
Lentils and Yeast Fibers: A New Strategy to Mitigate Enterotoxigenic Escherichia coli (ETEC) Strain H10407 Virulence?

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation and Characterization of Dietary-Fiber-Containing Products
2.2. ETEC Strain and Growth Conditions
2.3. Growth Kinetics Assays in Broth Media
2.4. LT Toxin Measurement in Broth Media
2.5. Mucin Bead Adhesion Assays
2.6. Caco-2 and HT29-MTX Cell Culture Assays
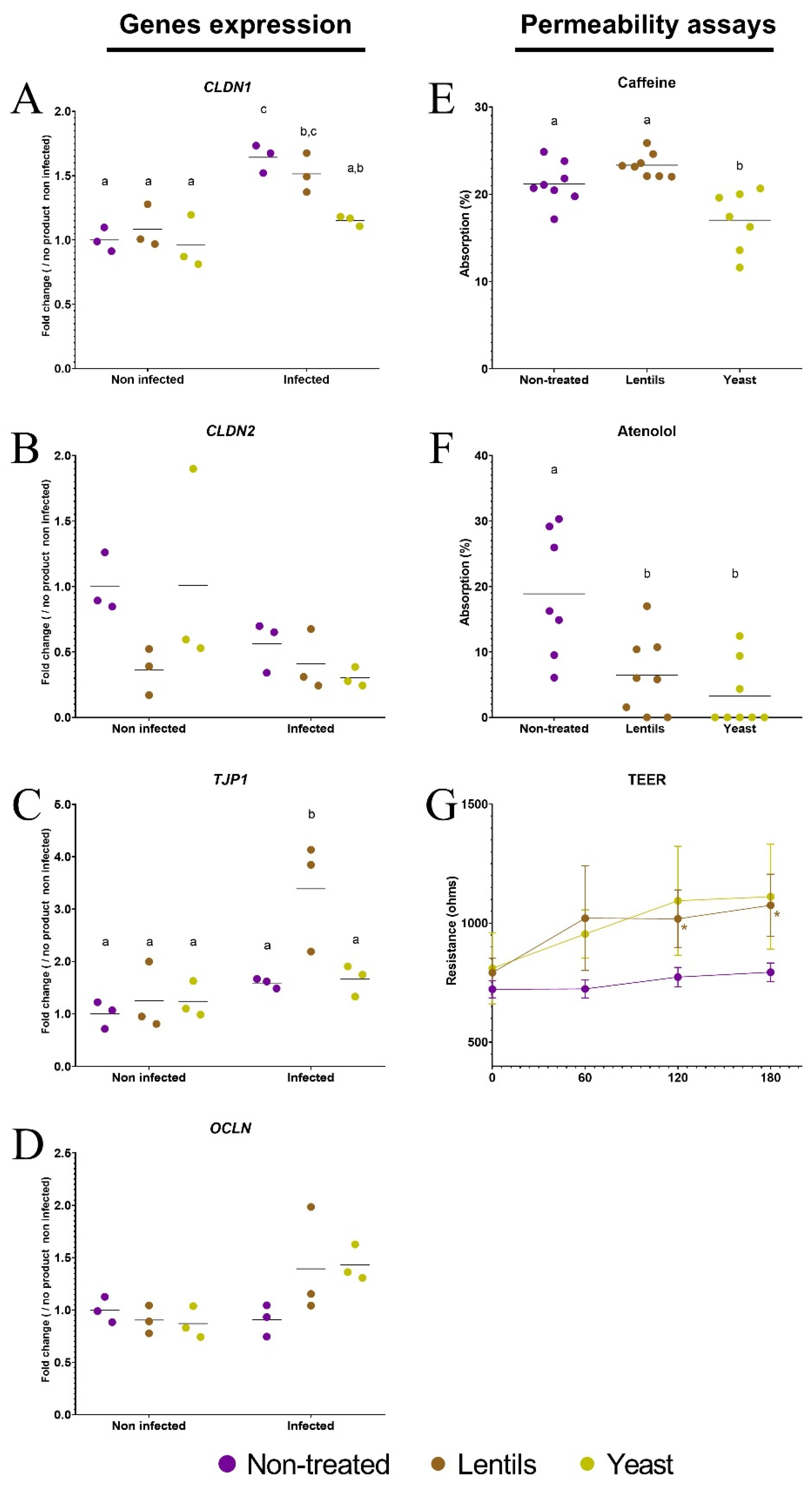
2.7. Measurement of Caco-2/HT29-MTX Permeability on Transwells
2.8. RNA Extractions
2.9. Quantitative Reverse Transcription (RT-qPCR) Analysis of ETEC Virulence Genes
2.10. Quantitative Reverse Transcription (RT-qPCR) Analysis of Selected Intestinal Cell Genes
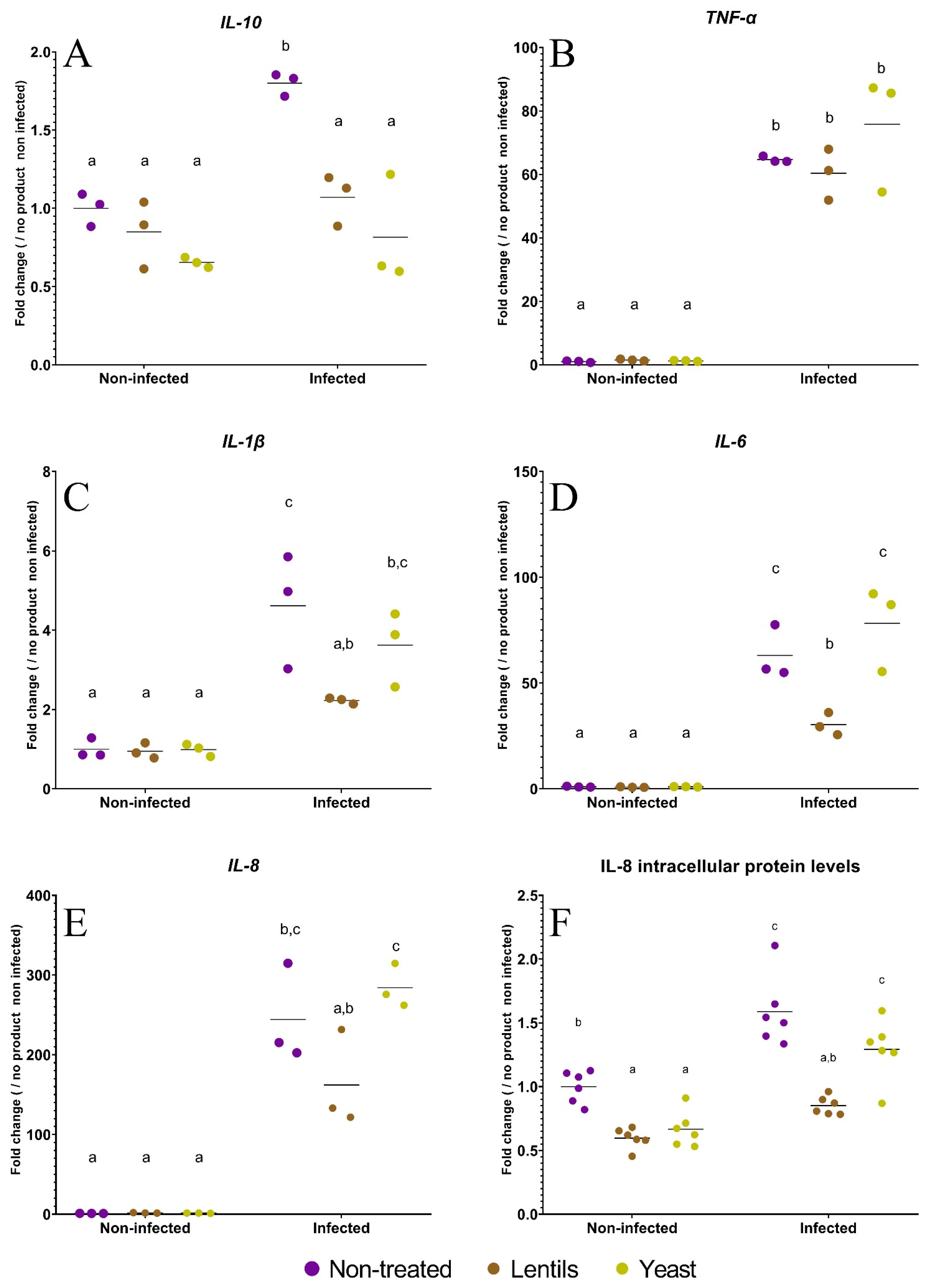
2.11. Measurement of Interleukin-8 by ELISA
2.12. Batch Experiments
2.13. Gut Microbiota Metabolite Analysis
2.14. DNA Extraction
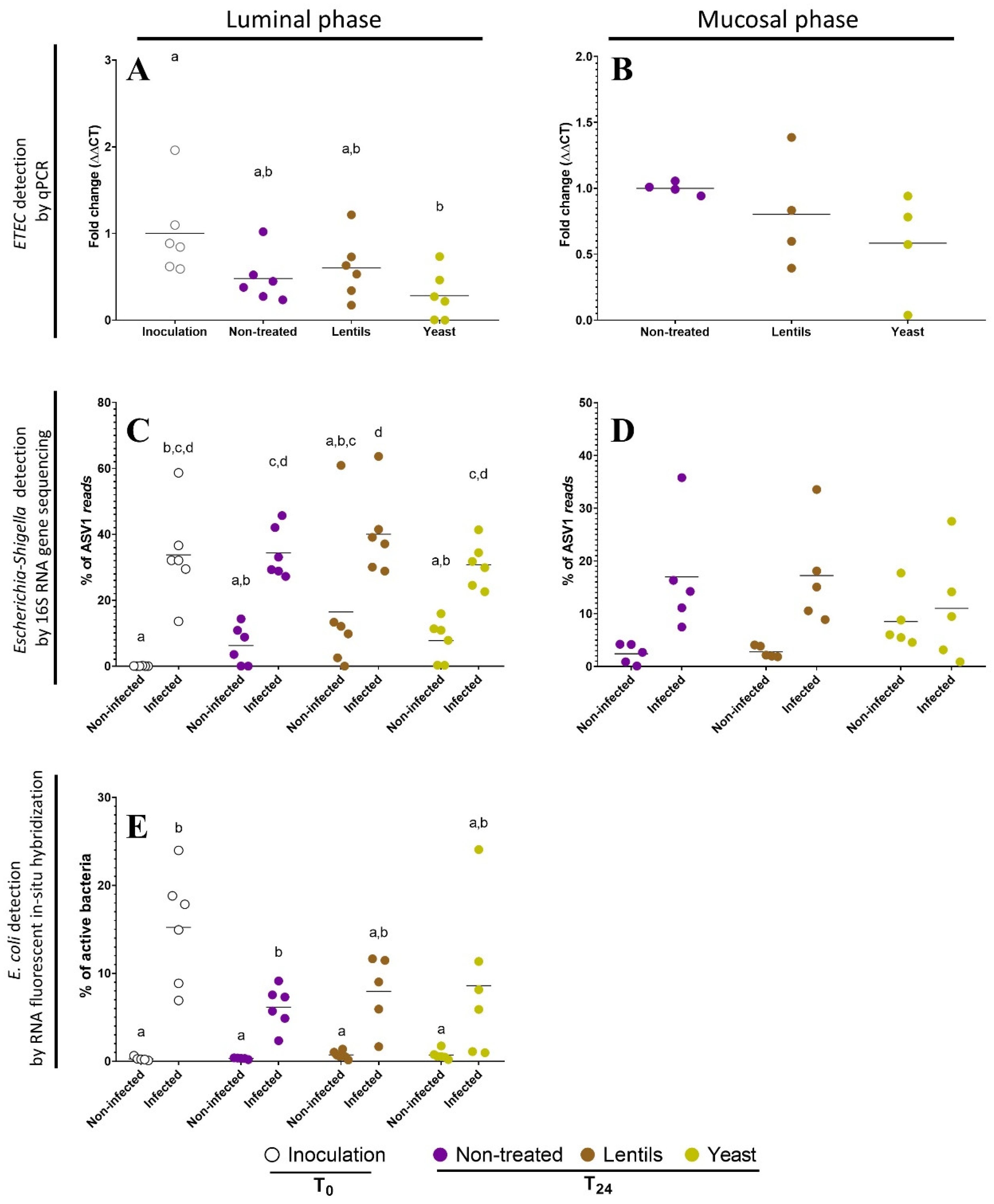
2.15. ETEC Quantification by qPCR
2.16. ETEC Quantification by RNA Fluorescent In Situ Hybridization
2.17. 16S Metabarcoding Analysis of Gut Microbial Communities
2.18. Statistical Analysis
3. Results
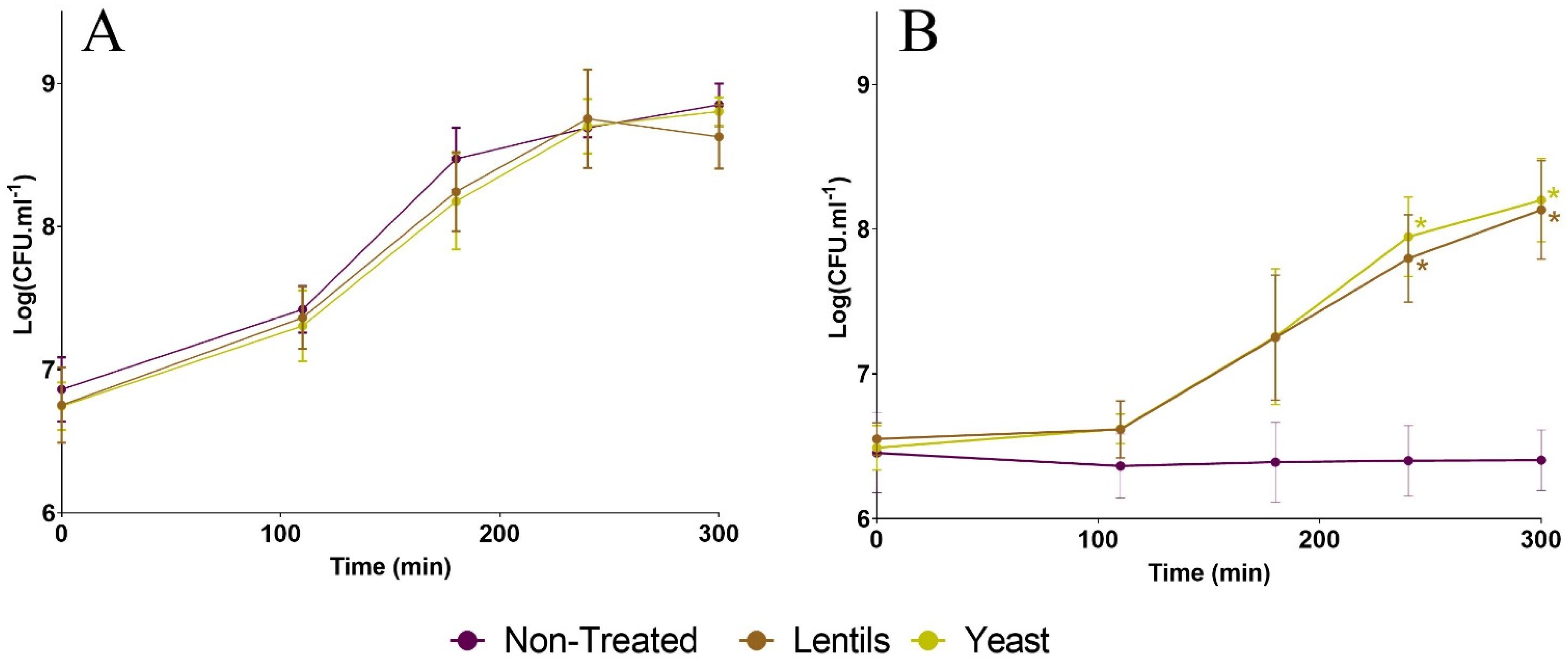
3.1. Fiber-Containing Products Do Not Impede ETEC Growth in Complete Culture Medium
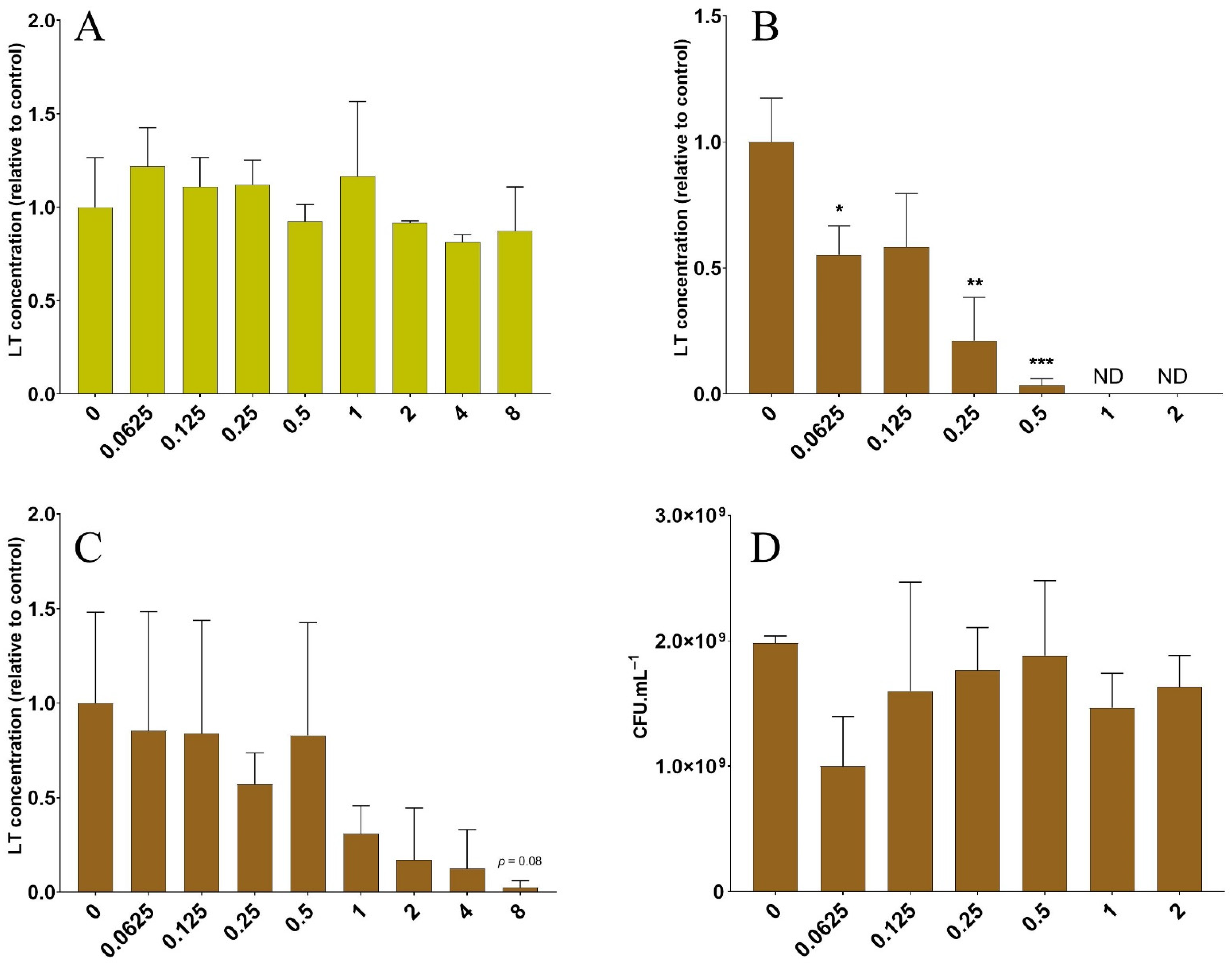
3.2. The Lentil Extract Decreases LT Toxin Concentrations in a Dose-Dependent Manner
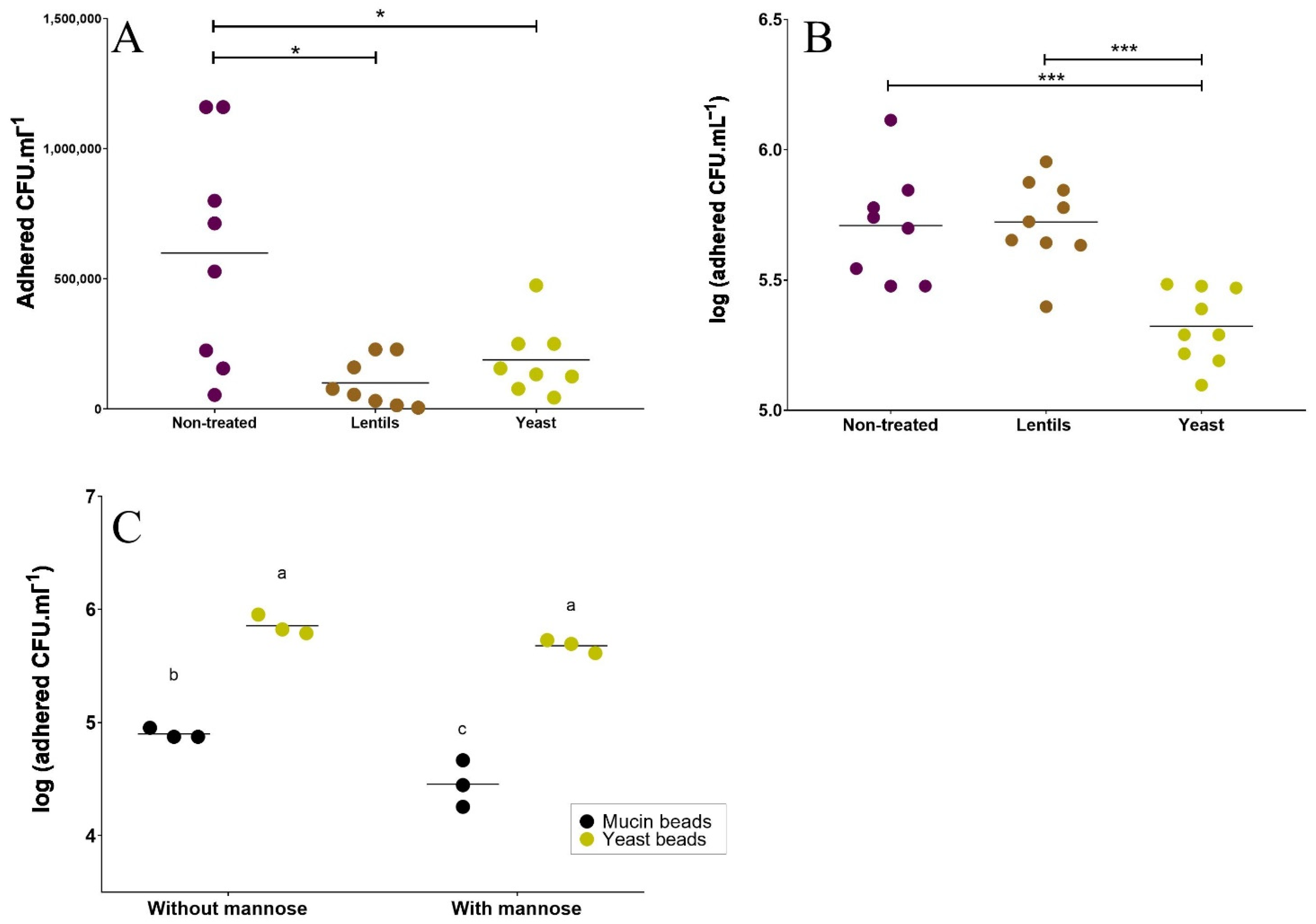
3.3. Yeast Cell Walls Inhibit ETEC Adhesion to Mucin and Mucus-Secreting Intestinal Cells
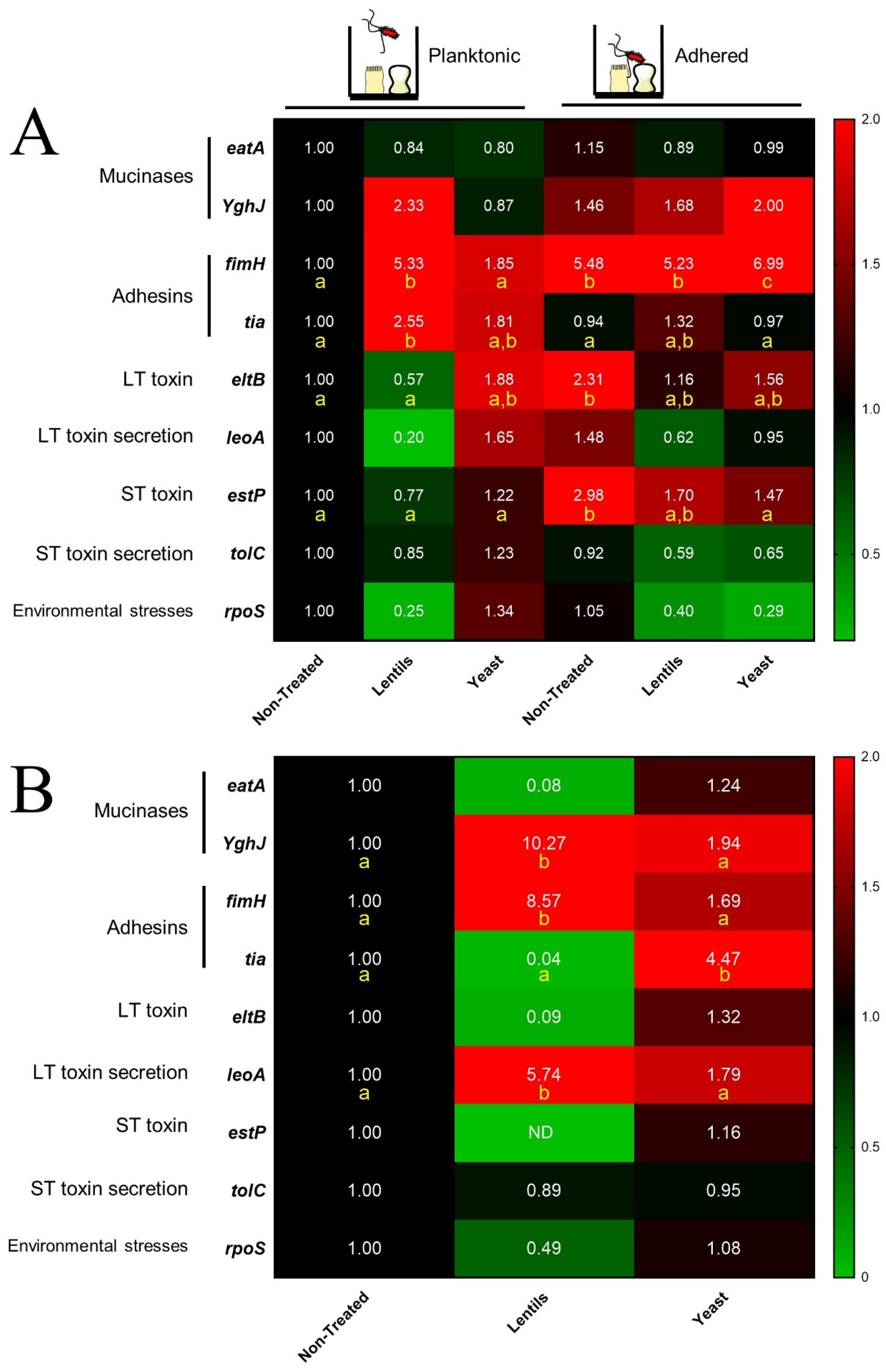
3.4. Both Fiber-Containing Products Modulate ETEC Toxin-Related Virulence Gene Expression
3.5. The lentil Extract Limits ETEC-Induced Inflammation
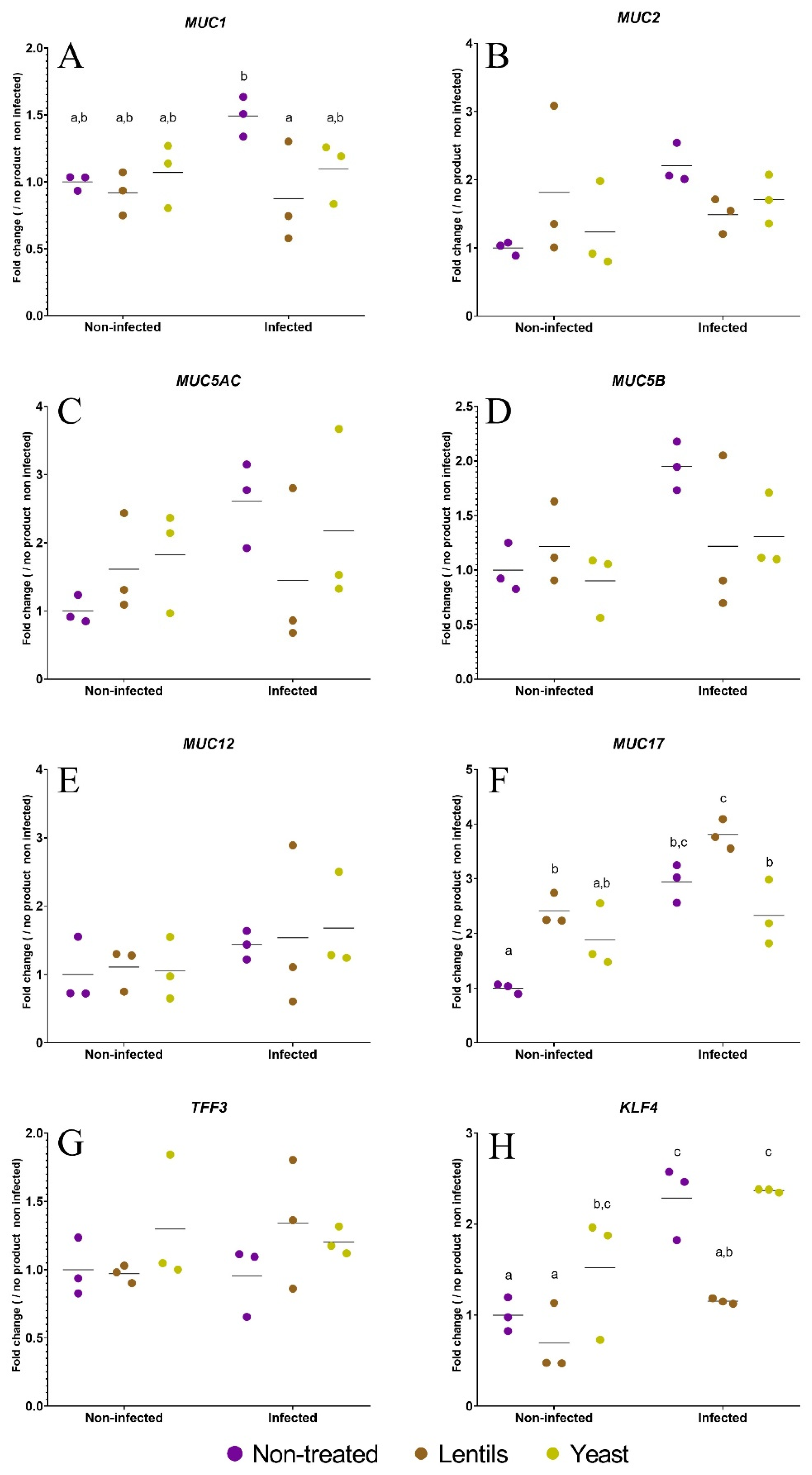
3.6. The Lentil Extract Modulates ETEC Induction of Mucus-Related Gene Expression
3.7. Yeast Cell Walls Strengthen Intestinal Barrier Function
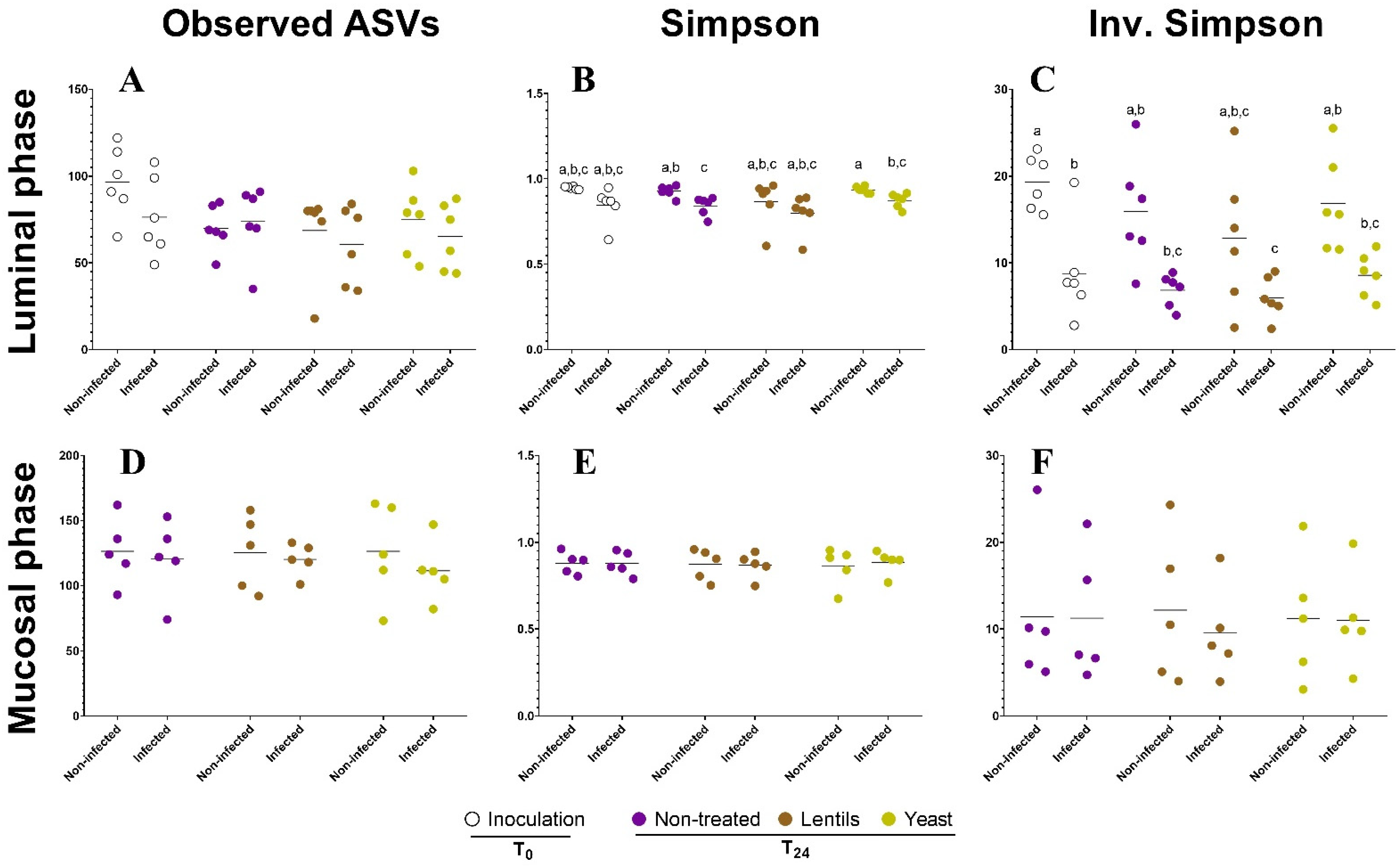
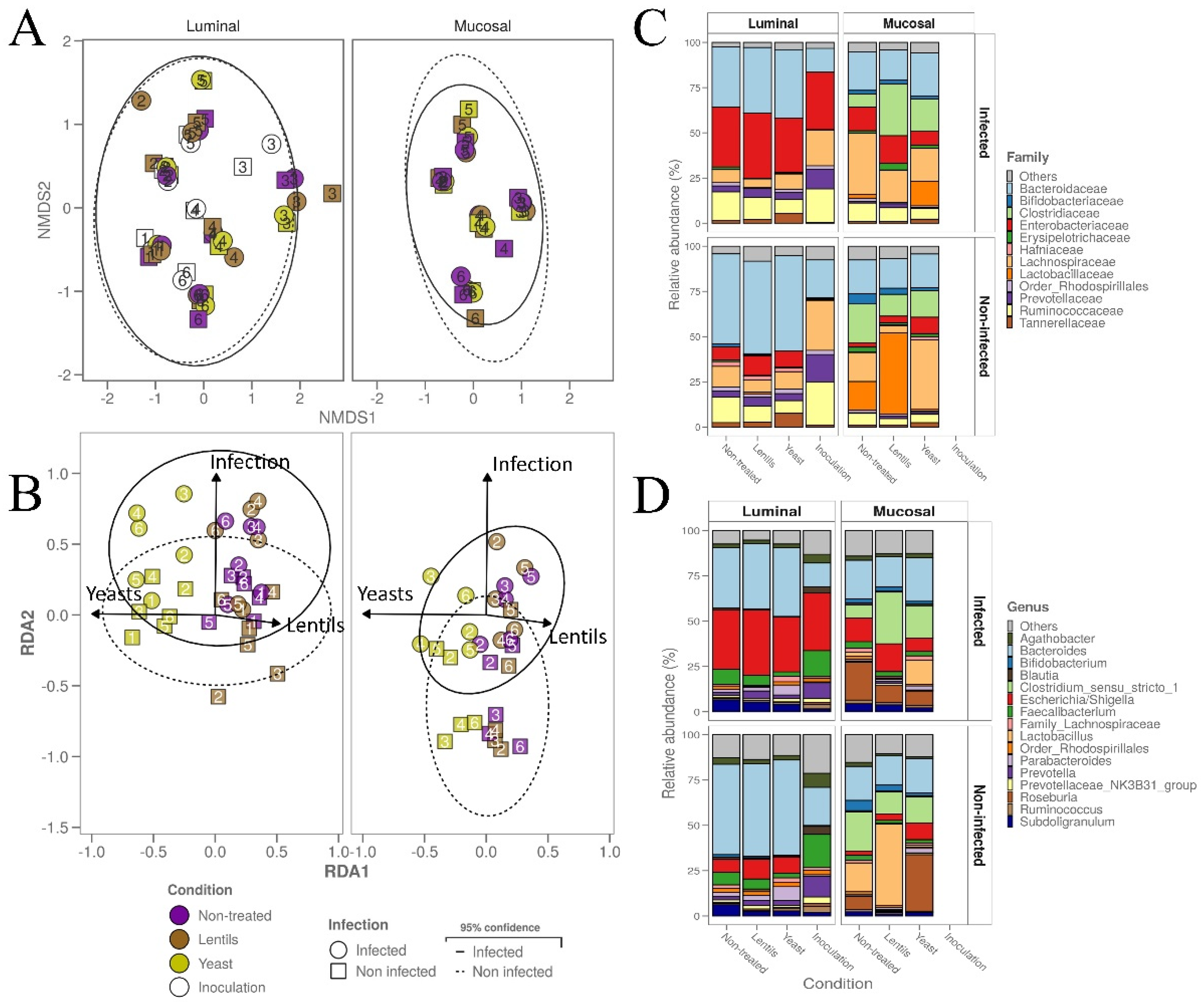
3.8. Yeast Cell Walls Mostly Impact Mucus-Associated Microbiota during ETEC Infection
3.9. Fiber Products Have No Significant Effect on ETEC Colonization in a Complex Microbial Background
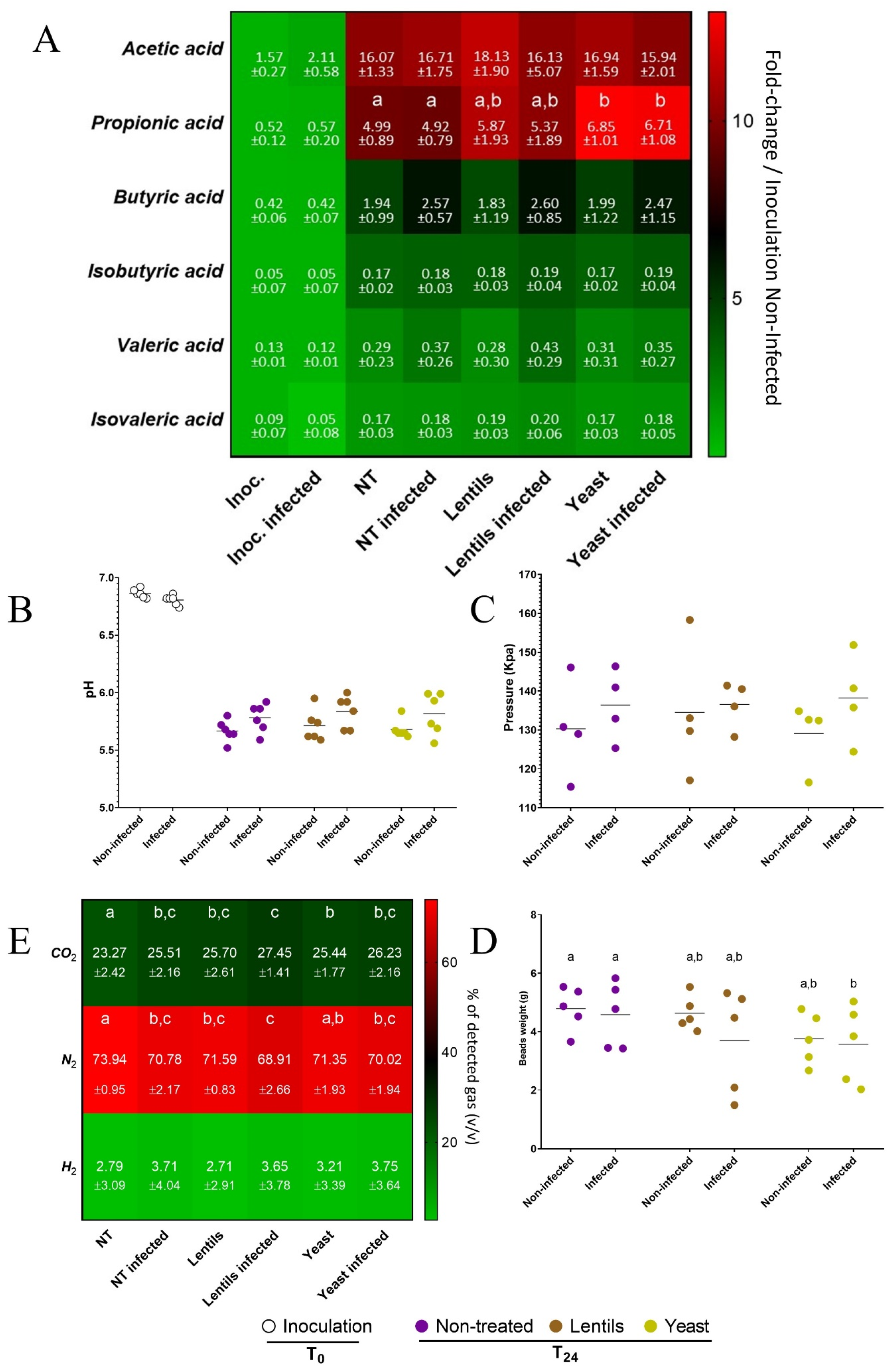
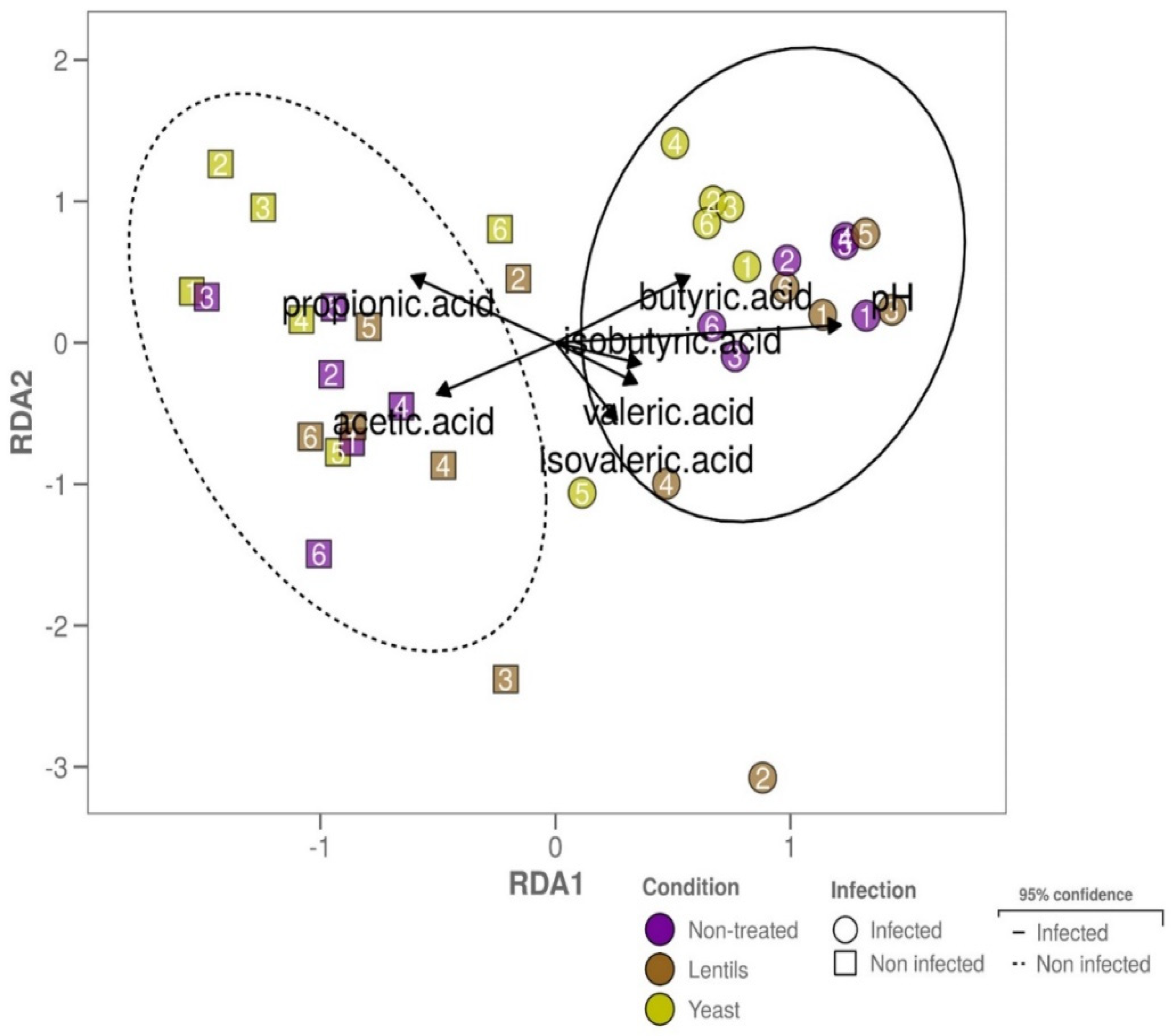
3.10. Fiber-Containing Products Slightly Affect Gut Microbial Activities during ETEC Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khalil, I.A.; Troeger, C.; Blacker, B.F.; Rao, P.C.; Brown, A.; Atherly, D.E.; Brewer, T.G.; Engmann, C.M.; Houpt, E.R.; Kang, G.; et al. Morbidity and mortality due to shigella and enterotoxigenic Escherichia coli diarrhoea: The Global Burden of Disease Study 1990–2016. Lancet Infect. Dis. 2018, 18, 1229–1240. [Google Scholar] [CrossRef] [Green Version]
- Stintzing, G.; Möllby, R. Colonization of the Upper Jejunum by Enteropathogenic and Enterotoxigenic Escherichia coli in Paediatric Diarrhoea. Acta Paediatr. 1982, 71, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.P.; Randolph, M.M.; Fleckenstein, J.M. Importance of Heat-Labile Enterotoxin in Colonization of the Adult Mouse Small Intestine by Human Enterotoxigenic Escherichia coli Strains. Infect. Immun. 2006, 74, 869–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodea, G.E.; Montiel-Infante, F.X.; Cruz-Córdova, A.; Saldaña-Ahuactzi, Z.; Ochoa, S.A.; Espinosa-Mazariego, K.; Hernández-Castro, R.; Xicohtencatl-Cortes, J. Tracking Bioluminescent ETEC during In vivo BALB/c Mouse Colonization. Front. Cell. Infect. Microbiol. 2017, 7, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vipin Madhavan, T.P.; Sakellaris, H. Colonization Factors of Enterotoxigenic Escherichia Coli. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2015; Volume 90, pp. 155–197. ISBN 978-0-12-802275-7. [Google Scholar]
- Mirhosseini, A.; Amani, J.; Nazarian, S. Review on pathogenicity mechanism of enterotoxigenic Escherichia coli and vaccines against it. Microb. Pathog. 2018, 117, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Luo, Q.; Vickers, T.J.; Sheikh, A.; Lewis, W.G.; Fleckenstein, J. EatA, an Immunogenic Protective Antigen of Enterotoxigenic Escherichia coli, Degrades Intestinal Mucin. Infect. Immun. 2014, 82, 500–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapader, R.; Bose, D.; Pal, A. YghJ, the secreted metalloprotease of pathogenic E. coli induces hemorrhagic fluid accumulation in mouse ileal loop. Microb. Pathog. 2017, 105, 96–99. [Google Scholar] [CrossRef]
- Turner, S.M.; Scott-Tucker, A.; Cooper, L.M.; Henderson, I.R. Weapons of mass destruction: Virulence factors of the global killer Enterotoxigenic Escherichia coli. FEMS Microbiol. Lett. 2006, 263, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Qadri, F.; Saha, A.; Ahmed, T.; Al Tarique, A.; Begum, Y.A.; Svennerholm, A.-M. Disease Burden Due to Enterotoxigenic Escherichia coli in the First 2 Years of Life in an Urban Community in Bangladesh. Infect. Immun. 2007, 75, 3961–3968. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, D.E.; Jiang, Z.; Steffen, R.; Verenker, M.P.; Dupont, H.L. Markers of Inflammation in Bacterial Diarrhea among Travelers, with a Focus on Enteroaggregative Escherichia coliPathogenicity. J. Infect. Dis. 2002, 185, 944–949. [Google Scholar] [CrossRef] [Green Version]
- Brubaker, J.; Zhang, X.; Bourgeois, A.L.; Harro, C.; Sack, D.A.; Chakraborty, S. Intestinal and systemic inflammation induced by symptomatic and asymptomatic enterotoxigenic E. coli infection and impact on intestinal colonization and ETEC specific immune responses in an experimental human challenge model. Gut Microbes 2021, 13, 1891852. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Weil, A.; Ryan, E.T.; Calderwood, S.B.; Harris, J.B.; Chowdhury, F.; Begum, Y.; Qadri, F.; LaRocque, R.C.; Turnbaugh, P.J. Gut Microbial Succession Follows Acute Secretory Diarrhea in Humans. mBio 2015, 6, e00381-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pop, M.; Paulson, J.N.; Chakraborty, S.; Astrovskaya, I.; Lindsay, B.; Li, S.; Bravo, H.C.; Harro, C.; Parkhill, J.; Walker, A.; et al. Individual-specific changes in the human gut microbiota after challenge with enterotoxigenic Escherichia coli and subsequent ciprofloxacin treatment. BMC Genom. 2016, 17, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, D.N.; Hamer, D.H.; Shlim, D.R. Medications for the prevention and treatment of travellers’ diarrhea. J. Travel Med. 2017, 24, S17–S22. [Google Scholar] [CrossRef]
- Piya, D.; Lessor, L.; Liu, M.; Gill, J.J. Complete Genome Sequence of Enterotoxigenic Escherichia coli Siphophage LL5. Microbiol. Resour. Announc. 2019, 8, e00674-19. [Google Scholar] [CrossRef] [Green Version]
- Roussel, C.; Sivignon, A.; de Vallée, A.; Garrait, G.; Denis, S.; Tsilia, V.; Ballet, N.; Vandekerckove, P.; Van de Wiele, T.; Barnich, N.; et al. Anti-infectious properties of the probiotic Saccharomyces cerevisiae CNCM I-3856 on enterotoxigenic E. coli (ETEC) strain H10407. Appl. Microbiol. Biotechnol. 2018, 102, 6175–6189. [Google Scholar] [CrossRef]
- Roberts, C.L.; Keita, V.; Parsons, B.N.; Prorok-Hamon, M.; Knight, P.; Winstanley, C.; O’kennedy, N.; Söderholm, J.D.; Rhodes, J.M.; Campbell, B.J. Soluble plantain fibre blocks adhesion and M-cell translocation of intestinal pathogens. J. Nutr. Biochem. 2013, 24, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Sauvaitre, T.; Durif, C.; Sivignon, A.; Chalancon, S.; Van de Wiele, T.; Etienne-Mesmin, L.; Blanquet-Diot, S. In Vitro Evaluation of Dietary Fiber Anti-Infectious Properties against Food-Borne Enterotoxigenic Escherichia Coli. Nutr. 2021, 13, 3188. [Google Scholar] [CrossRef]
- Jones, J.M. CODEX-aligned dietary fiber definitions help to bridge the ‘fiber gap’. Nutr. J. 2014, 13, 34. [Google Scholar] [CrossRef] [Green Version]
- Porter, N.; Martens, E.C. The Critical Roles of Polysaccharides in Gut Microbial Ecology and Physiology. Annu. Rev. Microbiol. 2017, 71, 349–369. [Google Scholar] [CrossRef]
- Deehan, E.C.; Duar, R.M.; Armet, A.M.; Perez-Muñoz, M.E.; Jin, M.; Walter, J. Modulation of the Gastrointestinal Microbiome with Nondigestible Fermentable Carbohydrates to Improve Human Health. Microbiol. Spectr. 2018, 5. [Google Scholar] [CrossRef]
- De Paepe, K.; Verspreet, J.; Rezaei, M.N.; Martinez, S.H.; Meysman, F.; Van De Walle, D.; Dewettinck, K.; Raes, J.; Courtin, C.; Van De Wiele, T. Isolation of wheat bran-colonizing and metabolizing species from the human fecal microbiota. PeerJ 2019, 7, e6293. [Google Scholar] [CrossRef] [PubMed]
- De Paepe, K.; Verspreet, J.; Courtin, C.M.; Van De Wiele, T. Microbial succession during wheat bran fermentation and colonisation by human faecal microbiota as a result of niche diversification. ISME J. 2020, 14, 584–596. [Google Scholar] [CrossRef]
- Hooper, L.V.; Littman, D.R.; MacPherson, A.J. Interactions Between the Microbiota and the Immune System. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef] [Green Version]
- Makki, K.; Deehan, E.C.; Walter, J.; Bäckhed, F. The Impact of Dietary Fiber on Gut Microbiota in Host Health and Disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chantarasataporn, P.; Tepkasikul, P.; Kingcha, Y.; Yoksan, R.; Pichyangkura, R.; Visessanguan, W.; Chirachanchai, S. Water-based oligochitosan and nanowhisker chitosan as potential food preservatives for shelf-life extension of minced pork. Food Chem. 2014, 159, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Kim, D.; Adesogan, A.T.; Ko, S.; Galvao, K.N.A.; Jeong, K.C. Chitosan Microparticles Exert Broad-Spectrum Antimicrobial Activity against Antibiotic-Resistant Micro-organisms without Increasing Resistance. ACS Appl. Mater. Interfaces 2016, 8, 10700–10709. [Google Scholar] [CrossRef]
- Vardaka, V.D.; Yehia, H.M.; Savvaidis, I.N. Effects of Citrox and chitosan on the survival of Escherichia coli O157: H7 and Salmonella enterica in vacuum-packaged turkey meat. Food Microbiol. 2016, 58, 128–134. [Google Scholar] [CrossRef]
- Garrido-Maestu, A.; Ma, Z.; Paik, S.-Y.; Chen, N.; Ko, S.; Tong, Z.; Jeong, K.C. Engineering of chitosan-derived nanoparticles to enhance antimicrobial activity against foodborne pathogen Escherichia coli O157: H7. Carbohydr. Polym. 2018, 197, 623–630. [Google Scholar] [CrossRef]
- Idota, T.; Kawakami, H.; Murakami, Y.; Sugawara, M. Inhibition of Cholera Toxin by Human Milk Fractions and Sialyllactose. Biosci. Biotechnol. Biochem. 1995, 59, 417–419. [Google Scholar] [CrossRef]
- Di, R.; Vakkalanka, M.S.; Onumpai, C.; Chau, H.K.; White, A.; Rastall, R.A.; Yam, K.; Hotchkiss, A. Pectic oligosaccharide structure-function relationships: Prebiotics, inhibitors of Escherichia coli O157: H7 adhesion and reduction of Shiga toxin cytotoxicity in HT29 cells. Food Chem. 2017, 227, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhang, Z.; Qiu, L.; Zhang, F.; Xu, X.; Wei, H.; Tao, X. Characterization and bioactivities of the exopolysaccharide from a probiotic strain of Lactobacillus plantarum WLPL04. J. Dairy Sci. 2017, 100, 6895–6905. [Google Scholar] [CrossRef] [PubMed]
- Leong, A.; Liu, Z.; Almshawit, H.; Zisu, B.; Pillidge, C.; Rochfort, S.; Gill, H. Oligosaccharides in goats’ milk-based infant formula and their prebiotic and anti-infection properties. Br. J. Nutr. 2019, 122, 441–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fooks, L.J.; Gibson, G.R. Mixed culture fermentation studies on the effects of synbiotics on the human intestinal pathogens Campylobacter jejuni and Escherichia coli. Anaerobe 2003, 9, 231–242. [Google Scholar] [CrossRef]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Terrapon, N.; Muller, A.; et al. A Dietary Fiber-Deprived Gut Microbiota Degrades the Colonic Mucus Barrier and Enhances Pathogen Susceptibility. Cell 2016, 167, 1339–1353.e21. [Google Scholar] [CrossRef] [Green Version]
- Martens, E.C.; Neumann, M.; Desai, M.S. Interactions of commensal and pathogenic microorganisms with the intestinal mucosal barrier. Nat. Rev. Genet. 2018, 16, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Idota, T.; Kawakami, H. Inhibitory Effects of Milk Gangliosides on the Adhesion of Escherichia colito Human Intestinal Carcinoma Cells. Biosci. Biotechnol. Biochem. 1995, 59, 69–72. [Google Scholar] [CrossRef] [Green Version]
- Salcedo, J.; Barbera, R.; Matencio, E.; Alegría, A.; Lagarda, M. Gangliosides and sialic acid effects upon newborn pathogenic bacteria adhesion: An in vitro study. Food Chem. 2013, 136, 726–734. [Google Scholar] [CrossRef]
- Evans, D.J.; DuPont, H.L.; Orskov, F.; Orskov, I. Patterns of loss of enterotoxigenicity by Escherichia coli isolated from adults with diarrhea: Suggestive evidence for an interrelationship with serotype. Infect. Immun. 1977, 17, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Deschamps, C.; Fournier, E.; Uriot, O.; Lajoie, F.; Verdier, C.; Comtet-Marre, S.; Thomas, M.; Kapel, N.; Cherbuy, C.; Alric, M.; et al. Comparative methods for fecal sample storage to preserve gut microbial structure and function in an in vitro model of the human colon. Appl. Microbiol. Biotechnol. 2020, 104, 10233–10247. [Google Scholar] [CrossRef]
- Gillois, K.; Stoffels, C.; Leveque, M.; Fourquaux, I.; Blesson, J.; Mils, V.; Cambier, S.; Vignard, J.; Terrisse, H.; Mirey, G.; et al. Repeated exposure of Caco-2 versus Caco-2/HT29-MTX intestinal cell models to (nano) silver in vitro: Comparison of two commercially available colloidal silver products. Sci. Total Environ. 2021, 754, 142324. [Google Scholar] [CrossRef] [PubMed]
- Roussel, C.; De Paepe, K.; Galia, W.; De Bodt, J.; Chalancon, S.; Leriche, F.; Ballet, N.; Denis, S.; Alric, M.; Van De Wiele, T.; et al. Spatial and temporal modulation of enterotoxigenic E. coli H10407 pathogenesis and interplay with microbiota in human gut models. BMC Biol. 2020, 18, 141. [Google Scholar] [CrossRef] [PubMed]
- Lothigius, A.; Janzon, A.; Begum, Y.; Sjöling, A.; Qadri, F.; Svennerholm, A.-M.; Bölin, I. Enterotoxigenic Escherichia coli is detectable in water samples from an endemic area by real-time PCR. J. Appl. Microbiol. 2008, 104, 1128–1136. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Delmas, J.; Gibold, L.; Faïs, T.; Batista, S.; Leremboure, M.; Sinel, C.; Vazeille, E.; Cattoir, V.; Buisson, A.; Barnich, N.; et al. Metabolic adaptation of adherent-invasive Escherichia coli to exposure to bile salts. Sci. Rep. 2019, 9, 2175. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Zhou, L.; Lim, Q.; Zou, R.; Stephanopoulos, G.; Too, H.-P. Novel reference genes for quantifying transcriptional responses of Escherichia coli to protein overexpression by quantitative PCR. BMC Mol. Biol. 2011, 12, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodas, C.; Iniguez, V.; Qadri, F.; Wiklund, G.; Svennerholm, A.-M.; Sjöling, A. Development of Multiplex PCR Assays for Detection of Enterotoxigenic Escherichia coli Colonization Factors and Toxins. J. Clin. Microbiol. 2009, 47, 1218–1220. [Google Scholar] [CrossRef] [Green Version]
- Swick, M.C.; Morgan-Linnell, S.K.; Carlson, K.M.; Zechiedrich, L. Expression of Multidrug Efflux Pump Genes acrAB-tolC, mdfA, and norE in Escherichia coli Clinical Isolates as a Function of Fluoroquinolone and Multidrug Resistance. Antimicrob. Agents Chemother. 2011, 55, 921–924. [Google Scholar] [CrossRef] [Green Version]
- Nicklasson, M.; Sjöling, A.; von Mentzer, A.; Qadri, F.; Svennerholm, A.-M. Expression of colonization factor CS5 of enterotoxigenic Escherichia coli (ETEC) is enhanced in vivo and by the bile component na glycocholate hydrate. PLoS ONE 2012, 7, e35827. [Google Scholar] [CrossRef] [Green Version]
- Hojati, Z.; Molaie, R.; Gholipour, A. The FimH Gene in Uropathogenic Escherichiacoli Strains Isolated from Patient with Urinary Tract Infection. Jundishapur J. Microbiol. 2015, 8, e17520. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Hasan, M.R.; Oba, T.; Shimizu, K. Effect OfrpoS gene knockout on the metabolism of Escherichia coli during exponential growth phase and early stationary phase based on gene expressions, enzyme activities and intracellular metabolite concentrations. Biotechnol. Bioeng. 2006, 94, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Amann, R.I.; Binder, B.J.; Olson, R.J.; Chisholm, S.W.; Devereux, R.; Stahl, D.A. Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. Appl. Environ. Microbiol. 1990, 56, 1919–1925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baudart, J.; LeBaron, P. Rapid detection of Escherichia coli in waters using fluorescent in situ hybridization, direct viable counting and solid phase cytometry. J. Appl. Microbiol. 2010, 109, 1253–1264. [Google Scholar] [CrossRef] [PubMed]
- Dorier, M.; Béal, D.; Tisseyre, C.; Marie-Desvergne, C.; Dubosson, M.; Barreau, F.; Houdeau, E.; Herlin-Boime, N.; Rabilloud, T.; Carriere, M. The food additive E171 and titanium dioxide nanoparticles indirectly alter the homeostasis of human intestinal epithelial cells in vitro. Environ. Sci. Nano 2019, 6, 1549–1561. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Gao, Y.; Li, S.; Wu, C.; Wang, J.; Zheng, N. Modulation of Mucin (MUC2, MUC5AC and MUC5B) mRNA Expression and Protein Production and Secretion in Caco-2/HT29-MTX Co-Cultures Following Exposure to Individual and Combined Aflatoxin M1 and Ochratoxin A. Toxins 2019, 11, 132. [Google Scholar] [CrossRef] [Green Version]
- de Leon, M.C.P. Développement D’un Modèle in Vitro D’inflammation Intestinale Par L’utilisation de Lignées Cellulaires Humaines En Co-Culture Pour L’étude Des Interactions Avec Les Micro-Constituants Alimentaires; Médecine humaine et pathologie. Doctoral Dissertation, Université Montpellier, Montpellier, France, 2019. [Google Scholar]
- Netsch, M.I.; Gutmann, H.; Aydogan, C.; Drewe, J. Green Tea Extract Induces Interleukin-8 (IL-8) mRNA and Protein Expression but Specifically Inhibits IL-8 Secretion in Caco-2 Cells. Planta Medica 2006, 72, 697–702. [Google Scholar] [CrossRef]
- Berger, E.; Nassra, M.; Atgié, C.; Plaisancié, P.; Géloën, A. Oleic Acid Uptake Reveals the Rescued Enterocyte Phenotype of Colon Cancer Caco-2 by HT29-MTX Cells in Co-Culture Mode. Int. J. Mol. Sci. 2017, 18, 1573. [Google Scholar] [CrossRef]
- Raveschot, C.; Coutte, F.; Frémont, M.; Vaeremans, M.; Dugersuren, J.; Demberel, S.; Drider, D.; Dhulster, P.; Flahaut, C.; Cudennec, B. Probiotic Lactobacillus strains from Mongolia improve calcium transport and uptake by intestinal cells in vitro. Food Res. Int. 2020, 133, 109201. [Google Scholar] [CrossRef]
- De Paepe, K.; Kerckhof, F.-M.; Verspreet, J.; Courtin, C.; Van De Wiele, T. Inter-individual differences determine the outcome of wheat bran colonization by the human gut microbiome. Environ. Microbiol. 2017, 19, 3251–3267. [Google Scholar] [CrossRef]
- Anderson, M.J.; Ellingsen, K.E.; McArdle, B.H. Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 2006, 9, 683–693. [Google Scholar] [CrossRef]
- Miclotte, L.; de Paepe, K.; Rymenans, L.; Callewaert, C.; Raes, J.; Rajkovic, A.; Van Camp, J.; Van de Wiele, T. Dietary Emulsifiers Alter Composition and Activity of the Human Gut Microbiota in vitro, Irrespective of Chemical or Natural Emulsifier Origin. Front. Microbiol. 2020, 11, 2733. [Google Scholar] [CrossRef]
- Huang, W.E.; Stoecker, K.; Griffiths, R.; Newbold, L.; Daims, H.; Whiteley, A.S.; Wagner, M. Raman-FISH: Combining stable-isotope Raman spectroscopy and fluorescence in situ hybridization for the single cell analysis of identity and function. Environ. Microbiol. 2007, 9, 1878–1889. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMurdie, P.; Holmes, S. Waste Not, Want Not: Why Rarefying Microbiome Data Is Inadmissible. PLOS Comput. Biol. 2014, 10, e1003531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Baselga, A.; Orme, C.D.L. Betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Ramette, A.N. Multivariate Analyses in Microbial Ecology. FEMS Microbiol. Ecol. 2007, 62, 142–160. [Google Scholar] [CrossRef] [Green Version]
- Legendre, P.; Borcard, D.; Peres-Neto, P.R. Analyzing Beta Diversity: Partitioning the Spatial Variation of Community Composition Data. Ecol. Monogr. 2005, 75, 435–450. [Google Scholar] [CrossRef]
- Legendre, P.; Anderson, M.J. Distance-Based Redundancy Analysis: Testing Multispecies Responses in Multifactorial Ecological Experiments. Ecol. Monogr. 1999, 69, 1–24. [Google Scholar] [CrossRef]
- Chen, X.; Slättengren, T.; De Lange, E.C.M.; Smith, D.E.; Hammarlund-Udenaes, M. Revisiting atenolol as a low passive permeability marker. Fluids Barriers CNS 2017, 14, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smetanova, L.; Stetinova, V.; Kholova, D.; Kvetina, J.; Smetana, J.; Svoboda, Z. Caco-2 cells and Biopharmaceutics Classification System (BCS) for prediction of transepithelial transport of xenobiotics (model drug: Caffeine). Neuro Endocrinol. Lett. 2009, 30 (Suppl. 1), 101–105. [Google Scholar]
- Drakoularakou, A.; Tzortzis, G.; Rastall, R.A.; Gibson, G.R. A double-blind, placebo-controlled, randomized human study assessing the capacity of a novel galacto-oligosaccharide mixture in reducing travellers’ diarrhoea. Eur. J. Clin. Nutr. 2010, 64, 146–152. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Liu, S.; Kling, D.; Leone, S.; Lawlor, N.; Huang, Y.; Feinberg, S.B.; Hill, D.; Newburg, D.S. The human milk oligosaccharide 2’-fucosyllactose modulates CD14 expression in human enterocytes, thereby attenuating LPS-induced inflammation. Gut 2016, 65, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Dodevska, M.S.; Djordjevic, B.I.; Sobajic, S.S.; Miletic, I.D.; Djordjevic, P.B.; Dimitrijevic-Sreckovic, V.S. Characterisation of dietary fibre components in cereals and legumes used in Serbian diet. Food Chem. 2013, 141, 1624–1629. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, Q.; Wu, X.; Algharib, S.A.; Gong, F.; Hu, J.; Luo, W.; Zhou, M.; Pan, Y.; Yan, Y.; et al. Structure, preparation, modification, and bioactivities of β-glucan and mannan from yeast cell wall: A review. Int. J. Biol. Macromol. 2021, 173, 445–456. [Google Scholar] [CrossRef]
- King, D.E.; Mainous, A.; Lambourne, C.A. Trends in Dietary Fiber Intake in the United States, 1999–2008. J. Acad. Nutr. Diet. 2012, 112, 642–648. [Google Scholar] [CrossRef]
- Holscher, H.D. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef]
- Kiela, P.R.; Ghishan, F.K. Physiology of Intestinal Absorption and Secretion. Best Pr. Res. Clin. Gastroenterol. 2016, 30, 145–159. [Google Scholar] [CrossRef] [Green Version]
- Wijemanne, P.; Moxley, R.A. Glucose Significantly Enhances Enterotoxigenic Escherichia coli Adherence to Intestinal Epithelial Cells through Its Effects on Heat-Labile Enterotoxin Production. PLoS ONE 2014, 9, e113230. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Gutiérrez, I.; Martinez, A. Polysaccharide hydrolysis with engineered Escherichia coli for the production of biocommodities. J. Ind. Microbiol. Biotechnol. 2013, 40, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Patnode, M.; Beller, Z.W.; Han, N.D.; Cheng, J.; Peters, S.L.; Terrapon, N.; Henrissat, B.; Le Gall, S.; Saulnier, L.; Hayashi, D.K.; et al. Interspecies Competition Impacts Targeted Manipulation of Human Gut Bacteria by Fiber-Derived Glycans. Cell 2019, 179, 59–73.e13. [Google Scholar] [CrossRef] [PubMed]
- Al-Majali, A.; Asem, E.; Lamar, C.; Robinson, J.; Freeman, M.; Saeed, A. Studies on the mechanism of diarrhoea induced by Escherichia coli heat-stable enterotoxin (STa) in newborn calves. Veter. Res. Commun. 2000, 24, 327–338. [Google Scholar] [CrossRef]
- Al-Majali, A.M.; Ababneh, M.M.; Shorman, M.; Saeed, A.M. Interaction of Escherichia coliheat-stable enterotoxin (STa) with its putative receptor on the intestinal tract of newborn kids. FEMS Immunol. Med Microbiol. 2007, 49, 35–40. [Google Scholar] [CrossRef] [Green Version]
- Al-Majali, A.M.; Khalifeh, M.S. Distribution and characterization of the Escherichia coli heat-stable enterotoxin (STa) receptor throughout the intestinal tract of newborn camels (Camelus dromedaries). Trop. Anim. Health Prod. 2010, 42, 1311–1314. [Google Scholar] [CrossRef]
- Gonzales, L.; Ali, Z.B.; Nygren, E.; Wang, Z.; Karlsson, S.; Zhu, B.; Quiding-Järbrink, M.; Sjöling, A. Alkaline pH Is a Signal for Optimal Production and Secretion of the Heat Labile Toxin, LT in Enterotoxigenic Escherichia coli (ETEC). PLoS ONE 2013, 8, e74069. [Google Scholar] [CrossRef] [Green Version]
- Sauvaitre, T.; Etienne-Mesmin, L.; Sivignon, A.; Mosoni, P.; Courtin, C.M.; Van de Wiele, T.; Blanquet-Diot, S. Tripartite relationship between gut microbiota, intestinal mucus and dietary fibers: Towards preventive strategies against enteric infections. FEMS Microbiol. Rev. 2021, 45, fuaa052. [Google Scholar] [CrossRef]
- Takashi, K.; Fluita, I.; Kobari, K. Effects of Short Chain Fatty Acids on the Production of Heat-Labile Enterotoxin from Enterotoxigenic Escherichia coli. Jpn. J. Pharmacol. 1989, 50, 495–498. [Google Scholar] [CrossRef]
- Verhelst, R.; Schroyen, M.; Buys, N.; Niewold, T. Selection of Escherichia coliHeat-Labile Toxin (LT) Inhibitors Using Both the GM1-ELISA and the cAMP Vero Cell Assay. Foodborne Pathog. Dis. 2013, 10, 603–607. [Google Scholar] [CrossRef]
- Kansal, R.; Rasko, D.A.; Sahl, J.W.; Munson, G.P.; Roy, K.; Luo, Q.; Sheikh, A.; Kuhne, K.J.; Fleckenstein, J.M. Transcriptional Modulation of Enterotoxigenic Escherichia coli Virulence Genes in Response to Epithelial Cell Interactions. Infect. Immun. 2013, 81, 259–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, K.P.; Martin, J.C.; Chassard, C.; Clerget, M.; Potrykus, J.; Campbell, G.; Mayer, C.-D.; Young, P.; Rucklidge, G.; Ramsay, A.G.; et al. Substrate-driven gene expression in Roseburia inulinivorans: Importance of inducible enzymes in the utilization of inulin and starch. Proc. Natl. Acad. Sci. USA 2010, 108, 4672–4679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagle, B.R.; Shrestha, S.; Arsi, K.; Upadhyaya, I.; Donoghue, A.M.; Donoghue, D.J. Pectin or chitosan coating fortified with eugenol reduces Campylobacter jejunion chicken wingettes and modulates expression of critical survival genes. Poult. Sci. 2019, 98, 1461–1471. [Google Scholar] [CrossRef] [PubMed]
- Ribet, D.; Cossart, P. How bacterial pathogens colonize their hosts and invade deeper tissues. Microbes Infect. 2015, 17, 173–183. [Google Scholar] [CrossRef]
- Luo, Q.; Kumar, P.; Vickers, T.J.; Sheikh, A.; Lewis, W.G.; Rasko, D.A.; Sistrunk, J.; Fleckenstein, J. Enterotoxigenic Escherichia coli Secretes a Highly Conserved Mucin-Degrading Metalloprotease to Effectively Engage Intestinal Epithelial Cells. Infect. Immun. 2014, 82, 509–521. [Google Scholar] [CrossRef] [Green Version]
- Dorier, M.; Béal, D.; Marie-Desvergne, C.; Dubosson, M.; Barreau, F.; Houdeau, E.; Herlin-Boime, N.; Carriere, M. Continuous in vitro exposure of intestinal epithelial cells to E171 food additive causes oxidative stress, inducing oxidation of DNA bases but no endoplasmic reticulum stress. Nanotoxicology 2017, 11, 1–11. [Google Scholar] [CrossRef] [Green Version]
- García-Rodríguez, A.; Vila, L.; Cortés, C.; Hernández, A.; Marcos, R. Exploring the usefulness of the complex in vitro intestinal epithelial model Caco-2/HT29/Raji-B in nanotoxicology. Food Chem. Toxicol. 2018, 113, 162–170. [Google Scholar] [CrossRef]
- Kim, Y.; Oh, S.; Kim, S.H. Released exopolysaccharide (r-EPS) produced from probiotic bacteria reduce biofilm formation of enterohemorrhagic Escherichia coli O157: H7. Biochem. Biophys. Res. Commun. 2009, 379, 324–329. [Google Scholar] [CrossRef]
- Wang, Y.; Gänzle, M.G.; Schwab, C. Exopolysaccharide Synthesized by Lactobacillus reuteri Decreases the Ability of Enterotoxigenic Escherichia coli To Bind to Porcine Erythrocytes. Appl. Environ. Microbiol. 2010, 76, 4863–4866. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.Y.; Woodward, A.; Zijlstra, R.T.; Gänzle, M.G. Exopolysaccharides Synthesized by Lactobacillus reuteri Protect against Enterotoxigenic Escherichia coli in Piglets. Appl. Environ. Microbiol. 2014, 80, 5752–5760. [Google Scholar] [CrossRef] [Green Version]
- Bouckenooghe, A.R.; Rodrigues, S.; Steffen, R.; Dupont, H.L.; Jiang, Z.D.; Mathewson, J.J.; Adachi, J.; Verenkar, M.P. Markers of enteric inflammation in enteroaggregative Escherichia coli diarrhea in travelers. Am. J. Trop. Med. Hyg. 2000, 62, 711–713. [Google Scholar] [CrossRef] [Green Version]
- Park, K.-S.; Choi, K.-H.; Kim, Y.-S.; Hong, B.S.; Kim, O.Y.; Kim, J.H.; Yoon, C.M.; Koh, G.-Y.; Kim, Y.-K.; Gho, Y.S. Outer Membrane Vesicles Derived from Escherichia coli Induce Systemic Inflammatory Response Syndrome. PLoS ONE 2010, 5, e11334. [Google Scholar] [CrossRef] [Green Version]
- Chutkan, H.; Kuehn, M.J. Context-Dependent Activation Kinetics Elicited by Soluble versus Outer Membrane Vesicle-Associated Heat-Labile Enterotoxin. Infect. Immun. 2011, 79, 3760–3769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loos, M.; Geens, M.; Schauvliege, S.; Gasthuys, F.; van der Meulen, J.; Dubreuil, J.D.; Goddeeris, B.M.; Niewold, T.; Cox, E. Role of Heat-Stable Enterotoxins in the Induction of Early Immune Responses in Piglets after Infection with Enterotoxigenic Escherichia coli. PLoS ONE 2012, 7, e41041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Gao, X.; Hardwidge, P.R. Heat-labile enterotoxin-induced activation of NF-κB and MAPK pathways in intestinal epithelial cells impacts enterotoxigenic Escherichia coli (ETEC) adherence. Cell. Microbiol. 2012, 14, 1231–1241. [Google Scholar] [CrossRef] [Green Version]
- Tapader, R.; Bose, D.; Basu, P.; Mondal, M.; Mondal, A.; Chatterjee, N.S.; Dutta, P.; Basu, S.; Bhadra, R.K.; Pal, A. Role in proinflammatory response of YghJ, a secreted metalloprotease from neonatal septicemic Escherichia coli. Int. J. Med. Microbiol. 2016, 306, 554–565. [Google Scholar] [CrossRef] [PubMed]
- Long, K.Z.; Rosado, J.L.; Santos, J.I.; Haas, M.; Al Mamun, A.; DuPont, H.L.; Nanthakumar, N.N.; Estrada-Garcia, T. Associations between Mucosal Innate and Adaptive Immune Responses and Resolution of Diarrheal Pathogen Infections. Infect. Immun. 2010, 78, 1221–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNamara, N.; Basbaum, C. Signaling networks controlling mucin production in response to Gram-positive and Gram-negative bacteria. Glycoconj. J. 2001, 18, 715–722. [Google Scholar] [CrossRef]
- Birchenough, G.M.H.; Nyström, E.E.L.; Johansson, M.E.V.; Hansson, G.C. A sentinel goblet cell guards the colonic crypt by triggering Nlrp6-dependent Muc2 secretion. Science 2016, 352, 1535–1542. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Kim, J.J.; Denou, E.; Gallagher, A.; Thornton, D.J.; Shajib, M.S.; Xia, L.; Schertzer, J.D.; Grencis, R.K.; Philpott, D.J.; et al. New Role of Nod Proteins in Regulation of Intestinal Goblet Cell Response in the Context of Innate Host Defense in an Enteric Parasite Infection. Infect. Immun. 2016, 84, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Farré, R.; Fiorani, M.; Rahiman, S.A.; Matteoli, G. Intestinal Permeability, Inflammation and the Role of Nutrients. Nutrients 2020, 12, 1185. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, R.; Kamata, Y.; Nishikawa, Y. Effects of Escherichia coli heat-stable enterotoxin and guanylin on the barrier integrity of intestinal epithelial T84 cells. Veter. Immunol. Immunopathol. 2013, 152, 78–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukiza, C.N.; Dubreuil, J.D. Escherichia coli Heat-Stable Toxin b Impairs Intestinal Epithelial Barrier Function by Altering Tight Junction Proteins. Infect. Immun. 2013, 81, 2819–2827. [Google Scholar] [CrossRef] [Green Version]
- Nassour, H.; Dubreuil, J.D. Escherichia coli STb Enterotoxin Dislodges Claudin-1 from Epithelial Tight Junctions. PLoS ONE 2014, 9, e113273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Ji, H.; Wang, S.; Liu, H.; Zhang, W.; Zhang, D.; Wang, Y. Probiotic Lactobacillus plantarum Promotes Intestinal Barrier Function by Strengthening the Epithelium and Modulating Gut Microbiota. Front. Microbiol. 2018, 9, 1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, W.; Huang, J.; Li, X.; Zhang, Z.; Jin, M.; Wang, J.; Xu, Y.; Wang, Z. Icariin and its phosphorylated derivatives alleviate intestinal epithelial barrier disruption caused by enterotoxigenic Escherichia coli through modulate p38 MAPK in vivo and in vitro. FASEB J. 2020, 34, 1783–1801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Yan, S.; Guo, Y.; Qiao, L.; Ma, L.; Dou, X.; Zhang, B. Lactobacillus casei ATCC 393 alleviates Enterotoxigenic Escherichia coli K88-induced intestinal barrier dysfunction via TLRs/mast cells pathway. Life Sci. 2020, 244, 117281. [Google Scholar] [CrossRef] [PubMed]
- Kreisberg, R.B.; Harper, J.; Clements, J.D.; Strauman, M.C.; Nataro, J.P.; Marohn, M. Induction of Increased Permeability of Polarized Enterocyte Monolayers by Enterotoxigenic Escherichia coli Heat-Labile Enterotoxin. Am. J. Trop. Med. Hyg. 2011, 84, 451–455. [Google Scholar] [CrossRef] [Green Version]
- Che, L.; Xu, Q.; Wu, C.; Luo, Y.; Huang, X.; Zhang, B.; Auclair, E.; Kiros, T.; Fang, Z.; Lin, Y.; et al. Effects of dietary live yeast supplementation on growth performance, diarrhoea severity, intestinal permeability and immunological parameters of weaned piglets challenged with enterotoxigenic Escherichia coli K88. Br. J. Nutr. 2017, 118, 949–958. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.; Gu, M.J.; Kye, Y.-C.; Ju, Y.-J.; Hong, R.; Bin Ju, D.; Pyung, Y.J.; Han, S.H.; Park, B.-C.; Yun, C.-H. Bacteriophage EK99P-1 alleviates enterotoxigenic Escherichia coli K99-induced barrier dysfunction and inflammation. Sci. Rep. 2022, 12, 941. [Google Scholar] [CrossRef]
- Schwarz, B.T.; Wang, F.; Shen, L.; Clayburgh, D.; Su, L.; Wang, Y.; Fu, Y.-X.; Turner, J.R. LIGHT Signals Directly to Intestinal Epithelia to Cause Barrier Dysfunction via Cytoskeletal and Endocytic Mechanisms. Gastroenterology 2007, 132, 2383–2394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, C.R.; Raleigh, D.R.; Su, L.; Shen, L.; Sullivan, E.A.; Wang, Y.; Turner, J.R. Epithelial Myosin Light Chain Kinase Activation Induces Mucosal Interleukin-13 Expression to Alter Tight Junction Ion Selectivity. J. Biol. Chem. 2010, 285, 12037–12046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, L.; Weber, C.R.; Raleigh, D.R.; Yu, D.; Turner, J.R. Tight Junction Pore and Leak Pathways: A Dynamic Duo. Annu. Rev. Physiol. 2011, 73, 283–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, B.; Sheng, B.; Zhang, Z.; Pu, A.; Yin, J.; Wang, Q.; Yang, K.; Sun, L.; Yu, M.; Qiu, Y.; et al. Aryl Hydrocarbon Receptor Activation in Intestinal Obstruction Ameliorates Intestinal Barrier Dysfunction via Suppression of MLCK-MLC Phosphorylation Pathway. Shock 2016, 46, 319–328. [Google Scholar] [CrossRef]
- Li, Q.; Burrough, E.R.; Gabler, N.K.; Loving, C.L.; Sahin, O.; Gould, S.A.; Patience, J.F. A soluble and highly fermentable dietary fiber with carbohydrases improved gut barrier integrity markers and growth performance in F18 ETEC challenged pigs1. J. Anim. Sci. 2019, 97, 2139–2153. [Google Scholar] [CrossRef]
- Wan, J.; Zhang, J.; Wu, G.; Chen, D.; Yu, B.; Huang, Z.; Luo, Y.; Zheng, P.; Luo, J.; Mao, X.; et al. Amelioration of Enterotoxigenic Escherichia coli-Induced Intestinal Barrier Disruption by Low-Molecular-Weight Chitosan in Weaned Pigs is Related to Suppressed Intestinal Inflammation and Apoptosis. Int. J. Mol. Sci. 2019, 20, 3485. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Chen, D.; Yu, B.; Yin, H.; Huang, Z.; Luo, Y.; Zheng, P.; Mao, X.; Yu, J.; Luo, J.; et al. Fructooligosaccharides improve growth performance and intestinal epithelium function in weaned pigs exposed to enterotoxigenic Escherichia coli. Food Funct. 2020, 11, 9599–9612. [Google Scholar] [CrossRef]
- van Hees, H.; Maes, D.; Millet, S.; Hartog, L.D.; van Kempen, T.; Janssens, G. Fibre supplementation to pre-weaning piglet diets did not improve the resilience towards a post-weaning enterotoxigenic E. coli challenge. J. Anim. Physiol. Anim. Nutr. 2020, 105, 260–271. [Google Scholar] [CrossRef]
- Youmans, B.P.; Ajami, N.J.; Jiang, Z.-D.; Campbell, F.; Wadsworth, W.D.; Petrosino, J.F.; Dupont, H.L.; Highlander, S.K. Characterization of the human gut microbiome during travelers’ diarrhea. Gut Microbes 2015, 6, 110–119. [Google Scholar] [CrossRef]
- Moens, F.; Duysburgh, C.; Abbeele, P.V.D.; Morera, M.; Marzorati, M. Lactobacillus rhamnosus GG and Saccharomyces cerevisiae boulardii exert synergistic antipathogenic activity in vitro against enterotoxigenic Escherichia coli. Benef. Microbes 2019, 10, 923–935. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Dai, C.; Brown, K.; Rajendiran, E.; Makarenko, S.; Baker, J.; Ma, C.; Halder, S.; Montero, M.; Ionescu, V.A.; et al. Colonic microbiota alters host susceptibility to infectious colitis by modulating inflammation, redox status, and ion transporter gene expression. Am. J. Physiol. Liver Physiol. 2011, 301, G39–G49. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, E.G.D.; Frankel, G. Overview of the Effect of Citrobacter Rodentium Infection on Host Metabolism and the Microbiota. In Shiga Toxin-Producing E. coli; Schüller, S., Bielaszewska, M., Eds.; Methods in Molecular Biology; Springer US: New York, NY, USA, 2021; Volume 2291, pp. 399–418. ISBN 978-1-07-161338-2. [Google Scholar]
- Wang, K.; Liao, M.; Zhou, N.; Bao, L.; Ma, K.; Zheng, Z.; Wang, Y.; Liu, C.; Wang, W.; Wang, J.; et al. Parabacteroides distasonis Alleviates Obesity and Metabolic Dysfunctions via Production of Succinate and Secondary Bile Acids. Cell Rep. 2019, 26, 222–235.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.-R.; Lin, C.-S.; Chang, C.-J.; Lin, T.-L.; Martel, J.; Ko, Y.-F.; Ojcius, D.M.; Lu, C.-C.; Young, J.D.; Lai, H.-C. Gut commensal Parabacteroides goldsteinii plays a predominant role in the anti-obesity effects of polysaccharides isolated from Hirsutella sinensis. Gut 2019, 68, 248–262. [Google Scholar] [CrossRef] [PubMed]
- Koh, G.Y.; Kane, A.V.; Wu, X.; Crott, J.W. Parabacteroides distasonis attenuates tumorigenesis, modulates inflammatory markers and promotes intestinal barrier integrity in azoxymethane-treated A/J mice. Carcinogenesis 2020, 41, 909–917. [Google Scholar] [CrossRef]
- Tsai, C.-C.; Lin, P.-P.; Hsieh, Y.-M. Three Lactobacillus strains from healthy infant stool inhibit enterotoxigenic Escherichia coli grown in vitro. Anaerobe 2008, 14, 61–67. [Google Scholar] [CrossRef]
- Osmanağaoğlu, O.; Kiran, F.; Ataoğlu, H. Evaluation of in vitro Probiotic Potential of Pediococcus pentosaceus OZF Isolated from Human Breast Milk. Probiotics Antimicrob. Proteins 2010, 2, 162–174. [Google Scholar] [CrossRef]
- Anand, S.; Mandal, S.; Tomar, S.K. Effect of Lactobacillus rhamnosus NCDC 298 with FOS in Combination on Viability and Toxin Production of Enterotoxigenic Escherichia coli. Probiotics Antimicrob. Proteins 2017, 11, 23–29. [Google Scholar] [CrossRef]
- Kanjee, U.; Houry, W.A. Mechanisms of Acid Resistance in Escherichia coli. Annu. Rev. Microbiol. 2013, 67, 65–81. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, B.O.; Birchenough, G.M.H.; Ståhlman, M.; Arike, L.; Johansson, M.E.; Hansson, G.C.; Bäckhed, F. Bifidobacteria or Fiber Protects against Diet-Induced Microbiota-Mediated Colonic Mucus Deterioration. Cell Host Microbe 2018, 23, 27–40.e7. [Google Scholar] [CrossRef] [Green Version]
- Neumann, M.; Steimle, A.; Grant, E.T.; Wolter, M.; Parrish, A.; Willieme, S.; Brenner, D.; Martens, E.C.; Desai, M.S. Deprivation of dietary fiber in specific-pathogen-free mice promotes susceptibility to the intestinal mucosal pathogen Citrobacter rodentium. Gut Microbes 2021, 13, 1966263. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters Tested | Results (+/−Incertitude) (g·100 g−1) | Analytical Methods | |
|---|---|---|---|
| Lentils | Specific Yeast Cell Wall | ||
| Moisture at 70 °C and low pressure | 6.1 (+/−0.5) | 7.7 (+/−0.5) | Steaming |
| Ashes | 4.1 (+/−0.2) | 3.0 (+/−0.2) | Incineration |
| Protein “N*6.25” | 36.1 (+/−1.0) | 13.3 (+/−0.6) | Kjeldahl |
| Fat “B” | 0.90 (+/−0.50) | 15.9 (+/−1.3) | Soxhlet |
| Total carbohydrates | 52.7 | 60.2 | Calculation |
| Total dietary fibers | 34.8 (+/−2.0) | 57.7 (+/−2.0) | Enzymatic method |
| Carbohydrates | 17.9 | 2.5 | Calculation |
| Gene | Target | Primer Sequence 5′-3′ | Amplicon Length (pb) | References |
|---|---|---|---|---|
| Genes to monitor ETEC survival by qPCR (in fecal batches) | ||||
| eltB | LT toxin | F-GGCAGGCAAAAGAGAAATGG R-TCCTTCATCCTTTCAATGGCT | 117 | [44] |
| 16S | Reference gene | F-NNNNNNNNNTCCTACGGGNGGCWGCAG R-NNNNNNNNNNTGACTACHVGGGTATCTAAKCC | 464 | [45] |
| Genes for RT-qPCR analysis of ETEC virulence genes | ||||
| tufA | Reference gene | F-GACATGGTTGATGACGAAGA R-GCTCTGGTTCCGGAATGTA | 199 | [46] |
| ihfB | Reference gene | F-CTGCGAGGCAGCTTCCAGTT R-GCAGCAACAGCAGCCGCTTA | 419 | [47] |
| eltB | LT toxin | F-GGCAGGCAAAAGAGAAATGG R-TCCTTCATCCTTTCAATGGCT | 117 | [44] |
| leoA | LT enterotoxin output | F-AAACGGTGCATATCCTCGTC R-AAATGCTGCCACCGAAATAC | 168 | [43] |
| estP | ST toxin | F-TCTTTCCCCTCTTTTAGTCAG R-ACAGGCAGGATTACAACAAAG | 165 | [48] |
| tolC | TolC outer membrane protein (ST toxin secretion) | F-AAGCCGAAAAACGCAACCT R-CAGAGTCGGTAAGTGACCATC | 101 | [49] |
| tia | Tia Adhesin | F-ACAGGCTTTTATGTGACCGGTAA R-GACGGAAGCGCTGGTCAGT | 67 | [50] |
| imH | Minor component of Type I pili | F-GTGCCAATTCCTCTTACCGTT R-TGGAATAATCGTACCGTTGCG | 164 | [51] |
| yghJ | Mucinase | F-CCCTGTTAGCCGGTTGTGAT R-GGTATCGGTTCTGGCGTAGG | 166 | This study |
| eatA | Mucinase | F-AACGGAAGCACCGTCATTCT R-CAGAGTCAGGGAGGCGTTTT | 363 | This study |
| rpoS | Environmental stresses response | F-GCGCGGTAGAGAAGTTTGAC R-GGCTTATCCAGTTGCTCTGC | 229 | [52] |
| Genes for bacterial quantification by RNA fluorescent in situ hybridization in batch fermentation | ||||
| 16S | Eubacteria 16S rRNA | 1-GCTGCCTCCCGTAGGAGT 2-CGGCGTCGCTGCGTCAGG 3-MCGCARACTCATCCCCAAA | N/A | [53] |
| 16S | E. coli 16S rRNA | 1-GCAAAGGTATTAACTTTACTCCC (Cy5 in 5′) 2-GCAGCAACAGCAGCCGCTTA (Helper probe) | N/A | [54] |
| Genes for RT-qPCR analysis of host response | ||||
| Mucin-related genes | ||||
| MUC1 | Mucin 1 | F-AGACGTCAGCGTGAGTGATG R-CAGCTGCCCGTAGTTCTTTC | 172 | [55] |
| MUC2 | Mucin 2 | F-CAGTGTGTCTGTAACGCTGG R-AATCGTTGTGGTCACCCTTG | 160 | This study |
| MUC5AC | Mucin 5AC | F-GTTTGACGGGAAGCAATACA R-CGATGATGAAGAAGGTTGAGG | 278 | [56] |
| MUC5B | Mucin 5B | F-GTGACAACCGTGTCGTCCTG R-TGCCGTCAAAGGTGGAATAG | 171 | [56] |
| MUC12 | Mucin 12 | F-ACCTTAGCACCAGGGTTGTG R-GGAGGATGCGTCATTCATCT | 204 | [55] |
| MUC17 | Mucin 17 | F-TGCAGAACAGGACCTCAGTG R- AGGTCATCTCAGGGTTGGTG | 206 | [55] |
| TFF3 | Trefoil factor 3 | F-AGGAGTACGTGGGCCTGTCT R-AAGGTGCATTCTGCTTCCTG | 175 | [55] |
| KLF4 | Kruppel-like-factor 4 | F-CTCACCCCACCTTCTTCACC R-AAGGTTTCTCACCTGTGTGG | 202 | This study |
| Tight-junction-related genes | ||||
| CLDN1 | Claudin 1 | F-TGGAAGACGATGAGGTGCA R-AAGGCAGAGAGAAGCAGCA | 206 | [55] |
| CLDN2 | Claudin 2 | F-CATTTGTACCTGCAAGGTCTTCT R-GCCTAGGATGTAGCCCACAA | 236 | This study |
| OCLN | Occludin | F-ACTTCAGGCAGCCTCGTTAC R-CCTGATCCAGTCCTCCTCCA | 170 | [55] |
| TJP1 | Zonula occludens 1 | F-GTGCTGGCTTGGTCTGTTTG R-TCTGTACATGCTGGCCAAGG | 159 | [55] |
| Inflammation-related genes | ||||
| TNF | Tumor necrosis factor α | F-GCCCATGTTGTAGCAAACCC R-AGGAGGTTGACCTTGGTCTG | 242 | This study |
| IL6 | Interleukin 6 | F-CCAGAGCTGTGCAGATGAGTACA R-GGCATTTGTGGTTGGGTCAGG | 101 | [57] |
| IL8 | Interleukin 8 | F-TCTGCAGCTCTGTGTGAAGG R-TGAATTCTCAGCCCTCTTCAA | 252 | This study |
| IL10 | Interleukin 10 | F-GGCGCTGTCATCGATTTCTTC R-CACTCATGGCTTTGTAGATGCC | 108 | [57] |
| IL1β | Interleukin 1β | F-AGCCATGGCAGAAGTACCTG R-TGGTGGTCGGAGATTCGTAG | 171 | [58] |
| Housekeeping genes | ||||
| GADPH | Housekeeping gene | F-GGAGTCCACTGGCGTCTT R-GAGTCCTTCCACGATACCAAA | 235 | [56] |
| HPRT | Housekeeping gene | F-TTGCTGACCTGCTGGATTAC R-AGTTGAGAGATCATCTCCAC | 149 | [59] |
| PPIA | Housekeeping gene | F-TGCTGACTGTGGACAACTCG F-TGCAGCGAGAGCACAAAGAT | 136 | [60] |
| Parameters of Static In Vitro Digestion | Gastric Vessel | Duodenum–Ileum Vessels |
|---|---|---|
| pH | from 6 (T0) to 2.1 | maintained at 6.8 |
| Volume (mL) | 50 | 90 |
| Secretions | (i) 5.36 mg of pepsin (727 U.mg−1) (ii) 4.28 mg of lipase (32 U.mg−1) (iii) HCl 0.3 M (iv) NaHCO3 0.5 M if necessary | (i) 0.9 g of bile salts (27.9 mM in solution) (ii) 1.8 g of pancreatin 4 USP (iii) Trypsin 2 mg·mL−1 (15156 U/mg protein) (iv) NaHCO3 0.5 M if necessary |
| Time period in batch (min) | 30 | 60 |
| Chyme mixing | 100 rpm (magnetic stirrer) | 100 rpm (magnetic stirrer) |
| [Total microbes] | sterile | sterile |
| Oxygen level (%) | 20 | 20 |
| Temperature (°C) | 37 | 37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sauvaitre, T.; Van Herreweghen, F.; Delbaere, K.; Durif, C.; Van Landuyt, J.; Fadhlaoui, K.; Huille, S.; Chaucheyras-Durand, F.; Etienne-Mesmin, L.; Blanquet-Diot, S.; et al. Lentils and Yeast Fibers: A New Strategy to Mitigate Enterotoxigenic Escherichia coli (ETEC) Strain H10407 Virulence? Nutrients 2022, 14, 2146. https://doi.org/10.3390/nu14102146
Sauvaitre T, Van Herreweghen F, Delbaere K, Durif C, Van Landuyt J, Fadhlaoui K, Huille S, Chaucheyras-Durand F, Etienne-Mesmin L, Blanquet-Diot S, et al. Lentils and Yeast Fibers: A New Strategy to Mitigate Enterotoxigenic Escherichia coli (ETEC) Strain H10407 Virulence? Nutrients. 2022; 14(10):2146. https://doi.org/10.3390/nu14102146
Chicago/Turabian StyleSauvaitre, Thomas, Florence Van Herreweghen, Karen Delbaere, Claude Durif, Josefien Van Landuyt, Khaled Fadhlaoui, Ségolène Huille, Frédérique Chaucheyras-Durand, Lucie Etienne-Mesmin, Stéphanie Blanquet-Diot, and et al. 2022. "Lentils and Yeast Fibers: A New Strategy to Mitigate Enterotoxigenic Escherichia coli (ETEC) Strain H10407 Virulence?" Nutrients 14, no. 10: 2146. https://doi.org/10.3390/nu14102146
APA StyleSauvaitre, T., Van Herreweghen, F., Delbaere, K., Durif, C., Van Landuyt, J., Fadhlaoui, K., Huille, S., Chaucheyras-Durand, F., Etienne-Mesmin, L., Blanquet-Diot, S., & Van de Wiele, T. (2022). Lentils and Yeast Fibers: A New Strategy to Mitigate Enterotoxigenic Escherichia coli (ETEC) Strain H10407 Virulence? Nutrients, 14(10), 2146. https://doi.org/10.3390/nu14102146