [CrossRef]
- Eren, O.C.; Ortiz, A.; Afsar, B.; Covic, A.; Kuwabara, M.; Lanaspa, M.A.; Johnson, R.J.; Kanbay, M. Multilayered Interplay Between Fructose and Salt in Development of Hypertension. Hypertension 2019, 73, 265–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, A.V.; Kiat, H. The mechanisms underlying fructose-induced hypertension: A review. J. Hypertens. 2015, 33, 912–920. [Google Scholar] [CrossRef] [Green Version]
- Perez-Pozo, S.E.; Schold, J.; Nakagawa, T.; Sanchez-Lozada, L.G.; Johnson, R.J.; Lillo, J.L. Excessive fructose intake induces the features of metabolic syndrome in healthy adult men: Role of uric acid in the hypertensive response. Int. J. Obes. (Lond.) 2010, 34, 454–461. [Google Scholar] [CrossRef] [Green Version]
- Dhingra, R.; Sullivan, L.; Jacques, P.F.; Wang, T.J.; Fox, C.S.; Meigs, J.B.; D’Agostino, R.B.; Gaziano, J.M.; Vasan, R.S. Soft drink consumption and risk of developing cardiometabolic risk factors and the metabolic syndrome in middle-aged adults in the community. Circulation 2007, 116, 480–488. [Google Scholar] [CrossRef] [Green Version]
- Fung, T.T.; Malik, V.; Rexrode, K.M.; Manson, J.E.; Willett, W.C.; Hu, F.B. Sweetened beverage consumption and risk of coronary heart disease in women. Am. J. Clin. Nutr. 2009, 89, 1037–1042. [Google Scholar] [CrossRef] [Green Version]
- Aeberli, I.; Zimmermann, M.B.; Molinari, L.; Lehmann, R.; l’Allemand, D.; Spinas, G.A.; Berneis, K. Fructose intake is a predictor of LDL particle size in overweight schoolchildren. Am. J. Clin. Nutr. 2007, 86, 1174–1178. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.K.; Yetley, E.A. Intakes and food sources of fructose in the United States. Am. J. Clin. Nutr. 1993, 58, 737S–747S. [Google Scholar] [CrossRef]
- Marriott, B.P.; Cole, N.; Lee, E. National estimates of dietary fructose intake increased from 1977 to 2004 in the United States. J. Nutr. 2009, 139, 1228S–1235S. [Google Scholar] [CrossRef] [Green Version]
- Adriani, W.; Laviola, G. Windows of vulnerability to psychopathology and therapeutic strategy in the adolescent rodent model. Behav. Pharmacol. 2004, 15, 341–352. [Google Scholar] [CrossRef]
- Sengupta, P. The Laboratory Rat: Relating Its Age With Human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar] [PubMed]
- Zicha, J.; Kunes, J. Ontogenetic aspects of hypertension development: Analysis in the rat. Physiol. Rev. 1999, 79, 1227–1282. [Google Scholar] [CrossRef] [PubMed]
- Harrap, S.B.; Van der Merwe, W.M.; Griffin, S.A.; Macpherson, F.; Lever, A.F. Brief angiotensin converting enzyme inhibitor treatment in young spontaneously hypertensive rats reduces blood pressure long-term. Hypertension 1990, 16, 603–614. [Google Scholar] [CrossRef] [Green Version]
- Rughani, A.; Friedman, J.E.; Tryggestad, J.B. Type 2 Diabetes in Youth: The Role of Early Life Exposures. Curr. Diab. Rep. 2020, 20, 45. [Google Scholar] [CrossRef]
- Varlinskaya, E.I.; Spear, L.P. Social interactions in adolescent and adult Sprague-Dawley rats: Impact of social deprivation and test context familiarity. Behav. Brain Res. 2008, 188, 398–405. [Google Scholar] [CrossRef] [Green Version]
- Coatney, R.W. Ultrasound imaging: Principles and applications in rodent research. ILAR J. 2001, 42, 233–247. [Google Scholar] [CrossRef] [Green Version]
- Lindsey, M.L.; Kassiri, Z.; Virag, J.A.I.; de Castro Bras, L.E.; Scherrer-Crosbie, M. Guidelines for measuring cardiac physiology in mice. Am. J. Physiol. Heart Circ. Physiol. 2018, 314, H733–H752. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, W.C. Renal resistive index: A case of mistaken identity. Hypertension 2014, 64, 915–917. [Google Scholar] [CrossRef] [PubMed]
- Komnenov, D.; Levanovich, P.E.; Perecki, N.; Chung, C.S.; Rossi, N.F. Aortic Stiffness and Diastolic Dysfunction in Sprague Dawley Rats Consuming Short-Term Fructose Plus High Salt Diet. Integr. Blood Press Control 2020, 13, 111–124. [Google Scholar] [CrossRef]
- D’Angelo, G.; Elmarakby, A.A.; Pollock, D.M.; Stepp, D.W. Fructose feeding increases insulin resistance but not blood pressure in Sprague-Dawley rats. Hypertension 2005, 46, 806–811. [Google Scholar] [CrossRef] [Green Version]
- Reaven, G.M.; Ho, H.; Hoffman, B.B. Attenuation of fructose-induced hypertension in rats by exercise training. Hypertension 1988, 12, 129–132. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.D.; Zhang, H.Y.; Kotchen, T.A. Sucrose does not raise blood pressure in rats maintained on a low salt intake. Hypertension 1993, 21, 779–785. [Google Scholar] [CrossRef] [Green Version]
- Paixao, A.D.; Alexander, B.T. How the kidney is impacted by the perinatal maternal environment to develop hypertension. Biol. Reprod. 2013, 89, 144. [Google Scholar] [CrossRef]
- Gray, C.; Gardiner, S.M.; Elmes, M.; Gardner, D.S. Excess maternal salt or fructose intake programmes sex-specific, stress- and fructose-sensitive hypertension in the offspring. Br. J. Nutr. 2016, 115, 594–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.; Lv, J.; Mao, C.; Zhang, H.; Wang, A.; Zhu, L.; Zhu, H.; Xu, Z. High-salt diet during pregnancy and angiotensin-related cardiac changes. J. Hypertens. 2010, 28, 1290–1297. [Google Scholar] [CrossRef] [Green Version]
- Gray, C.; Al-Dujaili, E.A.; Sparrow, A.J.; Gardiner, S.M.; Craigon, J.; Welham, S.J.; Gardner, D.S. Excess maternal salt intake produces sex-specific hypertension in offspring: Putative roles for kidney and gastrointestinal sodium handling. PLoS ONE 2013, 8, e72682. [Google Scholar] [CrossRef]
- Hocherl, K.; Endemann, D.; Kammerl, M.C.; Grobecker, H.F.; Kurtz, A. Cyclo-oxygenase-2 inhibition increases blood pressure in rats. Br. J. Pharmacol. 2002, 136, 1117–1126. [Google Scholar] [CrossRef] [Green Version]
- Arashi, H.; Ogawa, H.; Yamaguchi, J.; Kawada-Watanabe, E.; Hagiwara, N. Impact of visit-to-visit variability and systolic blood pressure control on subsequent outcomes in hypertensive patients with coronary artery disease (from the HIJ-CREATE substudy). Am. J. Cardiol. 2015, 116, 236–242. [Google Scholar] [CrossRef] [Green Version]
- Melgarejo, J.D.; Thijs, L.; Wei, D.M.; Bursztyn, M.; Yang, W.Y.; Li, Y.; Asayama, K.; Hansen, T.W.; Kikuya, M.; Ohkubo, T.; et al. Relative and Absolute Risk to Guide the Management of Pulse Pressure, an Age-Related Cardiovascular Risk Factor. Am. J. Hypertens. 2021. [Google Scholar] [CrossRef]
- Fyhrquist, F.; Metsarinne, K.; Tikkanen, I. Role of angiotensin II in blood pressure regulation and in the pathophysiology of cardiovascular disorders. J. Hum. Hypertens. 1995, 9 (Suppl. 5), S19–S24. [Google Scholar]
- Yang, N.; Hong, N.J.; Garvin, J.L. Dietary fructose enhances angiotensin II-stimulated Na(+) transport via activation of PKC-alpha in renal proximal tubules. Am. J. Physiol. Ren. Physiol. 2020, 318, F1513–F1519. [Google Scholar] [CrossRef]
- Gonzalez-Vicente, A.; Cabral, P.D.; Hong, N.J.; Asirwatham, J.; Saez, F.; Garvin, J.L. Fructose reabsorption by rat proximal tubules: Role of Na(+)-linked cotransporters and the effect of dietary fructose. Am. J. Physiol. Ren. Physiol. 2019, 316, F473–F480. [Google Scholar] [CrossRef]
- Gonzalez-Vicente, A.; Hong, N.J.; Yang, N.; Cabral, P.D.; Berthiaume, J.M.; Dominici, F.P.; Garvin, J.L. Dietary Fructose Increases the Sensitivity of Proximal Tubules to Angiotensin II in Rats Fed High-Salt Diets. Nutrients 2018, 10, 1244. [Google Scholar] [CrossRef] [Green Version]
- Cabral, P.D.; Hong, N.J.; Hye Khan, M.A.; Ortiz, P.A.; Beierwaltes, W.H.; Imig, J.D.; Garvin, J.L. Fructose stimulates Na/H exchange activity and sensitizes the proximal tubule to angiotensin II. Hypertension 2014, 63, e68–e73. [Google Scholar] [CrossRef] [Green Version]
- Ares, G.R.; Kassem, K.M.; Ortiz, P.A. Fructose acutely stimulates NKCC2 activity in rat thick ascending limbs by increasing surface NKCC2 expression. Am. J. Physiol. Ren. Physiol. 2019, 316, F550–F557. [Google Scholar] [CrossRef]
- Katakam, P.V.; Ujhelyi, M.R.; Hoenig, M.E.; Miller, A.W. Endothelial dysfunction precedes hypertension in diet-induced insulin resistance. Am. J. Physiol. 1998, 275, R788–R792. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.; Bhanot, S.; McNeill, J.H. Effect of chronic endothelin blockade in hyperinsulinemic hypertensive rats. Am. J. Physiol 1995, 269, H2017–H2021. [Google Scholar] [CrossRef]
- Nakagawa, T.; Hu, H.; Zharikov, S.; Tuttle, K.R.; Short, R.A.; Glushakova, O.; Ouyang, X.; Feig, D.I.; Block, E.R.; Herrera-Acosta, J.; et al. A causal role for uric acid in fructose-induced metabolic syndrome. Am. J. Physiol. Ren. Physiol. 2006, 290, F625–F631. [Google Scholar] [CrossRef] [Green Version]
- Zenner, Z.P.; Gordish, K.L.; Beierwaltes, W.H. Free radical scavenging reverses fructose-induced salt-sensitive hypertension. Integr. Blood Press Control 2018, 11, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, J.D.; Harrison, D.G.; Figueroa, C.A.; Lacolley, P.; Laurent, S. Central Artery Stiffness in Hypertension and Aging: A Problem With Cause and Consequence. Circ. Res. 2016, 118, 379–381. [Google Scholar] [CrossRef]
- Gazhonova, V.E.; Zykova, A.S.; Chistyakov, A.A.; Roshchupkina, S.V.; Romanova, M.D.; Krasnova, T.N. Prognostic value of renal resistance index in estimating the progression of chronic kidney disease. Ter. Arkh. 2015, 87, 29–33. [Google Scholar] [CrossRef]
- Radermacher, J.; Ellis, S.; Haller, H. Renal resistance index and progression of renal disease. Hypertension 2002, 39, 699–703. [Google Scholar] [CrossRef] [Green Version]
- Parolini, C.; Noce, A.; Staffolani, E.; Giarrizzo, G.F.; Costanzi, S.; Splendiani, G. Renal resistive index and long-term outcome in chronic nephropathies. Radiology 2009, 252, 888–896. [Google Scholar] [CrossRef]
- Mendonca, S.; Gupta, S. Resistive index predicts renal prognosis in chronic kidney disease. Nephrol. Dial. Transplant. 2010, 25, 644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heine, G.H.; Rogacev, K.S.; Fliser, D.; Krumme, B. Renal resistive index and cardiovascular and renal outcomes in essential hypertension. Hypertension 2013, 61, e22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doi, Y.; Iwashima, Y.; Yoshihara, F.; Kamide, K.; Hayashi, S.; Kubota, Y.; Nakamura, S.; Horio, T.; Kawano, Y. Renal resistive index and cardiovascular and renal outcomes in essential hypertension. Hypertension 2012, 60, 770–777. [Google Scholar] [CrossRef] [Green Version]
- Tublin, M.E.; Bude, R.O.; Platt, J.F. Review. The resistive index in renal Doppler sonography: Where do we stand? AJR Am. J. Roentgenol. 2003, 180, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Ikee, R.; Kobayashi, S.; Hemmi, N.; Imakiire, T.; Kikuchi, Y.; Moriya, H.; Suzuki, S.; Miura, S. Correlation between the resistive index by Doppler ultrasound and kidney function and histology. Am. J. Kidney Dis. 2005, 46, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Boddi, M.; Cecioni, I.; Poggesi, L.; Fiorentino, F.; Olianti, K.; Berardino, S.; La Cava, G.; Gensini, G. Renal resistive index early detects chronic tubulointerstitial nephropathy in normo- and hypertensive patients. Am. J. Nephrol. 2006, 26, 16–21. [Google Scholar] [CrossRef]
- Sugiura, T.; Nakamori, A.; Wada, A.; Fukuhara, Y. Evaluation of tubulointerstitial injury by Doppler ultrasonography in glomerular diseases. Clin. Nephrol. 2004, 61, 119–126. [Google Scholar] [CrossRef]
- Riemer, K.; Rowland, E.M.; Leow, C.H.; Tang, M.X.; Weinberg, P.D. Determining Haemodynamic Wall Shear Stress in the Rabbit Aorta In Vivo Using Contrast-Enhanced Ultrasound Image Velocimetry. Ann. Biomed. Eng. 2020, 48, 1728–1739. [Google Scholar] [CrossRef] [Green Version]
- Renna, N.F.; Lembo, C.; Diez, E.; Miatello, R.M. Role of Renin-Angiotensin system and oxidative stress on vascular inflammation in insulin resistence model. Int. J. Hypertens. 2013, 2013, 420979. [Google Scholar] [CrossRef]
- Boron, W.F.; Boulpaep, E.L. Medical Physiology, 3rd ed.; Elsevier: Philadelphia, PA, USA, 2017; Chapter 23; pp. 533–555. [Google Scholar]
- Tham, Y.K.; Bernardo, B.C.; Ooi, J.Y.; Weeks, K.L.; McMullen, J.R. Pathophysiology of cardiac hypertrophy and heart failure: Signaling pathways and novel therapeutic targets. Arch. Toxicol. 2015, 89, 1401–1438. [Google Scholar] [CrossRef]
- Katz, D.H.; Beussink, L.; Sauer, A.J.; Freed, B.H.; Burke, M.A.; Shah, S.J. Prevalence, clinical characteristics, and outcomes associated with eccentric versus concentric left ventricular hypertrophy in heart failure with preserved ejection fraction. Am. J. Cardiol. 2013, 112, 1158–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popovic, A.; Neskovic, N.; Marinkovic, J.; Lee, J.C.; Tan, M.; Thomas, J.D. Serial assessment of left ventricular chamber stiffness after acute myocardial infarction. Am. J. Cardiol. 1996, 77, 361–364. [Google Scholar] [CrossRef]
- Satpathy, C.; Mishra, T.K.; Satpathy, R.; Satpathy, H.K.; Barone, E. Diagnosis and management of diastolic dysfunction and heart failure. Am. Fam. Physician 2006, 73, 841–846. [Google Scholar] [PubMed]
- Huynh, K.; Bernardo, B.C.; McMullen, J.R.; Ritchie, R.H. Diabetic cardiomyopathy: Mechanisms and new treatment strategies targeting antioxidant signaling pathways. Pharmacol. Ther. 2014, 142, 375–415. [Google Scholar] [CrossRef] [PubMed]
- Schannwell, C.M.; Schneppenheim, M.; Perings, S.; Plehn, G.; Strauer, B.E. Left ventricular diastolic dysfunction as an early manifestation of diabetic cardiomyopathy. Cardiology 2002, 98, 33–39. [Google Scholar] [CrossRef]
- Zile, M.R.; Brutsaert, D.L. New concepts in diastolic dysfunction and diastolic heart failure: Part I: Diagnosis, prognosis, and measurements of diastolic function. Circulation 2002, 105, 1387–1393. [Google Scholar] [CrossRef] [Green Version]
- Abdelhaffez, A.S.; Abd El-Aziz, E.A.; Tohamy, M.B.; Ahmed, A.M. N-acetyl cysteine can blunt metabolic and cardiovascular effects via down-regulation of cardiotrophin-1 in rat model of fructose-induced metabolic syndrome. Arch. Physiol. Biochem. 2021, 1–16. [Google Scholar] [CrossRef]
- Wang, S.; Wang, E.; Chen, Q.; Yang, Y.; Xu, L.; Zhang, X.; Wu, R.; Hu, X.; Wu, Z. Uncovering Potential lncRNAs and mRNAs in the Progression From Acute Myocardial Infarction to Myocardial Fibrosis to Heart Failure. Front. Cardiovasc. Med. 2021, 8, 664044. [Google Scholar] [CrossRef]
- Galipeau, D.; Verma, S.; McNeill, J.H. Female rats are protected against fructose-induced changes in metabolism and blood pressure. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H2478–H2484. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.; Bhanot, S.; Yao, L.; McNeill, J.H. Vascular insulin resistance in fructose-hypertensive rats. Eur. J. Pharmacol. 1997, 322, R1–R2. [Google Scholar] [CrossRef]
- Galipeau, D.M.; Yao, L.; McNeill, J.H. Relationship among hyperinsulinemia, insulin resistance, and hypertension is dependent on sex. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H562–H567. [Google Scholar] [CrossRef] [Green Version]
- Song, D.; Arikawa, E.; Galipeau, D.M.; Yeh, J.N.; Battell, M.L.; Yuen, V.G.; McNeill, J.H. Chronic estrogen treatment modifies insulin-induced insulin resistance and hypertension in ovariectomized rats. Am. J. Hypertens. 2005, 18, 1189–1194. [Google Scholar] [CrossRef] [Green Version]
- Vasudevan, H.; Xiang, H.; McNeill, J.H. Differential regulation of insulin resistance and hypertension by sex hormones in fructose-fed male rats. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H1335–H1342. [Google Scholar] [CrossRef]
- Battiprolu, P.K.; Lopez-Crisosto, C.; Wang, Z.V.; Nemchenko, A.; Lavandero, S.; Hill, J.A. Diabetic cardiomyopathy and metabolic remodeling of the heart. Life Sci. 2013, 92, 609–615. [Google Scholar] [CrossRef] [Green Version]
- Lamacchia, O.; Sorrentino, M.R. Diabetes Mellitus, Arterial Stiffness and Cardiovascular Disease: Clinical Implications and the Influence of SGLT2i. Curr. Vasc. Pharmacol. 2021, 19, 233–240. [Google Scholar] [CrossRef]
- An, D.; Rodrigues, B. Role of changes in cardiac metabolism in development of diabetic cardiomyopathy. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H1489–H1506. [Google Scholar] [CrossRef] [PubMed]
- Nikolajevic Starcevic, J.; Janic, M.; Sabovic, M. Molecular Mechanisms Responsible for Diastolic Dysfunction in Diabetes Mellitus Patients. Int. J. Mol. Sci. 2019, 20, 1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Susic, D.; Varagic, J.; Ahn, J.; Frohlich, E.D. Crosslink breakers: A new approach to cardiovascular therapy. Curr. Opin. Cardiol. 2004, 19, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Nunes, S.; Soares, E.; Fernandes, J.; Viana, S.; Carvalho, E.; Pereira, F.C.; Reis, F. Early cardiac changes in a rat model of prediabetes: Brain natriuretic peptide overexpression seems to be the best marker. Cardiovasc. Diabetol. 2013, 12, 44. [Google Scholar] [CrossRef] [Green Version]
- Drazner, M.H. The progression of hypertensive heart disease. Circulation 2011, 123, 327–334. [Google Scholar] [CrossRef]
- Griffin, K.A. Hypertensive Kidney Injury and the Progression of Chronic Kidney Disease. Hypertension 2017, 70, 687–694. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dietary Regimen | n | Initial Weight (g) | Final Weight (g) | Heart Weight (g/kg) | Fasting Glucose (mg/dL) | Fasting Insulin (ng/mL) | G:I Ratio (×106) | PRA (ngAngI/mL/hr) |
|---|---|---|---|---|---|---|---|---|
| GCS-GCS | 9 | 125 ± 4 | 381 ± 9 | 3.3 ± 0.1 | 128 ± 13 | 1.14 ± 0.23 | 64.7 ± 6.4 | 1.82 ± 0.20 |
| FCS-GHS | 9 | 132 ± 4 | 347 ± 10 | 3.4 ± 0.1 | 129 ± 24 | 0.74 ± 0.12 | 74.6 ± 12.6 | 0.66 ± 0.12 * |
| FHS-GHS | 8 | 128 ± 3 | 362 ± 12 | 3.2 ± 0.1 | 126 ± 14 | 1.33 ± 0.12 | 31.6 ± 1.9 *,† | 1.35 ± 0.28 |
| FHS-FHS | 9 | 127 ± 5 | 366 ± 12 | 3.4 ± 0.1 | 118 ± 11 | 1.03 ± 0.17 | 39.1 ± 7.1 † | 1.09 ± 0.29 |
| PHASE I | |||||||
| WEEK 1 | WEEK 2 | WEEK 3 | |||||
| Dietary Regimen | n | Caloric Intake (kcal/day) | Sodium Intake (mmol/day) | Caloric Intake (kcal/day) | Sodium Intake (mmol/day) | Caloric Intake (kcal/day) | Sodium Intake (mmol/day) |
| GCS-GCS | 9 | 61.5 ± 1.7 | 3.0 ± 0.3 | 68.4 ± 1.3 | 3.0 ± 0.1 | 66.0 ± 2.1 | 2.9 ± 0.1 |
| FCS-GHS | 9 | 60.7 ± 2.6 | 2.7 ± 0.1 | 67.2 ± 1.4 | 3.0 ± 0.1 | 65.7 ± 2.0 | 3.2 ± 0.3 |
| FHS-GHS | 8 | 48.7 ± 1.6 *,† | 23.5 ± 0.8 *,† | 60.6 ± 2.2 * | 29.2 ± 1.1 *,† | 61.8 ± 2.1 | 29.3 ± 0.9 *,† |
| FHS-FHS | 9 | 51.9 ± 2.1 *,† | 24.9 ± 1.0 *,† | 63.9 ± 1.2 | 30.9 ± 0.6 *,† | 63.5 ± 1.5 | 31.0 ± 0.8 *,† |
| PHASE III | |||||||
| WEEK 1 | WEEK 2 | WEEK 3 | |||||
| Dietary Regimen | n | Caloric Intake (kcal/day) | Sodium Intake (mmol/day) | Caloric Intake (kcal/day) | Sodium Intake (mmol/day) | Caloric Intake (kcal/day) | Sodium Intake (mmol/day) |
| GCS-GCS | 9 | 62.6 ± 1.9 | 2.8 ± 0.1 | 63.5 ± 2.1 | 2.9 ± 0.1 | 64.3 ± 1.8 | 2.8 ± 0.1 |
| FCS-GHS | 9 | 45.9 ± 1.7 * | 22.1 ± 0.8 * | 54.4 ± 1.4 * | 26.2 ± 0.7 * | 57.1 ± 1.3 | 27.5 ± 0.6 * |
| FHS-GHS | 8 | 50.1 ± 2.5 * | 23.3 ± 1.2 * | 56.1 ± 2.2 | 26.0 ± 0.8 * | 59.9 ± 2.0 | 27.7 ± 0.6 * |
| FHS-FHS | 9 | 60.5 ± 1.6 †,§ | 27.5 ± 0.8 * | 63.9 ± 1.9 †,§ | 29.0 ± 0.3 * | 66.4 ± 2.6 † | 29.9 ± 1.0 * |
| (A) | ||||||
| Dietary Regimen | MAP (mmHg) | HEART RATE (BPM) | ||||
| n | Baseline | End Phase I | Baseline | End Phase I | ||
| GCS-GCS | 9 | 100.0 ± 1.0 | 108.4 ± 1.6 | 465 ± 8 | 391 ± 16 | |
| FCS-GHS | 9 | 100.7 ± 0.9 | 108.0 ± 0.9 | 458 ± 9 | 395 ± 15 | |
| FHS-GHS | 8 | 100.6 ± 1.3 | 111.1 ± 1.3 | 456 ± 11 | 381 ± 8 | |
| FHS-FHS | 9 | 99.4 ± 1.0 | 110.2 ± 1.4 | 451 ± 11 | 391 ± 11 | |
| (B) | ||||||
| Dietary Regimen | n | Δ MAP (mmHg) | Δ SBP (mmHg) | Δ DBP (mmHg) | Δ HR (BPM) | Δ PP (MMHG) |
| GCS-GCS | 7 | 10 ± 1.0 | 11 ± 2.2 | 11 ± 1.2 | −92 ± 11 | 2.4 ± 1.5 |
| FCS-GHS | 8 | 15 ± 0.9 * | 18 ± 0.9 * | 14 ± 1.3 | −103 ± 18 | 4.2 ± 1.6 |
| FHS-GHS | 8 | 15 ± 1.4 * | 18 ± 1.7 * | 13 ± 1.7 | −105 ± 6 | 4.2 ± 2.0 |
| FHS-FHS | 8 | 16 ± 2.0 ** | 19 ± 2.3 ** | 14 ± 1.9 | −89 ± 14 | 4.6 ± 1.4 |
| Groups | ||||
|---|---|---|---|---|
| GCS-GCS | FCS-GHS | FHS-GHS | FHS-FHS | |
| n | 9 | 8 | 8 | 9 |
| LVEF (%) | 74.9 ± 3.4 | 77.3 ± 4.4 | 80.3 ± 3.7 | 78.2 ± 2.3 |
| LVFS (%) | 46.0 ± 3.4 | 49.1 ± 4.4 | 51.9 ± 4.4 | 48.8 ± 2.4 |
| LVIDs (mm) | 3.7 ± 0.3 | 3.4 ± 0.4 | 3.0 ± 0.4 | 3.5 ± 0.3 |
| LVIDD (mm) | 6.7 ± 0.3 | 6.5 ± 0.3 | 6.1 ± 0.4 | 6.9 ± 0.4 |
| LVAWS (mm) | 3.2 ± 0.1 | 3.3 ± 0.2 | 3.5 ± 0.2 | 3.6 ± 0.1 |
| LVAWD (mm) | 1.9 ± 0.03 | 2.0 ± 0.1 | 2.5 ± 0.1 * | 2.2 ± 0.1 * |
| LVPWS (mm) | 3.5 ± 0.1 | 3.5 ± 0.2 | 4.0 ± 0.3 | 4.0 ± 0.2 |
| LVPWD (mm) | 2.3 ± 0.1 | 2.4 ± 0.1 | 3.0 ± 0.3 * | 2.7 ± 0.2 |
| LVTWS (mm) | 6.8 ± 0.2 | 6.8 ± 0.4 | 7.4 ± 0.3 | 7.6 ± 0.4 |
| LVTWD (mm) | 4.3 ± 0.1 | 4.4 ± 0.2 | 5.6 ± 0.4 * | 4.9 ± 0.3 |
| LV Mass (mg) | 1190 ± 73 | 1060 ± 59 | 1401 ± 56 * | 1373.0 ± 75 * |
| Group | ||||
|---|---|---|---|---|
| GCS-GCS | FCS-GHS | FHS-GHS | FHS-FHS | |
| n | 9 | 8 | 8 | 9 |
| Peak E (mm/s) | 706 ± 22 | 630 ± 45 | 652 ± 41 | 641 ± 39 |
| Peak A (mm/s) | 555 ± 27 | 550 ± 44 | 576 ± 34 | 621 ± 32 |
| E/A | 1.31 ± 0.06 | 1.16 ± 0.06 | 1.14 ± 0.06 | 1.03 ± 0.04 ** |
| DT (ms) | 39.5 ± 4.0 | 28.0 ± 2.2 * | 31.8 ± 3.0 | 25.4 ± 2.3 ** |
| IVRT (ms) | 25.5 ± 1.0 | 27.7 ± 1.5 | 27.6 ± 1.1 | 29.0 ± 0.9 |
| E’ (mm/s) | 34.1 ± 2.6 | 32.3 ± 4.6 | 34.2 ± 4.0 | 27.4 ± 1.8 |
| A’ (mm/s) | 45.3 ± 3.7 | 45.8 ± 4.7 | 48.0 ± 4.9 | 47.1 ± 4.7 |
| E/E’ | 22.5 ± 1.9 | 25.2 ± 6.2 | 21.1 ± 3.3 | 24.3 ± 2.3 |
| E’/A’ | 0.81 ± 0.01 | 0.71 ± 0.09 | 0.81 ± 0.15 | 0.64 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levanovich, P.E.; Chung, C.S.; Komnenov, D.; Rossi, N.F. Fructose plus High-Salt Diet in Early Life Results in Salt-Sensitive Cardiovascular Changes in Mature Male Sprague Dawley Rats. Nutrients 2021, 13, 3129. https://doi.org/10.3390/nu13093129
Levanovich PE, Chung CS, Komnenov D, Rossi NF. Fructose plus High-Salt Diet in Early Life Results in Salt-Sensitive Cardiovascular Changes in Mature Male Sprague Dawley Rats. Nutrients. 2021; 13(9):3129. https://doi.org/10.3390/nu13093129
Chicago/Turabian StyleLevanovich, Peter E., Charles S. Chung, Dragana Komnenov, and Noreen F. Rossi. 2021. "Fructose plus High-Salt Diet in Early Life Results in Salt-Sensitive Cardiovascular Changes in Mature Male Sprague Dawley Rats" Nutrients 13, no. 9: 3129. https://doi.org/10.3390/nu13093129
APA StyleLevanovich, P. E., Chung, C. S., Komnenov, D., & Rossi, N. F. (2021). Fructose plus High-Salt Diet in Early Life Results in Salt-Sensitive Cardiovascular Changes in Mature Male Sprague Dawley Rats. Nutrients, 13(9), 3129. https://doi.org/10.3390/nu13093129