
Antibacterial Activity and Epigenetic Remodeling of Essential Oils from Calabrian Aromatic Plants

 , ,
, ,  and
and
Abstract
:
1. Introduction
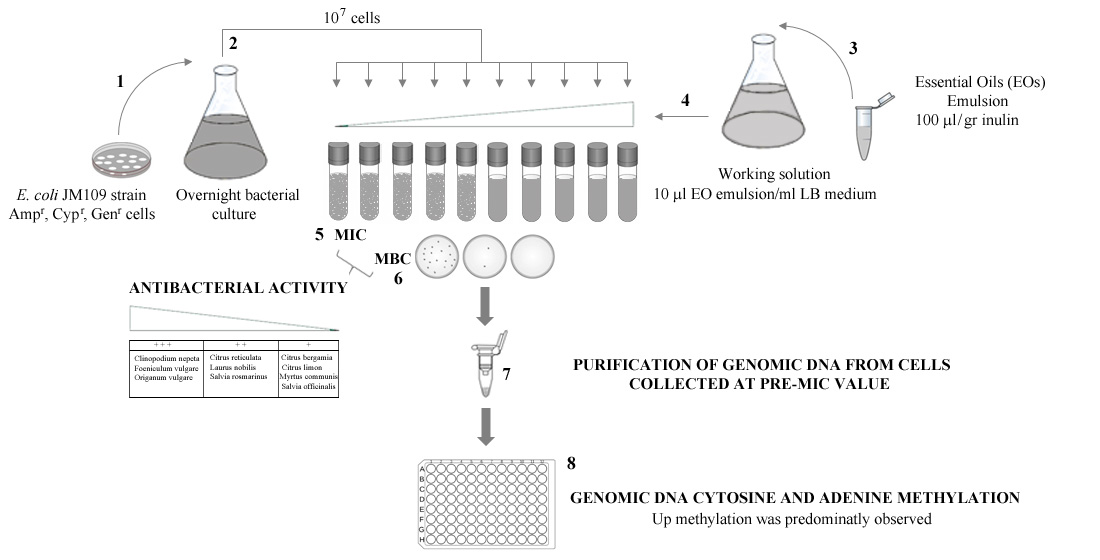
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Essential Oils (EOs) Extraction
2.3. Analysis of Chemical Composition of Essential Oils
2.4. Determination of Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
2.5. DNA Extraction
2.6. Quantification of Global 5-Methylcytosine and N6-Methyladenosine Levels
2.7. Statistical Analysis
3. Results
3.1. Antibacterial Activity of Essential Oils
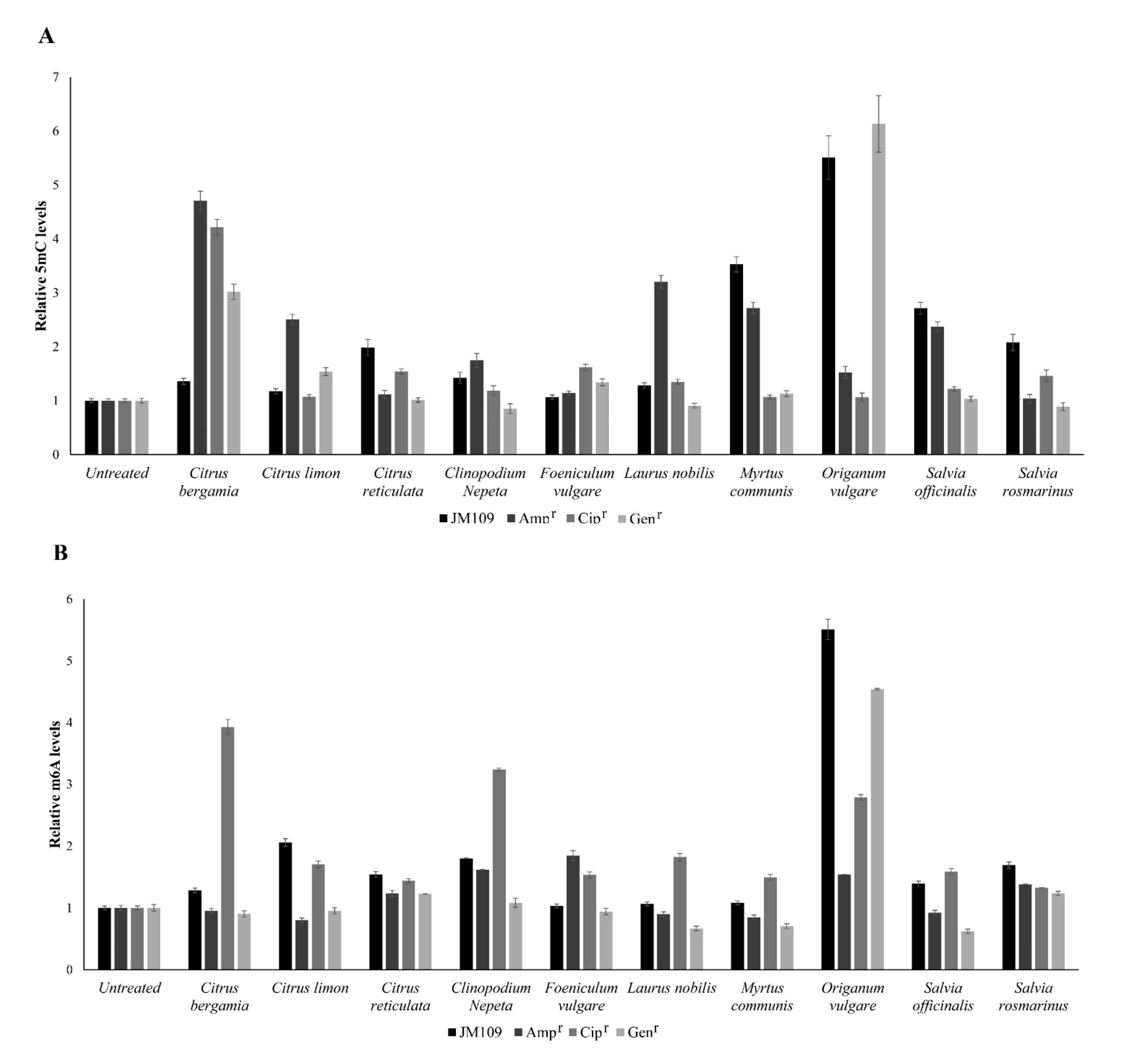
3.2. Effects of EOs on the Methylation Profiles of Citosine and Adenine
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cragg, G.M.; Newman, D.J. Natural Products: A Continuing Source of Novel Drug Leads. Biochim. Biophys. Acta 2013, 1830, 3670–3695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharifi-Rad, J.; Sureda, A.; Tenore, G.C.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.R.; Ademiluyi, A.O.; et al. Biological Activities of Essential Oils: From Plant Chemoecology to Traditional Healing Systems. Molecules 2017, 22, 70. [Google Scholar] [CrossRef] [PubMed]
- Lautié, E.; Russo, O.; Ducrot, P.; Boutin, J.A. Unraveling Plant Natural Chemical Diversity for Drug Discovery Purposes. Front. Pharmacol. 2020, 11, 397. [Google Scholar] [CrossRef] [PubMed]
- Dini, I.; Laneri, S. Spices, Condiments, Extra Virgin Olive Oil and Aromas as not only Flavorings, but Precious Allies for our Wellbeing. Antioxidants 2021, 10, 868. [Google Scholar] [CrossRef]
- Gavahian, M.; Chu, Y.H.; Lorenzo, J.M.; Mousavi Khaneghah, A.; Barba, F.J. Essential Oils as Natural Preservatives for Bakery Products: Understanding the Mechanisms of Action, Recent Findings, and Applications. Crit. Rev. Food Sci. Nutr. 2020, 60, 310–321. [Google Scholar] [CrossRef]
- Falleh, H.; Ben Jemaa, M.; Saada, M.; Ksouri, R. Essential Oils: A Promising Eco-Friendly Food Preservative. Food Chem. 2020, 330, 127268. [Google Scholar] [CrossRef]
- Valdivieso-Ugarte, M.; Gomez-Llorente, C.; Plaza-Díaz, J.; Gil, Á. Antimicrobial, Antioxidant, and Immunomodulatory Properties of Essential Oils: A Systematic Review. Nutrients 2019, 11, 2786. [Google Scholar] [CrossRef] [Green Version]
- Heghes, S.C.; Vostinaru, O.; Rus, L.M.; Mogosan, C.; Iuga, C.A.; Filip, L. Antispasmodic Effect of Essential Oils and their Constituents: A Review. Molecules 2019, 24, 1675. [Google Scholar] [CrossRef] [Green Version]
- Sandner, G.; Heckmann, M.; Weghuber, J. Immunomodulatory Activities of Selected Essential Oils. Biomolecules 2020, 10, 1139. [Google Scholar] [CrossRef]
- Barra, A. Factors Affecting Chemical Variability of Essential Oils: A Review of Recent Developments. Nat. Prod. Commun. 2009, 4, 1147–1154. [Google Scholar] [CrossRef] [Green Version]
- Wojtunik-Kulesza, K.A.; Kasprzak, K.; Oniszczuk, T.; Oniszczuk, A. Natural Monoterpenes: Much More than Only a Scent. Chem. Biodivers. 2019, 16, e1900434. [Google Scholar] [CrossRef]
- Abers, M.; Schroeder, S.; Goelz, L.; Sulser, A.; St Rose, T.; Puchalski, K.; Langland, J. Antimicrobial Activity of the Volatile Substances from Essential Oils. BMC Complement Med. Ther. 2021, 21, 124. [Google Scholar] [CrossRef]
- Bora, H.; Kamle, M.; Mahato, D.K.; Tiwari, P.; Kumar, P. Citrus Essential Oils (CEOS) and their Applications in Food: An Overview. Plants 2020, 9, 357. [Google Scholar] [CrossRef] [Green Version]
- Kaya, D.A.; Ghica, M.V.; Dănilă, E.; Öztürk, Ş.; Türkmen, M.; Albu Kaya, M.G.; Dinu-Pîrvu, C.E. Selection of Optimal Operating Conditions for Extraction of Myrtus Communis L. Essential Oil by the Steam Distillation Method. Molecules 2020, 25, 2399. [Google Scholar] [CrossRef]
- Ni, Z.-J.; Wang, X.; Shen, Y.; Thakur, K.; Han, J.; Zhang, J.-G.; Hu, F.; Wei, Z.-J. Recent Updates on the Chemistry, Bioactivities, Mode of Action, and Industrial Applications of Plant Essential Oils. Trends Food Sci. Technol. 2021, 110, 78–89. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological Effects of Essential Oils—A Review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential Oils’ Chemical Characterization and Investigation of Some Biological Activities: A Critical Review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Fung, F.; Wang, H.S.; Menon, S. Food Safety in the 21st Century. Biomed. J. 2018, 41, 88–95. [Google Scholar] [CrossRef]
- Mith, H.; Duré, R.; Delcenserie, V.; Zhiri, A.; Daube, G.; Clinquart, A. Antimicrobial Activities of Commercial Essential Oils and their Components against Food-Borne Pathogens and Food Spoilage Bacteria. Food Sci. Nutr. 2014, 2, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Bouarab Chibane, L.; Degraeve, P.; Ferhout, H.; Bouajila, J.; Oulahal, N. Plant Antimicrobial Polyphenols as Potential Natural Food Preservatives. J. Sci. Food Agric. 2019, 99, 1457–1474. [Google Scholar] [CrossRef] [Green Version]
- Larsson, D.G.J.; Flach, C.F. Antibiotic Resistance in the Environment. Nat. Rev. Microbiol. 2021, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Del-Río, I.; López-Ibáñez, S.; Magadán-Corpas, P.; Fernández-Calleja, L.; Pérez-Valero, Á.; Tuñón-Granda, M.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Terpenoids and Polyphenols as Natural Antioxidant Agents in Food Preservation. Antioxidants 2021, 10, 1264. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Martínez, F.J.; Barrajón-Catalán, E.; Encinar, J.A.; Rodríguez-Díaz, J.C.; Micol, V. Antimicrobial Capacity of Plant Polyphenols against Gram-positive Bacteria: A Comprehensive Review. Curr. Med. Chem. 2020, 27, 2576–2606. [Google Scholar] [CrossRef]
- Álvarez-Martínez, F.J.; Barrajón-Catalán, E.; Herranz-López, M.; Micol, V. Antibacterial Plant Compounds, Extracts and Essential Oils: An Updated Review on their Effects and Putative Mechanisms of Action. Phytomedicine 2021, 90, 153626. [Google Scholar] [CrossRef] [PubMed]
- Pattnaik, S.; Subramanyam, V.R.; Bapaji, M.; Kole, C.R. Antibacterial and Antifungal Activity of Aromatic Constituents of Essential Oils. Microbios 1997, 89, 39–46. [Google Scholar] [PubMed]
- Kalemba, D.; Kunicka, A. Antibacterial and Antifungal Properties of Essential Oils. Curr. Med. Chem. 2003, 10, 813–829. [Google Scholar] [CrossRef]
- Fournomiti, M.; Kimbaris, A.; Mantzourani, I.; Plessas, S.; Theodoridou, I.; Papaemmanouil, V.; Kapsiotis, I.; Panopoulou, M.; Stavropoulou, E.; Bezirtzoglou, E.E.; et al. Antimicrobial Activity of Essential Oils of Cultivated Oregano (Origanum vulgare), Sage (Salvia officinalis), and Thyme (Thymus vulgaris) against Clinical Isolates of Escherichia Coli, Klebsiella Oxytoca, and Klebsiella Pneumoniae. Microb. Ecol. Health Dis. 2015, 26, 23289–23295. [Google Scholar] [CrossRef]
- Tariq, S.; Wani, S.; Rasool, W.; Shafi, K.; Bhat, M.A.; Prabhakar, A.; Shalla, A.H.; Rather, M.A. A Comprehensive Review of the Antibacterial, Antifungal and Antiviral Potential of Essential Oils and their Chemical Constituents against Drug-Resistant Microbial Pathogens. Microb. Pathog. 2019, 134, 103580. [Google Scholar] [CrossRef]
- Lambert, R.J.; Skandamis, P.N.; Coote, P.J.; Nychas, G.J. A Study of the Minimum Inhibitory Concentration and Mode of Action of Oregano Essential Oil, Thymol and Carvacrol. J. Appl. Microbiol. 2001, 91, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Chandra, H.; Bishnoi, P.; Yadav, A.; Patni, B.; Mishra, A.P.; Nautiyal, A.R. Antimicrobial Resistance and the Alternative Resources with Special Emphasis on Plant-Based Antimicrobials—A Review. Plants 2017, 6, 16. [Google Scholar] [CrossRef]
- Khameneh, B.; Iranshahy, M.; Soheili, V.; Fazly Bazzaz, B.S. Review on Plant Antimicrobials: A Mechanistic Viewpoint. Antimicrob. Resist. Infect. Control. 2019, 8, 118. [Google Scholar] [CrossRef] [Green Version]
- Alibi, S.; Ben Selma, W.; Ramos-Vivas, J.; Smach, M.A.; Touati, R.; Boukadida, J.; Navas, J.; Ben Mansour, H. Anti-Oxidant, Antibacterial, Anti-Biofilm, and Anti-Quorum Sensing Activities of Four Essential Oils against Multidrug-Resistant Bacterial Clinical Isolates. Curr. Res. Transl. Med. 2020, 68, 59–66. [Google Scholar] [CrossRef]
- Sharifi, A.; Mohammadzadeh, A.; Zahraei Salehi, T.; Mahmoodi, P. Antibacterial, Antibiofilm and Antiquorum Sensing Effects of Thymus Daenensis and Satureja Hortensis Essential Oils against Staphylococcus Aureus Isolates. J. Appl. Microbiol. 2018, 124, 379–388. [Google Scholar] [CrossRef]
- Zhang, D.; Gan, R.Y.; Zhang, J.R.; Farha, A.K.; Li, H.B.; Zhu, F.; Wang, X.H.; Corke, H. Antivirulence Properties and Related Mechanisms of Spice Essential Oils: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1018–1055. [Google Scholar] [CrossRef] [Green Version]
- Reichling, J. Anti-Biofilm and Virulence Factor-Reducing Activities of Essential Oils and Oil Components as a Possible Option for Bacterial Infection Control. Planta Med. 2020, 86, 520–537. [Google Scholar] [CrossRef]
- Sandoval-Motta, S.; Aldana, M. Adaptive Resistance to Antibiotics in Bacteria: A Systems Biology Perspective. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 253–267. [Google Scholar] [CrossRef]
- Hao, Y.; Kang, J.; Yang, R.; Li, H.; Cui, H.; Bai, H.; Tsitsilin, A.; Li, J.; Shi, L. Multidimensional Exploration of Essential Oils Generated Via Eight Oregano Cultivars: Compositions, Chemodiversities, and Antibacterial Capacities. Food Chem. 2021, 374, 131629. [Google Scholar] [CrossRef]
- Pereira, C.; Barros, L.; Ferreira, I.C. A Comparison of the Nutritional Contribution of Thirty-Nine Aromatic Plants Used as Condiments and/or Herbal Infusions. Plant Foods Hum. Nutr. 2015, 70, 176–183. [Google Scholar] [CrossRef]
- Carlson, J.L.; Erickson, J.M.; Lloyd, B.B.; Slavin, J.L. Health Effects and Sources of Prebiotic Dietary Fiber. Curr. Dev. Nutr. 2018, 2, nzy005. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, W.; Rashid, S. Functional and Therapeutic Potential of Inulin: A Comprehensive Review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1–13. [Google Scholar] [CrossRef]
- Thielmann, J.; Muranyi, P.; Kazman, P. Screening Essential Oils for their Antimicrobial Activities against the Foodborne Pathogenic Bacteria Escherichia Coli and Staphylococcus Aureus. Heliyon 2019, 5, e01860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puškárová, A.; Bučková, M.; Kraková, L.; Pangallo, D.; Kozics, K. The Antibacterial and Antifungal Activity of Six Essential Oils and their Cyto/Genotoxicity to Human HEL 12469 Cells. Sci. Rep. 2017, 7, 8211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arantes, S.M.; Piçarra, A.; Guerreiro, M.; Salvador, C.; Candeias, F.; Caldeira, A.T.; Martins, M.R. Toxicological and Pharmacological Properties of Essential Oils of Calamintha Nepeta, Origanum Virens and Thymus Mastichina of Alentejo (Portugal). Food Chem. Toxicol. 2019, 133, 110747. [Google Scholar] [CrossRef] [PubMed]
- Gormez, A.; Bozari, S.; Yanmis, D.; Gulluce, M.; Sahin, F.; Agar, G. Chemical Composition and Antibacterial Activity of Essential Oils of Two Species of Lamiaceae against Phytopathogenic Bacteria. Pol. J. Microbiol. 2015, 64, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Dadalioglu, I.; Evrendilek, G.A. Chemical Compositions and Antibacterial Effects of Essential Oils of Turkish Oregano (Origanum minutiflorum), Bay Laurel (Laurus nobilis), Spanish Lavender (Lavandula Stoechas L.), and Fennel (Foeniculum vulgare) on Common Foodborne Pathogens. J. Agric. Food Chem. 2004, 52, 8255–8260. [Google Scholar] [CrossRef]
- Ruberto, G.; Baratta, M.T.; Deans, S.G.; Dorman, H.J. Antioxidant and Antimicrobial Activity of Foeniculum vulgare and Crithmum Maritimum Essential Oils. Planta Med. 2000, 66, 687–693. [Google Scholar] [CrossRef]
- Pellegrini, M.; Ricci, A.; Serio, A.; Chaves-López, C.; Mazzarrino, G.; D’Amato, S.; Lo Sterzo, C.; Paparella, A. Characterization of Essential Oils Obtained from Abruzzo Autochthonous Plants: Antioxidant and Antimicrobial Activities Assessment for Food Application. Foods 2018, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Cetin, B.; Ozer, H.; Cakir, A.; Polat, T.; Dursun, A.; Mete, E.; Oztürk, E.; Ekinci, M. Antimicrobial Activities of Essential Oil and Hexane Extract of Florence Fennel [Foeniculum vulgare Var. Azoricum (Mill.) Thell.] against Foodborne Microorganisms. J. Med. Food 2010, 13, 196–204. [Google Scholar] [CrossRef]
- Napoli, E.; Giovino, A.; Carrubba, A.; How Yuen Siong, V.; Rinoldo, C.; Nina, O.; Ruberto, G. Variations of Essential Oil Constituents in Oregano (Origanum vulgare subsp. viridulum (= O. heracleoticum) over Cultivation Cycles. Plants 2020, 9, 1174. [Google Scholar] [CrossRef]
- Novak, J.; Lukas, B.; Franz, C. Temperature Influences Thymol and Carvacrol Differentially in Origanum spp.(Lamiaceae). J. Essent. Oil Res. 2010, 22, 412–415. [Google Scholar] [CrossRef]
- Giuliani, C.; Maggi, F.; Papa, F.; Maleci Bini, L. Congruence of Phytochemical and Morphological Profiles along an Altitudinal Gradient in Origanum vulgare ssp. vulgare from Venetian Region (NE Italy). Chem. Biodivers. 2013, 10, 569–583. [Google Scholar] [CrossRef]
- Kokkini, S.; Karousou, R.; Dardioti, A.; Krigas, N.; Lanaras, T. Autumn essential oils of Greek Oregano. Phytochemistry 1997, 44, 883–886. [Google Scholar] [CrossRef]
- Carrubba, A.; Catalano, C. Essential Oil Crops for Sustainable Agriculture—A review. In Climate Change, Intercropping, Pest Control and Beneficial Microorganisms; Lichtfouse, E., Ed.; Springer: Dijon, France, 2009; Volume 55, pp. 137–188. [Google Scholar]
- Gullberg, E.; Cao, S.; Berg, O.G.; Ilbäck, C.; Sandegren, L.; Hughes, D.; Andersson, D.I. Selection of Resistant Bacteria at Very Low Antibiotic Concentrations. PLoS Pathog. 2011, 7, e1002158. [Google Scholar] [CrossRef] [Green Version]
- Unusan, N. Essential Oils and Microbiota: Implications for Diet and Weight Control. Trend Food Sci. Technol. 2020, 104, 60–71. [Google Scholar] [CrossRef]
- Sánchez-Romero, M.A.; Casadesús, J. The Bacterial Epigenome. Nat. Rev. Microbiol. 2020, 18, 7–20. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Essential Oils | JM109 | Ampr | Cipr | Genr | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | |||||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Clinopodium nepeta | 0.966 | 0.057 | 1.000 | 0.000 | 0.633 | 0.058 | 0.800 | 0.000 | 0.867 | 0.115 | 1.000 | 0.058 | 0.900 | 0.000 | 0.900 | 0.058 |
| Citrus bergamia | 6.333 | 0.577 | 5.667 | 0.577 | 4.667 | 0.577 | 4.667 | 0.577 | 10.000 | 0.000 | 10.000 | 0.000 | 10.000 | 0.000 | 10.000 | 0.000 |
| Citrus limon | 5.000 | 0.000 | 5.000 | 0.000 | 2.000 | 0.000 | 2.000 | 0.000 | 8.333 | 1.528 | 10.000 | 0.000 | 3.000 | 0.000 | 3.000 | 0.000 |
| Citrus reticulata | 2.333 | 0.577 | 2.000 | 0.000 | 2.000 | 0.000 | 2.000 | 0.000 | 2.667 | 1.155 | 2.333 | 0.577 | 3.000 | 1.000 | 4.667 | 0.577 |
| Foeniculum vulgare | 0.400 | 0.000 | 0.400 | 0.000 | 0.367 | 0.058 | 0.400 | 0.000 | 0.567 | 0.058 | 0.600 | 0.000 | 0.333 | 0.058 | 0.400 | 0.000 |
| Laurus nobilis | 2.333 | 0.577 | 3.000 | 0.000 | 2.000 | 0.000 | 2.000 | 0.000 | 2.000 | 0.000 | 2.000 | 0.000 | 2.000 | 0.000 | 2.000 | 0.000 |
| Myrtus communis | 4.333 | 0.577 | 5.000 | 0.000 | 3.667 | 0.577 | 5.000 | 0.000 | 10.000 | 0.000 | 10.000 | 0.000 | 5.667 | 0.577 | 6.333 | 0.577 |
| Origanum vulgare | 0.300 | 0.000 | 0.300 | 0.000 | 0.300 | 0.000 | 0.300 | 0.000 | 0.300 | 0.000 | 0.300 | 0.000 | 0.300 | 0.000 | 0.300 | 0.000 |
| Salvia officinalis | 10.000 | 0.000 | 10.000 | 0.000 | 2.000 | 0.000 | 2.000 | 0.000 | 4.333 | 0.577 | 6.333 | 1.155 | 4.000 | 0.000 | 4.000 | 0.000 |
| Salvia rosmarinus | 2.000 | 0.000 | 2.333 | 0.577 | 2.000 | 0.000 | 2.000 | 0.000 | 6.333 | 0.577 | 7.000 | 0.000 | 2.333 | 0.577 | 3.000 | 0.000 |
| Essential Oil | Component Name | % |
|---|---|---|
| Clinopodium nepeta (L.) Kuntze | piperitone oxide | 34.28 |
| piperitenone oxide | 18.23 | |
| (+)-limonene | 15.80 | |
| (+)-pulegone | 13.75 | |
| menthone | 8.32 | |
| isolegylacetate | 3.64 | |
| 1-terpine-4-ol | 1.40 | |
| (+)-neomenthol | 1.37 | |
| β-pinene | 1.22 | |
| Citrus bergamia, (Risso & Poit.) | (+)-limonene | 38.88 |
| lynalyl acetate | 34.28 | |
| (+)-linalool | 11.54 | |
| α-terpinene | 6.79 | |
| β-pinene | 5.49 | |
| α-pinene | 1.22 | |
| Citrus limon (L.) Osbeck | (+)-limonene | 74.41 |
| α-terpinene | 11.91 | |
| β-pinene | 4.34 | |
| α-terpineol | 3.01 | |
| α-terpinolene | 1.67 | |
| 1-terpine-4-ol | 1.26 | |
| Citrus reticulata (Blanco) | (+)-sabinene | 50.91 |
| (+)-linalool | 18.27 | |
| α-phellandrene | 6.54 | |
| β-cis-ocimene | 6.45 | |
| (+)-limonene | 5.04 | |
| β-myrcene | 2.37 | |
| β-pinene | 2.35 | |
| α-pinene | 1.93 | |
| β-citronella | 1.44 | |
| α-terpinolene | 1.37 | |
| α-terpinene | 1.14 | |
| Foeniculum vulgare subsp. piperitum (Ucria) Bég. | estragole | 45.33 |
| α-pinene | 14.71 | |
| anethal | 14.54 | |
| fenchone | 11.24 | |
| α-limonene | 8.49 | |
| α-phellandrene | 2.51 | |
| β-pinene | 1.65 | |
| β-myrcene | 1.05 | |
| Laurus nobilis L. | eucalyptol | 56.61 |
| (+)-sabinene | 15.74 | |
| (+)-linalool | 7.38 | |
| terpinyl acetate | 6.48 | |
| α-pinene | 5.65 | |
| methyleugenol | 1.51 | |
| 1-terpine-4-ol | 1.29 | |
| Myrtus communis L. | eucalyptol | 33.04 |
| (−)-myrtenylacetate | 17.04 | |
| α-pinene | 12.33 | |
| (+)-limonene | 10.81 | |
| (+)-linalool | 10.43 | |
| lynalyl acetate | 3.88 | |
| geraniol acetate | 1.88 | |
| α-terpineol | 2.10 | |
| β-ocimene | 1.58 | |
| α-phellandrene | 1.41 | |
| o-cymene | 1.41 | |
| terpinolene | 1.10 | |
| terpinene | 1.09 | |
| Origanum vulgare L. subsp. viridulum (Martrin-Donos) Nyman | p-thymol | 47.31 |
| terpinene | 18.52 | |
| p-cymene | 11.78 | |
| β-caryophyllene | 4.88 | |
| β-myrcene | 3.76 | |
| carvacrol | 3.52 | |
| terpinolene | 3.18 | |
| α-thujene (origanene) | 2.73 | |
| α-pinene | 1.23 | |
| Salvia officinalis L. | eucalyptol | 23.70 |
| (−)-α-thujone | 24.14 | |
| β-pinene | 15.10 | |
| (−)-camphor | 9.59 | |
| α-humulene | 5.54 | |
| (−)-β-thujone | 4.35 | |
| α-pinene | 3.99 | |
| (−)-β-caryophyllene | 2.85 | |
| β-myrcene | 2.26 | |
| camphene | 1.88 | |
| (+)-sabinene | 1.13 | |
| Salvia rosmarinus Spenn | eucalyptol | 49.29 |
| α-pinene | 22.84 | |
| β-pinene | 9.26 | |
| camphene | 6.70 | |
| (−)-camphor | 3.66 | |
| isoborneol | 2.28 | |
| β-myrcene | 1.79 | |
| (−)-β-caryophyllene | 1.17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Aquila, P.; Paparazzo, E.; Crudo, M.; Bonacci, S.; Procopio, A.; Passarino, G.; Bellizzi, D. Antibacterial Activity and Epigenetic Remodeling of Essential Oils from Calabrian Aromatic Plants. Nutrients 2022, 14, 391. https://doi.org/10.3390/nu14020391
D’Aquila P, Paparazzo E, Crudo M, Bonacci S, Procopio A, Passarino G, Bellizzi D. Antibacterial Activity and Epigenetic Remodeling of Essential Oils from Calabrian Aromatic Plants. Nutrients. 2022; 14(2):391. https://doi.org/10.3390/nu14020391
Chicago/Turabian StyleD’Aquila, Patrizia, Ersilia Paparazzo, Michele Crudo, Sonia Bonacci, Antonio Procopio, Giuseppe Passarino, and Dina Bellizzi. 2022. "Antibacterial Activity and Epigenetic Remodeling of Essential Oils from Calabrian Aromatic Plants" Nutrients 14, no. 2: 391. https://doi.org/10.3390/nu14020391
APA StyleD’Aquila, P., Paparazzo, E., Crudo, M., Bonacci, S., Procopio, A., Passarino, G., & Bellizzi, D. (2022). Antibacterial Activity and Epigenetic Remodeling of Essential Oils from Calabrian Aromatic Plants. Nutrients, 14(2), 391. https://doi.org/10.3390/nu14020391