
Astragalosides Supplementation Enhances Intrinsic Muscle Repair Capacity Following Eccentric Exercise-Induced Injury



Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Muscle Damage Protocol
2.3. Convalescence: Astragalosides and Placebo Supplement
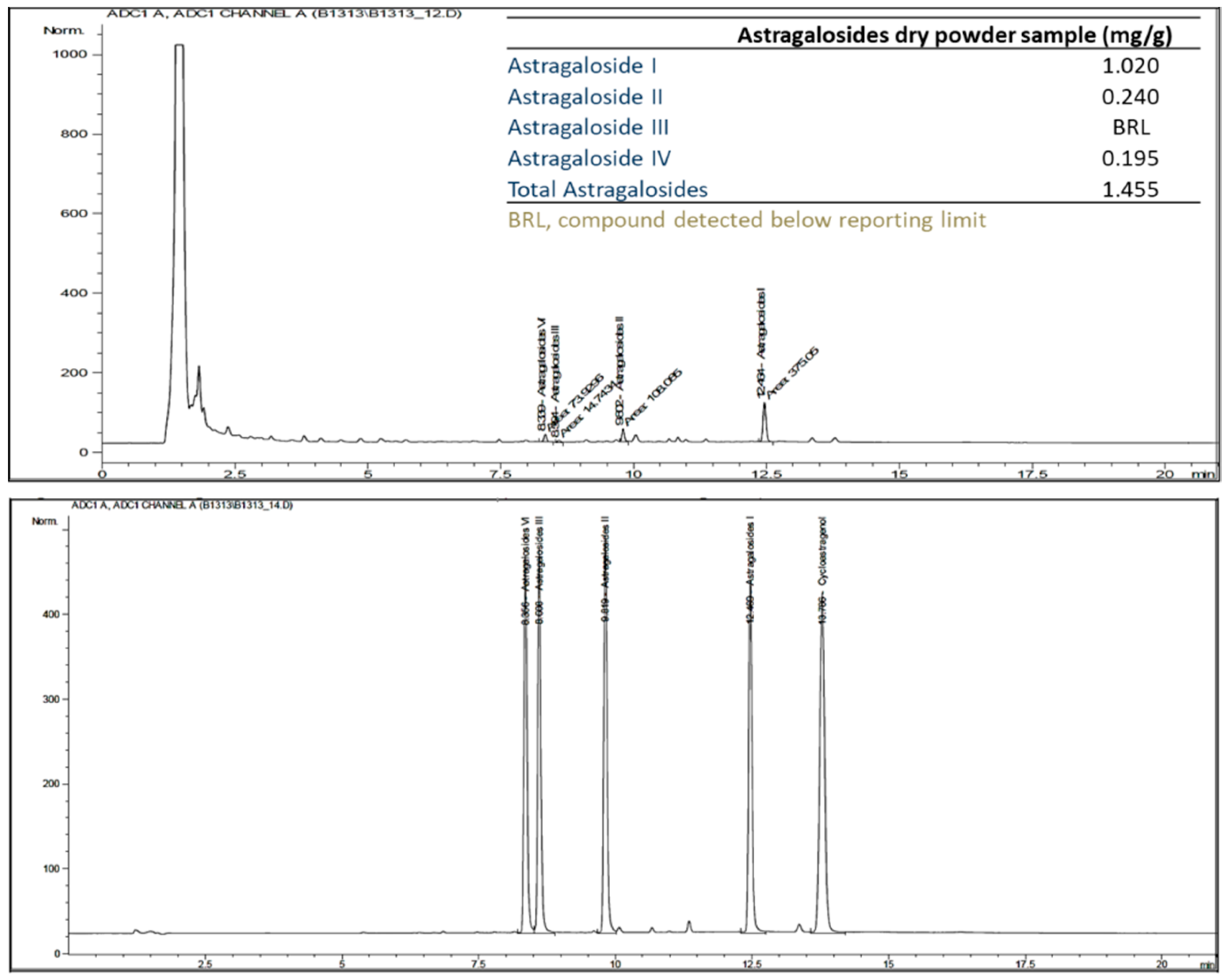
2.4. Quantitative Analysis of Astragalosides Dry Powder in Capsules
2.5. Assessment of Muscle Pain
2.6. Biochemical Analysis
2.7. Assessment of Muscular Strength
2.8. Muscle Oxygenation Monitoring
2.9. Statistical Analyses
3. Results
3.1. Subjects’ Characteristics
3.2. Astragalosides Quantification
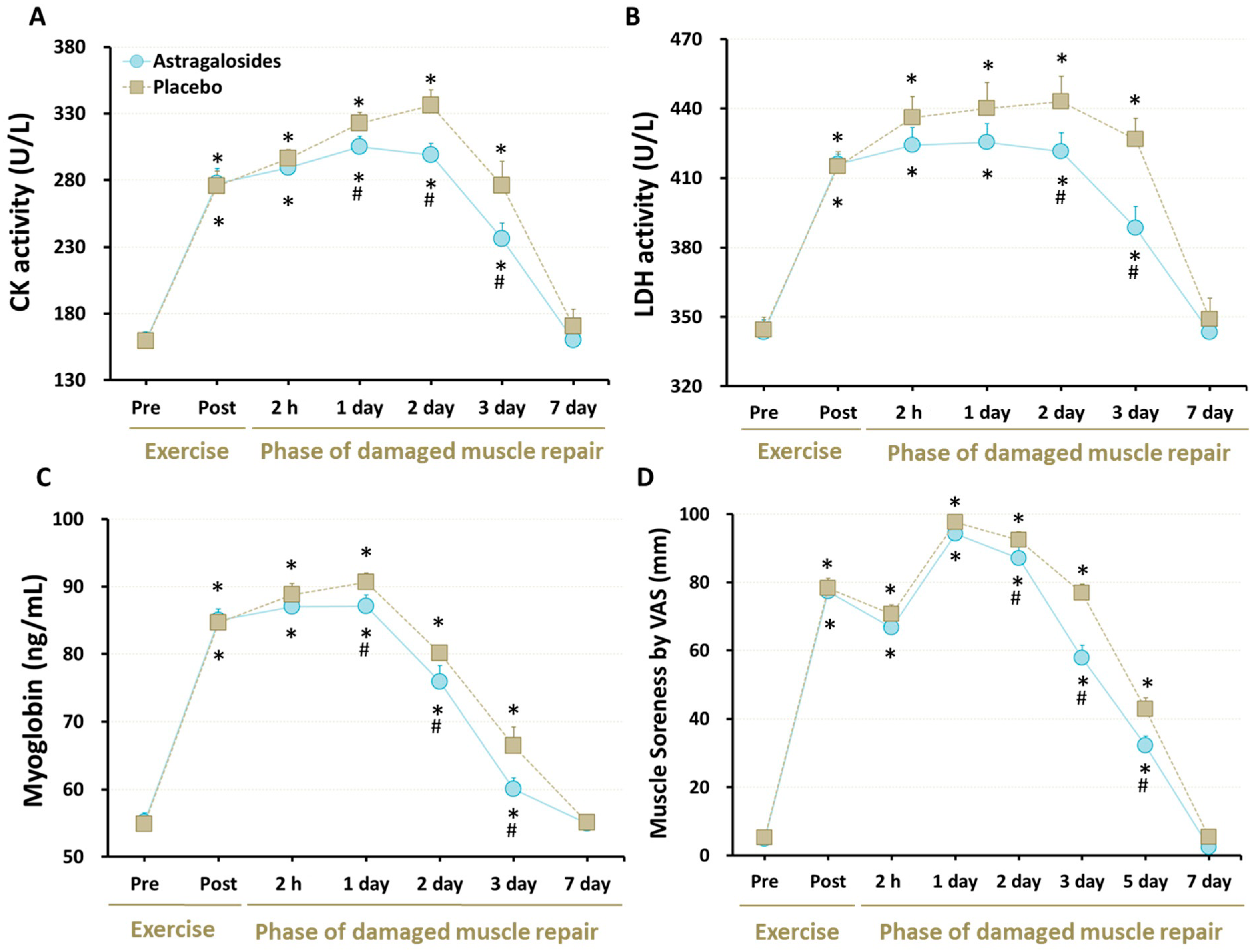
3.3. Cellular Markers of Muscle Damage
3.4. Muscle Soreness
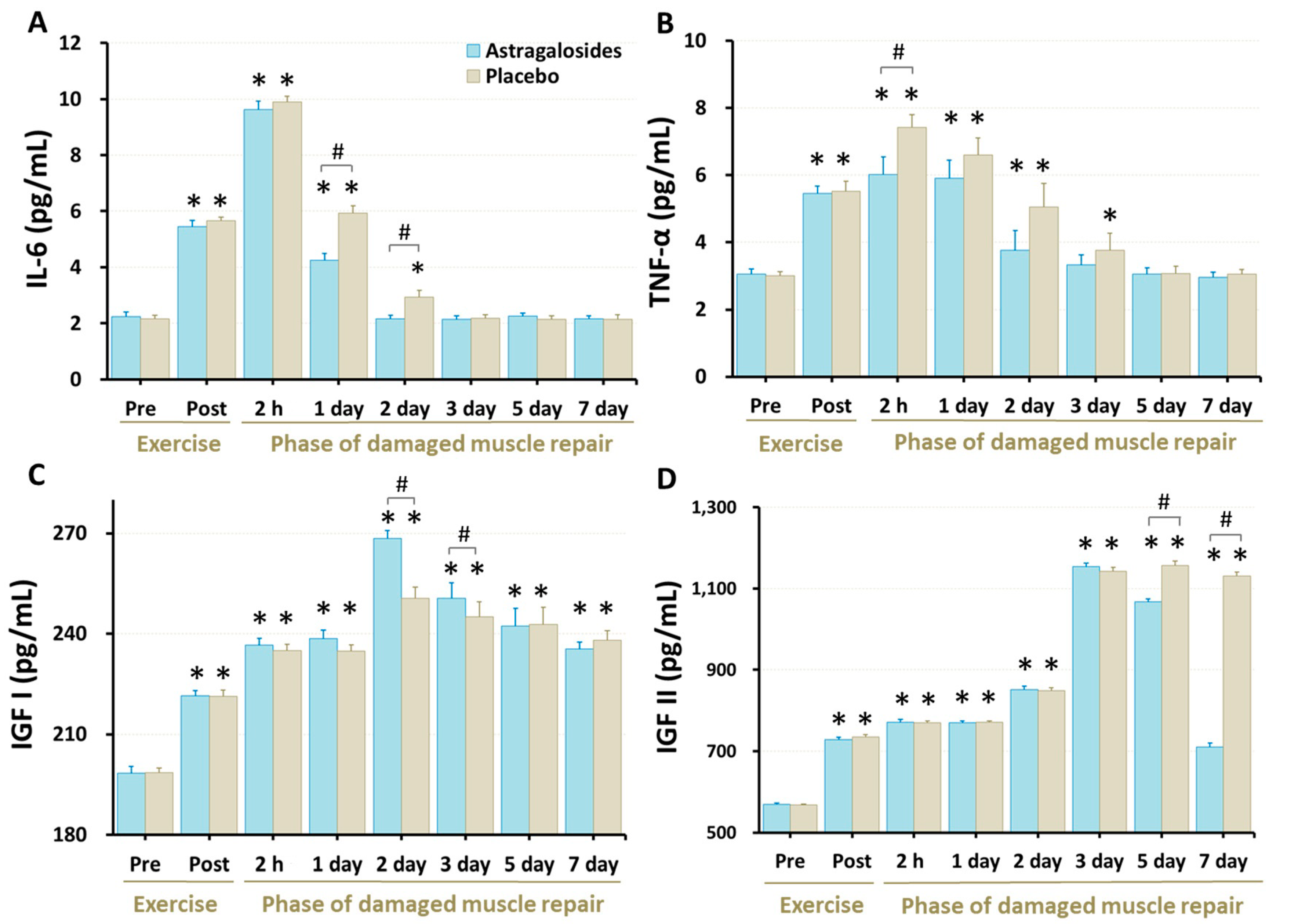
3.5. Inflammation Responses and Regeneration Regulatory Factors
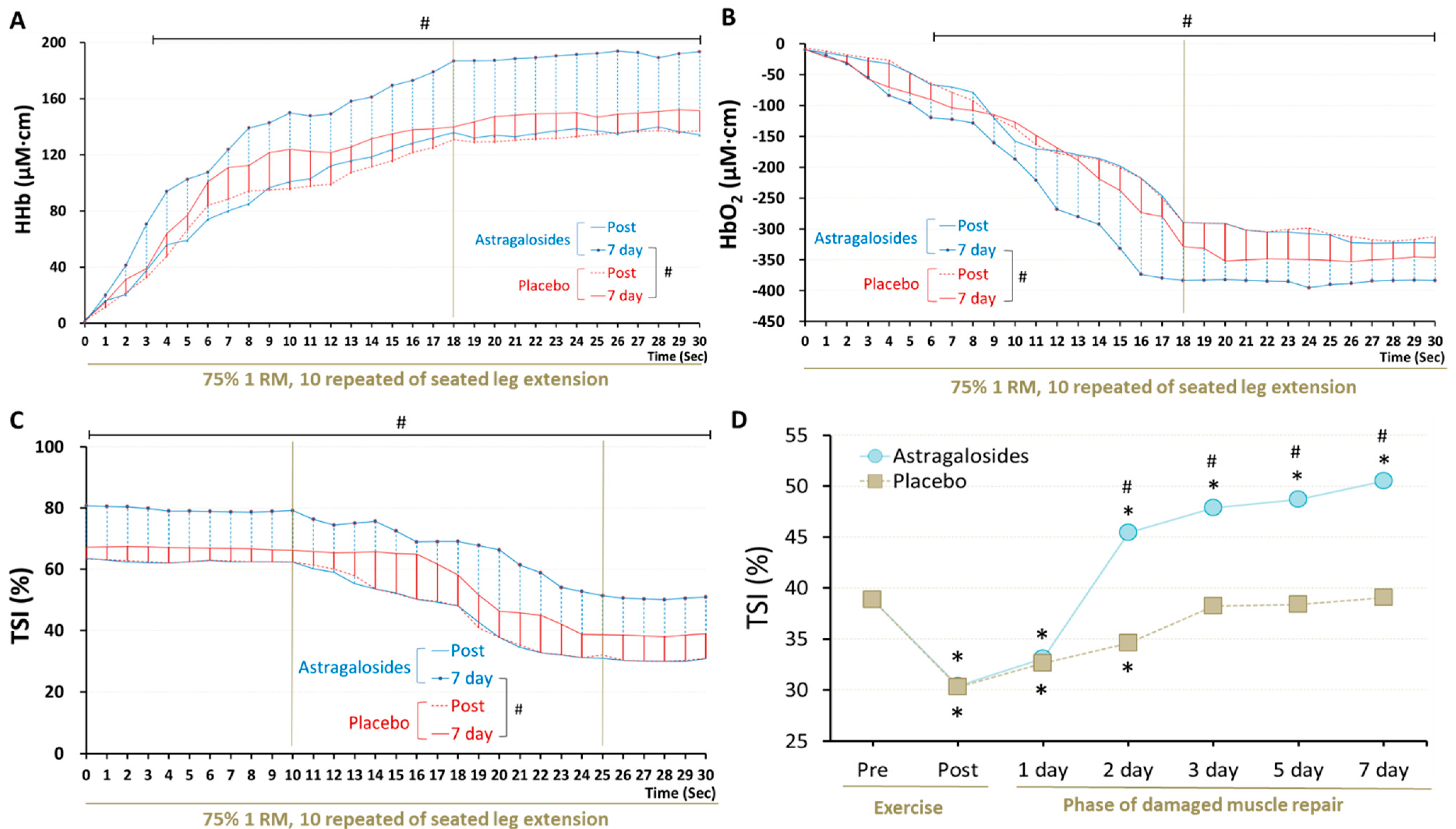
3.6. Recovery of Muscle Respiratory Capacity
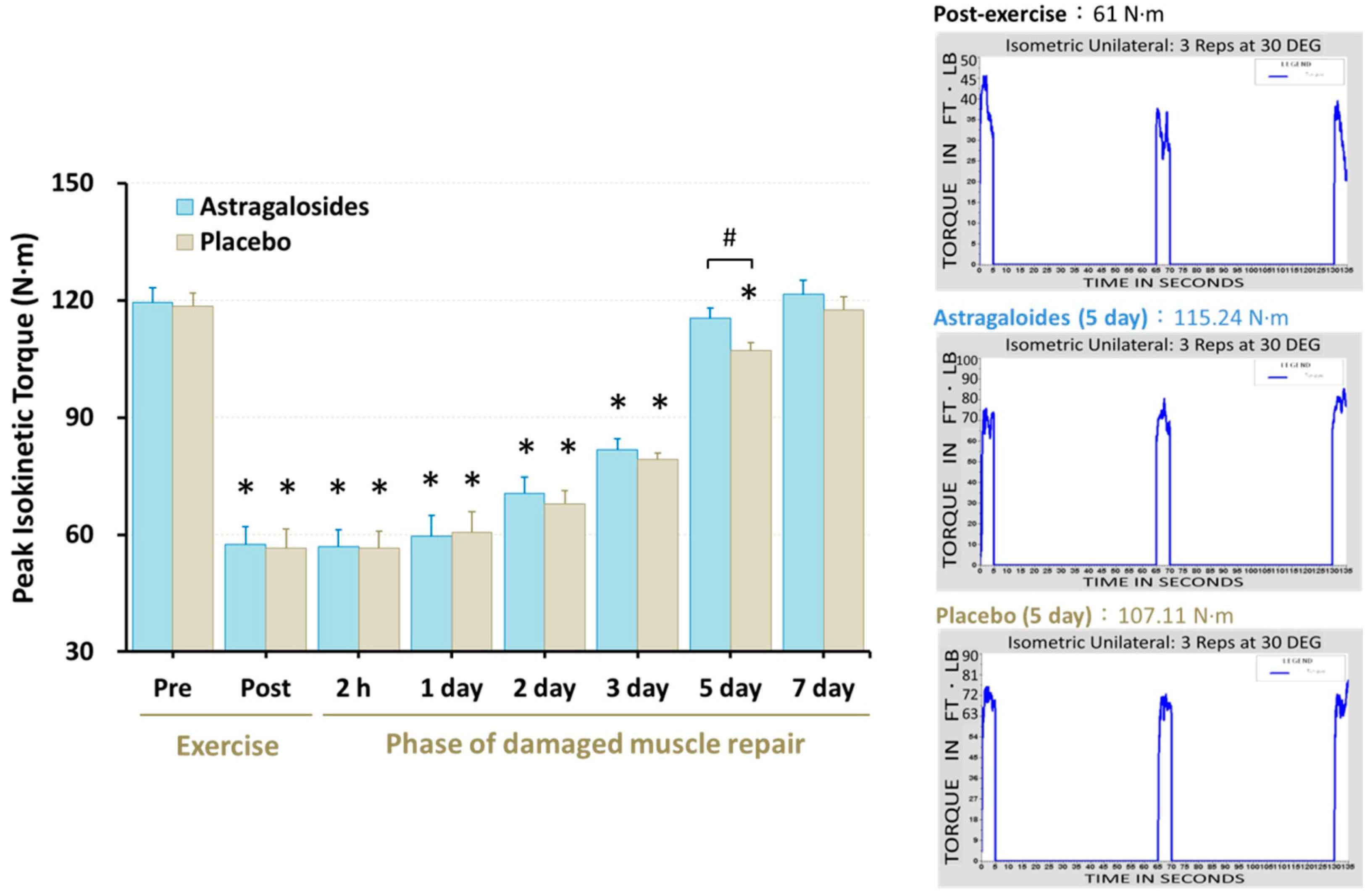
3.7. Recovery of Muscle Function
4. Discussion
4.1. Astragalosides Supplementation and Chemical Analysis
4.2. Astragalosides Supplementation and Muscle Damage Markers
4.3. Astragalosides Supplementation and Inflammatory Responses
4.4. Astragalosides Supplementation and Early Stage of Muscular Repair
4.5. Astragalosides Supplementation and Muscle Strength Performance
4.6. Astragalosides Supplementation and Muscle Respiratory Functional Performance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sciorati, C.; Rigamonti, E.; Manfredi, A.A.; Rovere-Querini, P. Cell death, clearance and immunity in the skeletal muscle. Cell Death Differ. 2016, 23, 927–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schertzer, J.D.; Lynch, G.S. Comparative evaluation of IGF-I gene transfer and IGF-I protein administration for enhancing skeletal muscle regeneration after injury. Gene Ther. 2006, 13, 1657–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, S.; Ren, H.; Li, Y.; Zhou, J.; Duan, C.; Lu, L. Differential regulation of IGF-I and IGF-II gene expression in skeletal muscle cells. Mol. Cell. Biochem. 2013, 373, 107–113. [Google Scholar] [CrossRef]
- Zanou, N.; Gailly, P. Skeletal muscle hypertrophy and regeneration: Interplay between the myogenic regulatory factors (MRFs) and insulin-like growth factors (IGFs) pathways. Cell. Mol. Life Sci. 2013, 70, 4117–4130. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Lázaro, D.; Mielgo-Ayuso, J.; Calvo, J.S.; Martínez, A.C.; García, A.C.; Fernandez-Lazaro, C.I. Modulation of exercise-induced muscle Damage, Inflammation, and Oxidative Markers by Curcumin Supplementation in a physically active population: A systematic review. Nutrients 2020, 12, 501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupuy, O.; Douzi, W.; Theurot, D.; Bosquet, L.; Dugué, B. An evidence-based approach for choosing post-exercise recovery techniques to reduce markers of muscle damage, soreness, fatigue, and inflammation: A systematic review with meta-analysis. Front. Physiol. 2018, 9, 403. [Google Scholar] [CrossRef]
- Paulsen, G.; Mikkelsen, U.R.; Raastad, T.; Peake, J.M. Leucocytes, cytokines and satellite cells: What role do they play in muscle damage and regeneration following eccentric exercise? Exerc. Immunol. Rev. 2012, 18, 42–97. [Google Scholar] [PubMed]
- Peake, J.M.; Neubauer, O.; Gatta, P.A.D.; Nosaka, K. Muscle damage and inflammation during recovery from exercise. J. Appl. Physiol. 2017, 122, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Reid, W.D.; Clarke, T.J.; Wallace, A.M. Respiratory muscle injury: Evidence to date and potential mechanisms. Can. J. Appl. Physiol. 2001, 26, 356–387. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, J.A.; La Gerche, A.; Hull, J.H. Is the healthy respiratory system built just right, overbuilt, or underbuilt to meet the demands imposed by exercise? J. Appl. Physiol. 2020, 129, 1235–1256. [Google Scholar] [CrossRef]
- Owens, D.J.; Twist, C.; Cobley, J.N.; Howatson, G.; Close, G.L. Exercise-induced muscle damage: What is it, what causes it and what are the nutritional solutions? Eur. J. Sport Sci. 2019, 19, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, Y.; Fujii, N.; Suzuki, K. Dietary supplementation for attenuating exercise-induced muscle damage and delayed-onset muscle soreness in humans. Nutrients 2021, 14, 70. [Google Scholar] [CrossRef] [PubMed]
- Simeonova, R.; Bratkov, V.M.; Kondeva-Burdina, M.; Vitcheva, V.; Manov, V.; Krasteva, I. Experimental liver protection of n-butanolic extract of Astragalus monspessulanus L. on carbon tetrachloride model of toxicity in rat. Redox Rep. 2015, 20, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Wang, R.; Jiang, W.; Wang, H.; Xu, A.; Lu, G.; Ren, Y.; Xu, Y.; Song, Y.; Yong, S.; et al. Protective effect of astragaloside IV against paraquat-induced lung injury in mice by suppressing Rho signaling. Inflammation 2016, 39, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Luo, Y.; Zou, C.; Huang, L.; Tian, R.; Lu, Z. Effect of astragaloside IV on indoxyl sulfate-induced kidney injury in mice via attenuation of oxidative stress. BMC Pharmacol. Toxicol. 2018, 19, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, T.S.; Lei, T.H.; Liu, J.F.; Hsu, M.C. Astragalus membranaceus enhances myotube hypertrophy through PI3K-mediated Akt/mTOR signaling phosphorylation. Nutrients 2022, 14, 1670. [Google Scholar] [CrossRef]
- Yeh, T.S.; Chuang, H.L.; Huang, W.C.; Chen, Y.M.; Huang, C.C.; Hsu, M.C. Astragalus membranaceus improves exercise performance and ameliorates exercise-induced fatigue in trained mice. Molecules 2014, 19, 2793–2807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vardiman, J.P.; Moodie, N.; Siedlik, J.A.; Kudrna, R.A.; Graham, Z.; Gallagher, P. Short-wave diathermy pretreatment and inflammatory myokine response after high-intensity eccentric exercise. J. Athl. Train. 2015, 50, 612–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philippou, A.; Bogdanis, G.C.; Nevill, A.M.; Maridaki, M. Changes in the angle-force curve of human elbow flexors following eccentric and isometric exercise. Eur. J. Appl. Physiol. 2004, 93, 237–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado, D.A.; Lambert, B.S.; Boutris, N.; McCulloch, P.C.; Robbins, A.B.; Moreno, M.R.; Harris, J.D. Validation of digital visual analog scale pain scoring with a traditional paper-based visual analog scale in adults. J. Am. Acad. Orthop. Surg. Glob. Res. Rev. 2018, 2, e088. [Google Scholar] [CrossRef] [PubMed]
- Nemoto, E.M. Near-infrared spectroscopy (NIRS) of muscle HbO2 and MbO2 desaturation during exercise. Adv. Exp. Med. Biol. 2021, 1269, 83–86. [Google Scholar] [PubMed]
- Gurley, K.; Shang, Y.; Yu, G. Noninvasive optical quantification of absolute blood flow, blood oxygenation, and oxygen consumption rate in exercising skeletal muscle. J. Biomed. Opt. 2012, 17, 075010. [Google Scholar] [PubMed] [Green Version]
- Qi, Y.; Gao, F.; Hou, L.; Wan, C. Anti-Inflammatory and Immunostimulatory Activities of Astragalosides. Am. J. Chin. Med. 2017, 45, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, C.; Gao, L.; Du, G.; Qin, X. Astragaloside IV derived from Astragalus membranaceus: A research review on the pharmacological effects. Adv. Pharmacol. 2020, 87, 89–112. [Google Scholar]
- Li, L.; Hou, X.; Xu, R.; Liu, C.; Tu, M. Research review on the pharmacological effects of astragaloside IV. Fundam. Clin. Pharmacol. 2017, 31, 17–36. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Zhang, H.; Mu, Y.; Sun, M.; Liu, P. Pharmacological effects of Astragaloside IV: A literature review. J. Tradit. Chin. Med. 2013, 33, 413–416. [Google Scholar] [CrossRef]
- Cheng, X.; Wei, B.; Sun, L.; Hu, X.; Liang, J.; Chen, Y. Astragaloside I Stimulates Osteoblast Differentiation Through the Wnt/β-catenin Signaling Pathway. Phytother. Res. 2016, 30, 1680–1688. [Google Scholar] [CrossRef]
- Guo, T.; Liu, Z.L.; Zhao, Q.; Zhao, Z.M.; Liu, C.H. A combination of astragaloside I, levistilide A and calycosin exerts anti-liver fibrosis effects in vitro and in vivo. Acta Pharmacol. Sin. 2018, 39, 1483–1492. [Google Scholar] [CrossRef] [Green Version]
- Wan, C.P.; Gao, L.X.; Hou, L.F.; Yang, X.Q.; He, P.L.; Yang, Y.F.; Tang, W.; Yue, J.M.; Li, J.; Zuo, J.P. Astragaloside II triggers T cell activation through regulation of CD45 protein tyrosine phosphatase activity. Acta Pharmacol. Sin. 2013, 34, 522–530. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Tsai, W.C.; Lin, J.C.; Ahmetaj-Shala, B.; Huang, S.F.; Chang, W.L.; Chang, T.C. Astragaloside II promotes intestinal epithelial repair by enhancing L-arginine uptake and activating the mTOR pathway. Sci. Rep. 2017, 7, 12302. [Google Scholar] [CrossRef] [Green Version]
- Ross, M.L.; Halson, S.L.; Suzuki, K.; Garnham, A.; Hawley, J.A.; Cameron-Smith, D.; Peake, J.M. Cytokine responses to carbohydrate ingestion during recovery from exercise-induced muscle injury. J. Interferon Cytokine Res. 2010, 30, 329–337. [Google Scholar] [CrossRef]
- Ciciliot, S.; Schiaffino, S. Regeneration of mammalian skeletal muscle. Basic mechanisms and clinical implications. Curr. Pharm. Des. 2010, 16, 906–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, T.; Delafontaine, P. Mechanisms of IGF-1-mediated regulation of skeletal muscle hypertrophy and atrophy. Cells 2020, 9, 1970. [Google Scholar] [CrossRef]
- Wosczyna, M.N.; Rando, T.A. A Muscle Stem Cell Support Group: Coordinated Cellular Responses in Muscle Regeneration. Dev. Cell 2018, 46, 135–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonkin, J.; Temmerman, L.; Sampson, R.D.; Gallego-Colon, E.; Barberi, L.; Bilbao, D.; Schneider, M.D.; Musarò, A.; Rosenthal, N. Monocyte/Macrophage-derived IGF-1 Orchestrates Murine Skeletal Muscle Regeneration and Modulates Autocrine Polarization. Mol. Ther. 2015, 23, 1189–1200. [Google Scholar] [CrossRef] [Green Version]
- Saclier, M.; Yacoub-Youssef, H.; Mackey, A.L.; Arnold, L.; Ardjoune, H.; Magnan, M.; Sailhan, F.; Chelly, J.; Pavlath, G.K.; Mounier, R.; et al. Differentially activated macrophages orchestrate myogenic precursor cell fate during human skeletal muscle regeneration. Stem Cells 2013, 31, 384–396. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Gu, W.; Shi, Y.; Chen, Y.; Tan, Y. Protective effects of astragaloside IV on IL-8-treated diaphragmatic muscle cells. Exp. Ther. Med. 2019, 17, 519–524. [Google Scholar] [CrossRef]
- McKay, B.R.; O’Reilly, C.E.; Phillips, S.M.; Tarnopolsky, M.A.; Parise, G. Co-expression of IGF-1 family members with myogenic regulatory factors following acute damaging muscle-lengthening contractions in humans. J. Physiol. 2008, 586, 5549–5560. [Google Scholar] [CrossRef]
- Velloso, C.P. Regulation of muscle mass by growth hormone and IGF-I. Br. J. Pharmacol. 2008, 154, 557–568. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.S.; Ahmad, K.; Lee, E.J.; Lee, Y.H.; Choi, I. Implications of Insulin-Like Growth Factor-1 in Skeletal Muscle and Various Diseases. Cells 2020, 9, 1773. [Google Scholar] [CrossRef]
- Jarmusch, S.; Baber, L.; Bidlingmaier, M.; Ferrari, U.; Hofmeister, F.; Hintze, S.; Mehaffey, S.; Meinke, P.; Neuerburg, C.; Schoser, B.; et al. Influence of IGF-I serum concentration on muscular regeneration capacity in patients with sarcopenia. BMC Musculoskelet. Disord. 2021, 22, 807. [Google Scholar] [CrossRef]
- Wu, H.L.; Zhou, H.J. Astragalus membranaceus promote expression of insulin-like growth factor 1 in rat model of olivo-cerebellar degeneration. Zhongguo Zhong Yao Za Zhi 2007, 32, 242–245. [Google Scholar] [PubMed]
- Florini, J.R.; Magri, K.A.; Ewton, D.Z.; James, P.L.; Grindstaff, K.; Rotwein, P.S. “Spontaneous” differentiation of skeletal myoblasts is dependent upon autocrine secretion of insulin-like growth factor-II. J. Biol. Chem. 1991, 266, 15917–15923. [Google Scholar] [CrossRef]
- Levinovitz, A.; Jennische, E.; Oldfors, A.; Edwall, D.; Norstedt, G. Activation of insulin-like growth factor II expression during skeletal muscle regeneration in the rat: Correlation with myotube formation. Mol. Endocrinol. 1992, 6, 1227–1234. [Google Scholar]
- Erbay, E.; Park, I.H.; Nuzzi, P.D.; Schoenherr, C.J.; Chen, J. IGF-II transcription in skeletal myogenesis is controlled by mTOR and nutrients. J. Cell Biol. 2003, 163, 931–936. [Google Scholar] [CrossRef] [Green Version]
- Michaut, A.; Pousson, M.; Millet, G.; Belleville, J.; Van Hoecke, J. Maximal voluntary eccentric, isometric and concentric torque recovery following a concentric isokinetic exercise. Int. J. Sports Med. 2003, 24, 51–56. [Google Scholar] [CrossRef]
- Rhodes, D.; McNaughton, L.; Greig, M. The temporal pattern of recovery in eccentric hamstring strength post-soccer specific fatigue. Res. Sports Med. 2019, 27, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, G.; Crameri, R.; Benestad, H.B.; Fjeld, J.G.; Mørkrid, L.; Hallén, J.; Raastad, T. Time course of leukocyte accumulation in human muscle after eccentric exercise. Med. Sci. Sports Exerc. 2010, 42, 75–85. [Google Scholar] [CrossRef]
- Broome, S.; Atiola, R.D.; Braakhuis, A.; Mitchell, C.; Merry, T.L. Mitochondria-targeted antioxidant supplementation does not affect muscle soreness or recovery of maximal voluntary isometric contraction force following muscle-damaging exercise in untrained men: A randomised clinical trial. Appl. Physiol. Nutr. Metab. 2022, 47, 762–774. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Chiesa, S.T.; Chaturvedi, N.; Hughes, A.D. Recent developments in near-infrared spectroscopy (NIRS) for the assessment of local skeletal muscle microvascular function and capacity to utilise oxygen. Artery Res. 2016, 16, 25–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barstow, T.J. Understanding near infrared spectroscopy and its application to skeletal muscle research. J. Appl. Physiol. 2019, 126, 1360–1376. [Google Scholar] [CrossRef]
- Cornelis, N.; Chatzinikolaou, P.; Buys, R.; Fourneau, I.; Claes, J.; Cornelissen, V. The use of near infrared spectroscopy to evaluate the effect of exercise on peripheral muscle oxygenation in patients with lower extremity artery disease: A systematic review. Eur. J. Vasc. Endovasc. Surg. 2021, 61, 837–847. [Google Scholar] [CrossRef]
- Bailey, S.J.; Blackwell, J.R.; Williams, E.; Vanhatalo, A.; Wylie, L.J.; Winyard, P.G.; Jones, A.M. Two weeks of watermelon juice supplementation improves nitric oxide bioavailability but not endurance exercise performance in humans. Nitric. Oxide 2016, 59, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Theodorou, A.A.; Zinelis, P.T.; Malliou, V.J.; Chatzinikolaou, P.N.; Margaritelis, N.V.; Mandalidis, D.; Geladas, N.D.; Paschalis, V. Acute L-citrulline supplementation increases nitric oxide bioavailability but not inspiratory muscle oxygenation and respiratory performance. Nutrients 2021, 13, 3311. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.L.; Chen, X.J.; Ji, H.; Li, P.; Bian, Y.Y.; Yang, D.; Xu, J.D.; Bian, Z.P.; Zhang, J.N. Astragaloside IV improved intracellular calcium handling in hypoxia-reoxygenated cardiomyocytes via the sarcoplasmic reticulum Ca-ATPase. Pharmacology 2008, 81, 325–332. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | (n = 11) | |
|---|---|---|
| Age (year) | 23 ± 0.9 | |
| Body weight (Kg) | 73.4 ± 1.9 | |
| Height (m) | 1.75 ± 0.02 | |
| Body mass index (kg/m2) | 24.0 ± 0.7 | |
| Leg volume (liters) | 7.5 ± 0.3 | |
| 1RM leg press (kg) | 217.7 ± 7.0 | |
| 1RM leg extension (kg) | 120.2 ± 3.4 | |
| Astragalosides period | Placebo period | |
| Total energy (Kcal) | 1987 ± 65 | 1984 ± 77 |
| Carbohydrate (% of energy) | 51.4 ± 2.3 | 51.6 ± 2.2 |
| Fat (% of energy) | 30.8 ± 1.4 | 30.6 ± 1.3 |
| Protein (% of energy) | 17.8 ± 1.2 | 17.8 ± 1.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeh, T.-S.; Lei, T.-H.; Barnes, M.J.; Zhang, L. Astragalosides Supplementation Enhances Intrinsic Muscle Repair Capacity Following Eccentric Exercise-Induced Injury. Nutrients 2022, 14, 4339. https://doi.org/10.3390/nu14204339
Yeh T-S, Lei T-H, Barnes MJ, Zhang L. Astragalosides Supplementation Enhances Intrinsic Muscle Repair Capacity Following Eccentric Exercise-Induced Injury. Nutrients. 2022; 14(20):4339. https://doi.org/10.3390/nu14204339
Chicago/Turabian StyleYeh, Tzu-Shao, Tze-Huan Lei, Matthew J. Barnes, and Lei Zhang. 2022. "Astragalosides Supplementation Enhances Intrinsic Muscle Repair Capacity Following Eccentric Exercise-Induced Injury" Nutrients 14, no. 20: 4339. https://doi.org/10.3390/nu14204339
APA StyleYeh, T. -S., Lei, T. -H., Barnes, M. J., & Zhang, L. (2022). Astragalosides Supplementation Enhances Intrinsic Muscle Repair Capacity Following Eccentric Exercise-Induced Injury. Nutrients, 14(20), 4339. https://doi.org/10.3390/nu14204339