
Effect of Type 2 Diabetes and Impaired Glucose Tolerance on Digestive Enzymes and Glucose Absorption in the Small Intestine of Young Rats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and the Type 2 Diabetes Model
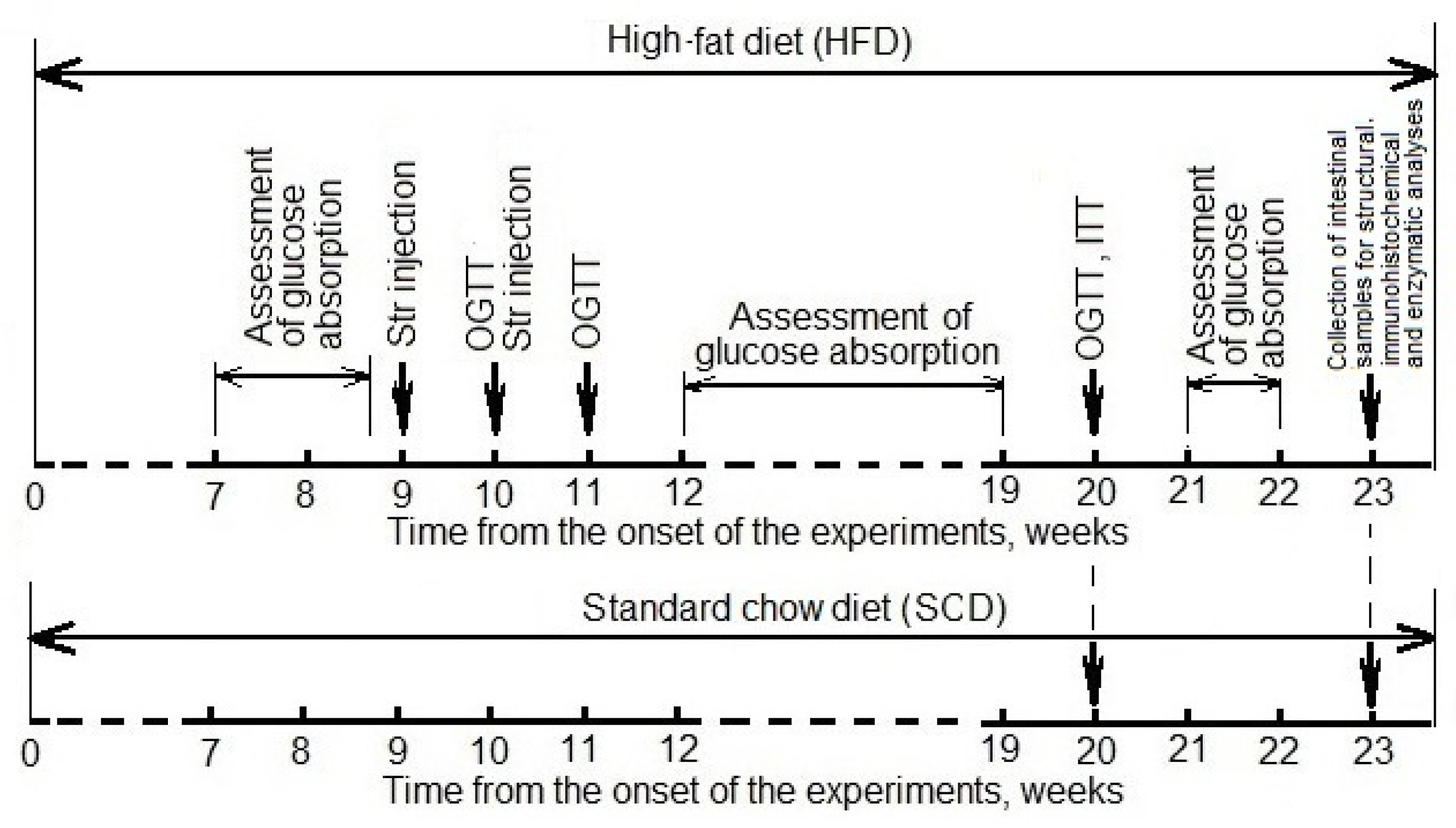
2.2. Experimental Design
2.3. Intestinal Glucose Absorption
2.4. Oral Glucose Tolerance Test (OGTT) and Insulin Tolerance Test (ITT)
2.5. Collection of Intestinal Tissues from Various Areas and Contents of the Colon
2.6. The Count of Enterocytes and Goblet Cells
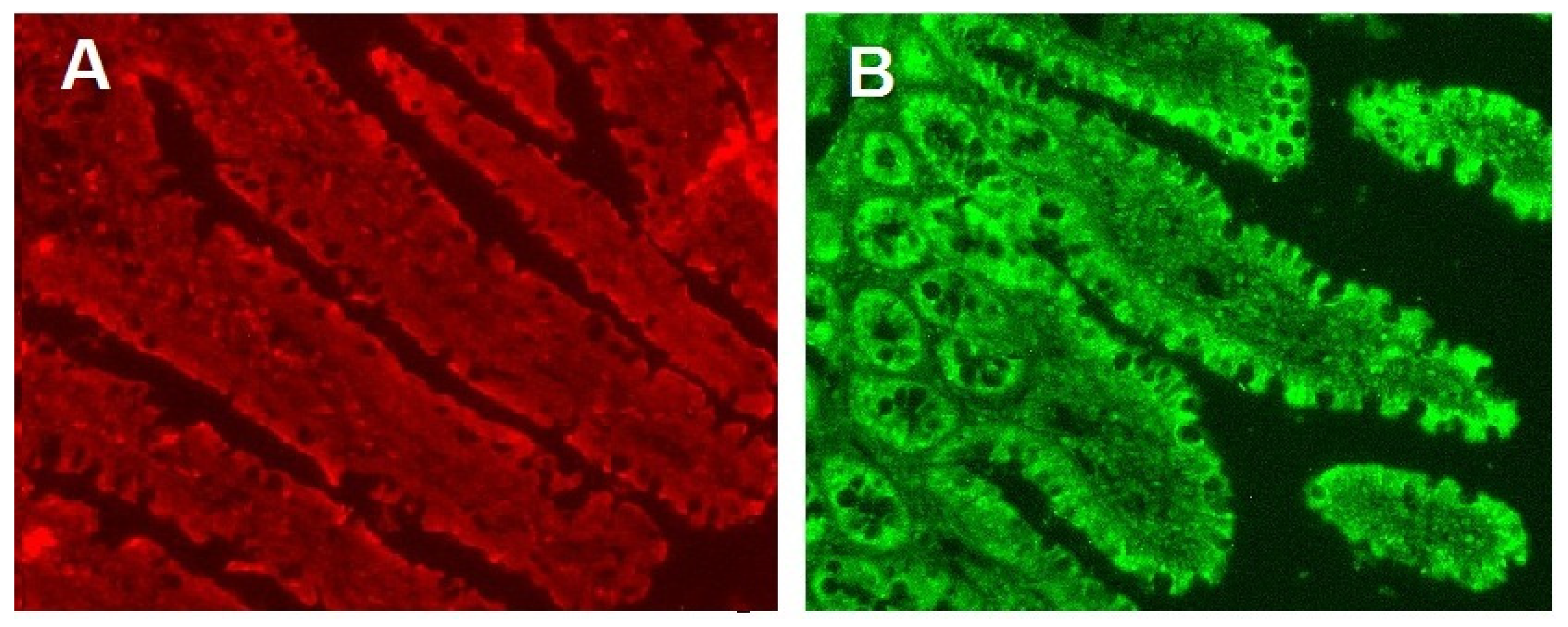
2.7. Determination of the Content of Glucose Transporters in Enterocytes
2.8. Determination of the Activity of Intestinal Digestive Enzymes
2.9. Statistical Analysis
3. Results
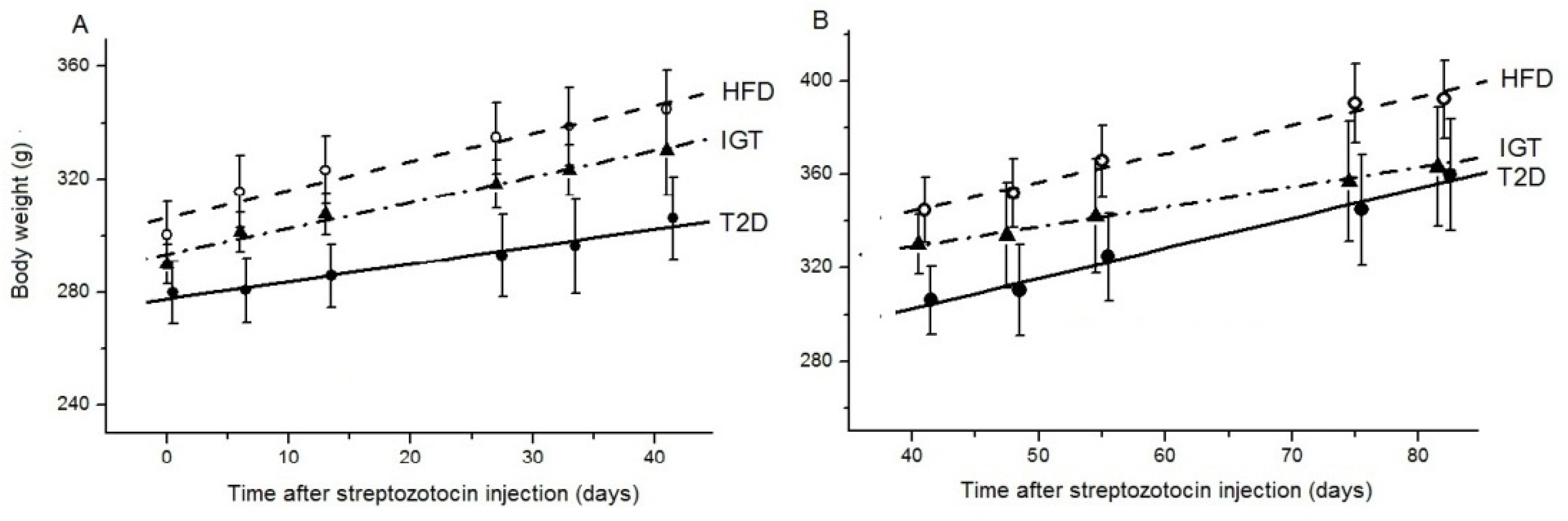
3.1. Rat Body Weight
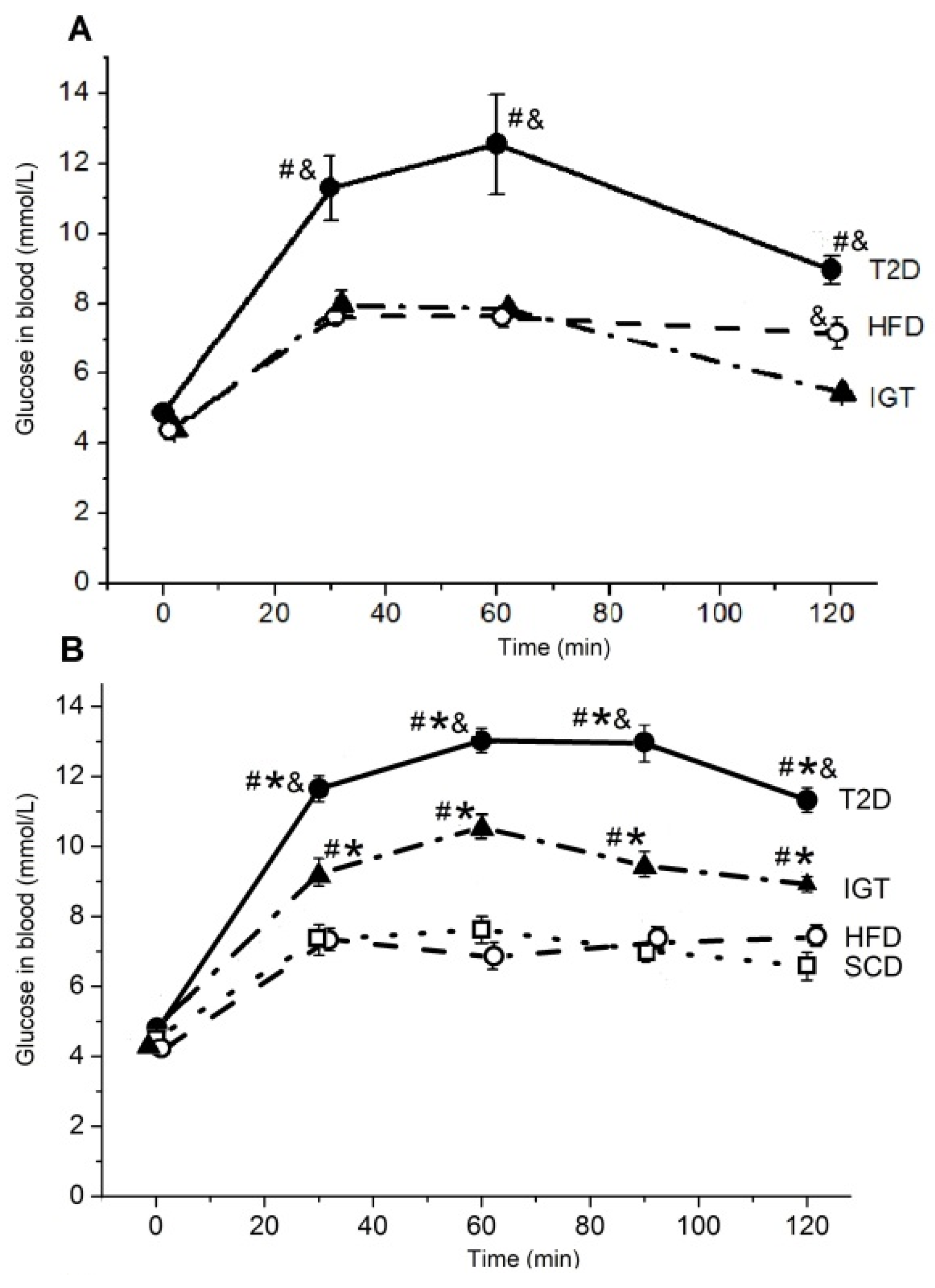
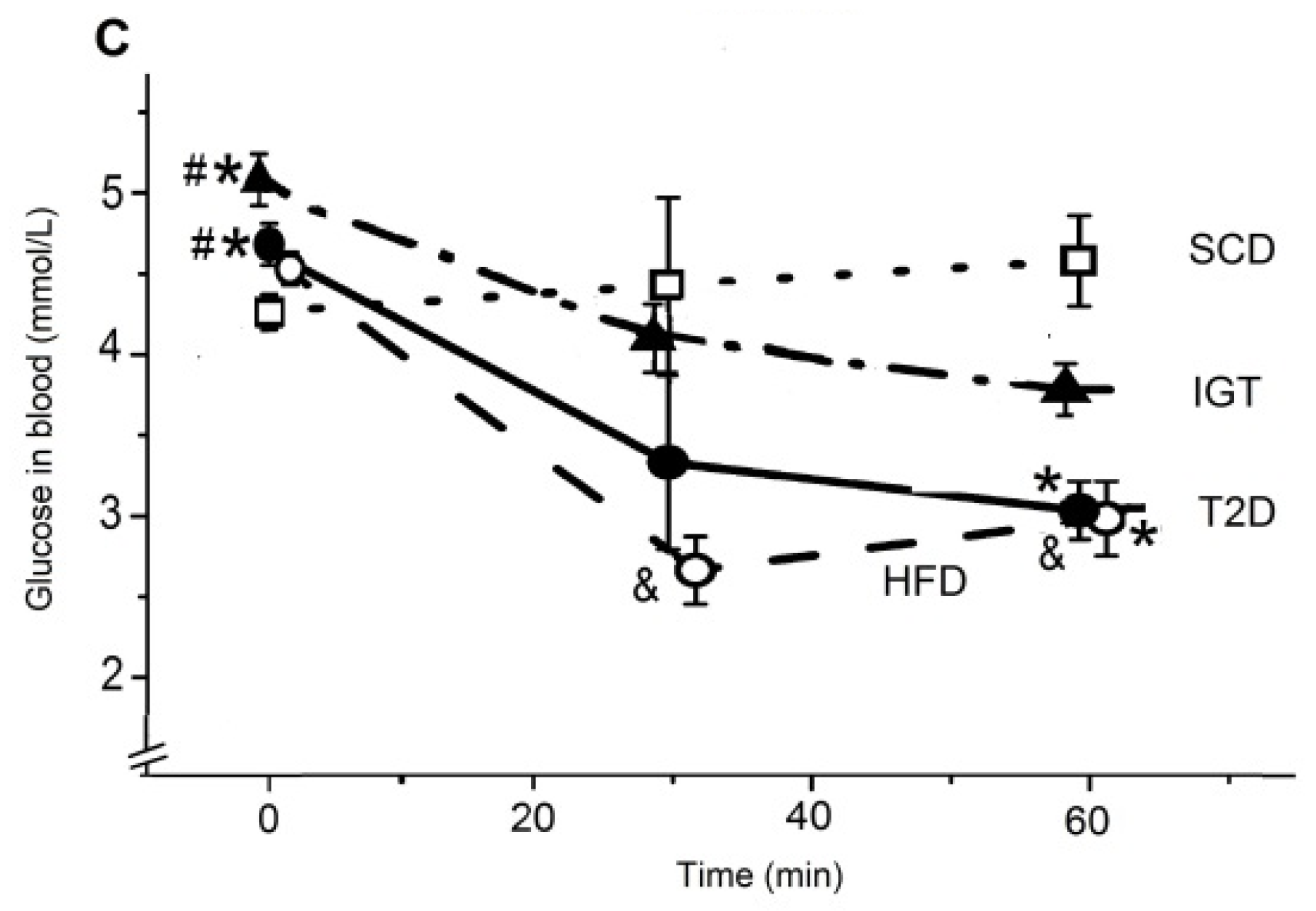
3.2. The Oral Glucose Tolerance and the Insulin Tolerance Tests in Rats
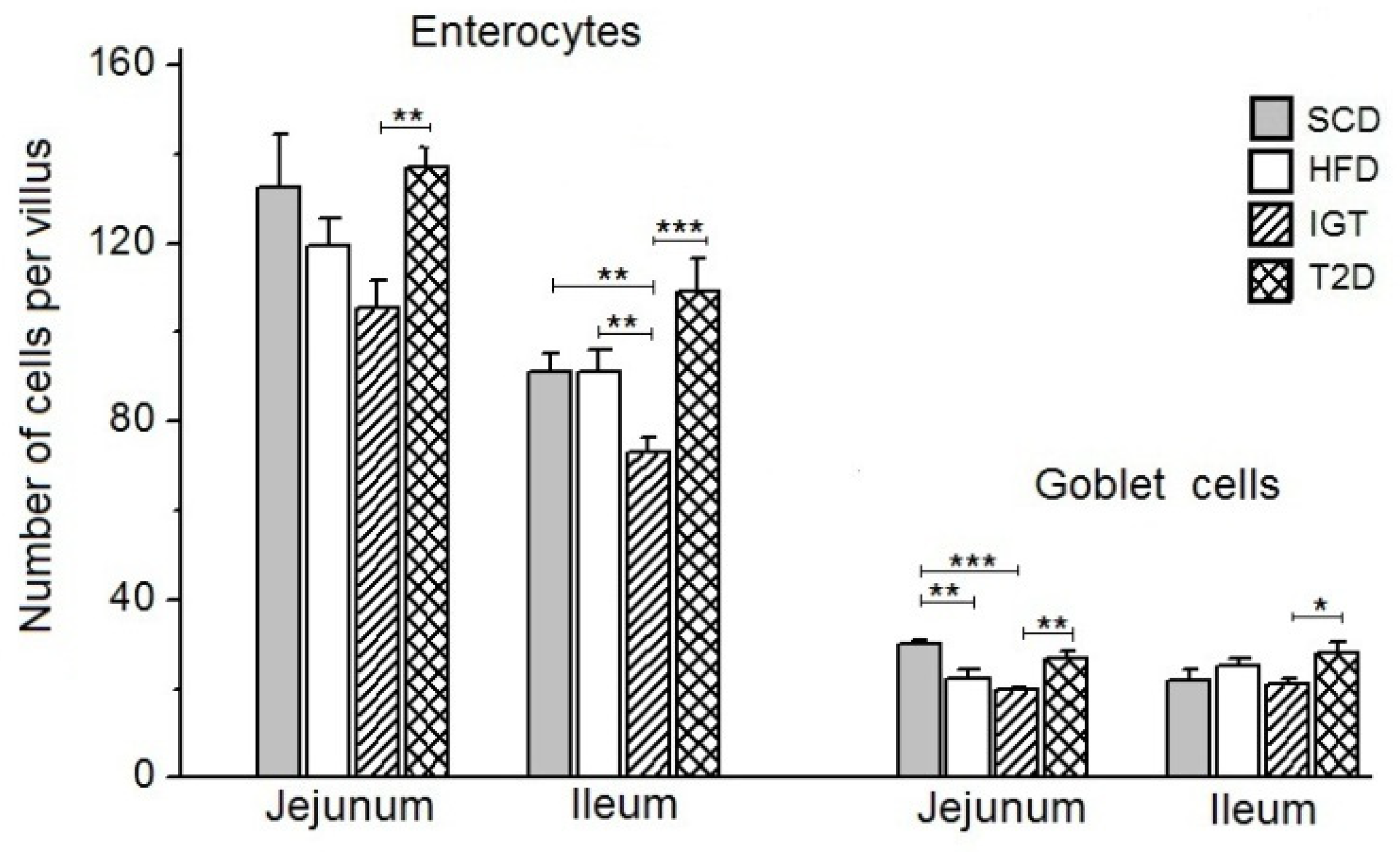
3.3. The Number of Enterocytes and Goblet Cells on the Villi of the Jejunum and Ileum
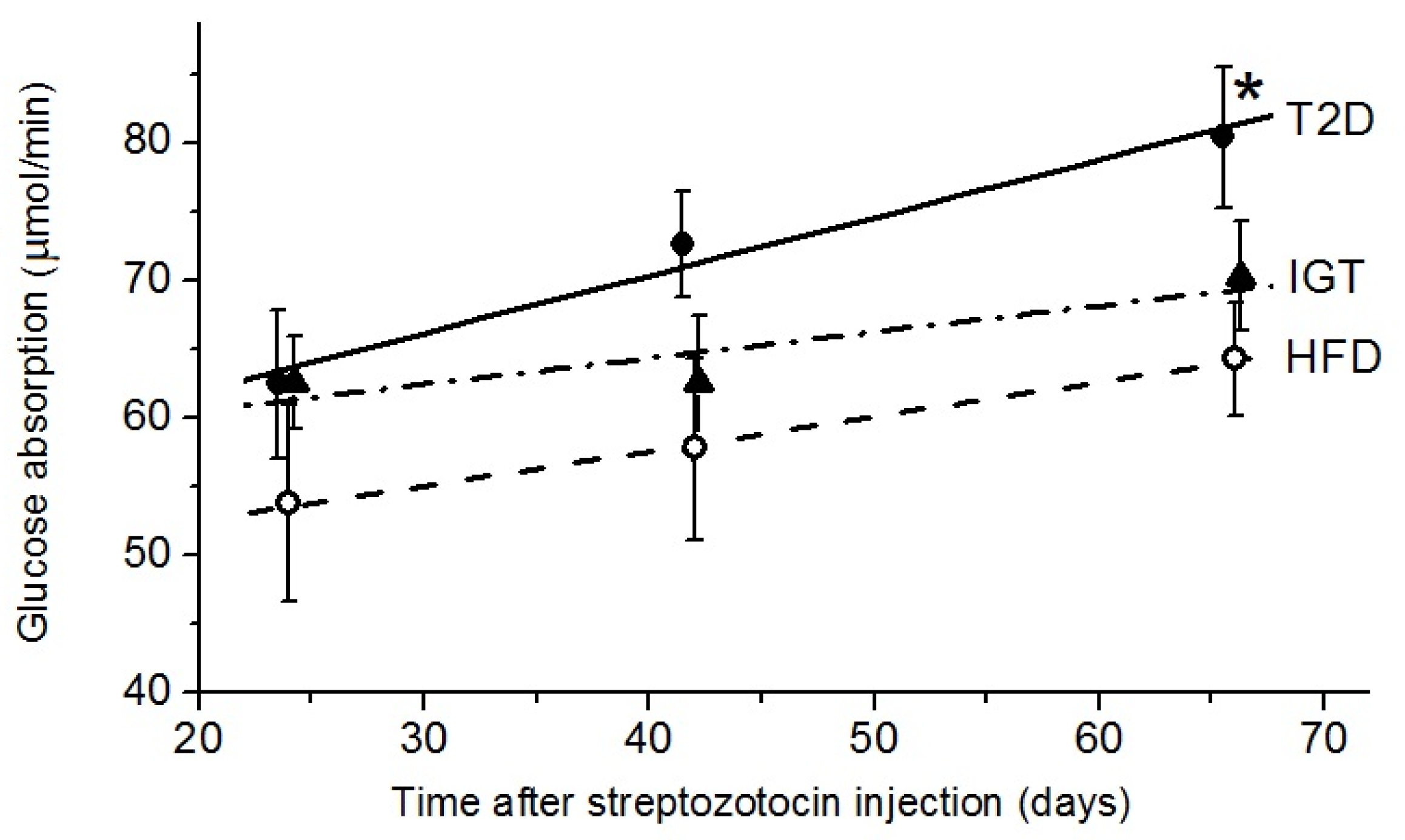
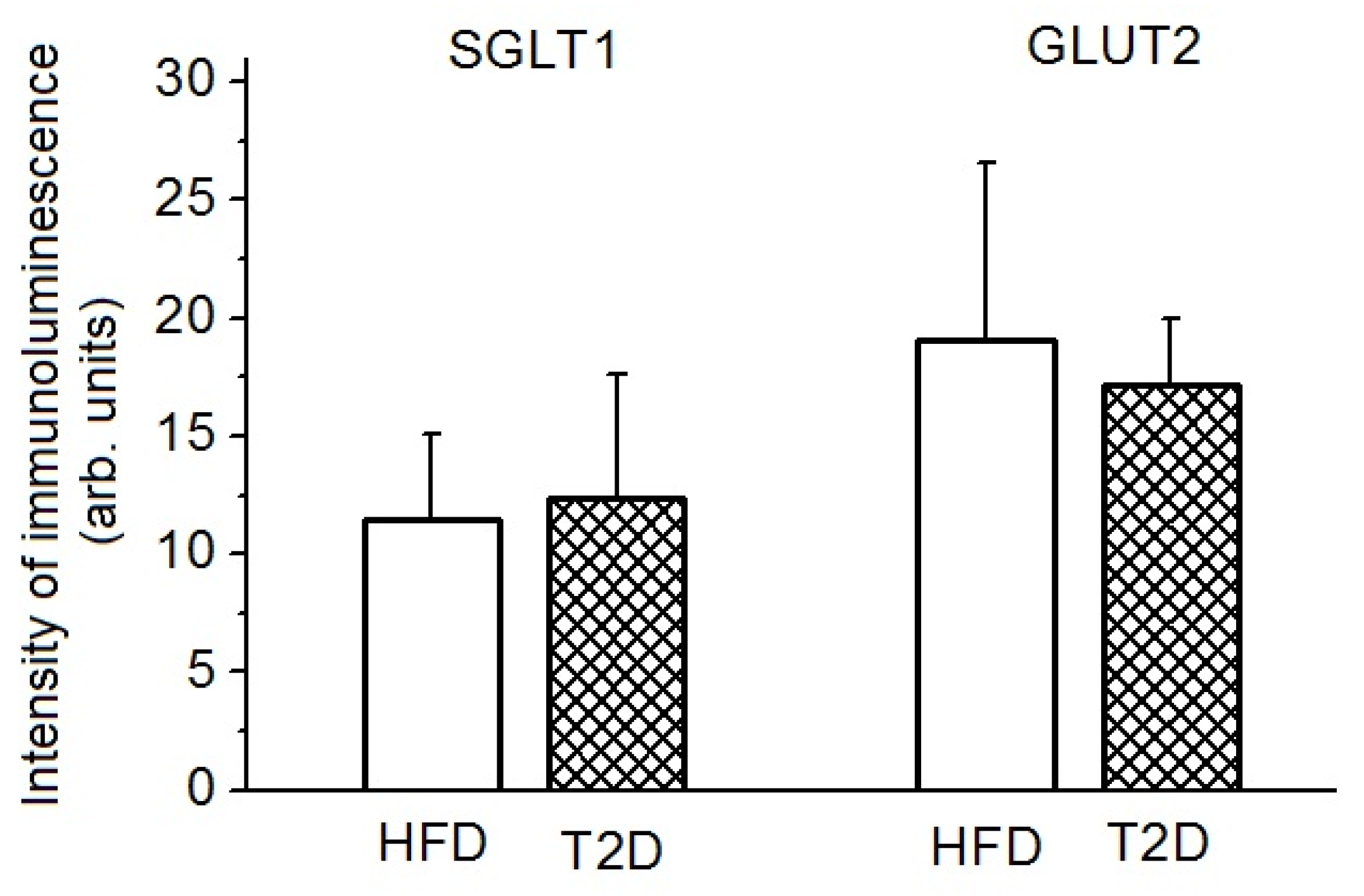
3.4. Glucose Absorption, SGLT1 and GLUT2 Transporters
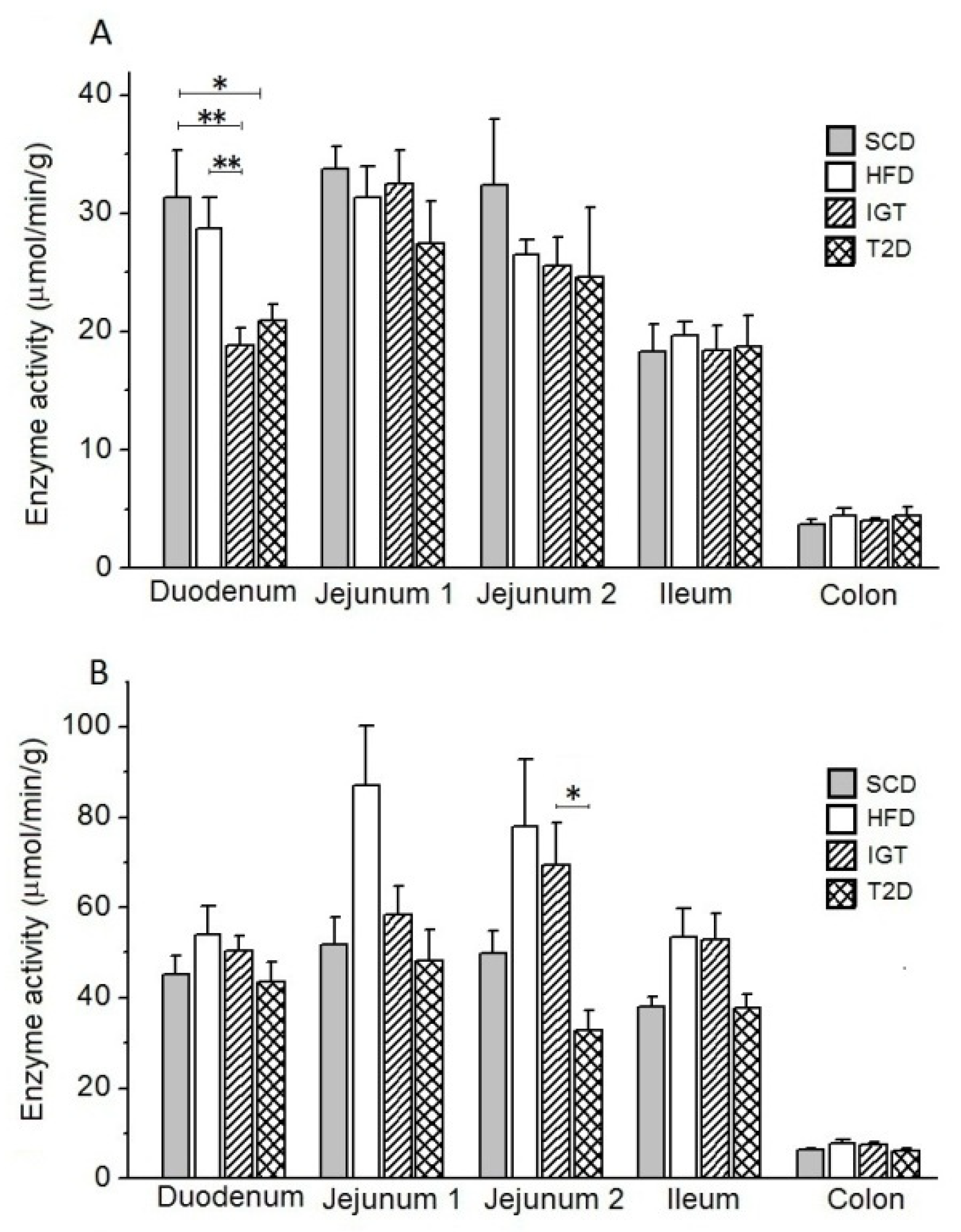
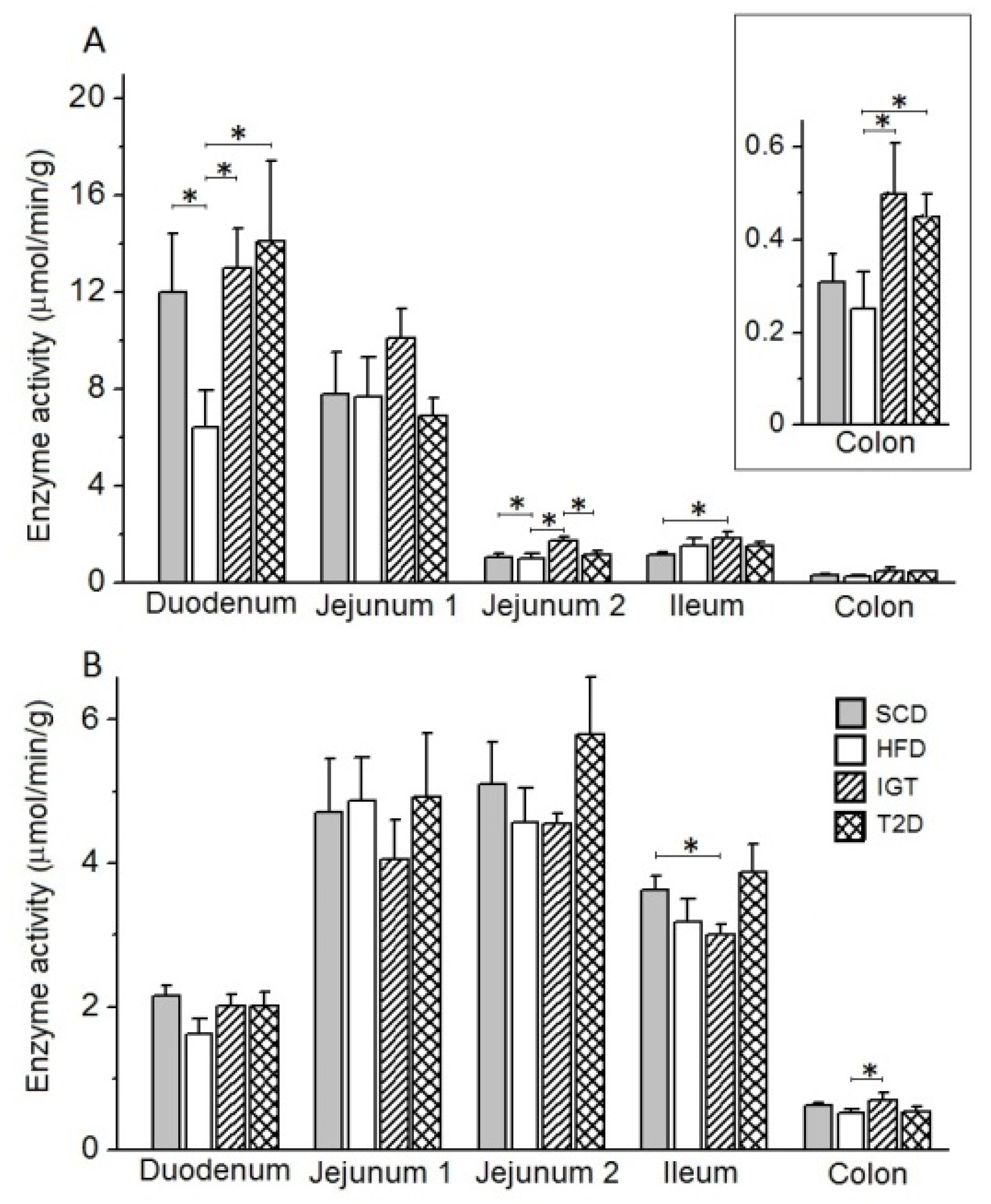
3.5. Intestinal Enzyme Activities
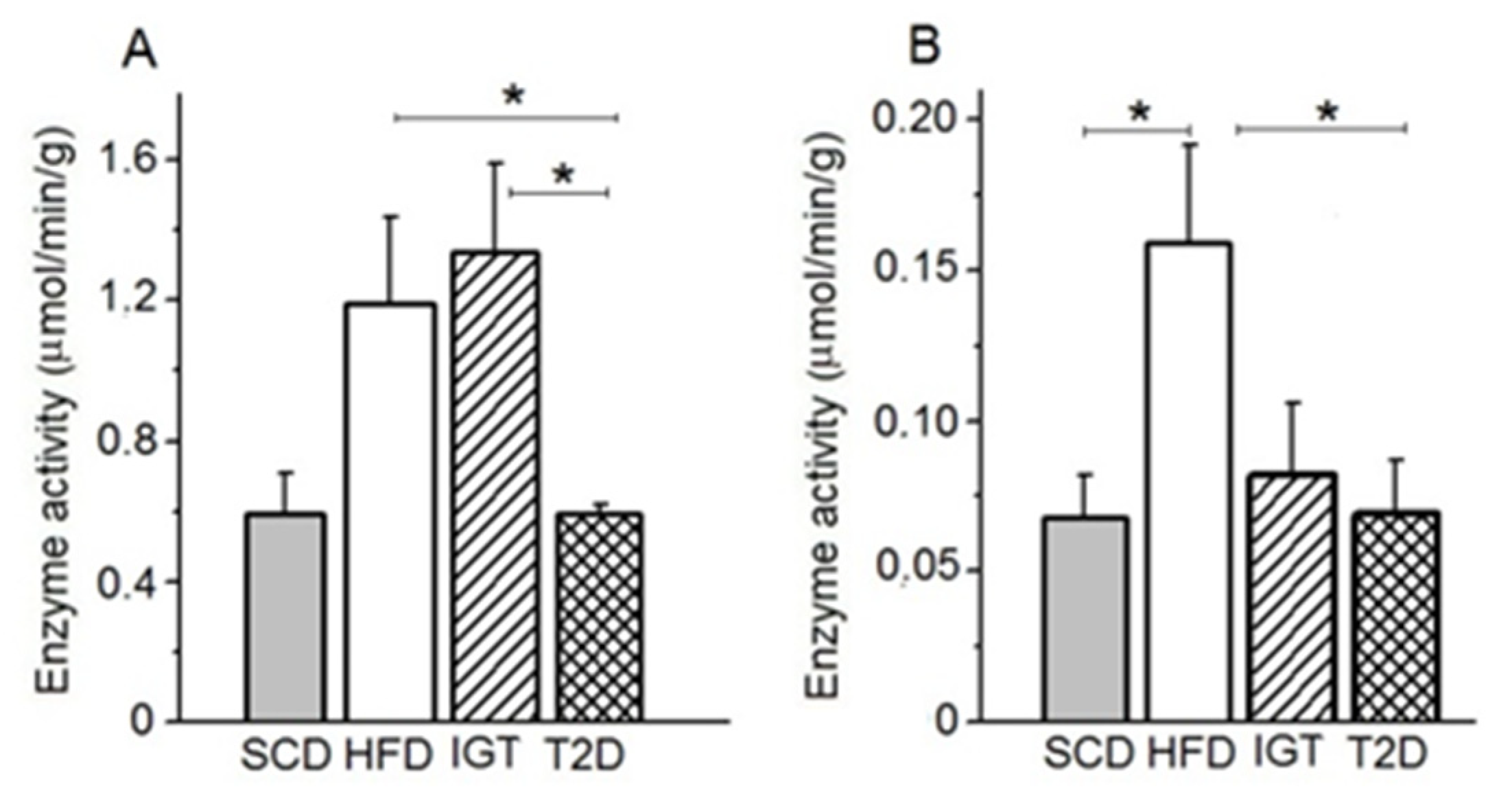
3.6. AP and APN Activities in Colon Content
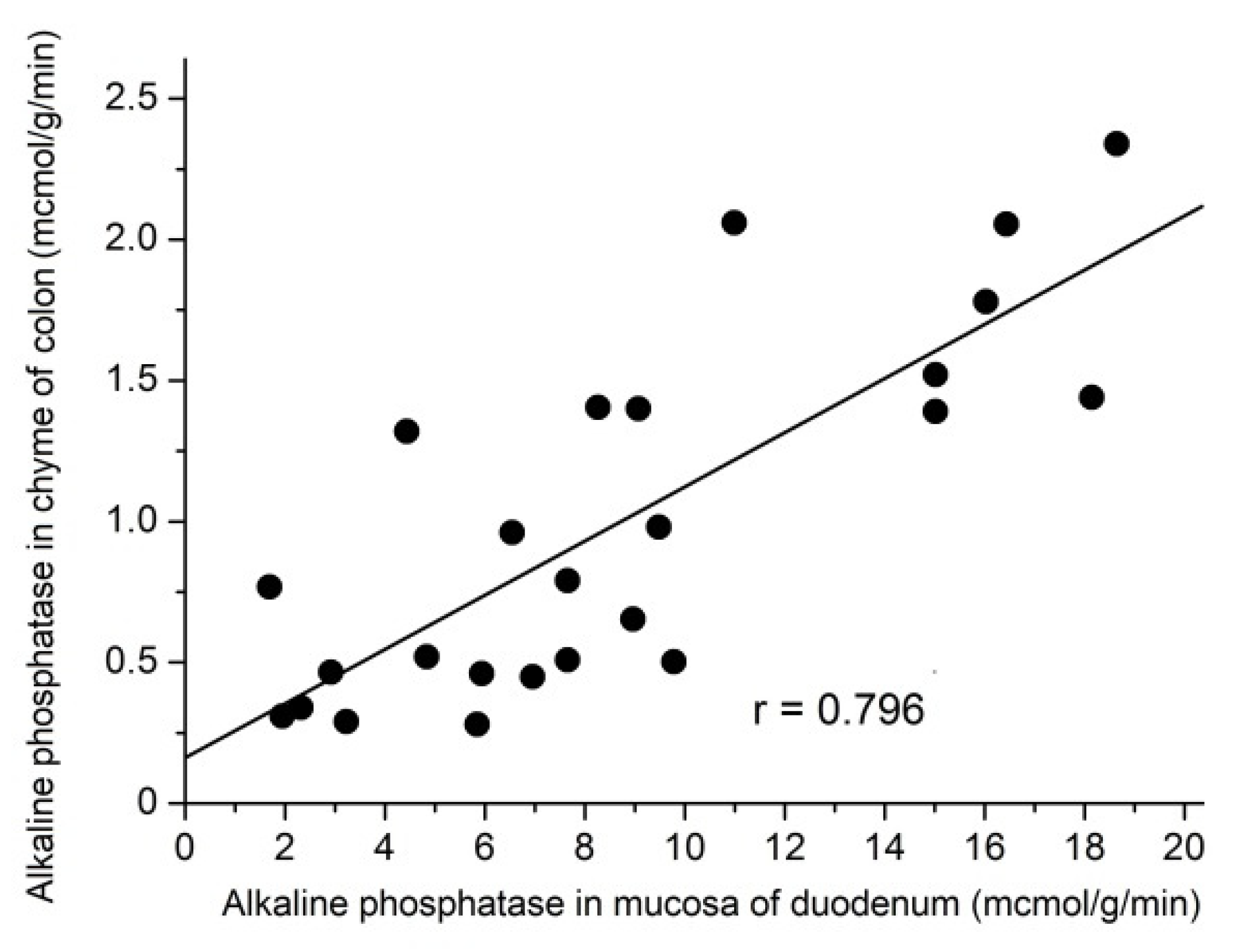
3.7. Relationship between the AP Activity in the Chyme of the Colon and its Activity in the Mucosa of the Duodenum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Elder, D.A.; Hornung, L.N.; Herbers, P.M.; Prigeon, R.; Woo, J.G.; D’Alessio, D.A. Rapid deterioration of insulin secretion in obese adolescents preceding the onset of type 2 diabetes. J. Pediatr. 2015, 166, 672–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valaiyapathi, B.; Gower, B.; Ashraf, A.P. Pathophysiology of Type 2 Diabetes in Children and Adolescents. Curr. Diabetes Rev. 2020, 16, 220–229. [Google Scholar] [CrossRef]
- Baud, G.; Raverdy, V.; Bonner, C.; Daoudi, M.; Caiazzo, R.; Pattou, F. Sodium glucose transport modulation in type 2 diabetes and gastric bypass surgery. Surg. Obes. Relat. Dis. 2016, 12, 1206–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, A.; Hornby, P.J. Intestinal SGLT1 in metabolic health and disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G887–G898. [Google Scholar] [CrossRef] [Green Version]
- Khawandanah, J. Double or Hybrid Diabetes: A Systematic Review on Disease Prevalence, Characteristics and Risk Factors. Nutr. Diabetes 2019, 9, 33. [Google Scholar] [CrossRef] [PubMed]
- Skovsø, S. Modeling type 2 diabetes in rats using high fat diet and streptozotocin. J. Diabetes Investig. 2014, 5, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Kojima, H.; Hidaka, H.; Fujimiya, M.; Kashiwagi, A.; Kikkawa, R. Increased intestinal glucose absorption and postprandial hyperglycaemia at the early step of glucose intolerance in Otsuka Long-Evans Tokushima Fatty rats. Diabetologia 1998, 41, 1459–1466. [Google Scholar] [CrossRef] [Green Version]
- Corpe, C.; Sreenan, S.; Burant, C. Effects of type-2 diabetes and troglitazone on the expression patterns of small intestinal sugar transporters and PPAR-gamma in the Zucker diabetic fatty rat. Digestion 2001, 63, 116–123. [Google Scholar] [CrossRef]
- Chukwuma, C.I.; Islam, M.S. Effects of xylitol on carbohydrate digesting enzymes activity, intestinal glucose absorption and muscle glucose uptake: A multi-mode study. Food Funct. 2015, 6, 955–962. [Google Scholar] [CrossRef]
- Burant, C.F.; Flink, S.; DePaoli, A.M.; Chen, J.; Lee, W.S.; Hediger, M.A.; Buse, J.B.; Chang, E.B. Small Intestine Hexose Transport in Experimental Diabetes. Increased Transporter mRNA and Protein Expression in Enterocytes. J. Clin. Investig. 1994, 93, 578–585. [Google Scholar] [CrossRef] [Green Version]
- Debnam, E.S.; Smith, M.V.; Sharp, P.A.; Srai, S.K.; Turvey, A.; Keable, S.J. The effects of streptozotocin diabetes on sodium-glucose transporter (SGLT1) expression and function in rat jejunal and ileal villus-attached enterocytes. Pflügers Arch. 1995, 430, 151–159. [Google Scholar] [CrossRef]
- Dyer, J.; Wood, I.S.; Palejwala, A.; Ellis, A.; Shirazi-Beechey, S.P. Expression of monosaccharide transporters in intestine of diabetic humans. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 282, G241–G248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koepsell, H. Glucose transporters in the small intestine in health and disease. Pflugers Arch. 2020, 472, 1207–1248. [Google Scholar] [CrossRef]
- Gorboulev, V.; Schurmann, A.; Volker, V.; Kipp, H.; Jaschke, A.; Klessen, D.; Friedrich, A.; Scherneck, S.; Rieg, T.; Cunard, R.; et al. Na(+)-D-glucose Cotransporter SGLT1 is pivotal for intestinal glucose absorption and glucose-dependent incretin secretion. Diabetes 2012, 61, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruzdkov, A.A.; Gromova, L.V. Glucose absorption in the rat small intestine in vivo after various levels of local substrate load. Ross. Fiziol. Zhurnal Im. I. M. Sechenova 2013, 99, 630–641. (In Russian) [Google Scholar]
- Röder, P.V.; Geillinger, K.E.; Zietek, T.S.; Thorens, B.; Koepsell, H.; Daniel, H. The role of SGLT1 and GLUT2 in intestinal glucose transport and sensing. PLoS ONE 2014, 9, e89977. [Google Scholar] [CrossRef]
- Röder, P.V.; Wu, B.; Liu, Y.; Han, W. Pancreatic regulation of glucose homeostasis. Exp. Mol. Med. 2016, 48, e219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellett, G.L.; Brot-Laroche, E.; Mace, O.J.; Leturque, A. Sugar Absorption in the Intestine: The Role of GLUT2. Annu. Rev. Nutr. 2008, 28, 35–54. [Google Scholar] [CrossRef]
- Tobin, V.; Le Gall, M.; Fioramonti, X.; Stolarczyk, E.; Blazquez, A.G.; Klein, C.; Prigent, M.; Serradas, P.; Cuif, M.-H.; Magnan, C.; et al. Insulin internalizes GLUT2 in the enterocytes of healthy but not insulin-resistant mice. Diabetes 2008, 57, 555–562. [Google Scholar] [CrossRef]
- Ait-Omar, A.; Monteiro-Sepulveda, M.; Poitou, C.; Le Gall, M.; Cotillard, A.; Gilet, J.; Garbin, K.; Houllier, A.; Château, D.; Lacombe, A.; et al. GLUT2 accumulation in enterocyte apical and intracellular membranes: A study in morbidly obese human subjects and ob/ob and high fat–fed mice. Diabetes 2011, 60, 12598–12607. [Google Scholar] [CrossRef] [Green Version]
- Gromova, L.V.; Polozov, A.S.; Kornyushin, O.V.; Grefner, N.M.; Dmitrieva, Y.V.; Alekseeva, A.S.; Gruzdkov, A.A. Glucose Absorption in the Rat Small Intestine under Experimental Type 2 Diabetes Mellitus. J. Evol. Biochem. Physiol. 2019, 55, 155–157. [Google Scholar] [CrossRef]
- Cani, P.D.; Delzenne, N.M. The role of the gut microbiota in energy metabolism and metabolic disease. Curr. Pharm. Des. 2009, 15, 1546–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalles, J.P.; Orozco-Solis, R.; Bolanos-Jimenez, F. Perinatal Undernutrition Alters Intestinal Alkaline Phosphatase and Its Main Transcription Factors KLF4 and Cdx1 in Adult Offspring Fed a High-Fat Diet. J. Nutr. Biochem. 2012, 23, 1490–1497. [Google Scholar] [CrossRef]
- Lallès, J.P. Microbiota-host interplay at the gut epithelial level, health and nutrition. J. Anim. Sci. Biotechnol. 2016, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.B.; Carroll-Portillo, A.; Coffman, C.; Ritz, N.L.; Lin, H.C. Intestinal Alkaline Phosphatase Exerts Anti-Inflammatory Effects Against Lipopolysaccharide by Inducing Autophagy. Sci. Rep. 2020, 10, 3107. [Google Scholar] [CrossRef] [Green Version]
- Kaliannan, K.; Hamarneh, S.R.; Economopoulos, K.O.; Alam, S.N.; Moaven, O.; Patel, P.; Malo, N.S.; Ray, M.; Abtahi, S.M.; Muhammad, N.; et al. Intestinal alkaline phosphatase prevents metabolic syndrome in mice. Proc. Natl. Acad. Sci. USA 2013, 110, 7003–7008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malo, M.S. A high level of intestinal alkaline phosphatase is protective against type 2 diabetes mellitus irrespective of obesity. EBioMedicine 2015, 2, 2016–2023. [Google Scholar] [CrossRef] [Green Version]
- Lassenius, M.I.; Fogarty, C.L.; Blaut, M.; Haimila, K.; Riittinen, L.; Paju, A.; Kirveskari, J.; Järvelä, J.; Ahola, A.J.; Gordin, D.; et al. Intestinal alkaline phosphatase at the crossroad of intestinal health and disease—A putative role in type 1 diabetes. J. Intern. Med. 2017, 281, 586–600. [Google Scholar] [CrossRef]
- Kramer, W.; Girbig, F.; Corsiero, D.; Pfenninger, A.; Frick, W.; Jähne, G.; Rhein, M.; Wendler, W.; Lottspeich, F.; Hochleitner, E.O.; et al. Aminopeptidase N (CD13) is a molecular target of the cholesterol absorption inhibitor ezetimibe in the enterocyte brush border membrane. J. Biol. Chem. 2005, 280, 1306–1320. [Google Scholar] [CrossRef] [Green Version]
- Cong, Y.; Li, X.; Bai, Y.; Lv, X.; Herrler, G.; Enjuanesm, L.; Zhou, X.; Qu, B.; Meng, F.; Cong, C.; et al. Porcine aminopeptidase N mediated polarized infection by porcine epidemic diarrhea virus in target cells. Virology 2015, 478, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Santiago, C.; Mudgal, G.; Reguera, J.; Recacha, R.; Albrecht, S.; Enjuanes, L.; Casasnovas, J.M. Allosteric inhibition of aminopeptidase N functions related to tumor growth and virus infection. Sci. Rep. 2017, 7, 46045. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Amin, M.A.; Fox, D.A. CD13/Aminopeptidase N Is a Potential Therapeutic Target for Inflammatory Disorders. J. Immunol. 2020, 204, 3–11. [Google Scholar] [CrossRef]
- Gruzdkov, A.A.; Gromova, L.V.; Dmitrieva, Y.V.; Alekseeva, A.S. Free consumption of glucose solution by rats as a criterion for evaluation its absorption in the small intestine (Experimental study and mathematical modeling). Ross. Fiziol. Zhurnal Im. I. M. Sechenova 2015, 101, 708–720. (In Russian) [Google Scholar]
- Gruzdkov, A.A.; Dmitrieva, Y.V.; Alekseeva, A.S.; Polozov, A.S.; Gromova, L.V. Evaluation of Glucose Absorption Level in the Small Intestine of Different Rat Strains under Natural Conditions. J. Evol. Biochem. Physiol. 2018, 54, 308–315. [Google Scholar] [CrossRef]
- Maljaars, P.W.; Maljaars, P.H.P.; Mela, D.J.; Masclee, A.A. Ileal brake: A sensible food target for appetite control, a review. Physiol. Behav. 2008, 95, 271–281. [Google Scholar] [CrossRef]
- Grefner, N.M.; Gromova, L.V.; Gruzdkov, A.A.; Komissarchik, Y.Y. Comparative analysis of SGLT1 and GLUT2 transportovers distribution in rat small intestine enterocytes and Caco 2 cells during hexose absorption. Cell Tissue Biol. 2010, 4, 354–361. [Google Scholar] [CrossRef]
- Timofeeva, N.M.; Egorova, V.V.; Nikitina, A.A.; Dmitrieva, Y.V. Digestive enzyme activities in rats kept on standard or surplus breast-feeding and on low-protein diet directly after weaning. Zhurnal Evoliutsionnoi Biokhimii Fiziol. 2009, 45, 290–297. (In Russian) [Google Scholar]
- Dahlqvist, A. Method for assay of intestinal disaccharidases. Anal. Biochem. 1964, 7, 18–25. [Google Scholar] [CrossRef]
- Farr, W.; Rehfeld, N.; Reichelt, D.; Haschen, R.J. Comparative studies on the determination of amino acid aryl amidase in human serum. Z. Med. Labortechnik 1968, 9, 78–86. (In German) [Google Scholar]
- Ugolev, A.M.; Zaripov, B.Z.; Iezuitova, N.N.; Gruzdkov, A.A.; Rybin, I.S.; Voloshenovich, M.I.; Nikitina, A.A.; Punin, M.Y.; Tokgaevet, N.T. A revision of current data and views on membrane hydrolysis and transport in the mammalian small intestine based on a comparison of techniques of chronic and acute experiments: Experimental re-investigation and critical review. Comp. Biochem. Physiol. 1986, 85, 593–612. [Google Scholar] [CrossRef]
- Uhing, M.R.; Kimura, R.E. The effect of surgical bowel manipulation and anesthesia on intestinal glucose absorption in rats. J. Clin. Investig. 1995, 95, 2790–2798. [Google Scholar] [CrossRef] [PubMed]
- Gromova, L.V.; Gruzdkov, A.A. Hydrolysis-dependent absorption of disaccharides in the rat small intestine (chronic experiments and mathematical modeling). Gen. Physiol. Biophys. 1999, 18, 209–224. [Google Scholar]
- Mu, Q.; Kirby, J.; Reilly, C.M.; Luo, X.M. Leaky Gut As a Danger Signal for Autoimmune Diseases. Front Immunol. 2017, 8, 598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paray, B.A.; Albeshr, M.F.; Jan, A.T.; Rather, I.A. Leaky Gut and Autoimmunity: An Intricate Balance in Individuals Health and the Diseased State. Int. J. Mol. Sci. 2020, 21, 9770. [Google Scholar] [CrossRef] [PubMed]
- Hansen, G.H.; Niels-Christiansen, L.L.; Immerdal, L.; Nystrøm, B.T.; Danielsen, E.M. Intestinal alkaline phosphatase: Selective endocytosis from the enterocyte brush border during fat absorption. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G1325–G1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gromova, L.V.; Polozov, A.S.; Savochkina, E.V.; Alekseeva, A.S.; Dmitrieva, Y.V.; Kornyushin, O.V.; Gruzdkov, A.A. Effect of Type 2 Diabetes and Impaired Glucose Tolerance on Digestive Enzymes and Glucose Absorption in the Small Intestine of Young Rats. Nutrients 2022, 14, 385. https://doi.org/10.3390/nu14020385
Gromova LV, Polozov AS, Savochkina EV, Alekseeva AS, Dmitrieva YV, Kornyushin OV, Gruzdkov AA. Effect of Type 2 Diabetes and Impaired Glucose Tolerance on Digestive Enzymes and Glucose Absorption in the Small Intestine of Young Rats. Nutrients. 2022; 14(2):385. https://doi.org/10.3390/nu14020385
Chicago/Turabian StyleGromova, Lyudmila V., Alexandr S. Polozov, Elizaveta V. Savochkina, Anna S. Alekseeva, Yulia V. Dmitrieva, Oleg V. Kornyushin, and Andrey A. Gruzdkov. 2022. "Effect of Type 2 Diabetes and Impaired Glucose Tolerance on Digestive Enzymes and Glucose Absorption in the Small Intestine of Young Rats" Nutrients 14, no. 2: 385. https://doi.org/10.3390/nu14020385
APA StyleGromova, L. V., Polozov, A. S., Savochkina, E. V., Alekseeva, A. S., Dmitrieva, Y. V., Kornyushin, O. V., & Gruzdkov, A. A. (2022). Effect of Type 2 Diabetes and Impaired Glucose Tolerance on Digestive Enzymes and Glucose Absorption in the Small Intestine of Young Rats. Nutrients, 14(2), 385. https://doi.org/10.3390/nu14020385