
Therapeutic Role of Curcumin in Diabetes: An Analysis Based on Bioinformatic Findings



Abstract
:1. Introduction
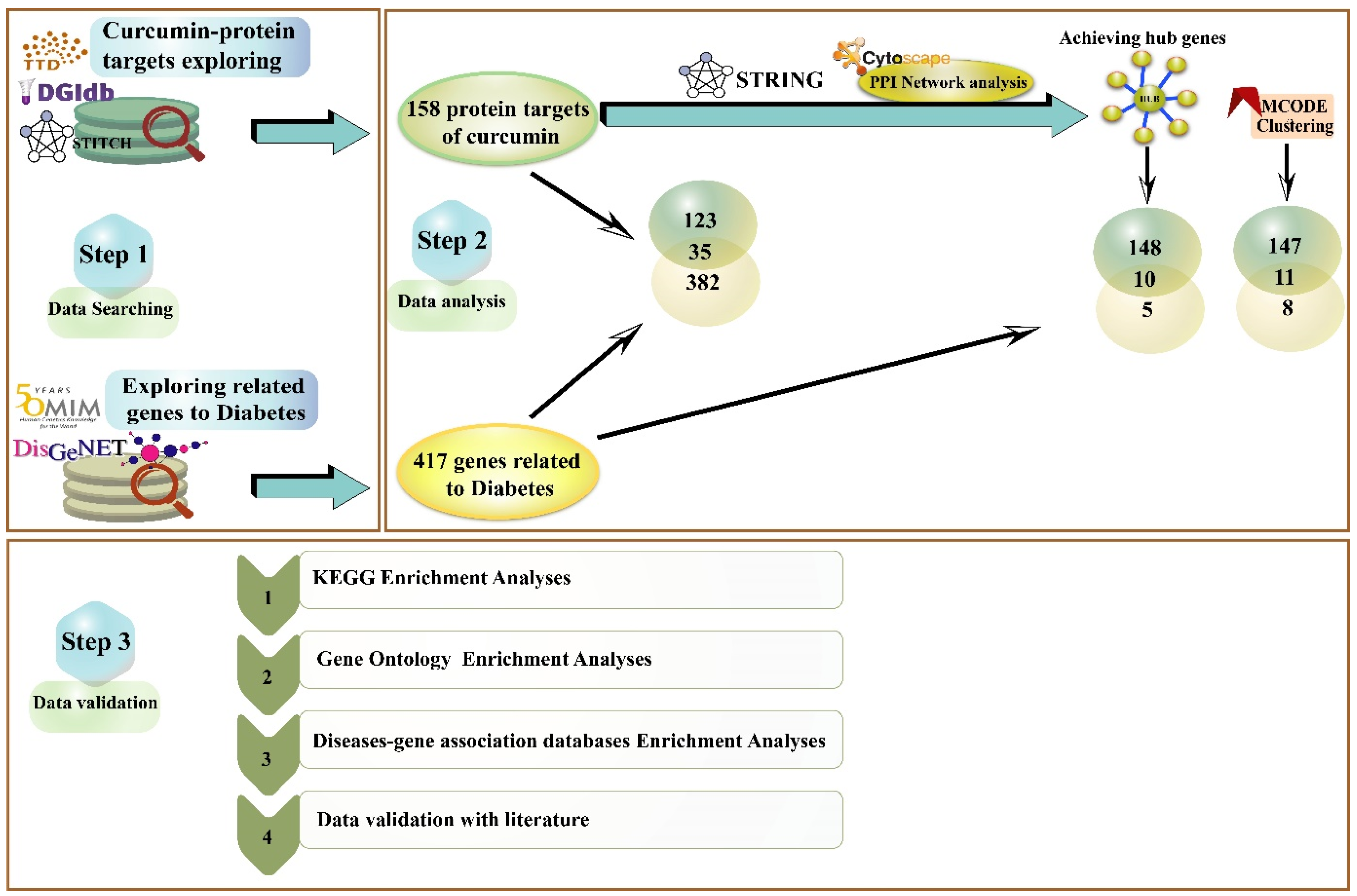
2. Methods
2.1. Curcumin Targets Exploring
2.2. Exploring Critical Diabetes-Related Genes in OMIM and DisGeNET Databases
2.3. Protein-Protein Interaction (PPI) Network
2.4. Evaluate Curcumin against Protein Targets of Diabetes
2.5. Biological Pathways and Process Enrichment Analysis
2.6. Validation of Shared Genes in Diseases-Genes Databases
3. Results
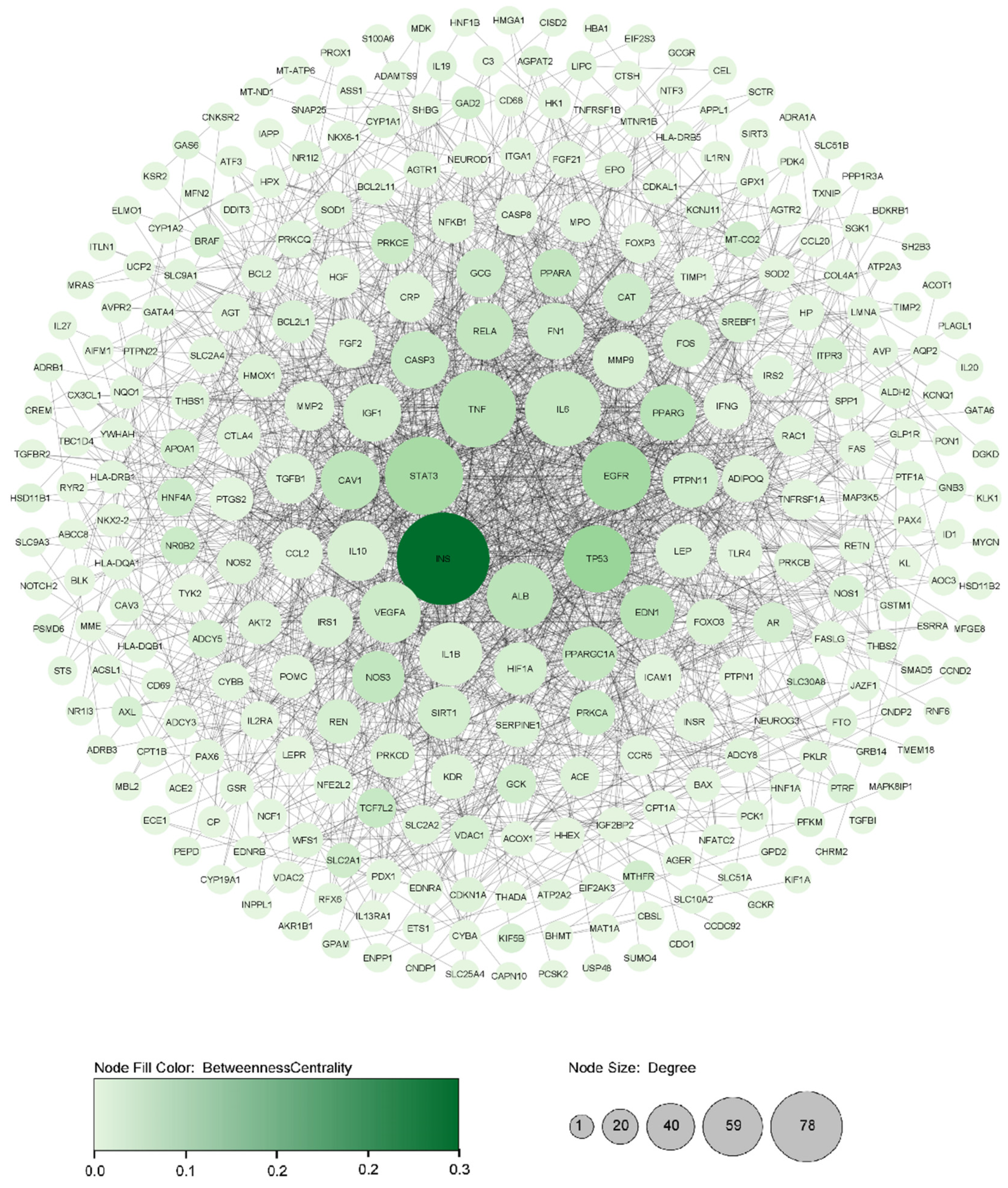
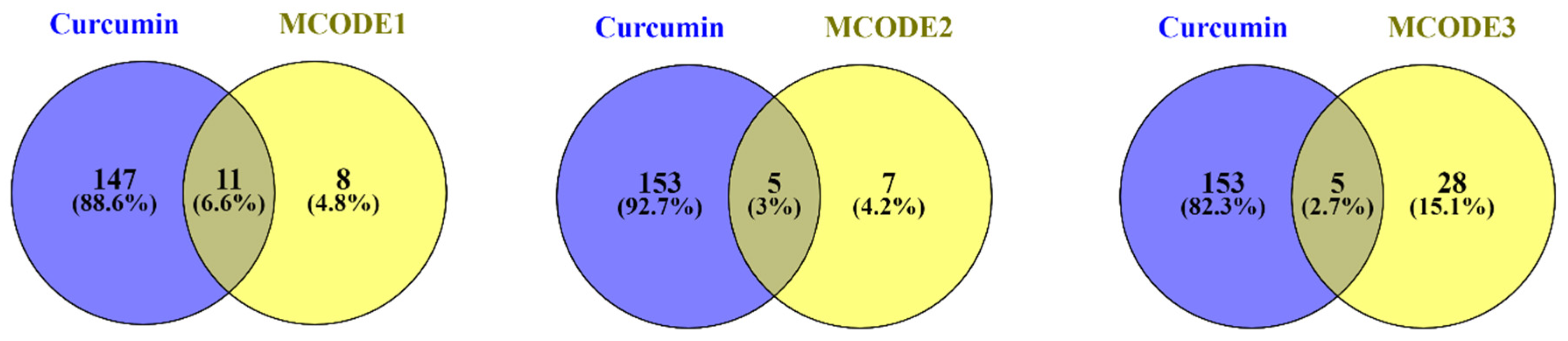
3.1. Assembly and Analyzing PPI Network
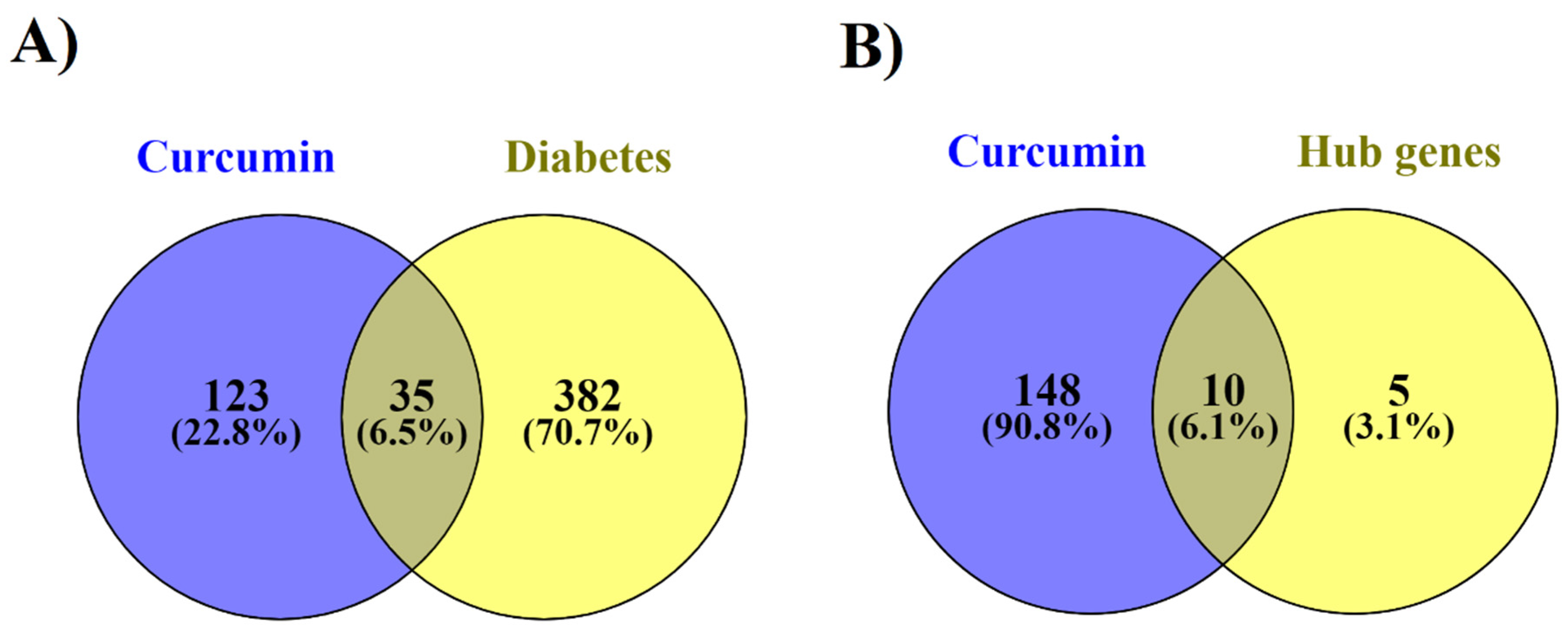
3.2. Curcumin and their Possible Targets in Diabetes
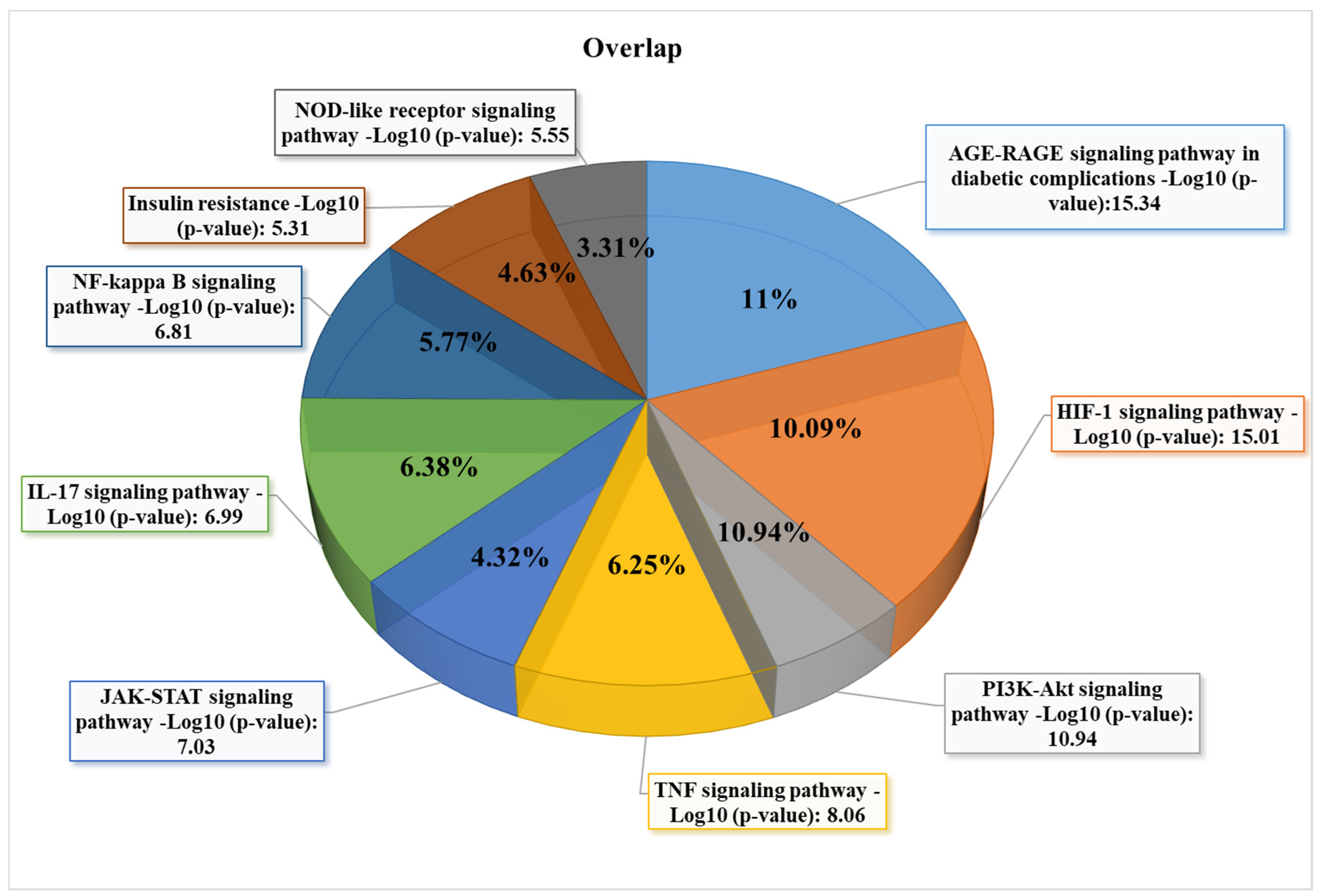
3.3. GO and KEGG Enrichment Analyses of Shared Proteins
3.4. Enrichment Analysis of the Different Types of Diabetes in Gene-Diseases Database
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Acronyms | Full Name |
| IDF | International Diabetes Federation |
| VLDL | Very low density lipoprotein |
| TTD | Therapeutic target database |
| DGIdb | Drug-gene interaction database |
| OMIM | Online Mendelian Inheritance in Man |
| DisGeNET | Gene-disease Associations |
| PPI | Protein-Protein interaction |
| STRING | Search Tool For The Retrieval Of Interacting Genes |
| STITCH | Search Tool For Interactions Of Chemicals |
| MCODE | Molecular Complex Detection |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| HIF-1 | Hypoxia-inducible Factor-1 |
| TNF | Tumor Necrosis Factor |
| JAK-STAT | Janus Kinase-signal Transducer And Activator Of Transcription |
| ROS | Reactive oxygen species |
| AGE-RAGE | Advanced glycation end-Receptor for Advanced Glycation End |
| PKC | Protein Kinase C |
| MAPK | Mitogen-Activated Protein Kinase |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| TGF-β | Transforming Growth Factor-β |
| ERK1/2 | Extracellular Signal-Regulated Kinases 1/2 |
| SOD-1 | Superoxide Dismutase-1 |
| VSMCs | Vascular Smooth Muscle Cells |
| HSC | Hepatic stellate cells |
| SGLT2 | Sodium-glucose Cotransporter 2 |
| GLUT4 | Glucose transporter 4 membrane translocation |
| STZ | Streptozotocin |
| STAT3 | Signal Transducer And Activator Of Transcription 3 |
| HG | High glucose |
| EGFR | Epidermal growth factor receptor |
| ErbB | Erythroblastosis Oncogene B |
| IRS1 | Insulin receptor substrate-1 |
| T2D | Type 2 diabetes |
| NQO1 | NAD(P)H Quinone Dehydrogenase 1 |
| INS | Insulin |
| TP53 | Tumor protein p53 |
| EGFR | Epidermal growth factor receptor |
| PPARG | Peroxisome proliferator activated receptor gamma |
| ALB | Albumin |
| CAV1 | Caveolin 1 |
| RELA | RELA proto-oncogene, NF-kB subunit |
| IL6 | Interleukin 6 |
| CASP3 | Caspase 3 |
| VEGFA | Vascular endothelial growth factor A |
| NOS3 | Nitric oxide synthase 3 |
| PPARA | Peroxisome proliferator activated receptor alpha |
| FN1 | Fibronectin 1 |
| THBS1 | Thrombospondin 1 |
| TLR4 | toll like receptor 4 |
| FOXO3 | forkhead box O3 |
| HMOX1 | heme oxygenase 1 |
| ICAM1 | intercellular adhesion molecule 1 |
| IL1B | interleukin 1 beta |
| MMP2 | matrix metallopeptidase 2(MMP2) |
| BCL2 | BCL2 apoptosis regulator( |
| CDKN1A | cyclin dependent kinase inhibitor 1A |
References
- Wang, H.; Li, N.; Chivese, T.; Werfalli, M.; Sun, H.; Yuen, L.; Hoegfeldt, C.A.; Elise Powe, C.; Immanuel, J.; Karuranga, S.; et al. IDF Diabetes Atlas: Estimation of Global and Regional Gestational Diabetes Mellitus Prevalence for 2021 by International Association of Diabetes in Pregnancy Study Group’s Criteria. Diabetes Res. Clin. Pract. 2022, 183, 109050. [Google Scholar] [CrossRef]
- American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care 2014, 37 (Suppl. S1), S81–S90. [Google Scholar] [CrossRef] [Green Version]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Patho-physiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef]
- American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care 2013, 36 (Suppl. S1), S67–S74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genuth, S.M.; Palmer, J.P.; Nathan, D.M. Classification and Diagnosis of Diabetes. In Diabetes in America; Cowie, C.C., Casagrande, S.S., Menke, A., Cissell, M.A., Eberhardt, M.S., Meigs, J.B., Gregg, E.W., Knowler, W.C., Barrett-Connor, E., Becker, D.J., et al., Eds.; National Institute of Diabetes and Digestive and Kidney Diseases: Bethesda, MD, USA, 2018. [Google Scholar]
- Centers for Disease Control and Prevention. Prevention. National Diabetes Statistics Report: Estimates of Diabetes and Its Burden in the United States; Department of Health and Human Services: Atlanta, GA, USA, 2014. [Google Scholar]
- Javed Shaikh, M.A.; Roshan, S.; Singh, H.; Rawat, S.; Pathak, S.; Mishra, A.; Gupta, G. Role of Various Gene Expressions in Etiopathogenesis of Type 2 Diabetes Mellitus. Adv. Mind-Body Med. 2021, 35, 31–39. [Google Scholar] [PubMed]
- Marucci, A.; Rutigliano, I.; Fini, G.; Pezzilli, S.; Menzaghi, C.; Di Paola, R.; Trischitta, V. Role of Actionable Genes in Pursuing a True Approach of Precision Medicine in Monogenic Diabetes. Genes 2022, 13, 117. [Google Scholar] [CrossRef] [PubMed]
- Miao, C.; Chen, H.; Li, Y.; Guo, Y.; Xu, F.; Chen, Q.; Zhang, Y.; Hu, M.; Chen, G. Curcumin and its analog alleviate diabetes-induced damages by regulating inflammation and oxidative stress in brain of diabetic rats. Diabetol. Metab. Syndr. 2021, 13, 21. [Google Scholar] [CrossRef] [PubMed]
- Sanidad, K.Z.; Sukamtoh, E.; Xiao, H.; McClements, D.J.; Zhang, G. Curcumin: Recent Advances in the Development of Strategies to Improve Oral Bioavailability. Annu. Rev. Food Sci. Technol. 2019, 10, 597–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmoudi, A.; Kesharwani, P.; Majeed, M.; Teng, Y.; Sahebkar, A. Recent advances in nanogold as a promising nanocarrier for curcumin delivery. Colloids Surf. B Biointerfaces 2022, 215, 112481. [Google Scholar] [CrossRef]
- Bagherniya, M.; Nobili, V.; Blesso, C.N.; Sahebkar, A. Medicinal plants and bioactive natural compounds in the treatment of non-alcoholic fatty liver disease: A clinical review. Pharmacol. Res. 2018, 130, 213–240. [Google Scholar] [CrossRef] [PubMed]
- Farhood, B.; Mortezaee, K.; Goradel, N.H.; Khanlarkhani, N.; Salehi, E.; Nashtaei, M.S.; Najafi, M.; Sahebkar, A. Curcumin as an anti-inflammatory agent: Implications to radiotherapy and chemotherapy. J. Cell. Physiol. 2019, 234, 5728–5740. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, A.; Cicero, A.F.G.; Panahi, Y.; Mohajeri, M.; Sahebkar, A. Curcumin: A naturally occurring autophagy modulator. J. Cell. Physiol. 2019, 234, 5643–5654. [Google Scholar] [CrossRef] [PubMed]
- Afshari, A.R.; Jalili-Nik, M.; Abbasinezhad-Moud, F.; Javid, H.; Karimi, M.; Mollazadeh, H.; Jamialahmadi, T.; Sathyapalan, T.; Sahebkar, A. Anti-tumor effects of curcuminoids in glioblastoma multiforme: An updated literature review. Curr. Med. Chem. 2021, 28, 8116–8138. [Google Scholar] [CrossRef] [PubMed]
- Gorabi, A.M.; Kiaie, N.; Hajighasemi, S.; Jamialahmadi, T.; Majeed, M.; Sahebkar, A. The effect of curcumin on the differentiation of mesenchymal stem cells into mesodermal lineage. Molecules 2019, 24, 4029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidari, Z.; Daei, M.; Boozari, M.; Jamialahmadi, T.; Sahebkar, A. Curcumin supplementation in pediatric patients: A systematic review of current clinical evidence. Phytother. Res. 2022, 36, 1442–1458. [Google Scholar] [CrossRef]
- Shishodia, S. Molecular mechanisms of curcumin action: Gene expression. BioFactors 2013, 39, 37–55. [Google Scholar] [CrossRef]
- Mahmoudi, A.; Butler, A.E.; Majeed, M.; Banach, M.; Sahebkar, A. Investigation of the Effect of Curcumin on Protein Targets in NAFLD Using Bioinformatic Analysis. Nutrients 2022, 14, 1331. [Google Scholar] [CrossRef]
- Radbakhsh, S.; Momtazi-Borojeni, A.A.; Mahmoudi, A.; Sarborji, M.R.; Hatamipour, M.; Moallem, S.A.; Atkin, S.L.; Sahebkar, A. Investigation of the Effects of Difluorinated Curcumin on Glycemic Indices in Streptozotocin-Induced Diabetic Rats. In Natural Products and Human Diseases: Pharmacology, Molecular Targets, and Therapeutic Benefits; Sahebkar, A., Sathyapalan, T., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 131–141. [Google Scholar] [CrossRef]
- Panahi, Y.; Khalili, N.; Sahebi, E.; Namazi, S.; Reiner, Ž.; Majeed, M.; Sahbekar, A. Curcuminoids modify lipid profile in type 2 diabetes mellitus: A randomized controlled trial. Complementary Ther. Med. 2017, 33, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Parsamanesh, N.; Moossavi, M.; Bahrami, A.; Butler, A.E.; Sahebkar, A. Therapeutic potential of curcumin in diabetic complications. Pharmacol. Res. 2018, 136, 181–193. [Google Scholar] [CrossRef]
- Marton, L.T.; Pescinini-e-Salzedas, L.M.; Camargo, M.E.C.; Barbalho, S.M.; Haber, J.F.D.S.; Sinatora, R.V.; Detregiachi, C.R.P.; Girio, R.J.S.; Buchaim, D.V.; Cincotto dos Santos Bueno, P. The Effects of Curcumin on Diabetes Mellitus: A Systematic Review. Front. Endocrinol. 2021, 12, 669448. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, A.; Butler, A.E.; Jamialahmadi, T.; Sahebkar, A. Target Deconvolution of Fenofibrate in Nonalcoholic Fatty Liver Disease Using Bioinformatics Analysis. BioMed Res. Int. 2021, 2021, 3654660. [Google Scholar] [CrossRef]
- Mahmoudi, A.; Heydari, S.; Markina, Y.V.; Barreto, G.E.; Sahebkar, A. Role of statins in regulating molecular pathways following traumatic brain injury: A system pharmacology study. Biomed. Pharmacother. 2022, 153, 113304. [Google Scholar] [CrossRef] [PubMed]
- Likić, V.A.; McConville, M.J.; Lithgow, T.; Bacic, A. Systems Biology: The Next Frontier for Bioinformatics. Adv. Bioinform. 2010, 2010, 268925. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef] [PubMed]
- Daffu, G.; del Pozo, C.H.; O’Shea, K.M.; Ananthakrishnan, R.; Ramasamy, R.; Schmidt, A.M. Radical roles for RAGE in the pathogenesis of oxidative stress in cardiovascular diseases and beyond. Int. J. Mol. Sci. 2013, 14, 19891–19910. [Google Scholar] [CrossRef] [Green Version]
- Hutchinson, K.R.; Lord, C.K.; West, T.A.; Stewart, J.A., Jr. Cardiac fibroblast-dependent extracellular matrix accumulation is associated with diastolic stiffness in type 2 diabetes. PLoS ONE 2013, 8, e72080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kay, A.M.; Simpson, C.L.; Stewart, J.A., Jr. The Role of AGE/RAGE Signaling in Diabetes-Mediated Vascular Calcification. J. Diabetes Res. 2016, 2016, 6809703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, T.; Chen, X.; Chen, W.; Huang, S.; Peng, X.; Tian, L.; Wu, X.; Huang, Y. Curcumin is a Potential Adjuvant to Alleviates Diabetic Retinal Injury via Reducing Oxidative Stress and Maintaining Nrf2 Pathway Homeostasis. Front. Pharmacol. 2021, 12, 796565. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Chen, A. Curcumin eliminates the effect of advanced glycation end-products (AGEs) on the divergent regulation of gene expression of receptors of AGEs by interrupting leptin signaling. Lab. Investig. 2014, 94, 503–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, F.J.; Xie, C.; Jiang, C. The role of hypoxia-inducible factors in metabolic diseases. Nat. Rev. Endocrinol. 2018, 15, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Kim, J.H.; Li, F.; Qu, A.; Gavrilova, O.; Shah, Y.M.; Gonzalez, F.J. Hypoxia-inducible factor 1α regulates a SOCS3-STAT3-adiponectin signal transduction pathway in adipocytes. J. Biol. Chem. 2013, 288, 3844–3857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanatani, Y.; Usui, I.; Ishizuka, K.; Bukhari, A.; Fujisaka, S.; Urakaze, M.; Haruta, T.; Kishimoto, T.; Naka, T.; Kobayashi, M. Effects of pioglitazone on suppressor of cytokine signaling 3 expression: Potential mechanisms for its effects on insulin sensitivity and adiponectin expression. Diabetes 2007, 56, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Gunton, J.E.; Kulkarni, R.N.; Yim, S.; Okada, T.; Hawthorne, W.J.; Tseng, Y.H.; Roberson, R.S.; Ricordi, C.; O’Connell, P.J.; Gonzalez, F.J.; et al. Loss of ARNT/HIF1beta mediates altered gene expression and pancreatic-islet dysfunction in human type 2 diabetes. Cell 2005, 122, 337–349. [Google Scholar] [CrossRef] [Green Version]
- Cheng, K.; Ho, K.; Stokes, R.; Scott, C.; Lau, S.M.; Hawthorne, W.J.; O’Connell, P.J.; Loudovaris, T.; Kay, T.W.; Kulkarni, R.N.; et al. Hypoxia-inducible factor-1alpha regulates beta cell function in mouse and human islets. J. Clin. Investig. 2010, 120, 2171–2183. [Google Scholar] [CrossRef] [Green Version]
- Packer, M. Mechanisms Leading to Differential Hypoxia-Inducible Factor Signaling in the Diabetic Kidney: Modulation by SGLT2 Inhibitors and Hypoxia Mimetics. Am. J. Kidney Dis. 2021, 77, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Li, H.X.; Li, X.H.; Jiang, J.; Shi, P.X.; Zhang, X.G.; Tian, M. Effect of miR-26b on gestational diabetes mellitus in rats via PI3K/Akt signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 1609–1615. [Google Scholar] [CrossRef] [PubMed]
- Elghazi, L.; Rachdi, L.; Weiss, A.J.; Cras-Méneur, C.; Bernal-Mizrachi, E. Regulation of beta-cell mass and function by the Akt/protein kinase B signalling pathway. Diabetes Obes. Metab. 2007, 9 (Suppl. S2), 147–157. [Google Scholar] [CrossRef] [PubMed]
- Dickson, L.M.; Rhodes, C.J. Pancreatic beta-cell growth and survival in the onset of type 2 diabetes: A role for protein kinase B in the Akt? Am. J. Physiol. Endocrinol. Metab. 2004, 287, E192–E198. [Google Scholar] [CrossRef] [PubMed]
- Bernal-Mizrachi, E.; Fatrai, S.; Johnson, J.D.; Ohsugi, M.; Otani, K.; Han, Z.; Polonsky, K.S.; Permutt, M.A. Defective insulin secretion and increased susceptibility to experimental diabetes are induced by reduced Akt activity in pancreatic islet beta cells. J. Clin. Investig. 2004, 114, 928–936. [Google Scholar] [CrossRef] [Green Version]
- Cui, W.; Zhang, Y.; Lu, D.; Ren, M.; Yuan, G. Upregulation of p-Akt by glial cell line-derived neurotrophic factor ameliorates cell apoptosis in the hippocampus of rats with streptozotocin-induced diabetic encephalopathy. Mol. Med. Rep. 2016, 13, 543–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernal-Mizrachi, E.; Wen, W.; Stahlhut, S.; Welling, C.M.; Permutt, M.A. Islet beta cell expression of constitutively active Akt1/PKB alpha induces striking hypertrophy, hyperplasia, and hyperinsulinemia. J. Clin. Investig. 2001, 108, 1631–1638. [Google Scholar] [CrossRef]
- Ren, B.C.; Zhang, Y.F.; Liu, S.S.; Cheng, X.J.; Yang, X.; Cui, X.G.; Zhao, X.R.; Zhao, H.; Hao, M.F.; Li, M.D.; et al. Curcumin alleviates oxidative stress and inhibits apoptosis in diabetic cardiomyopathy via Sirt1-Foxo1 and PI3K-Akt signalling pathways. J. Cell. Mol. Med. 2020, 24, 12355–12367. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.H.; Zhang, S.Y.; Chen, Y.S.; Li, K.; Chen, W.B.; Liu, Y.Q. Curcumin anti-diabetic effect mainly correlates with its anti-apoptotic actions and PI3K/Akt signal pathway regulation in the liver. Food Chem. Toxicol. 2020, 146, 111803. [Google Scholar] [CrossRef] [PubMed]
- Senn, J.J.; Klover, P.J.; Nowak, I.A.; Zimmers, T.A.; Koniaris, L.G.; Furlanetto, R.W.; Mooney, R.A. Suppressor of cytokine signaling-3 (SOCS-3), a potential mediator of interleukin-6-dependent insulin resistance in hepatocytes. J. Biol. Chem. 2003, 278, 13740–13746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashili, F.; Chibalin, A.V.; Krook, A.; Zierath, J.R. Constitutive STAT3 phosphorylation contributes to skeletal muscle insulin resistance in type 2 diabetes. Diabetes 2013, 62, 457–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, C.; Huang, L.; Luo, W.; Yu, W.; Hu, X.; Guan, X.; Cai, Y.; Zou, C.; Yin, H.; Xu, Z.; et al. Inhibition of STAT3 in tubular epithelial cells prevents kidney fibrosis and nephropathy in STZ-induced diabetic mice. Cell Death Dis. 2019, 10, 848. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.D.; Kim, Y.H.; Cho, Y.M.; Kim, D.K.; Ahn, S.W.; Lee, J.M.; Chanda, D.; Shong, M.; Lee, C.H.; Choi, H.S. Metformin ameliorates IL-6-induced hepatic insulin resistance via induction of orphan nuclear receptor small heterodimer partner (SHP) in mouse models. Diabetologia 2012, 55, 1482–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reda, E.; Hassaneen, S.; El-Abhar, H.S. Novel Trajectories of Bromocriptine Antidiabetic Action: Leptin-IL-6/ JAK2/p-STAT3/SOCS3, p-IR/p-AKT/GLUT4, PPAR-γ/Adiponectin, Nrf2/PARP-1, and GLP-1. Front. Pharmacol. 2018, 9, 771. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, X.; Zeng, S.; Zhang, X.; Zhao, J.; Zhang, X.; Chen, X.; Yang, W.; Yang, Y.; Dong, Z.; et al. The natural polyphenol curcumin induces apoptosis by suppressing STAT3 signaling in esophageal squamous cell carcinoma 06 Biological Sciences 0601 Biochemistry and Cell Biology 11 Medical and Health Sciences 1112 Oncology and Carcinogenesis. J. Exp. Clin. Cancer Res. 2018, 37, 303. [Google Scholar] [CrossRef] [PubMed]
- Alexandrow, M.G.; Song, L.J.; Altiok, S.; Gray, J.; Haura, E.B.; Kumar, N.B. Curcumin: A novel Stat3 pathway inhibitor for chemoprevention of lung cancer. Eur.J. Cancer Prev. 2012, 21, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.Q.; Ahmed, E.I.; Elareer, N.; Fathima, H.; Prabhu, K.S.; Siveen, K.S.; Kulinski, M.; Azizi, F.; Dermime, S.; Ahmad, A.; et al. Curcumin-mediated apoptotic cell death in papillary thyroid cancer and cancer stem-like cells through targeting of the JAK/STAT3 signaling pathway. Int. J. Mol. Sci. 2020, 21, 438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Tian, S.; Liu, Y.; Gao, F.; Yin, Q. Inhibition of curcumin on proliferation and invasion in giant cell tumor of bone (GCTB) by targeting STAT3. Int. J. Clin. Exp. Med. 2016, 9, 21474–21481. [Google Scholar]
- Xu, X.; Zhu, Y. Curcumin inhibits human non-small cell lung cancer xenografts by targeting STAT3 pathway. Am. J. Transl. Res. 2017, 9, 3633–3641. [Google Scholar] [PubMed]
- Lu, M.; Tao, L.; Mei, W.; Luo, R.; Fu, X.; Wang, L.; Yang, W.; Liu, C. Effect of curcumin on the expression of P-STAT3 and IkB in db/db mice. J. Cent. South Univ. Med. Sci. 2014, 39, 591–597. [Google Scholar] [CrossRef]
- Hutchinson, R.A.; Adams, R.A.; McArt, D.G.; Salto-Tellez, M.; Jasani, B.; Hamilton, P.W. Epidermal growth factor receptor immunohistochemistry: New opportunities in metastatic colorectal cancer. J. Transl. Med. 2015, 13, 217. [Google Scholar] [CrossRef] [PubMed]
- Vairaktaris, E.; Goutzanis, L.; Yapijakis, C.; Vassiliou, S.; Spyridonidou, S.; Vylliotis, A.; Nkenke, E.; Lazaris, A.C.; Strantzias, P.; Patsouris, E. Diabetes enhances the expression of H-ras and suppresses the expression of EGFR leading to increased cell proliferation. Histol. Histopathol. 2009, 24, 531–539. [Google Scholar] [CrossRef]
- Kyohara, M.; Shirakawa, J.; Okuyama, T.; Togashi, Y.; Inoue, R.; Li, J.; Miyashita, D.; Terauchi, Y. Soluble EGFR, a hepatokine, and adipsin, an adipokine, are biomarkers correlated with distinct aspects of insulin resistance in type 2 diabetes subjects. Diabetol. Metab. Syndr. 2020, 12, 83. [Google Scholar] [CrossRef] [PubMed]
- Limonte, C.P.; Valo, E.; Montemayor, D.; Afshinnia, F.; Ahluwalia, T.S.; Costacou, T.; Darshi, M.; Forsblom, C.; Hoofnagle, A.N.; Groop, P.H.; et al. A Targeted Multiomics Approach to Identify Biomarkers Associated with Rapid eGFR Decline in Type 1 Diabetes. Am. J. Nephrol. 2020, 51, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, S.; Almubrad, T.; Bron, A.J.; Yousif, M.H.M.; Benter, I.F.; Akhtar, S. Role of epidermal growth factor receptor (EGFR) in corneal remodelling in diabetes. Acta Ophthalmol. 2009, 87, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, Z.; Xiao, X.; Han, L.; Wu, Z.; Ma, Q.; Cao, L. Curcumin attenuates hyperglycemia-driven EGF-induced invasive and migratory abilities of pancreatic cancer via suppression of the ERK and AKT pathways. Oncol. Rep. 2019, 41, 650–658. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Lee, Y.M.; Chang, G.C.; Yu, S.L.; Hsieh, W.Y.; Chen, J.J.W.; Chen, H.W.; Yang, P.C. Curcumin induces EGFR degradation in lung adenocarcinoma and modulates p38 activation in intestine: The versatile adjuvant for gefitinib therapy. PLoS ONE 2011, 6, e23756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, A.; Uechi, T.; Kenmochi, N. Guarding the ’translation apparatus’: Defective ribosome biogenesis and the p53 signaling pathway. Wiley Interdiscip. Rev. RNA 2011, 2, 507–522. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, X.; Gao, X.; Mei, Y.; Wu, M. A new role of p53 in regulating lipid metabolism. J. Mol. Cell Biol. 2013, 5, 147–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Secchiero, P.; Toffoli, B.; Melloni, E.; Agnoletto, C.; Monasta, L.; Zauli, G. The MDM2 inhibitor Nutlin-3 attenuates streptozotocin-induced diabetes mellitus and increases serum level of IL-12p40. Acta Diabetol. 2013, 50, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Armata, H.L.; Golebiowski, D.; Jung, D.Y.; Ko, H.J.; Kim, J.K.; Sluss, H.K. Requirement of the ATM/p53 tumor suppressor pathway for glucose homeostasis. Mol. Cell. Biol. 2010, 30, 5787–5794. [Google Scholar] [CrossRef] [Green Version]
- Franck, D.; Tracy, L.; Armata, H.L.; Delaney, C.L.; Jung, D.Y.; Ko, H.J.; Ong, H.; Kim, J.K.; Scrable, H.; Sluss, H.K. Glucose Tolerance in Mice is Linked to the Dose of the p53 Transactivation Domain. Endocr. Res. 2013, 38, 139–150. [Google Scholar] [CrossRef]
- Qu, L.; He, B.; Pan, Y.; Xu, Y.; Zhu, C.; Tang, Z.; Bao, Q.; Tian, F.; Wang, S. Association between polymorphisms in RAPGEF1, TP53, NRF1 and type 2 diabetes in Chinese Han population. Diabetes Res. Clin. Pract. 2011, 91, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Burgdorf, K.S.; Grarup, N.; Justesen, J.M.; Harder, M.N.; Witte, D.R.; Jørgensen, T.; Sandbæk, A.; Lauritzen, T.; Madsbad, S.; Hansen, T.; et al. Studies of the association of Arg72Pro of tumor suppressor protein p53 with type 2 diabetes in a combined analysis of 55,521 Europeans. PLoS ONE 2011, 6, e15813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonfigli, A.R.; Sirolla, C.; Testa, R.; Cucchi, M.; Spazzafumo, L.; Salvioli, S.; Ceriello, A.; Olivieri, F.; Festa, R.; Procopio, A.D.; et al. The p53 codon 72 (Arg72Pro) polymorphism is associated with the degree of insulin resistance in type 2 diabetic subjects: A cross-sectional study. Acta Diabetol. 2013, 50, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Chan, J.Y.; Wong, T.S. Curcumin exerts inhibitory effects on undifferentiated nasopharyngeal carcinoma by inhibiting the expression of miR-125a-5p. Clin. Sci. 2014, 127, 571–579. [Google Scholar] [CrossRef]
- Ciftci, G.; Aksoy, A.; cenesiz, S.; sogut, M.u.; Yarim, G.F.; Nisbet, C.; Guvenc, D.; Ertekin, A. Therapeutic role of curcumin in oxidative DNA damage caused by formaldehyde. Microsc. Res. Tech. 2015, 78, 391–395. [Google Scholar] [CrossRef]
- Patiño-Morales, C.C.; Soto-Reyes, E.; Arechaga-Ocampo, E.; Ortiz-Sánchez, E.; Antonio-Véjar, V.; Pedraza-Chaverri, J.; García-Carrancá, A. Curcumin stabilizes p53 by interaction with NAD(P)H:quinone oxidoreductase 1 in tumor-derived cell lines. Redox Biol. 2020, 28, 101320. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Full Name | Protein Class | DSI g | Score GDA | Network Analyser | ||
|---|---|---|---|---|---|---|---|
| Degree | Betweenness | Closeness | |||||
| INS | Insulin | Plasma proteins | 0.445 | 0.70 | 78 | 0.2223 | 0.5201 |
| TP53 | Tumour protein p53 | Transcription factors | 0.236 | 0.50 | 48 | 0.0746 | 0.4714 |
| EGFR | Epidermal growth factor receptor | Enzymes | 0.295 | 0.37 | 50 | 0.0632 | 0.4669 |
| STAT3 | Signal transducer and activator of transcription 3 | Transcription factors | 0.320 | 0.35 | 61 | 0.0604 | 0.4752 |
| TNF | Tumour necrosis factor | Plasma proteins | 0.231 | 0.50 | 60 | 0.0451 | 0.4845 |
| PPARG | Peroxisome proliferator-activated receptor gamma | Nuclear receptors | 0.358 | 0.50 | 35 | 0.0411 | 0.4506 |
| ALB | Albumin | Plasma proteins | 0.317 | 0.60 | 47 | 0.0406 | 0.4655 |
| CAV1 | Caveolin 1 | Transporters | 0.388 | 0.50 | 38 | 0.0387 | 0.4534 |
| RELA | RELA proto-oncogene, NF-kB subunit | Transcription factors | 0.406 | 0.50 | 36 | 0.0338 | 0.4291 |
| IL6 | Interleukin 6 | Plasma proteins | 0.248 | 0.50 | 58 | 0.0302 | 0.4744 |
| CASP3 | Caspase 3 | Enzymes | 0.351 | 0.50 | 37 | 0.0295 | 0.4439 |
| VEGFA | Vascular endothelial growth factor A | Plasma proteins | 0.266 | 0.50 | 41 | 0.0237 | 0.4562 |
| NOS3 | Nitric oxide synthase 3 | Enzymes | 0.378 | 0.40 | 30 | 0.0349 | 0.4400 |
| PPARA | Peroxisome proliferator activated receptor alpha | Nuclear receptors | 0.432 | 0.30 | 25 | 0.0337 | 0.4267 |
| FN1 | Fibronectin 1 | Plasma proteins | 0.365 | 0.40 | 35 | 0.0233 | 0.4273 |
| Shared Protein Targets | STITCH-Score | Action |
|---|---|---|
| TP53 | 0.962 | Activation/inhibition |
| EGFR | 0.987 | inhibition |
| STAT3 | 0.959 | inhibition |
| PPARG | 0.957 | Activation |
| IL6 | 0.869 | inhibition |
| CASP3 | 0.959 | Activation/inhibition |
| VEGFA | 0.868 | inhibition |
| NOS3 | 0.820 | Activation/inhibition |
| PPARA | 0.866 | Activation |
| FN1 | 0.844 | inhibition |
| Jensen Diseases | ||
|---|---|---|
| Diseases | Adj. p-Value | Gene Name |
| Diabetic retinopathy | 8.32 × 10−8 | IL6; NOS3; AKR1B1; ICAM1; VEGFA |
| Diabetes mellitus (1,2) | 5.67 × 10−5 | LEP; STAT3; PPARG; SLC2A4 |
| Type 2 diabetes mellitus | 0.027 | IL1B; PPARG; VEGFA |
| GWAS Catalog | ||
| Type 2 diabetes | 0.01278 | LEP; STAT3; PPARG; VEGFA; BCL2 |
| DisGeNET | ||
| Diseases | Adj. p-value | Gene Name |
| Diabetes Mellitus, Non-Insulin-Dependent | 2.33 × 10−31 | CDKN1A; AKR1B1; SLC2A4; PTGS2; HIF1A; EGFR; ICAM1; CASP3; HMOX1; CCL2; GSTM1; NOS2; NOS3; MMP2; STAT3; FN1; MMP9; VEGFA; AR; IL6; IL1B; LEP; DDIT3; CYP1A2; BCL2; CYP1A1; IAPP; PPARG; PPARA; TP53; TLR4; BCL2L1; NFE2L2 |
| Diabetes Mellitus, Insulin-Dependent | 1.56 × 10−27 | AKR1B1; SLC2A4; PTGS2; EGFR; ICAM1; CASP3; HMOX1; CCL2; GSTM1; NOS2; NOS3; STAT3; FN1; MMP9; VEGFA; AR; IL6; IL1B; LEP; DDIT3; BCL2; IAPP; PPARG; TP53; TLR4; BCL2L1 |
| Diabetic Nephropathy | 1.73 × 10−29 | CDKN1A; AKR1B1; PTGS2; HIF1A; THBS1; EGFR; ICAM1; CCL2; GSTM1; NOS2; NOS3; MMP2; STAT3; FN1; MMP9; VEGFA; IL6; IL1B; LEP; BCL2; IAPP; PPARG; PPARA; TLR4; NFE2L2 |
| Diabetic Retinopathy | 5.79 × 10−26 | GSTM1; NOS2; NOS3; MMP2; FN1; AKR1B1; PTGS2; HIF1A; THBS1; MMP9; ICAM1; VEGFA; IL6; CASP3; CCL2; PPARG; PPARA; TLR4; NFE2L2 |
| Gestational Diabetes | 2.02 × 10−12 | AR; IL6; NOS3; IL1B; LEP; CCL2; PPARG; VEGFA |
| Prediabetes syndrome | 5.82 × 10−9 | IL6; IAPP; PPARG; SLC2A4; TP53; TLR4 |
| Brittle diabetes | 1.54 × 10−4 | NOS3; DDIT3; STAT3 |
| OMIM Disease | ||
| Diseases | Adj. p-value | Gene Name |
| diabetes mellitus, type 2 | 0.567 × 10−3 | SLC2A4; PPARG |
| Rare Diseases GeneRIF ARCHS4 Predictions | ||
| Diseases | Adj. p-value | Gene Name |
| Diabetic mastopathy | 5.515 × 10−6 | STAT3; IL6; IL1B; TLR4; NFE2L2; PTGS2 |
| Rare Diseases AutoRIF Gene Lists | ||
| Diseases | Adj. p-value | Gene Name |
| Insulin-resistance type B | 4.197 × 10−28 | NOS2; NOS3; STAT3; SLC2A4; PTGS2; EGFR; ICAM1; IL6; CASP3; IL1B; LEP; HMOX1; CCL2; IAPP; PPARG; TLR4; NFE2L2 |
| Diabetic mastopathy | 1.58 × 10−22 | CDKN1A; STAT3; AKR1B1; FOXO3; PTGS2; HIF1A; THBS1; MMP9; ICAM1; VEGFA; IL6; CASP3; DDIT3; BCL2; CYP1A1; TP53; BCL2L1 |
| Nephrogenic diabetes insipidus | 2.45 × 10−5 | NOS2; NOS3; PTGS2; EGFR; NFE2L2 |
| Cardiomyopathy diabetes deafness | 6.12 × 10−4 | NOS3; HMOX1 |
| Maturity-onset diabetes of the young | 9.82 × 10−4 | CASP3; IAPP; PPARG |
| Neurogenic diabetes insipidus | 0.006854232 | CASP3; PTGS2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmoudi, A.; Atkin, S.L.; Nikiforov, N.G.; Sahebkar, A. Therapeutic Role of Curcumin in Diabetes: An Analysis Based on Bioinformatic Findings. Nutrients 2022, 14, 3244. https://doi.org/10.3390/nu14153244
Mahmoudi A, Atkin SL, Nikiforov NG, Sahebkar A. Therapeutic Role of Curcumin in Diabetes: An Analysis Based on Bioinformatic Findings. Nutrients. 2022; 14(15):3244. https://doi.org/10.3390/nu14153244
Chicago/Turabian StyleMahmoudi, Ali, Stephen L. Atkin, Nikita G. Nikiforov, and Amirhossein Sahebkar. 2022. "Therapeutic Role of Curcumin in Diabetes: An Analysis Based on Bioinformatic Findings" Nutrients 14, no. 15: 3244. https://doi.org/10.3390/nu14153244
APA StyleMahmoudi, A., Atkin, S. L., Nikiforov, N. G., & Sahebkar, A. (2022). Therapeutic Role of Curcumin in Diabetes: An Analysis Based on Bioinformatic Findings. Nutrients, 14(15), 3244. https://doi.org/10.3390/nu14153244