
Long-Term Dietary Patterns Are Reflected in the Plasma Inflammatory Proteome of Patients with Inflammatory Bowel Disease

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Data Collection and Definitions
2.3. Proteomic Profiling
2.4. Dietary Data
2.5. Statistical Analysis
2.5.1. General Descriptive and Inferential Statistics
2.5.2. Principal Component Analysis
2.5.3. Permutational Analysis of Variance
2.5.4. Association Analyses
3. Results
3.1. Cohort Description
3.2. Habitual Dietary Intake
3.3. Data-Driven Identification of Dietary Patterns
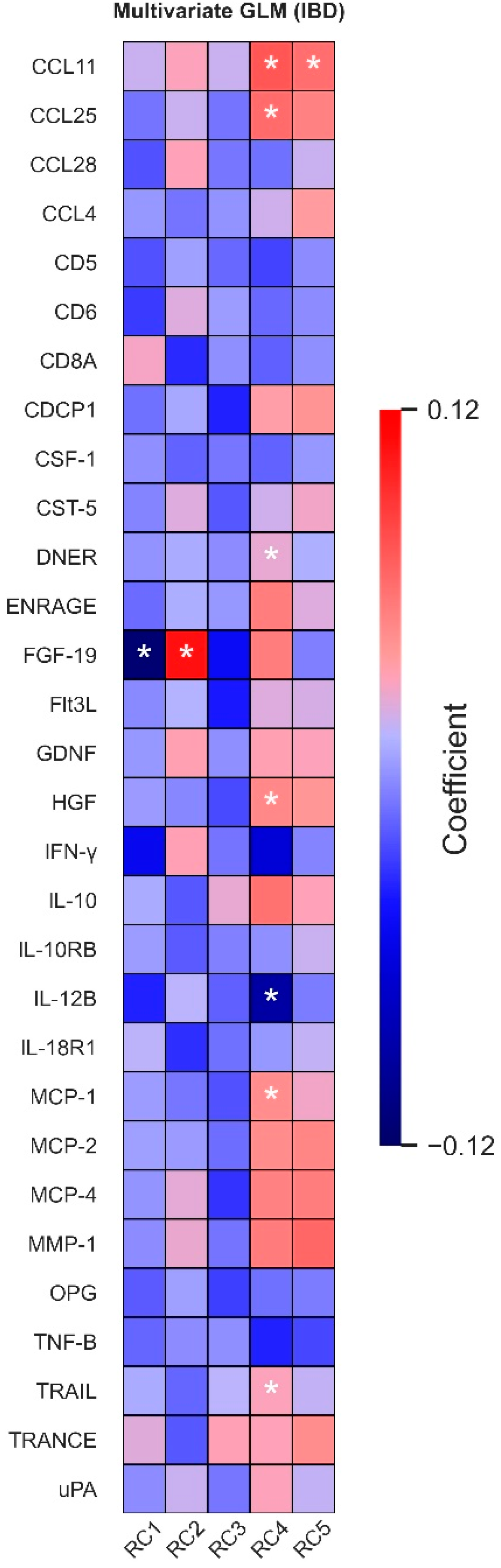
3.4. Associations between Dietary Intake Patterns and Plasma Protein Levels
3.5. Disease Activity Affects Most Dietary Pattern–Protein Associations, except for FGF-19
3.6. Protein Intake Influences Associations between Dietary Patterns and Plasma Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Colombel, J.F.; Narula, N.; Peyrin-Biroulet, L. Management Strategies to Improve Outcomes of Patients with Inflammatory Bowel Diseases. Gastroenterology 2017, 152, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Peyrin-Biroulet, L.; Eisenhut, M.; Shin, J.I. IBD immunopathogenesis: A comprehensive review of inflammatory molecules. Autoimmun. Rev. 2017, 16, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Bourgonje, A.R.; von Martels, J.Z.H.; Gabriëls, R.Y.; Blokzijl, T.; Buist-Homan, M.; Heegsma, J.; Jansen, B.H.; van Dullemen, H.M.; Festen, E.A.M.; ter Steege, R.W.F.; et al. A Combined Set of Four Serum Inflammatory Biomarkers Reliably Predicts Endoscopic Disease Activity in Inflammatory Bowel Disease. Front. Med. 2019, 6, 251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, B.B.; Maranville, J.C.; Peters, J.E.; Stacey, D.; Staley, J.R.; Blackshaw, J.; Burgess, S.; Jiang, T.; Paige, E.; Surendran, P.; et al. Genomic atlas of the human plasma proteome. Nature 2018, 558, 73–79. [Google Scholar] [CrossRef]
- Enroth, S.; Johansson, A.; Enroth, S.B.; Gyllensten, U. Strong effects of genetic and lifestyle factors on biomarker variation and use of personalized cutoffs. Nat. Commun. 2014, 5, 4684. [Google Scholar] [CrossRef] [PubMed]
- Peters, V.; Bolte, L.; Schuttert, E.; Andreu-Sánchez, S.; Dijkstra, G.; Weersma, R.; Campmans-Kuijpers, M.J.E. Western and carnivorous dietary patterns are associated with greater likelihood of IBD-development in a large prospective population-based cohort. J. Crohns Colitis 2021, jjab219. [Google Scholar] [CrossRef]
- Bolte, L.A.; Vich Vila, A.; Imhann, F.; Collij, V.; Gacesa, R.; Peters, V.; Wijmenga, C.; Kurilshikov, A.; Campmans-Kuijpers, M.J.E.; Fu, J.; et al. Long-term dietary patterns are associated with pro-inflammatory and anti-inflammatory features of the gut microbiome. Gut 2021, 70, 1287–1298. [Google Scholar] [CrossRef]
- Peters, V.; Spooren, C.E.G.M.; Pierik, M.J.; Weersma, R.K.; van Dullemen, H.M.; Festen, E.A.M.; Visschedijk, M.C.; Masclee, A.A.M.; Hendrix, E.M.B.; Almeida, R.J.; et al. Dietary Intake Pattern is Associated with Occurrence of Flares in IBD Patients. J. Crohns Colitis 2021, 15, 1305–1315. [Google Scholar] [CrossRef]
- Jowett, S.L.; Seal, C.J.; Pearce, M.S.; Phillips, E.; Gregory, W.; Barton, J.R.; Welfare, M.R. Influence of dietary factors on the clinical course of ulcerative colitis: A prospective cohort study. Gut 2004, 53, 1479–1484. [Google Scholar] [CrossRef]
- Opstelten, J.L.; de Vries, J.H.M.; Wools, A.; Siersema, P.D.; Oldenburg, B.; Witteman, B.J.M. Dietary intake of patients with inflammatory bowel disease: A comparison with individuals from a general population and associations with relapse. Clin. Nutr. 2019, 38, 1892–1898. [Google Scholar] [CrossRef]
- Shivappa, N.; Hébert, J.R.; Rashvand, S.; Rashidkhani, B.; Hekmatdoost, A. Inflammatory Potential of Diet and Risk of Ulcerative Colitis in a Case-Control Study from Iran. Nutr. Cancer 2016, 68, 404–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naqvi, S.A.; Taylor, L.M.; Panaccione, R.; Ghosh, S.; Barkema, H.W.; Hotte, N.; Shommu, N.; Kaur, S.; Reimer, R.A.; Madsen, K.L.; et al. Dietary patterns, food groups and nutrients in Crohn’s disease: Associations with gut and systemic inflammation. Sci. Rep. 2021, 11, 1674. [Google Scholar] [CrossRef] [PubMed]
- Campmans-Kuijpers, M.J.E.; Dijkstra, G. Food and Food Groups in Inflammatory Bowel Disease (IBD): The Design of the Groningen Anti-Inflammatory Diet (GrAID). Nutrients 2021, 13, 1067. [Google Scholar] [CrossRef] [PubMed]
- Forbes, A.; Escher, J.; Hébuterne, X.; Klęk, S.; Krznaric, Z.; Schneider, S.; Shamir, R.; Stardelova, K.; Wierdsma, N.; Wiskin, A.E.; et al. ESPEN guideline: Clinical nutrition in inflammatory bowel disease. Clin. Nutr. 2017, 36, 321–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imhann, F.; van der Velde, K.J.; Barbieri, R.; Alberts, R.; Voskuil, M.D.; Vich Vila, A.; Collij, V.; Spekhorst, L.M.; van der Sloot, K.W.J.; Peters, V.; et al. The 1000IBD project: Multi-omics data of 1000 inflammatory bowel disease patients; data release 1. BMC Gastroenterol. 2019, 19, 5. [Google Scholar]
- Assarsson, E.; Lundberg, M.; Holmquist, G.; Björkesten, J.; Thorsen, S.B.; Ekman, D.; Eriksson, A.; Dickens, E.R.; Ohlsson, S.; Edfeldt, G.; et al. Homogenous 96-plex PEA immunoassay exhibiting high sensitivity, specificity, and excellent scalability. PLoS ONE 2014, 9, e95192. [Google Scholar] [CrossRef] [Green Version]
- Berkhout, L.C.; L’Ami, M.J.; Ruwaard, J.; Hart, M.H.; Ooijevaar-de Heer, P.; Bloem, K.; Nurmohamed, M.T.; van Vollenhoven, R.F.; Boers, M.; Alvarez, D.F.; et al. Dynamics of circulating TNF during adalimumab treatment using a drug-tolerant TNF assay. Sci. Transl. Med. 2019, 11, eaat3356. [Google Scholar] [CrossRef] [Green Version]
- van Schie, K.A.; Ooijevaar-de Heer, P.; Dijk, L.; Kruithof, S.; Wolbink, G.; Rispens, T. Therapeutic TNF Inhibitors can Differentially Stabilize Trimeric TNF by Inhibiting Monomer Exchange. Sci. Rep. 2016, 6, 32747. [Google Scholar] [CrossRef] [Green Version]
- Siebelink, E.; Geelen, A.; de Vries, J.H.M. Self-reported energy intake by FFQ compared with actual energy intake to maintain body weight in 516 adults. Br. J. Nutr. 2011, 106, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Willett, W.C.; Sampson, L.; Stampfer, M.J.; Rosner, B.; Bain, C.; Witschi, J.; Hennekens, C.H.; Speizer, F.E. Reproducibility and validity of a semiquantitative food frequency questionnaire. Am. J. Epidemiol. 1985, 122, 51–65. [Google Scholar] [CrossRef]
- Banna, J.C.; McCrory, M.A.; Fialkowski, M.K.; Boushey, C. Examining Plausibility of Self-Reported Energy Intake Data: Considerations for Method Selection. Front. Nutr. 2017, 4, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourgonje, A.R.; Hu, S.; Spekhorst, L.M.; Zhernakova, D.V.; Vich Vila, A.; Li, Y.; Voskuil, M.D.; van Berkel, L.A.; Bley Folly, B.; Charrout, M.; et al. The Effect of Phenotype and Genotype on the Plasma Proteome in Patients with Inflammatory Bowel Disease. J. Crohns Colitis 2021, 16, 414–429. [Google Scholar] [CrossRef] [PubMed]
- Peters, V.; Tigchelaar-Feenstra, E.F.; Imhann, F.; Dekens, J.A.M.; Swertz, M.A.; Franke, L.H.; Wijmenga, C.; Weersma, R.K.; Alizadeh, B.Z.; Dijkstra, G.; et al. Habitual dietary intake of IBD patients differs from population controls: A case-control study. Eur. J. Nutr. 2021, 60, 345–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, K.L. Dietary patterns, approaches, and multicultural perspective. Appl. Physiol. Nutr. Metab. 2010, 35, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.S.; Jess, T.; Vind, I.; Elkjaer, M.; Nielsen, M.F.; Gamborg, M.; Munkholm, P. Environmental factors in inflammatory bowel disease: A case-control study based on a Danish inception cohort. J. Crohns Colitis 2011, 5, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Gadaleta, R.M.; Moschetta, A. Metabolic Messengers: Fibroblast growth factor 15/19. Nat. Metab. 2019, 1, 588–594. [Google Scholar] [CrossRef]
- Nolan, J.D.; Johnston, I.M.; Pattni, S.S.; Dew, T.; Orchard, T.R.; Walters, J.R.F. Diarrhea in Crohn’s disease: Investigating the role of the ileal hormone fibroblast growth factor 19. J. Crohns Colitis 2015, 9, 125–131. [Google Scholar] [CrossRef]
- Vítek, L. Bile acid malabsorption in inflammatory bowel disease. Inflamm. Bowel Dis. 2015, 21, 476–483. [Google Scholar] [CrossRef]
- Pavlidis, P.; Powell, N.; Vincent, R.P.; Ehrlich, D.; Bjarnason, I.; Hayee, B. Systematic review: Bile acids and intestinal inflammation-luminal aggressors or regulators of mucosal defence? Aliment. Pharmacol. Ther. 2015, 42, 802–817. [Google Scholar] [CrossRef]
- Friedrich, D.; Marschall, H.U.; Lammert, F. Response of fibroblast growth factor 19 and bile acid synthesis after a body weight-adjusted oral fat tolerance test in overweight and obese NAFLD patients: A non-randomized controlled pilot trial. BMC Gastroenterol. 2018, 18, 76. [Google Scholar] [CrossRef] [Green Version]
- Wojcik, M.; Janus, D.; Dolezal-Oltarzewska, K.; Kalicka-Kasperczyk, A.; Poplawska, K.; Drozdz, D.; Sztefko, K.; Starzyk, J.B. A decrease in fasting FGF19 levels is associated with the development of non-alcoholic fatty liver disease in obese adolescents. J. Pediatr. Endocrinol. Metab. 2012, 25, 1089–1093. [Google Scholar] [CrossRef] [PubMed]
- van Schaik, F.D.M.; Gadaleta, R.M.; Schaap, F.G.; van Mil, S.W.C.; Siersema, P.D.; Oldenburg, B.; van Erpecum, K.J. Pharmacological activation of the bile acid nuclear farnesoid X receptor is feasible in patients with quiescent Crohn’s colitis. PLoS ONE 2012, 7, e49706. [Google Scholar] [CrossRef] [PubMed]
- Gadaleta, R.M.; Garcia-Irigoyen, O.; Cariello, M.; Scialpi, N.; Peres, C.; Vetrano, S.; Fiorino, G.; Danese, S.; Ko, B.; Luo, J.; et al. Fibroblast Growth Factor 19 modulates intestinal microbiota and inflammation in presence of Farnesoid X Receptor. EBioMedicine 2020, 54, 102719. [Google Scholar] [CrossRef] [PubMed]
- Gadaleta, R.M.; van Erpecum, K.J.; Oldenburg, B.; Willemsen, E.C.L.; Renooij, W.; Murzilli, S.; Klomp, L.W.J.; Siersema, P.D.; Schipper, M.E.I.; Danese, S.; et al. Farnesoid X receptor activation inhibits inflammation and preserves the intestinal barrier in inflammatory bowel disease. Gut 2011, 60, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Cai, C.; Chen, Q.; Jin, S.; Yang, B.; Li, N. High-Fat Diet Promotes DSS-Induced Ulcerative Colitis by Downregulated FXR Expression through the TGFB Pathway. Biomed. Res. Int. 2020, 2020, 3516128. [Google Scholar] [CrossRef] [PubMed]
- Morton, G.J.; Kaiyala, K.J.; Foster-Schubert, K.E.; Cummings, D.E.; Schwartz, M.W. Carbohydrate feeding dissociates the postprandial FGF19 response from circulating bile acid levels in humans. J. Clin. Endocrinol. Metab. 2014, 99, E241–E245. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, I.; Ceulemans, M.; Wauters, L.; Breynaert, C.; Vermeire, S.; Verstockt, B.; Vanuytsel, T. Role of Eosinophils in Intestinal Inflammation and Fibrosis in Inflammatory Bowel Disease: An Overlooked Villain? Front. Immunol. 2021, 12, 754413. [Google Scholar] [CrossRef]
- Ahrens, R.; Waddell, A.; Seidu, L.; Blanchard, C.; Carey, R.; Forbes, E.; Lampinen, M.; Wilson, T.; Cohen, E.; Stringer, K.; et al. Intestinal macrophage/epithelial cell-derived CCL11/eotaxin-1 mediates eosinophil recruitment and function in pediatric ulcerative colitis. J. Immunol. 2008, 181, 7390–7399. [Google Scholar] [CrossRef]
- Mir, A.; Minguez, M.; Tatay, J.; Pascual, I.; Peña, A.; Sanchiz, V.; Almela, P.; Mora, F.; Benages, A. Elevated serum eotaxin levels in patients with inflammatory bowel disease. Am. J. Gastroenterol. 2002, 97, 1452–1457. [Google Scholar] [CrossRef]
- Huang, M.C.; Chung, R.H.; Lin, P.H.; Kuo, H.W.; Liu, T.H.; Chen, Y.Y.; Chen, A.C.H.; Liu, Y.L. Increase in plasma CCL11 (Eotaxin-1) in patients with alcohol dependence and changes during detoxification. Brain Behav. Immun. 2022, 99, 83–90. [Google Scholar] [CrossRef]
- Jeon, J.I.; Ko, S.H.; Kim, Y.J.; Choi, S.M.; Kang, K.K.; Kim, H.; Yoon, H.J.; Kim, J.M. The flavone eupatilin inhibits eotaxin expression in an NF-κB-dependent and STAT6-independent manner. Scand. J. Immunol. 2015, 81, 166–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasudevan, A.R.; Wu, H.; Xydakis, A.M.; Jones, P.H.; O’Brian Smith, E.; Sweeney, J.F.; Corry, D.B.; Ballantyne, C.M. Eotaxin and obesity. J. Clin. Endocrinol. Metab. 2006, 91, 256–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Sloot, K.W.J.; Joshi, A.D.; Bellavance, D.R.; Gilpin, K.K.; Stewart, K.O.; Lochhead, P.; Garber, J.J.; Giallourakis, C.; Yajnik, V.; Ananthakrishnan, A.N.; et al. Visceral Adiposity, Genetic Susceptibility, and Risk of Complications Among Individuals with Crohn’s Disease. Inflamm. Bowel Dis. 2017, 23, 82–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoury, T.; Rotnemer-Golinkin, D.; Zolotarev, L.; Ilan, Y. Orally administered anti-eotaxin-1 monoclonal antibody is biologically active in the gut and alleviates immune-mediated hepatitis: A novel anti-inflammatory personalized therapeutic approach. Int. J. Immunopathol. Pharmacol. 2021, 35, 20587384211021215. [Google Scholar] [CrossRef]
- Mandrekar, P.; Catalano, D.; Dolganiuc, A.; Kodys, K.; Szabo, G. Inhibition of myeloid dendritic cell accessory cell function and induction of T cell anergy by alcohol correlates with decreased IL-12 production. J. Immunol. 2004, 173, 3398–3407. [Google Scholar] [CrossRef] [Green Version]
- Kuang, A.; Erlund, I.; Herder, C.; Westerhuis, J.A.; Tuomilehto, J.; Cornelis, M.C. Targeted proteomic response to coffee consumption. Eur. J. Nutr. 2020, 59, 1529–1539. [Google Scholar] [CrossRef]
- Link, A.A.; Kino, T.; Worth, J.A.; McGuire, J.L.; Crane, M.L.; Chrousos, G.P.; Wilder, R.L.; Elenkov, I.J. Ligand-activation of the adenosine A2a receptors inhibits IL-12 production by human monocytes. J. Immunol. 2000, 164, 436–442. [Google Scholar] [CrossRef]
- Glas, J.; Seiderer, J.; Wagner, J.; Olszak, T.; Fries, C.; Tillack, C.; Friedrich, M.; Beigel, F.; Stallhofer, J.; Steib, C.; et al. Analysis of IL12B gene variants in inflammatory bowel disease. PLoS ONE 2012, 7, e34349. [Google Scholar] [CrossRef]
- Opstad, T.B.; Brusletto, B.S.; Arnesen, H.; Pettersen, A.A.; Seljeflot, I. Cigarette smoking represses expression of cytokine IL-12 and its regulator miR-21-An observational study in patients with coronary artery disease. Immunobiology 2017, 222, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Zeevi, D.; Korem, T.; Zmora, N.; Israeli, D.; Rothschild, D.; Weinberger, A.; Ben-Yacov, O.; Lador, D.; Avnit-Sagi, T.; Lotan-Pompan, M.; et al. Personalized Nutrition by Prediction of Glycemic Responses. Cell 2015, 163, 1079–1084. [Google Scholar] [CrossRef] [Green Version]
- Continuous Monitoring of Inflammatory Biomarkers, Seerave Foundation. Available online: https://seerave.org/projects/feasibility-of-continuous-inflammation-monitoring/ (accessed on 13 June 2022).




{kind=link}
{kind=link}
{kind=link}
| Variable | Total | CD | UC | p-Value |
|---|---|---|---|---|
| n = 454 | n = 264 | n = 190 | ||
| Age (years) | 41.4 ± 14.4 | 39.2 ± 14.1 | 44.5 ± 14.2 | <0.01 |
| Sex, n (%) | <0.01 | |||
| Male | 173 (38.1) | 87 (33.0) | 86 (45.3) | |
| Female | 281 (61.9) | 177 (67.0) | 104 (54.7) | |
| BMI (kg/m2) | 24.7 [21.9–28.1] | 23.9 [21.4–27.6] | 25.5 [22.8–29.1] | <0.01 |
| Current smoking, n (%) | 441 (97.1) | 257 (97.3) | 184 (96.8) | <0.01 |
| Yes | 93 (20.5) | 73 (28.4) | 20 (10.9) | |
| No | 348 (76.7) | 184 (71.6) | 164 (89.1) | |
| Montreal classification | ||||
| Montreal Age (A), n (%) | 453 (99.8) | 263 (99.6) | 190 (100) | <0.01 |
| A1 (≤16 years) | 58 (12.8) | 42 (16.0) | 16 (8.4) | |
| A2 (17–40 years) | 298 (65.6) | 179 (68.1) | 119 (62.6) | |
| A3 (>40 years) | 97 (21.4) | 42 (16.0) | 55 (28.9) | |
| Montreal Location (L), n (%) | - | 264 (100) | - | |
| L1 (ileal disease) | - | 92 (34.8) | - | |
| L2 (colonic disease) | - | 58 (22.0) | - | |
| L3 (ileocolonic disease) | - | 91 (34.5) | - | |
| L4 (upper GI disease) | - | 5 (1.9) | - | |
| L1 + L4 | - | 8 (3.0) | - | |
| L2 + L4 | - | 6 (2.3) | - | |
| L3 + L4 | - | 4 (1.5) | - | |
| Montreal Behaviour (B), n (%) | - | 264 (100) | - | |
| B1 (non-stricturing, non-penetrating) | - | 105 (39.8) | - | |
| B2 (stricturing) | - | 50 (18.9) | - | |
| B3 (penetrating) | - | 24 (9.1) | - | |
| B1 + P (perianal disease) | - | 32 (12.1) | - | |
| B2 + P (perianal disease) | - | 39 (14.8) | - | |
| B3 + P (perianal disease) | - | 14 (5.3) | - | |
| Montreal Extension (E), n (%) | - | - | 187 (98.4) | |
| E1 (proctitis) | - | - | 29 (15.5) | |
| E2 (left-sided colitis) | - | - | 61 (32.6) | |
| E3 (pancolitis) | - | - | 97 (51.9) | |
| Medication use | ||||
| Aminosalicylates, n (%) | 158 (34.8) | 27 (10.2) | 131 (68.9) | <0.01 |
| Thiopurines, n (%) | 169 (37.2) | 111 (42.0) | 58 (30.5) | 0.01 |
| Steroids, n (%) | 106 (23.3) | 65 (24.6) | 41 (21.6) | 0.45 |
| Calcineurin inhibitors, n (%) | 8 (1.8) | 2 (0.8) | 6 (3.2) | 0.06 |
| Methotrexate, n (%) | 32 (7.0) | 28 (10.6) | 4 (2.1) | <0.01 |
| TNF-α-antagonists, n (%) † | 115 (25.3) | 100 (37.9) | 15 (7.9) | <0.01 |
| Disease activity | ||||
| HBI (CD), n (%) | - | 252 (95.5) | - | |
| <5 | - | 161 (63.9) | - | |
| ≥5 | - | 91 (36.1) | - | |
| SCCAI (UC), n (%) | - | - | 183 (96.3) | |
| ≤2 | - | - | 130 (71.4) | |
| >2 | - | - | 53 (28.6) | |
| CRP, n (%) | 382 (84.1) | 224 (84.8) | 158 (83.2) | <0.05 |
| ≤5 mg/L | 279 (61.5) | 153 (68.3) | 126 (79.7) | |
| >5 mg/L | 103 (22.7) | 71 (31.7) | 32 (20.3) | |
| Surgical history | ||||
| Ileocecal resection, n (%) | 87 (19.2) | 87 (33.0) | 0 (0.0) | <0.01 |
| Colon resection (or partial), n (%) | 76 (16.7) | 43 (16.3) | 33 (17.4) | 0.76 |
| IBD | CD | UC | p-Value | |
|---|---|---|---|---|
| n = 454 | n = 264 | n = 190 | ||
| Macronutrients intake per day | ||||
| Energy intake (kcal/day) | 1,824 (1,519;2,258) | 1,780 (1,480;2,194) | 1,917 (1,552;2,364) | 0.059 |
| EI/BMR | 1.14 (0.93–1.42) | 1.17 (0.93–1.42) | 1.11 (0.93–1.41) | 0.994 |
| Total protein (g/day) | 65.7 (54.3;80.5) | 63.6 (52.2–76.5) | 70.6 (57.5–83.5) | 0.001 * |
| Energy (%) | 14.3 (12.7–15.9) | 14.2 (12.7–15.6) | 14.5 (12.7–16.2) | 0.050 |
| Protein (g/kg) | 0.87 (0.70–1.06) | 0.89 (0.70–1.07) | 0.84 (0.70–1.06) | 0.768 |
| Plant protein (g/day) | 27.3 (22.2;34.6) | 26.2 (21.4–33.3) | 28.1 (23.2–35.8) | 0.015 |
| Animal protein (g/day) | 37.9 (29.4;47.6) | 36.1 (28.2–45.8) | 41.1 (32.2–49.5) | 0.003 * |
| Total fat (g/day) | 72.1 (57.2;90.7) | 69.3 (55.4–89.5) | 75.2 (59.7–98.2) | 0.038 |
| Fat Energy (%) | 35.7 (31.8–39.4) | 35.6 (31.6–39.1) | 36.0 (32.3–39.9) | 0.275 |
| Carbohydrates (g/day) | 208 (165;267) | 211 (161–261) | 207 (171–283) | 0.405 |
| Carbohydrate Energy (%) † | 45.9 (41.8–49.7) | 46.3 (42.5–50.4) | 45.6 (41.3–48.6) | 0.028 |
| Alcohol (g/day) | 1.3 (0.0;5.9) | 1.2 (0.0–5.0) | 1.5 (0.0–6.7) | 0.323 |
| Alcohol Energy (%) | 0.5 (0.0–2.0) | 0.4 (0.0–1.9) | 0.6 (0.0–2.0) | 0.467 |
| Food groups (g/day) | ||||
| Alcoholic beverages | 13.4 (0.0–71.1) | 11.9 (0.0–63.9) | 16.2 (0.0–77.6) | 0.276 |
| Breads | 130 (80.0–164) | 127 (77.4–159) | 133 (90.1–174) | 0.069 |
| Cereals | 0.0 (0.0–3.5) | 0.0 (0.0–2.9) | 0.0 (0.0–4.8) | 0.309 |
| Cheese | 21.1 (8.5–36.9) | 20.7 (8.1–33.0) | 21.8 (9.2–38.9) | 0.236 |
| Coffee | 232 (17.9–465) | 232 (11.1–465) | 232 (22.3–465) | 0.925 |
| Dairy | 182 (96.5–330) | 159 (81.9–298) | 242 (123–348) | 0.001 * |
| Eggs | 8.9 (4.5–17.9) | 8.9 (4.5–17.9) | 8.9 (4.5–17.9) | 0.931 |
| Fish | 11.1 (4.4–17.7) | 11.0 (4.2–17.3) | 11.3 (4.9–18.3) | 0.271 |
| Fruits | 110(42.3–220) | 84.6 (42.3–220) | 119 (50.9–220) | 0.020 |
| Juice | 26.7 (0.0–107) | 26.7 (0.0–139) | 21.5 (0.0–96.5) | 0.056 |
| Legumes | 2.2 (0.0–11.0) | 0.0 (0.0–11.0) | 4.4 (0.0–16.4) | 0.021 |
| Meat | 86.1 (58.5–111) | 84.1 (49.7–109) | 91.6 (65.9–113) | 0.082 |
| Non-alcoholic beverages | 104 (20.9–278) | 136 (26.2–284) | 52.9 (13.0–271) | <0.001 * |
| Nuts | 5.4 (1.8–13.2) | 4.3 (1.4–12.6) | 6.7 (2.1–14.1) | 0.063 |
| Pasta | 12.7 (7.9–25.5) | 12.7 (7.9–25.5) | 15.9 (7.9–31.8) | 0.065 |
| Pastry | 23.8 (12.4–40.3) | 21.8 (11.3–39.0) | 26.7 (14.7–44.2) | 0.015 |
| Potatoes | 72.3 (40.8–111) | 71.3 (39.6–104) | 85.6 (41.3–119) | 0.181 |
| Prepared meals | 32.0 (12.9–58.8) | 32.4 (12.9–59.9) | 27.6 (12.9–54.2) | 0.265 |
| Rice | 14.9 (4.7–24.8) | 14.9 (4.0–24.8) | 14.9 (5.5–24.8) | 0.574 |
| Sauces | 10.0 (4.6–20.8) | 10.5 (4.3–21.3) | 9.2 (4.7–20.5) | 0.860 |
| Savoury snacks | 13.1 (5.4–23.9) | 13.4 (5.1–24.7) | 12.8 (5.8–23.6) | 0.760 |
| Soup | 35.8 (9.0–71.5) | 35.8 (9.0–44.5) | 35.8 (15.8–71.5) | 0.335 |
| Spreads | 20.6 (8.3–31.4) | 19.1 (7.1–30.8) | 22.5 (9.5–32.6) | 0.146 |
| Sugar/Sweets | 29.8 (14.1–49.6) | 30.3 (12.9–48.7) | 28.9 (15.2–50.0) | 0.916 |
| Tea | 232 (44.6–465) | 232 (44.6–348) | 232 (44.6–465) | 0.841 |
| Vegetables | 107 (62.4–147) | 81.8 (61.5–114) | 108 (62.8–151) | 0.225 |
| PC1 | PC2 | PC3 | PC4 | PC5 | |
|---|---|---|---|---|---|
| Alcohol | −0.150 | −0.001 | 0.366 | 0.494 | −0.037 |
| Breads | 0.703 | 0.065 | 0.025 | 0.249 | 0.146 |
| Cereals | 0.050 | 0.343 | 0.049 | 0.121 | −0.170 |
| Cheese | 0.047 | 0.156 | 0.190 | 0.264 | 0.209 |
| Coffee | 0.039 | 0.066 | −0.079 | 0.672 | 0.129 |
| Dairy | 0.288 | 0.133 | −0.246 | 0.304 | 0.066 |
| Eggs | 0.184 | 0.382 | 0.237 | −0.155 | 0.180 |
| Fish | −0.038 | 0.560 | −0.012 | −0.029 | 0.114 |
| Fruit | −0.015 | 0.632 | −0.264 | −0.087 | 0.105 |
| Juice | 0.338 | −0.109 | −0.065 | −0.116 | 0.028 |
| Legumes | 0.009 | 0.061 | 0.146 | 0.016 | 0.514 |
| Meat | 0.257 | −0.386 | −0.001 | 0.189 | 0.550 |
| Non-alcoholic beverages | 0.257 | −0.466 | 0.138 | −0.257 | 0.065 |
| Nuts | 0.031 | 0.426 | 0.351 | −0.026 | −0.072 |
| Pasta | 0.032 | 0.028 | 0.655 | 0.074 | 0.219 |
| Pastry | 0.585 | 0.102 | 0.170 | −0.168 | 0.060 |
| Potatoes | 0.290 | −0.247 | −0.079 | 0.136 | 0.651 |
| Prepared meals | 0.163 | −0.062 | 0.519 | 0.143 | −0.263 |
| Rice | −0.104 | 0.201 | 0.402 | −0.157 | 0.204 |
| Sauces | 0.249 | −0.164 | 0.659 | 0.036 | 0.130 |
| Savoury snacks | 0.510 | −0.228 | 0.325 | −0.195 | 0.101 |
| Soup | 0.110 | 0.242 | 0.097 | 0.180 | 0.256 |
| Spreads | 0.707 | 0.087 | −0.009 | 0.325 | 0.090 |
| Sugar/Sweets | 0.532 | −0.007 | 0.106 | 0.024 | −0.145 |
| Tea | 0.002 | 0.366 | −0.051 | −0.572 | 0.107 |
| Vegetables | −0.173 | 0.258 | 0.058 | −0.119 | 0.659 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bourgonje, A.R.; Bolte, L.A.; Vranckx, L.L.C.; Spekhorst, L.M.; Gacesa, R.; Hu, S.; van Dullemen, H.M.; Visschedijk, M.C.; Festen, E.A.M.; Samsom, J.N.; et al. Long-Term Dietary Patterns Are Reflected in the Plasma Inflammatory Proteome of Patients with Inflammatory Bowel Disease. Nutrients 2022, 14, 2522. https://doi.org/10.3390/nu14122522
Bourgonje AR, Bolte LA, Vranckx LLC, Spekhorst LM, Gacesa R, Hu S, van Dullemen HM, Visschedijk MC, Festen EAM, Samsom JN, et al. Long-Term Dietary Patterns Are Reflected in the Plasma Inflammatory Proteome of Patients with Inflammatory Bowel Disease. Nutrients. 2022; 14(12):2522. https://doi.org/10.3390/nu14122522
Chicago/Turabian StyleBourgonje, Arno R., Laura A. Bolte, Lianne L. C. Vranckx, Lieke M. Spekhorst, Ranko Gacesa, Shixian Hu, Hendrik M. van Dullemen, Marijn C. Visschedijk, Eleonora A. M. Festen, Janneke N. Samsom, and et al. 2022. "Long-Term Dietary Patterns Are Reflected in the Plasma Inflammatory Proteome of Patients with Inflammatory Bowel Disease" Nutrients 14, no. 12: 2522. https://doi.org/10.3390/nu14122522
APA StyleBourgonje, A. R., Bolte, L. A., Vranckx, L. L. C., Spekhorst, L. M., Gacesa, R., Hu, S., van Dullemen, H. M., Visschedijk, M. C., Festen, E. A. M., Samsom, J. N., Dijkstra, G., Weersma, R. K., & Campmans-Kuijpers, M. J. E. (2022). Long-Term Dietary Patterns Are Reflected in the Plasma Inflammatory Proteome of Patients with Inflammatory Bowel Disease. Nutrients, 14(12), 2522. https://doi.org/10.3390/nu14122522