
The Antidiabetic Activities of Neocryptotanshinone: Screened by Molecular Docking and Related to the Modulation of PTP1B


Abstract
:1. Introduction
2. Materials and Methods
2.1. High-Throughput Virtual Screening and Molecular Docking
2.2. Animal Feeding and NCTS Administration
2.3. Oral Glucose Tolerance Test (OGTT)
2.4. Food and Water Intake Measurement
2.5. Hematoxylin and Eosin (H&E) Staining
2.6. Enzyme-Linked Immunosorbent Assay (ELISA) Analyses
2.7. Western Blotting
2.8. Immunohistochemical Examination
2.9. Statistical Analysis
3. Results
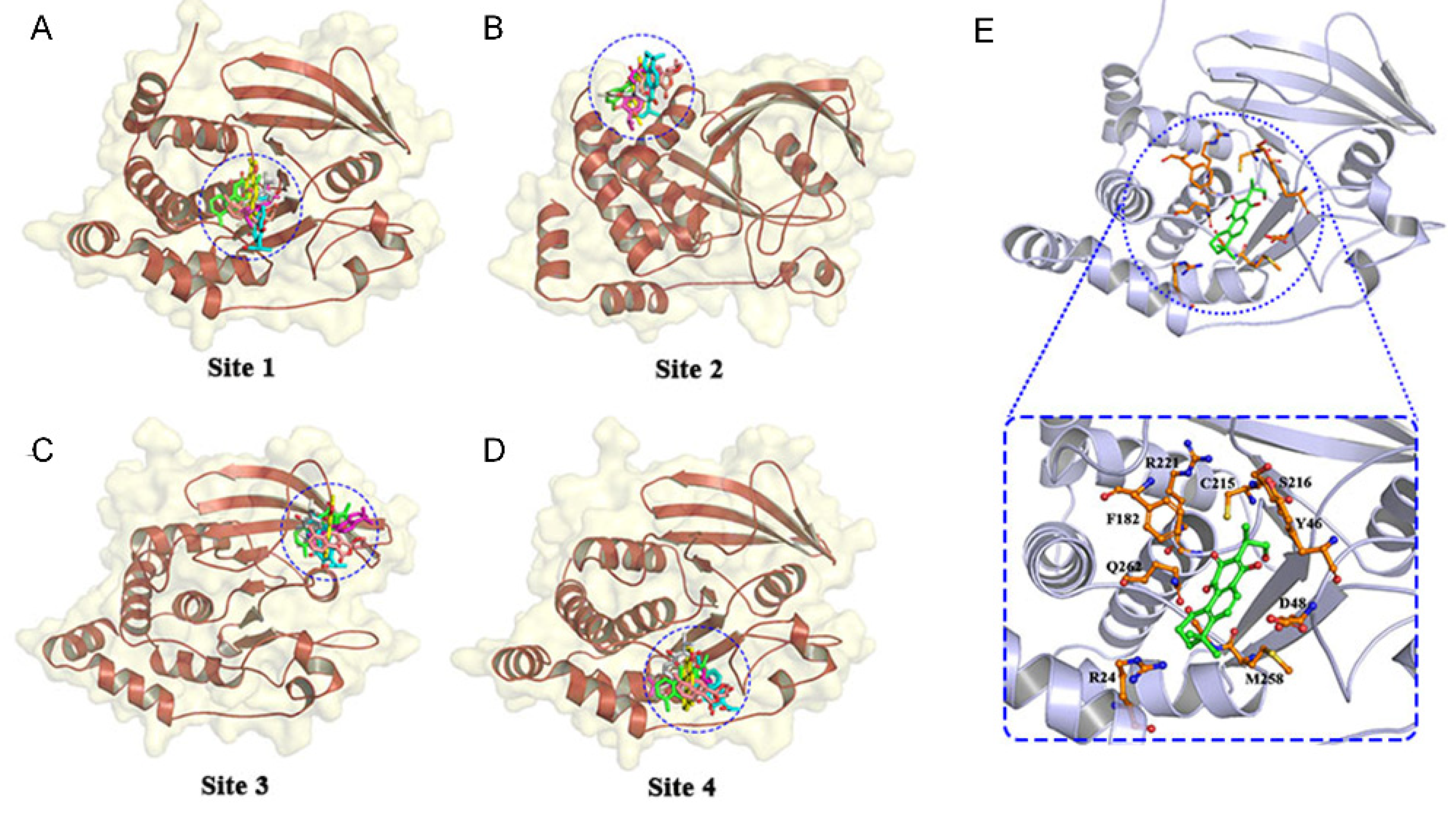
3.1. High-Throughput Virtual Screening and Molecular Docking
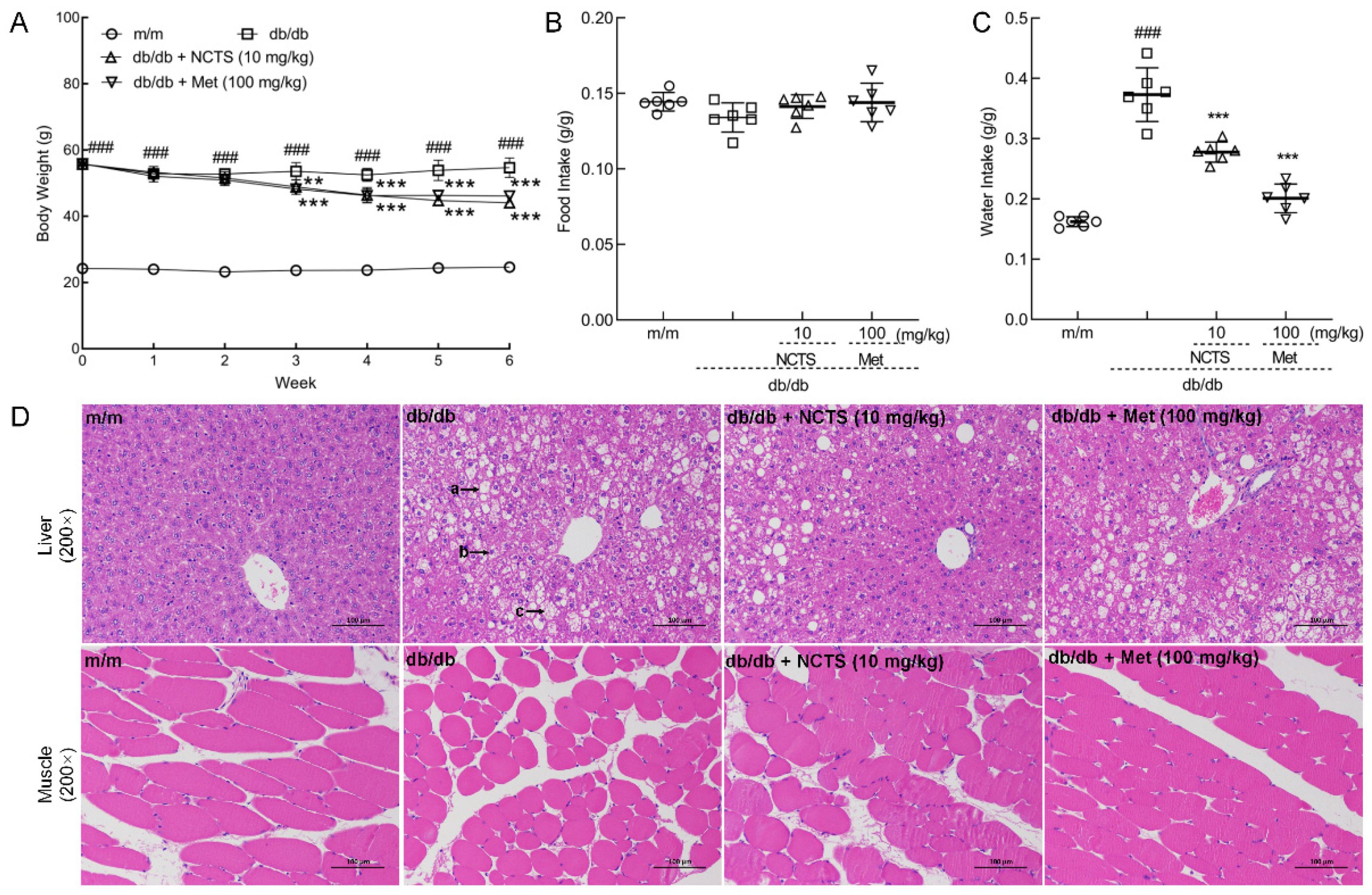
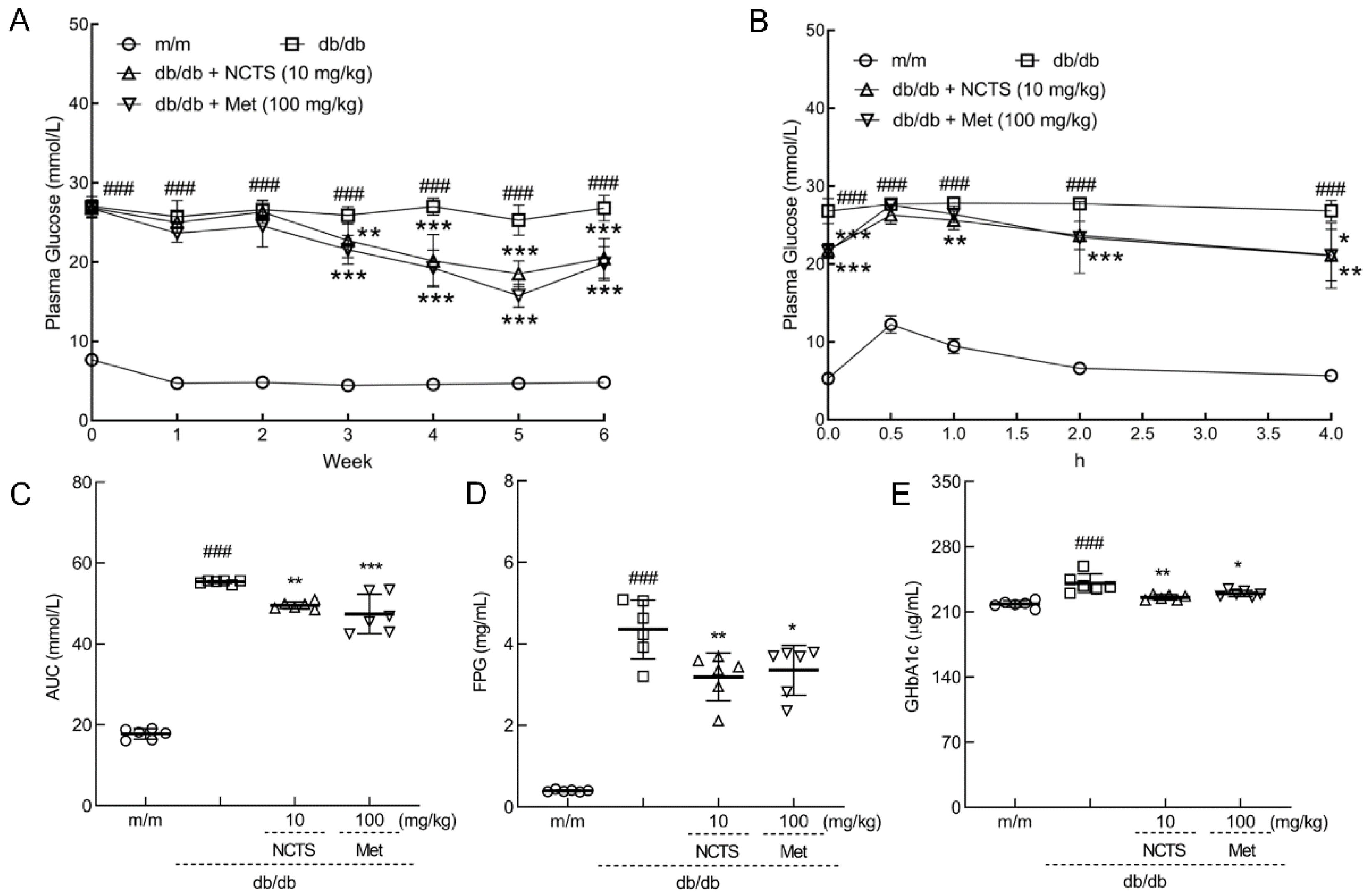
3.2. Effect of NCTS on Hypoglycemia in db/db Mice
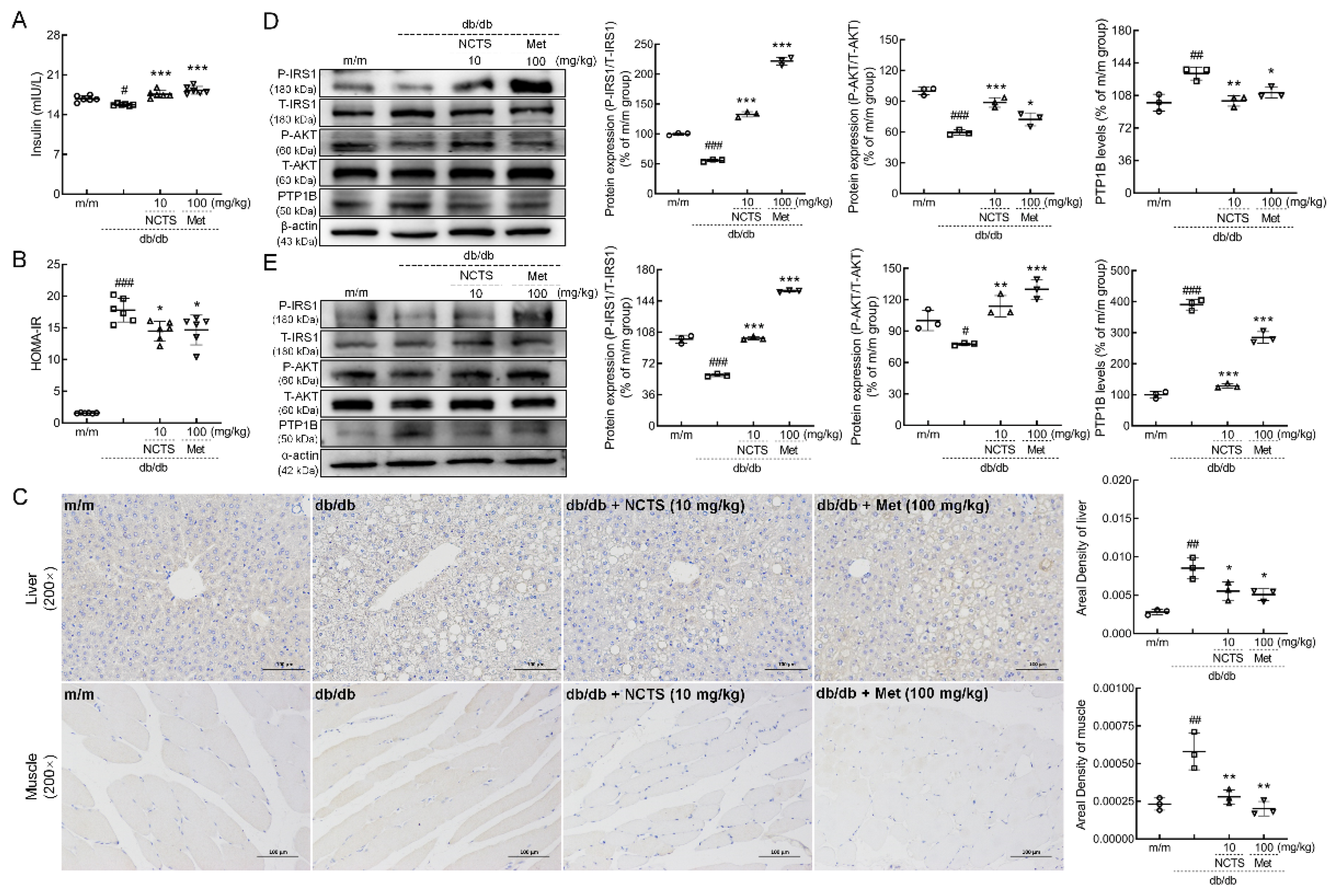
3.3. NCTS-Modulated IR Related to PTP1B in db/db Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ho, N.; Sommers, M.S.; Lucki, I. Effects of diabetes on hippocampal neurogenesis: Links to cognition and depression. Neurosci. Biobehav. Rev. 2013, 37, 1346–1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Z.; Peng, Y.; Qiu, J.; Cao, A.; Wang, G.; Peng, Z. Synthesis, In Vitro α-Glucosidase Inhibitory Activity and Molecular Docking Studies of Novel Benzothiazole-Triazole Derivatives. Molecules 2017, 22, 1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahrani, A.A.; Barnett, A.H.; Bailey, C.J. Pharmacology and therapeutic implications of current drugs for type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2016, 12, 566–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeFronzo, R.A.; Ferrannini, E.; Groop, L.; Henry, R.R.; Herman, W.H.; Holst, J.J.; Hu, F.B.; Kahn, C.R.; Raz, I.; Shulman, G.I.; et al. Type 2 diabetes mellitus. Nat. Rev. Dis. Primers 2015, 1, 15019. [Google Scholar] [CrossRef]
- Jonietz, E. Pathology: Cause and effect. Nature 2012, 485, S10–S11. [Google Scholar] [CrossRef] [Green Version]
- He, R.-J.; Yu, Z.-H.; Zhang, R.-Y.; Zhang, Z.-Y. Protein tyrosine phosphatases as potential therapeutic targets. Acta Pharm. Sin. 2014, 35, 1227–1246. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Wang, L.; Xie, J.; Li, X.; Liu, S.; Qiu, S.; Hu, Y.; Shen, X. Treatment of type 2 diabetes mellitus via reversing insulin resistance and regulating lipid homeostasis in vitro and in vivo using cajanonic acid A. Int. J. Mol. Med. 2018, 42, 2329–2342. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Tang, Z.; Shen, A.; Tao, T.; Wan, C.; Zhu, X.; Huang, J.; Zhang, W.; Xia, N.; Wang, S.; et al. The Role of PTP1B O-GlcNAcylation in Hepatic Insulin Resistance. Int. J. Mol. Sci 2015, 16, 22856–22869. [Google Scholar] [CrossRef] [Green Version]
- Haj, F.G.; Zabolotny, J.M.; Kim, Y.B.; Kahn, B.B.; Neel, B.G. Liver-specific protein-tyrosine phosphatase 1B (PTP1B) re-expression alters glucose homeostasis of PTP1B-/-mice. J. Biol. Chem. 2005, 280, 15038–15046. [Google Scholar] [CrossRef] [Green Version]
- Ito, M.; Fukuda, S.; Sakata, S.; Morinaga, H.; Ohta, T. Pharmacological effects of JTT-551, a novel protein tyrosine phosphatase 1B inhibitor, in diet-induced obesity mice. J. Diabetes Res. 2014, 2014, 680348. [Google Scholar] [CrossRef] [Green Version]
- Kuga, G.K.; Muñoz, V.R.; Gaspar, R.C.; Nakandakari, S.C.B.R.; da Silva, A.S.R.; Botezelli, J.D.; Leme, J.A.C.d.A.; Gomes, R.J.; de Moura, L.P.; Cintra, D.E.; et al. Impaired insulin signaling and spatial learning in middle-aged rats: The role of PTP1B. Exp. Gerontol. 2018, 104, 66–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Moura, L.P.; Souza Pauli, L.S.; Cintra, D.E.; de Souza, C.T.; da Silva, A.S.R.; Marinho, R.; de Melo, M.A.R.; Ropelle, E.R.; Pauli, J.R. Acute exercise decreases PTP-1B protein level and improves insulin signaling in the liver of old rats. Immun. Ageing 2013, 10, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauli, J.R.; Ropelle, E.R.; Cintra, D.E.; De Souza, C.T.; da Silva, A.S.R.; Moraes, J.C.; Prada, P.O.; de Almeida Leme, J.A.C.; Luciano, E.; Velloso, L.A.; et al. Acute exercise reverses aged-induced impairments in insulin signaling in rodent skeletal muscle. Mech. Ageing Dev. 2010, 131, 323–329. [Google Scholar] [CrossRef]
- Da Cruz Rodrigues, K.C.; Martins Pereira, R.; Peruca, G.F.; Torres Barbosa, L.W.; Ramos Sant’Ana, M.; Rosetto Muñoz, V.; Morelli, A.P.; Moreira Simabuco, F.; Sanchez Ramos da Silva, A.; Esper Cintra, D.; et al. Short-Term Strength Exercise Reduces Hepatic Insulin Resistance in Obese Mice by Reducing PTP1B Content, Regardless of Changes in Body Weight. Int. J. Mol. Sci. 2021, 22, 6402. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Konstantopoulos, N.; Lee, J.; Hansen, L.; Li, Z.W.; Karin, M.; Shoelson, S.E. Reversal of obesity- and diet-induced insulin resistance with salicylates or targeted disruption of Ikkbeta. Science 2001, 293, 1673–1677. [Google Scholar] [CrossRef] [PubMed]
- Inzucchi, S.E.; Bergenstal, R.M.; Buse, J.B.; Diamant, M.; Ferrannini, E.; Nauck, M.; Peters, A.L.; Tsapas, A.; Wender, R.; Matthews, D.R. Management of hyperglycaemia in type 2 diabetes: A patient-centered approach. Position statement of the American Diabetes Association (ADA) and the European Association for the Study of Diabetes (EASD). Diabetologia 2012, 55, 1577–1596. [Google Scholar] [CrossRef]
- Drzazga, A.; Okulus, M.; Rychlicka, M.; Biegała, Ł.; Gliszczyńska, A.; Gendaszewska-Darmach, E. Lysophosphatidylcholine Containing Anisic Acid Is Able to Stimulate Insulin Secretion Targeting G Protein Coupled Receptors. Nutrients 2020, 12, 1173. [Google Scholar] [CrossRef]
- Kang, L.; Mokshagundam, S.; Reuter, B.; Lark, D.S.; Sneddon, C.C.; Hennayake, C.; Williams, A.S.; Bracy, D.P.; James, F.D.; Pozzi, A.; et al. Integrin-Linked Kinase in Muscle Is Necessary for the Development of Insulin Resistance in Diet-Induced Obese Mice. Diabetes 2016, 65, 1590–1600. [Google Scholar] [CrossRef] [Green Version]
- Mosa, R.A.; Cele, N.D.; Mabhida, S.E.; Shabalala, S.C.; Penduka, D.; Opoku, A.R. In vivo antihyperglycemic activity of a lanosteryl triterpene from Protorhus longifolia. Molecules 2015, 20, 13374–13383. [Google Scholar] [CrossRef]
- Keedy, D.A.; Hill, Z.B.; Biel, J.T.; Kang, E.; Rettenmaier, T.J.; Brandão-Neto, J.; Pearce, N.M.; von Delft, F.; Wells, J.A.; Fraser, J.S. An expanded allosteric network in PTP1B by multitemperature crystallography, fragment screening, and covalent tethering. eLife 2018, 7, e36307. [Google Scholar] [CrossRef]
- Wu, C.; Zhao, W.; Zhang, X.; Chen, X. Neocryptotanshinone inhibits lipopolysaccharide-induced inflammation in RAW264.7 macrophages by suppression of NF-κB and iNOS signaling pathways. Acta Pharm. Sin. B 2015, 5, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.R.; Wu, W.L.; Chang, W.L.; Lin, H.C.; King, M.L. Isolation and bioactivity of new tanshinones. J. Nat. Prod. 1987, 50, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Lehnen, A.M.; Rodrigues, B.; Irigoyen, M.C.; De Angelis, K.; Schaan, B.D.A. Cardiovascular changes in animal models of metabolic syndrome. J. Diabetes Res. 2013, 2013, 761314. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Bradley, D.; Blaszczak, A.; Yin, Z.; Liu, J.; Joseph, J.J.; Wright, V.; Anandani, K.; Needleman, B.; Noria, S.; Renton, D.; et al. Clusterin Impairs Hepatic Insulin Sensitivity and Adipocyte Clusterin Associates With Cardiometabolic Risk. Diabetes Care 2019, 42, 466–475. [Google Scholar] [CrossRef] [Green Version]
- Saadeldeen, F.S.A.; Niu, Y.; Wang, H.; Zhou, L.; Meng, L.; Chen, S.; Sun-Waterhouse, D.; Waterhouse, G.I.N.; Liu, Z.; Kang, W. Natural products: Regulating glucose metabolism and improving insulin resistance. Food Sci. Hum. Wellness 2020, 9, 214–228. [Google Scholar] [CrossRef]
- Liu, H.-W.; Chen, Y.-J.; Chang, Y.-C.; Chang, S.-J. Oligonol, a Low-Molecular Weight Polyphenol Derived from Lychee, Alleviates Muscle Loss in Diabetes by Suppressing Atrogin-1 and MuRF1. Nutrients 2017, 9, 1040. [Google Scholar] [CrossRef]
- Jeong, S.-Y.; Kang, S.; Kim, D.S.; Park, S. Codonopsis lanceolata Water Extract Increases Hepatic Insulin Sensitivity in Rats with Experimentally-Induced Type 2 Diabetes. Nutrients 2017, 9, 1200. [Google Scholar] [CrossRef] [Green Version]
- Lees, E.K.; Krol, E.; Shearer, K.; Mody, N.; Gettys, T.W.; Delibegovic, M. Effects of hepatic protein tyrosine phosphatase 1B and methionine restriction on hepatic and whole-body glucose and lipid metabolism in mice. Metabolism 2015, 64, 305–314. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.-Y.; Sim, X.; Wu, Y.; Liang, J.; Tabara, Y.; Hu, C.; Hara, K.; Tam, C.H.T.; Cai, Q.; Zhao, Q.; et al. Genome-wide association meta-analysis identifies novel variants associated with fasting plasma glucose in East Asians. Diabetes 2015, 64, 291–298. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Takeuchi, F.; Lee, J.-Y.; Li, H.; Wu, J.-Y.; Liang, J.; Long, J.; Tabara, Y.; Goodarzi, M.O.; Pereira, M.A.; et al. Multiple nonglycemic genomic loci are newly associated with blood level of glycated hemoglobin in East Asians. Diabetes 2014, 63, 2551–2562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Wu, B.; Ward, M.G.; Chong, A.C.N.; Mukherjee, S.; Chen, S.; Hao, M. Multifunctional in vivo imaging of pancreatic islets during diabetes development. J. Cell Sci. 2016, 129, 2865–2875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buzova, D.; Maugeri, A.; Liguori, A.; Napodano, C.; Lo Re, O.; Oben, J.; Alisi, A.; Gasbarrini, A.; Grieco, A.; Cerveny, J.; et al. Circulating histone signature of human lean metabolic-associated fatty liver disease (MAFLD). Clin. Epigenetics 2020, 12, 126. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Luo, W.; Wang, J.; Guo, C.; Wang, X.; Wolffe, S.L.; Bodary, P.F.; Eitzman, D.T. Obesity-induced endothelial dysfunction is prevented by deficiency of P-selectin glycoprotein ligand-1. Diabetes 2012, 61, 3219–3227. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Han, Y.; Kwon, E.-Y.; Choi, M.-S. d-allulose Ameliorates Metabolic Dysfunction in C57BL/KsJ-db/db Mice. Molecules 2020, 25, 3656. [Google Scholar] [CrossRef]
- Stull, A.J.; Wang, Z.Q.; Zhang, X.H.; Yu, Y.; Johnson, W.D.; Cefalu, W.T. Skeletal muscle protein tyrosine phosphatase 1B regulates insulin sensitivity in African Americans. Diabetes 2012, 61, 1415–1422. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Tian, J.; Li, J.; Huang, L.; Wu, S.; Liang, W.; Zhong, L.; Ye, J.; Ye, F. A novel protein tyrosine phosphatase 1B inhibitor with therapeutic potential for insulin resistance. Br. J. Pharmacol. 2016, 173, 1939–1949. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.D.; Schwarzer, M.; Schrepper, A.; Amorim, P.A.; Blum, D.; Hain, C.; Faerber, G.; Haendeler, J.; Altschmied, J.; Doenst, T. Increased Protein Tyrosine Phosphatase 1B (PTP1B) Activity and Cardiac Insulin Resistance Precede Mitochondrial and Contractile Dysfunction in Pressure-Overloaded Hearts. J. Am. Heart Assoc. 2018, 7, e008865. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Liu, M.; Chen, Y.; Ma, C.; Liu, L.; Zhao, B.; Wang, Y.; Li, X.; Zhu, Y.; Gao, X.; et al. NaoXinTong Capsules inhibit the development of diabetic nephropathy in db/db mice. Sci. Rep. 2018, 8, 9158. [Google Scholar] [CrossRef]
- Caruso, M.; Ma, D.; Msallaty, Z.; Lewis, M.; Seyoum, B.; Al-janabi, W.; Diamond, M.; Abou-Samra, A.B.; Højlund, K.; Tagett, R.; et al. Increased interaction with insulin receptor substrate 1, a novel abnormality in insulin resistance and type 2 diabetes. Diabetes 2014, 63, 1933–1947. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Zhang, S.; Yang, C.; Li, Q.; Wang, S.; Xu, X.; Hao, J.; Li, C. A Novel PTP1B Inhibitor-Phosphate of Polymannuronic Acid Ameliorates Insulin Resistance by Regulating IRS-1/Akt Signaling. Int. J. Mol. Sci. 2021, 22, 12693. [Google Scholar] [CrossRef] [PubMed]
- Salem, H.F.; Nafady, M.M.; Ali, A.A.; Khalil, N.M.; Elsisi, A.A. Evaluation of Metformin Hydrochloride Tailoring Bilosomes as an Effective Transdermal Nanocarrier. Int. J. Nanomed. 2022, 17, 1185–1201. [Google Scholar] [CrossRef] [PubMed]
- Abd-El Hafeez, S.I.; Eleraky, N.E.; Hafez, E.; Abouelmagd, S.A. Design and optimization of metformin hydrophobic ion pairs for efficient encapsulation in polymeric drug carriers. Sci. Rep. 2022, 12, 5737. [Google Scholar] [CrossRef]
- Silva, T.S.; Silva, D.A.K.; Nogueira, A.L. Metformin hydrochloride sustained release biopolymeric system composed by PLLA-CMC microparticles. J. Appl. Polym. Sci. 2021, 138, 50806. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Free Energy (kcal/mol) | |||
|---|---|---|---|---|
| Site 1 | Site 2 | Site 3 | Site 4 | |
| 1,2-Dihydrotanshiiquinone | −6.1 | −5.2 | −6.0 | −5.4 |
| NCTS | −6.2 | −5.9 | −5.3 | −5.1 |
| Dehydromiltirone | −5.4 | −5.3 | −5.2 | −5.2 |
| Tanshinonel | −6.1 | −5.9 | −5.3 | −5.6 |
| Danshen Spiroketallactone | −6.0 | −5.5 | −5.2 | −5.1 |
| Neotanshinone | −5.6 | −5.3 | −5.3 | −5.2 |
| Organs (%) | m/m | db/db | db/db + NCTS (10 mg/kg) | db/db + Met (100 mg/kg) |
|---|---|---|---|---|
| Liver | 4.046 ± 0.279 | 6.866 ± 0.184 ### | 5.925 ± 0.687 ** | 5.874 ± 0.282 ** |
| Pancreas | 1.218 ± 0.073 | 0.319 ± 0.047 ### | 0.525 ± 0.037 *** | 0.585 ± 0.072 *** |
| Heart | 0.517 ± 0.025 | 0.254 ± 0.022 ### | 0.355 ± 0.023 *** | 0.332 ± 0.031 *** |
| Spleen | 0.244 ± 0.019 | 0.136 ± 0.012 ### | 0.147 ± 0.028 | 0.132 ± 0.015 |
| Kidney | 1.255 ± 0.016 | 0.793 ± 0.067 ### | 1.020 ± 0.076 *** | 0.864 ± 0.075 |
| Lung | 0.537 ± 0.025 | 0.245 ± 0.021 ### | 0.330 ± 0.017 *** | 0.307 ± 0.043 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, J.; Qian, Z.; Liu, Z.; Zhang, G.; Wang, D.; Han, W. The Antidiabetic Activities of Neocryptotanshinone: Screened by Molecular Docking and Related to the Modulation of PTP1B. Nutrients 2022, 14, 3031. https://doi.org/10.3390/nu14153031
Hao J, Qian Z, Liu Z, Zhang G, Wang D, Han W. The Antidiabetic Activities of Neocryptotanshinone: Screened by Molecular Docking and Related to the Modulation of PTP1B. Nutrients. 2022; 14(15):3031. https://doi.org/10.3390/nu14153031
Chicago/Turabian StyleHao, Jie, Zhiming Qian, Zijian Liu, Guirong Zhang, Di Wang, and Weiwei Han. 2022. "The Antidiabetic Activities of Neocryptotanshinone: Screened by Molecular Docking and Related to the Modulation of PTP1B" Nutrients 14, no. 15: 3031. https://doi.org/10.3390/nu14153031
APA StyleHao, J., Qian, Z., Liu, Z., Zhang, G., Wang, D., & Han, W. (2022). The Antidiabetic Activities of Neocryptotanshinone: Screened by Molecular Docking and Related to the Modulation of PTP1B. Nutrients, 14(15), 3031. https://doi.org/10.3390/nu14153031