

The Immunological Role of Milk Fat Globule Membrane

Abstract
:
1. Introduction
2. Composition of MFGM
3. Immuno-Metabolism of MFGM
3.1. Gastric Environment
3.2. Small and Large Intestines
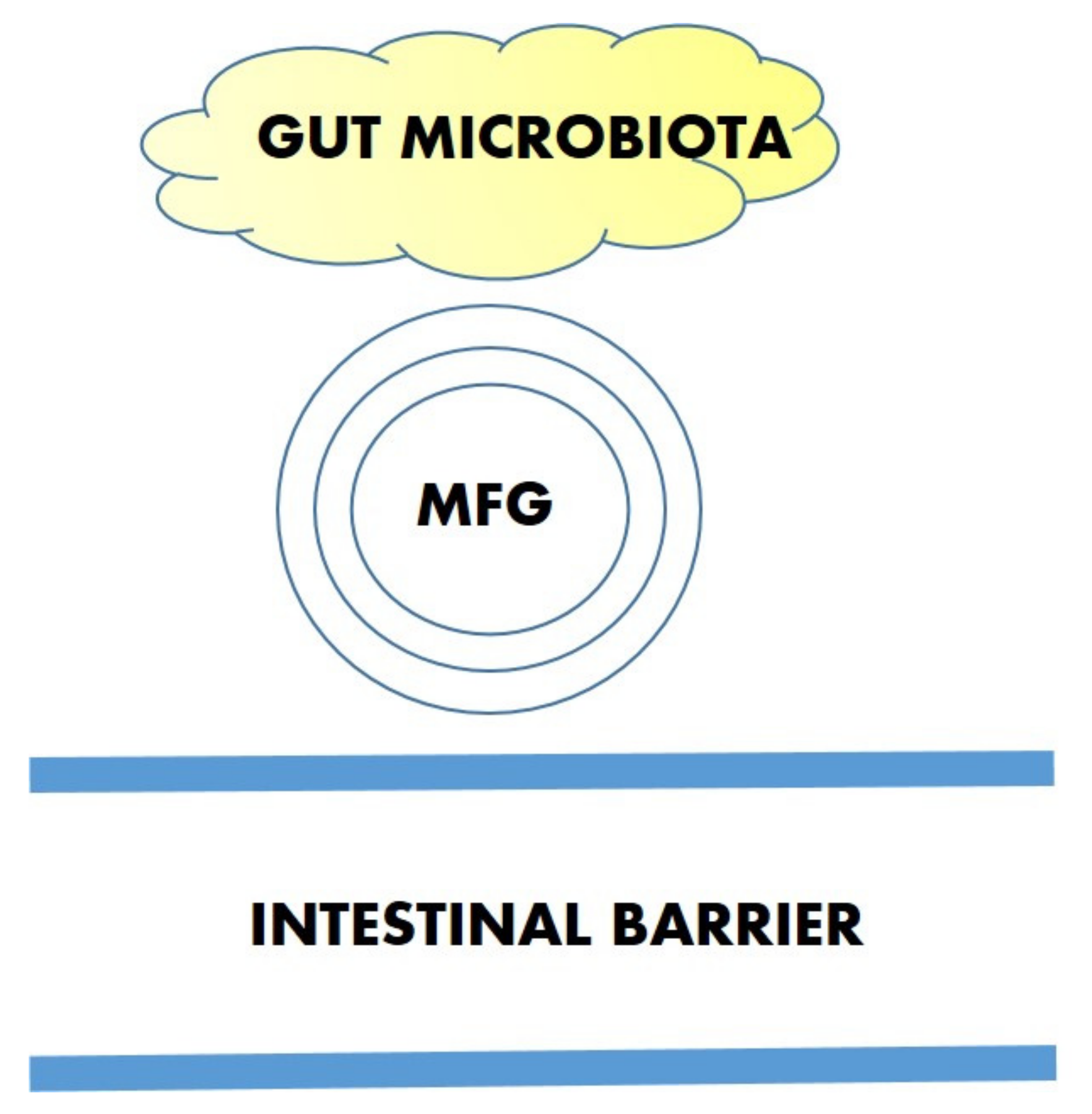
3.3. Interaction with the Microbiota
3.4. Formula Milk Added with Bovine MFGM, an Important Supplement
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Recommendations on Post-Natal Care of the Mother and Newborn; World Health Organization: Geneva, Switzerland, 2013. [Google Scholar]
- Brink, L.R.; Lönnerdal, B. Milk fat globule membrane: The role of its various components in infant health and development. J. Nutr. Biochem. 2020, 85, 108465. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.B.; Nickel, N.C.; Bode, L.; Brockway, M.; Brown, A.; Chambers, C.; Goldhammer, C.; Hinde, K.; McGuire, M.; Munblit, D.; et al. Breastfeeding and the origins of health: Interdisciplinary perspectives and priorities. Matern. Child Nutr. 2021, 17, e13109. [Google Scholar] [CrossRef] [PubMed]
- Noel, G.; In, J.G.; Lemme-Dumit, J.M.; DeVine, L.R.; Cole, R.N.; Guerrerio, A.L.; Campbell, J.D.; Kovbasnjuk, O.; Pasetti, M.F. Human Breast Milk Enhances Intestinal Mucosal Barrier Function and Innate Immunity in a Healthy Pediatric Human Enteroid Model. Front. Cell Dev. Biol. 2021, 9, 685171. [Google Scholar] [CrossRef] [PubMed]
- Cavaletto, M.; Giuffrida, M.G.; Conti, A. The proteomic approach to analysis of human milk fat globule membrane. Clin. Chim. Acta 2004, 347, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Ballard, O.; Morrow, A.L. Human milk composition: Nutrients and bioactive factors. Pediatr. Clin. 2013, 60, 49–74. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Parenti, M.; Grip, T.; Domellöf, M.; Lönnerdal, B.; Hernell, O.; Timby, N.; Slupsky, C.M. Metabolic phenotype of breast-fed infants, and infants fed standard formula or bovine MFGM supplemented formula: A randomized controlled trial. Sci. Rep. 2019, 9, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, L.A.; Kishton, R.J.; Rathmell, J. A guide to immunometabolism for immunologists. Nat. Rev. Immunol. 2016, 16, 553–565. [Google Scholar] [CrossRef] [Green Version]
- Arts, R.J.W.; Carvalho, A.; La Rocca, C.; Palma, C.; Rodrigues, F.; Silvestre, R.; Kleinnijenhuis, J.; Lachmandas, E.; Gonçalves, L.G.; Belinha, A.; et al. Immunometabolic Pathways in BCG-Induced Trained Immunity. Cell Rep. 2016, 17, 2562–2571. [Google Scholar] [CrossRef] [Green Version]
- Conti, M.G.; Angelidou, A.; Diray-Arce, J.; Smolen, K.K.; Lasky-Su, J.; De Curtis, M.; Levy, O. Immunometabolic approaches to prevent, detect, and treat neonatal sepsis. Pediatr. Res. 2020, 87, 399–405. [Google Scholar] [CrossRef]
- Ramalho, R.; Rao, M.; Zhang, C.; Agrati, C.; Ippolito, G.; Wang, F.S.; Zumla, A.; Maeurer, M. Immunometabolism: New insights and lessons from antigen-directed cellular immune responses. Semin. Immunopathol. 2020, 42, 279–313. [Google Scholar] [CrossRef]
- Lisboa, P.C.; Miranda, R.A.; Souza, L.L.; Moura, E.G. Can breastfeeding affect the rest of our life? Neuropharmacology 2021, 200, 108821. [Google Scholar] [CrossRef] [PubMed]
- Giuffrida, F.; Cruz-Hernandez, C.; Flück, B.; Tavazzi, I.; Thakkar, S.K.; Destaillats, F.; Braun, M. Quantification of phospholipids classes in human milk. Lipids 2013, 48, 1051–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rueda, R. The role of dietary gangliosides on immunity and the prevention of infection. Br. J. Nutr. 2007, 98 (Suppl. S1), S68–S73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thum, C.; Wall, C.; Day, L.; Szeto, I.M.Y.; Li, F.; Yan, Y.; Barnett, M.P.G. Changes in Human Milk Fat Globule Composition Throughout Lactation: A Review. Front. Nutr. 2022, 9, 835856. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, C.; Franco, L.; Regal, P.; Lamas, A.; Cepeda, A.; Fente, C. Breast milk: A source of functional compounds with potential application in nutrition and therapy. Nutrients 2021, 13, 1026. [Google Scholar] [CrossRef]
- Chai, C.; Oh, S.; Imm, J.Y. Roles of Milk Fat Globule Membrane on Fat Digestion and Infant Nutrition. Food Sci. Anim. Resour. 2022, 42, 351–371. [Google Scholar] [CrossRef]
- Zou, X.Q.; Guo, Z.; Huang, J.H.; Jin, Q.Z.; Cheong, L.Z.; Wang, X.G.; Xu, X.B. Human milk fat globules from different stages of lactation: A lipid composition analysis and microstructure characterization. J. Agric. Food Chem. 2012, 60, 7158–7167. [Google Scholar] [CrossRef]
- Manoni, M.; Di Lorenzo, C.; Ottoboni, M.; Tretola, M.; Pinotti, L. Comparative Proteomics of Milk Fat Globule Membrane (MFGM) Proteome across Species and Lactation Stages and the Potentials of MFGM Fractions in Infant Formula Preparation. Foods 2020, 9, 1251. [Google Scholar] [CrossRef]
- Lu, J.; Wang, X.; Zhang, W.; Liu, L.; Pang, X.; Zhang, S.; Lv, J. Comparative proteomics of milk fat globule membrane in different species reveals variations in lactation and nutrition. Food Chem. 2016, 196, 665–672. [Google Scholar] [CrossRef]
- UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Cattaneo, C.; Caramaschi, A.; Uga, E.; Braghin, M.; Cosi, G.; Peila, C.; Strozzi, M.C.; Sabatini, M.; Gazzolo, D.; Manfredi, M.; et al. Analysis of Toll-Like Receptors in Human Milk: Detection of Membrane-Bound and Soluble Forms. J. Immunol. Res. 2019, 2019, 4078671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timby, N.; Hernell, O.; Vaarala, O.; Melin, M.; Lönnerdal, B.; Domellöf, M. Infections in infants fed formula supplemented with bovine milk fat globule membranes. J. Pediatr. Gastroenterol. Nutr. 2015, 60, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Harris, D.L.; Weston, P.J.; Harding, J.E. Lactate, rather than ketones, may provide alternative cerebral fuel in hypoglycaemic newborns. Arch. Dis. Child. Fetal Neonatal Ed. 2015, 100, F161–F164. [Google Scholar] [CrossRef] [PubMed]
- Cahill, G.F., Jr. Fuel metabolism in starvation. Annu. Rev. Nutr. 2006, 26, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Slupsky, C.M.; Heckmann, A.B.; Christensen, B.; Peng, Y.; Li, X.; Hernell, O.; Lönnerdal, B.; Li, Z. Milk Fat Globule Membrane as a Modulator of Infant Metabolism and Gut Microbiota: A Formula Supplement Narrowing the Metabolic Differences between Breastfed and Formula-Fed Infants. Mol. Nutr. Food Res. 2021, 65, e2000603. [Google Scholar] [CrossRef]
- Le, T.T.; Van de Wiele, T.; Do, T.N.; Debyser, G.; Struijs, K.; Devreese, B.; Dewettinck, K.; Van Camp, J. Stability of milk fat globule membrane proteins toward human enzymatic gastrointestinal digestion. J. Dairy Sci. 2012, 95, 2307–2318. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Lin, S.; Ma, S.; Sun, Y.; Li, X.; Liang, S. A Comparative Analysis of Lipid Digestion in Human Milk and Infant Formulas Based on Simulated In Vitro Infant Gastrointestinal Digestion. Foods. 2022, 11, 200. [Google Scholar] [CrossRef]
- Nilsson, Å. Role of Sphingolipids in Infant Gut Health and Immunity. J. Pediatr. 2016, 173, S53–S59. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Padhi, E.; Hasegawa, Y.; Larke, J.; Parenti, M.; Wang, A.; Hernell, O.; Lönnerdal, B.; Slupsky, C. Compositional Dynamics of the Milk Fat Globule and Its Role in Infant Development. Front. Pediatr. 2018, 6, 313. [Google Scholar] [CrossRef] [Green Version]
- Jiang, R.; Du, X.; Brink, L.; Lönnerdal, B. The role of orally ingested milk fat globule membrane on intestinal barrier functions evaluated with a suckling rat pup supplementation model and a human enterocyte model. J. Nutr. Biochem. 2022, 108, 109084. [Google Scholar] [CrossRef]
- Gallier, S.; Zhu, X.Q.; Rutherfurd, S.M.; Ye, A.; Moughan, P.J.; Singh, H. In vivo digestion of bovine milk fat globules: Effect of processing and interfacial structural changes. II. Upper digestive tract digestion. Food Chem. 2013, 141, 3215–3223. [Google Scholar] [CrossRef] [PubMed]
- Socha-Banasiak, A.; Pawłowska, M.; Czkwianianc, E.; Pierzynowska, K. From Intrauterine to Extrauterine Life-The Role of Endogenous and Exogenous Factors in the Regulation of the Intestinal Microbiota Community and Gut Maturation in Early Life. Front. Nutr. 2021, 8, 696966. [Google Scholar] [CrossRef] [PubMed]
- Younge, N.; McCann, J.R.; Ballard, J.; Plunkett, C.; Akhtar, S.; Araújo-Pérez, F.; Murtha, A.; Brandon, D.; Seed, P.C. Fetal exposure to the maternal microbiota in humans and mice. JCI Insight. 2019, 4, e127806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Guryn, K.; Hubert, N.; Frazier, K.; Urlass, S.; Musch, M.W.; Ojeda, P.; Pierre, J.F.; Miyoshi, J.; Sontag, T.J.; Cham, C.M.; et al. Small Intestine Microbiota Regulate Host Digestive and Absorptive Adaptive Responses to Dietary Lipids. Cell Host Microbe. 2018, 23, 458–469.e5. [Google Scholar] [CrossRef] [Green Version]
- Torow, N.; Marsland, B.J.; Hornef, M.W.; Gollwitzer, E.S. Neonatal mucosal immunology. Mucosal Immunol. 2017, 10, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Denning, T.L.; Bhatia, A.M.; Kane, A.F.; Patel, R.M.; Denning, P.W. Pathogenesis of NEC: Role of the innate and adaptive immune response. Semin. Perinatol. 2017, 41, 15–28. [Google Scholar] [CrossRef] [Green Version]
- Tirone, C.; Pezza, L.; Paladini, A.; Tana, M.; Aurilia, C.; Lio, A.; D’Ippolito, S.; Tersigni, C.; Posteraro, B.; Sanguinetti, M.; et al. Gut and Lung Microbiota in Preterm Infants: Immunological Modulation and Implication in Neonatal Outcomes. Front. Immunol. 2019, 10, 2910. [Google Scholar] [CrossRef] [Green Version]
- Garg, P.M.; Pippin, M.; Zhang, M.; Ware, J.; Nelin, S.; Paschal, J.; Varshney, N.; Hillegass, W.B. Clinical Correlates of Moderate-to-Severe Bronchopulmonary Dysplasia in Preterm Infants following Surgical Necrotizing Enterocolitis. Am. J. Perinatol. 2022. [Google Scholar] [CrossRef]
- Ma, B.; Sundararajan, S.; Nadimpalli, G.; France, M.; McComb, E.; Rutt, L.; Lemme-Dumit, J.M.; Janofsky, E.; Roskes, L.S.; Gajer, P.; et al. Highly Specialized Carbohydrate Metabolism Capability in Bifidobacterium Strains Associated with Intestinal Barrier Maturation in Early Preterm Infants. mBio 2022, 13, e0129922. [Google Scholar] [CrossRef]
- Mohamed, H.J.J.; Lee, E.K.H.; Woo, K.C.K.; Sarvananthan, R.; Lee, Y.Y.; Mohd Hussin, Z.A. Brain-immune-gut benefits with early life supplementation of milk fat globule membrane. JGH Open 2022, 6, 454–461. [Google Scholar] [CrossRef]
- Mou, Y.; Du, Y.; Zhou, L.; Yue, J.; Hu, X.; Liu, Y.; Chen, S.; Lin, X.; Zhang, G.; Xiao, H.; et al. Gut Microbiota Interact With the Brain Through Systemic Chronic Inflammation: Implications on Neuroinflammation, Neurodegeneration, and Aging. Front. Immunol. 2022, 13, 796288. [Google Scholar] [CrossRef] [PubMed]
- Owino, V.; Ahmed, T.; Freemark, M.; Kelly, P.; Loy, A.; Manary, M.; Loechl, C. Environmental Enteric Dysfunction and Growth Failure/Stunting in Global Child Health. Pediatrics 2016, 138, e20160641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Hakim, Y.; Bake, S.; Mani, K.K.; Sohrabji, F. Impact of intestinal disorders on central and peripheral nervous system diseases. Neurobiol. Dis. 2022, 165, 105627. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Li, Y.; Niu, Y.; Tang, Q.; Wu, J. Milk Fat Globule Membrane Enhances Colonic-Mucus-Barrier Function in a Rat Model of Short-Bowel Syndrome. J. Parenter. Enteral Nutr. 2021, 45, 916–925. [Google Scholar] [CrossRef]
- Anto, L.; Warykas, S.W.; Torres-Gonzalez, M.; Blesso, C.N. Milk Polar Lipids: Underappreciated Lipids with Emerging Health Benefits. Nutrients 2020, 12, 1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.; Hosozawa, M.; Kudo, N.; Yoshikawa, N.; Hisata, K.; Shoji, H.; Shinohara, K.; Shimizu, T. The pilot study: Sphingomyelin-fortified milk hashas a positive association with the neurobehavioural development of very low birth weight infants during infancy, randomized control trial. Brain Dev. 2013, 35, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Qi, C.; Wang, X.; Jin, Q.; McClements, D.J. Influence of Homogenization and Thermal Processing on the Gastrointestinal Fate of Bovine Milk Fat: In Vitro Digestion Study. J. Agric. Food Chem. 2017, 65, 11109–11117. [Google Scholar] [CrossRef]
- Ambrożej, D.; Dumycz, K.; Dziechciarz, P.; Ruszczyński, M. Milk Fat Globule Membrane Supplementation in Children: Systematic Review with Meta-Analysis. Nutrients 2021, 13, 714. [Google Scholar] [CrossRef]
- Mozaffarian, D. Dairy Foods, Obesity, and Metabolic Health: The Role of the Food Matrix Compared with Single Nutrients. Adv. Nutr. 2019, 10, 917S–923S. [Google Scholar] [CrossRef]
- Lind, T.; Johansson, U.; Öhlund, I.; Lindberg, L.; Lönnerdal, B.; Tennefors, C.; Hernell, O. Study protocol: Optimized complementary feeding study (OTIS): A randomized controlled trial of the impact of a protein-reduced complementary diet based on Nordic foods. BMC Public Health 2019, 19, 134. [Google Scholar] [CrossRef]
- Toro-Campos, R.; Algarín, C.; Peirano, P.; Peña, M.; Murguia-Peniche, T.; Wu, S.S.; Uauy, R. Effect of feeding mode on infant growth and cognitive function: Study protocol of the Chilean infant Nutrition randomized controlled Trial (ChiNuT). BMC Pediatr. 2020, 20, 225. [Google Scholar] [CrossRef] [PubMed]
- Zavaleta, N.; Kvistgaard, A.S.; Graverholt, G.; Respicio, G.; Guija, H.; Valencia, N.; Lönnerdal, B. Efficacy of an MFGM-enriched complementary food in diarrhea, anemia, and micronutrient status in infants. J. Pediatr. Gastroenterol. Nutr. 2011, 53, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Zavaleta, N.; Chen, S.Y.; Lönnerdal, B.; Slupsky, C. Effect of bovine milk fat globule membranes as a complementary food on the serum metabolome and immune markers of 6-11-month-old Peruvian infants. NPJ Sci. Food. 2018, 2, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurnida, D.A.; Rowan, A.M.; Idjradinata, P.; Muchtadi, D.; Sekarwana, N. Association of complex lipids containing gangliosides with cognitive development of 6-month-old infants. Early Hum. Dev. 2012, 88, 595–601. [Google Scholar] [CrossRef]
- Veereman-Wauters, G.; Staelens, S.; Rombaut, R.; Dewettinck, K.; Deboutte, D.; Brummer, R.J.; Boone, M.; Le Ruyet, P. Milk fat globule membrane (INPULSE) enriched formula milk decreases febrile episodes and may improve behavioral regulation in young children. Nutrition 2012, 28, 749–752. [Google Scholar] [CrossRef]
- Poppitt, S.D.; McGregor, R.A.; Wiessing, K.R.; Goyal, V.K.; Chitkara, A.J.; Gupta, S.; Palmano, K.; Kuhn-Sherlock, B.; McConnell, M.A. Bovine complex milk lipid containing gangliosides for prevention of rotavirus infection and diarrhoea in northern Indian infants. J. Pediatr. Gastroenterol. Nutr. 2014, 59, 167–171. [Google Scholar] [CrossRef] [Green Version]
- Timby, N.; Domellöf, E.; Hernell, O.; Lönnerdal, B.; Domellöf, M. Neurodevelopment, nutrition, and growth until 12 mo of age in infants fed a low-energy, low-protein formula supplemented with bovine milk fat globule membranes: A randomized controlled trial. Am. J. Clin. Nutr. 2014, 99, 860–868. [Google Scholar] [CrossRef] [Green Version]
- Timby, N.; Lönnerdal, B.; Hernell, O.; Domellöf, M. Cardiovascular risk markers until 12 mo of age in infants fed a formula supplemented with bovine milk fat globule membranes. Pediatr. Res. 2014, 76, 394–400. [Google Scholar] [CrossRef] [Green Version]
- Billeaud, C.; Puccio, G.; Saliba, E.; Guillois, B.; Vaysse, C.; Pecquet, S.; Steenhout, P. Safety and tolerance evaluation of milk fat globule membrane-enriched infant formulas: A randomized controlled multicenter non-inferiority trial in healthy term infants. Clin. Med. Insights Pediatr. 2014, 8, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Timby, N.; Domellöf, M.; Holgerson, P.L.; West, C.E.; Lönnerdal, B.; Hernell, O.; Johansson, I. Oral Microbiota in Infants Fed a Formula Supplemented with Bovine Milk Fat Globule Membranes—A Randomized Controlled Trial. PLoS ONE 2017, 12, e0169831. [Google Scholar] [CrossRef]
- Grip, T.; Dyrlund, T.S.; Ahonen, L.; Domellöf, M.; Hernell, O.; Hyötyläinen, T.; Knip, M.; Lönnerdal, B.; Orešič, M.; Timby, N. Serum, plasma and erythrocyte membrane lipidomes in infants fed formula supplemented with bovine milk fat globule membranes. Pediatr. Res. 2018, 84, 726–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Parenti, M.; Grip, T.; Lönnerdal, B.; Timby, N.; Domellöf, M.; Hernell, O.; Slupsky, C.M. Fecal microbiome and metabolome of infants fed bovine MFGM supplemented formula or standard formula with breast-fed infants as reference: A randomized controlled trial. Sci. Rep. 2019, 9, 11589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Wu, S.S.; Berseth, C.L.; Harris, C.L.; Richards, J.D.; Wampler, J.L.; Zhuang, W.; Cleghorn, G.; Rudolph, C.D.; Liu, B.; et al. Improved Neurodevelopmental Outcomes Associated with Bovine Milk Fat Globule Membrane and Lactoferrin in Infant Formula: A Randomized, Controlled Trial. J. Pediatr. 2019, 215, 24–31.e8. [Google Scholar] [CrossRef] [PubMed]
- Chichlowski, M.; Bokulich, N.; Harris, C.L.; Wampler, J.L.; Li, F.; Berseth, C.L.; Rudolph, C.; Wu, S.S. Effect of Bovine Milk Fat Globule Membrane and Lactoferrin in Infant Formula on Gut Microbiome and Metabolome at 4 Months of Age. Curr. Dev. Nutr. 2021, 5, nzab027. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Peng, Y.; Li, Z.; Christensen, B.; Heckmann, A.B.; Stenlund, H.; Lönnerdal, B.; Hernell, O. Feeding Infants Formula With Probiotics or Milk Fat Globule Membrane: A Double-Blind, Randomized Controlled Trial. Front. Pediatr. 2019, 7, 347. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Peng, Y.; Li, Z.; Christensen, B.; Heckmann, A.B.; Lagerqvist, C.; Stenlund, H.; Lönnerdal, B.; Hernell, O.; West, C.E. Serum cytokine patterns are modulated in infants fed formula with probiotics or milk fat globule membranes: A randomized controlled trial. PLoS ONE 2021, 16, e0251293. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Jiang, B.; Zhou, L.; Ma, J.; Yang, L.; Wang, F.; Liu, H.; Zhang, N.; Li, X.; Petocz, P.; et al. Neurodevelopmental outcomes of healthy Chinese term infants fed infant formula enriched in bovine milk fat globule membrane for 12 months—A randomized controlled trial. Asia Pac. J. Clin. Nutr. 2021, 30, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, J.; Yeiser, M.; Harris, C.L.; Wampler, J.L.; London, H.E.; Patterson, A.C.; Wu, S.S. Infant Formula with Added Bovine Milk Fat Globule Membrane and Modified Iron Supports Growth and Normal Iron Status at One Year of Age: A Randomized Controlled Trial. Nutrients 2021, 13, 4541. [Google Scholar] [CrossRef]
- Jiang, B.; Xia, Y.; Zhou, L.; Liang, X.; Chen, X.; Chen, M.; Li, X.; Lin, S.; Zhang, N.; Zheng, L.; et al. Safety and tolerance assessment of milk fat globule membrane-enriched infant formulas in healthy term Chinese infants: A randomised multicenter controlled trial. BMC Pediatr. 2022, 22, 465. [Google Scholar] [CrossRef]
- Zhao, J.; Yi, W.; Liu, B.; Dai, Y.; Jiang, T.; Chen, S.; Wang, J.; Feng, B.; Qiao, W.; Liu, Y.; et al. MFGM components promote gut Bifidobacterium growth in infant and in vitro. Eur. J. Nutr. 2022, 61, 277–288. [Google Scholar] [CrossRef]
- Nieto-Ruiz, A.; García-Santos, J.A.; Verdejo-Román, J.; Diéguez, E.; Sepúlveda-Valbuena, N.; Herrmann, F.; Cerdó, T.; De-Castellar, R.; Jiménez, J.; Bermúdez, M.G.; et al. Infant Formula Supplemented With Milk Fat Globule Membrane, Long-Chain Polyunsaturated Fatty Acids, and Synbiotics Is Associated With Neurocognitive Function and Brain Structure of Healthy Children Aged 6 Years: The COGNIS Study. Front. Nutr. 2022, 9, 820224. [Google Scholar] [CrossRef]
- Mudd, A.T.; Alexander, L.S.; Berding, K.; Waworuntu, R.V.; Berg, B.M.; Donovan, S.M.; Dilger, R.N. Dietary Prebiotics, Milk Fat Globule Membrane, and Lactoferrin Affects Structural Neurodevelopment in the Young Piglet. Front. Pediatr. 2016, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Bhinder, G.; Allaire, J.M.; Garcia, C.; Lau, J.T.; Chan, J.M.; Ryz, N.R.; Bosman, E.S.; Graef, F.A.; Crowley, S.M.; Celiberto, L.S.; et al. Milk Fat Globule Membrane Supplementation in Formula Modulates the Neonatal Gut Microbiome and Normalizes Intestinal Development. Sci. Rep. 2017, 7, 45274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Huërou-Luron, I.; Bouzerzour, K.; Ferret-Bernard, S.; Ménard, O.; Le Normand, L.; Perrier, C.; Le Bourgot, C.; Jardin, J.; Bourlieu, C.; Carton, T.; et al. A mixture of milk and vegetable lipids in infant formula changes gut digestion, mucosal immunity and microbiota composition in neonatal piglets. Eur. J. Nutr. 2018, 57, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Moukarzel, S.; Dyer, R.A.; Garcia, C.; Wiedeman, A.M.; Boyce, G.; Weinberg, J.; Keller, B.O.; Elango, R.; Innis, S.M. Milk Fat Globule Membrane Supplementation in Formula-fed Rat Pups Improves Reflex Development and May Alter Brain Lipid Composition. Sci. Rep. 2018, 8, 15277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Gao, J.; Du, M.; Song, J.; Mao, X. Milk Fat Globule Membrane Attenuates High-Fat Diet-Induced Obesity by Inhibiting Adipogenesis and Increasing Uncoupling Protein 1 Expression in White Adipose Tissue of Mice. Nutrients 2018, 10, 331. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wu, Y.; Ye, H.; Feng, C.; Han, D.; Tao, S.; Pi, Y.; Zhao, J.; Chen, L.; Wang, J. Dietary milk fat globule membrane supplementation during late gestation increased the growth of neonatal piglets by improving their plasma parameters, intestinal barriers, and fecal microbiota. RSC Adv. 2020, 10, 16987–16998. [Google Scholar] [CrossRef]
- Li, T.; Gong, H.; Yuan, Q.; Du, M.; Ren, F.; Mao, X. Supplementation of polar lipids-enriched milk fat globule membrane in high-fat diet-fed rats during pregnancy and lactation promotes brown/beige adipocyte development and prevents obesity in male offspring. FASEB J. 2020, 34, 4619–4634. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Q.C.; Gong, H.; Du, M.; Mao, X.Y. Supplementation of milk polar lipids to obese dams improves neurodevelopment and cognitive function in male offspring. FASEB J. 2021, 35, e21454. [Google Scholar] [CrossRef]
- O’Mahony, S.M.; McVey Neufeld, K.A.; Waworuntu, R.V.; Pusceddu, M.M.; Manurung, S.; Murphy, K.; Strain, C.; Laguna, M.C.; Peterson, V.L.; Stanton, C.; et al. The enduring effects of early-life stress on the microbiota-gut-brain axis are buffered by dietary supplementation with milk fat globule membrane and a prebiotic blend. Eur. J. Neurosci. 2020, 51, 1042–1058. [Google Scholar] [CrossRef]
- Gong, H.; Yuan, Q.; Pang, J.; Li, T.; Li, J.; Zhan, B.; Chang, R.; Mao, X. Dietary Milk Fat Globule Membrane Restores Decreased Intestinal Mucosal Barrier Development and Alterations of Intestinal Flora in Infant-Formula-Fed Rat Pups. Mol. Nutr. Food Res. 2020, 64, e2000232. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zhang, Q.; Xin, F.; Cao, B.; Qian, L.; Dong, Y. Neonatal Milk Fat Globule Membrane Supplementation During Breastfeeding Ameliorates the Deleterious Effects of Maternal High-Fat Diet on Metabolism and Modulates Gut Microbiota in Adult Mice Offspring in a Sex-Specific Way. Front. Cell. Infect. Microbiol. 2021, 11, 621957. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Huang, S.; Li, Y.; Niu, Y.; Chen, H.; Wu, J. Milk Fat Globule Membrane Alleviates Short Bowel Syndrome-Associated Liver Injury in Rats Through Inhibiting Autophagy and NLRP3 Inflammasome Activation. Front. Nutr. 2022, 9, 758762. [Google Scholar] [CrossRef] [PubMed]
- Fraser, K.; Ryan, L.; Dilger, R.N.; Dunstan, K.; Armstrong, K.; Peters, J.; Stirrat, H.; Haggerty, N.; MacGibbon, A.K.H.; Dekker, J.; et al. Impacts of Formula Supplemented with Milk Fat Globule Membrane on the Neurolipidome of Brain Regions of Piglets. Metabolites 2022, 12, 689. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Gong, H.; Du, M.; Li, T.; Mao, X. Milk fat globule membrane supplementation to obese rats during pregnancy and lactation promotes neurodevelopment in offspring via modulating gut microbiota. Front. Nutr. 2022, 9, 945052. [Google Scholar] [CrossRef]
- Wu, Z.; Liu, X.; Huang, S.; Li, T.; Zhang, X.; Pang, J.; Zhao, J.; Chen, L.; Zhang, B.; Wang, J.; et al. Milk Fat Globule Membrane Attenuates Acute Colitis and Secondary Liver Injury by Improving the Mucus Barrier and Regulating the Gut Microbiota. Front. Immunol. 2022, 13, 865273. [Google Scholar] [CrossRef]
- Collins, J.M.; Caputi, V.; Manurung, S.; Gross, G.; Fitzgerald, P.; Golubeva, A.V.; Popov, J.; Deady, C.; Dinan, T.G.; Cryan, J.F.; et al. Supplementation with milk fat globule membrane from early life reduces maternal separation-induced visceral pain independent of enteric nervous system or intestinal permeability changes in the rat. Neuropharmacology 2022, 210, 109026. [Google Scholar] [CrossRef]
- Fontecha, J.; Brink, L.; Wu, S.; Pouliot, Y.; Visioli, F.; Jiménez-Flores, R. Sources, Production, and Clinical Treatments of Milk Fat Globule Membrane for Infant Nutrition and Well-Being. Nutrients 2020, 12, 1607. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Type of Study/Year | MFGM Formulation | Health Effects | Reference |
|---|---|---|---|
| RCT/2011 e 2018 | MFGM-fortified complementary food | Lower incidence of bloody diarrhea Abnormalities removal in protein and energy metabolism, toward an enhanced immunity, decrease of T helper type 1 response, decrease of trimethylamine-N-oxide (Peruvian infants) | [53,54] |
| RCT/2012 | Bovine gangliosides enriched formula | Better cognitive function, similar to breastfed controls (Indonesian infants) | [55] |
| RCT/2012 | MFGM-enriched formula | Lower febrile episodes (Belgian preschool children) | [56] |
| RCT/2014 | Bovine ganglioside enriched formula | No difference in diarrhea incidence (Indian infants) | [57] |
| RCT/2014 e 2015 | MFGM-enriched formula | Positive effect on cognitive scores; lower incidence of otitis; higher serum cholesterol (Swedish infants) | [23,58,59] |
| RCT/2014 | Formula enriched with a protein rich MFGM and Formula enriched with a lipid rich MFGM | For both formula no weight gain; some atopic dermatitis in protein rich MFGM group (French and Italian infants) | [60] |
| RCT/2017 | MFGM-enriched formula | Moderate effect on oral microbiome, Moraxella catarrhalis was less prevalent (Swedish infants) | [61] |
| RCT/2018 | MFGM-enriched formula | Differences in the serum/plasma lipidome at 4 and 6 months of age (Swedish infants) | [62] |
| RCT/2019, 2019 | MFGM-enriched formula | Enriched-formula infants are more metabolically similar to BF infants. Limited differences in fecal microbiome and metabolome between Enriched-formula fed and standard formula fed infants (Swedish infants) | [7,63] |
| RCT/2019, 2021 | MFGM plus lactoferrin enriched formula | Accelerated neurodevelopmental profile and fewer diarrhea and respiratory-associated adverse events. Increased prevalence of Bacteroides species n gut microbiome, modest effect on stool metabolome (Chinese infants) | [64,65] |
| RCT/2019, 2021 | MFGM- or probiotic enriched formula | Fewer fever episodes compared to standard formula, the outcomes of MFGM group were close to those of the breastfed group, Cytokine profile of the MFGM group approached that of breastfed infants, (Chinese infants) | [66,67] |
| RCT/2021 | MFGM-enriched formula | Serum gangliosides increase and improvement of cognitive development (Chinese infants) | [68] |
| RCT/2021 | MFGM plus iron enriched formula | Well-tolerated, adequate growth, normal iron status until one year of age (US infants) | [69] |
| RCT/2021 | MFGM-enriched formula | Serum metabolome shifts to fatty acid oxidation and ketogenesis (Chinese infants) | [26] |
| RCT/2022 | MFGM-enriched formula | The MFGM-formula had a similar digestive tolerance to the standard formula and was different from the breastfeeding in stool color (Chinese infants) | [70] |
| RCT/2022 | MFGM-enriched formula | Supported growth of Bifidobacterium in gut microbiome (Chinese infants, limited number) | [71] |
| RCT/2022 | MFGM plus PUFA plus symbiotic-enriched formula | Neurocognitive positive effect. Long term effect at 6 years of age of supplementation in the first months of life. Bifidogenic and lactogenic effect on gut microbiota (Spanish infants) | [72] |
| Animal study/2016 | Supplement of MFGM plus prebiotic plus lactoferrin | Advance in neurodevelopment (piglets) | [73] |
| Animal study/2017 | MFGM-enriched formula | Protection against inflammation, development of intestinal barrier and microbiome (rats) | [74] |
| Animal study/2018 | Mixture of vegetable oils plus milk fat plus MFGM | Developmental profile of intestinal physiology and mucosal immunity were positively modified (piglets) | [75] |
| Animal study/2018 | MFGM-enriched formula | Promotion of reflex development, change in brain phospholipid composition (rats) | [76] |
| Animal study/2018 | MFGM addition to high fat diet | Protection against diet-induced adiposity by suppressing adipogenesis and promoting brown-like transformation of white adipose tissue (mice) | [77] |
| Animal study/2020 | MFGM-supplemented diet in late gestation | Positive effects on intestinal barrier (neonatal piglets), improvement of plasma parameters and gut microbiota (sows) | [78] |
| Animal study/2020, 2021 | MFGM plus polar lipids supplementation in basal and high fat diet | Maternal treatment in high fat diet reduced adiposity and induced thermogenesis in offspring. Neurodevelopment and alleviation of cognitive impairment in the offspring (pregnant rats and their offspring) | [79,80] |
| Animal study/2020 | MFGM plus prebiotic blend | Modification of the gut microbiota, improvements of the long-term effects of early-life stress (maternal separation in rats) | [81] |
| Animal study/2020 | Intragastrically administered formula with MFGM | Positive effects on intestinal physiology and gut microbiota (rats) | [82] |
| Animal study/2021 | MFGM supplementation | Modulation of gut microbiota and protective effect on glucose and lipid metabolism (mice offspring exposed to maternal high fat diet) | [83] |
| Animal study/2021, 2022 | MFGM supplementation | Improvement of the colonic mucus barrier and regulation of NLRP6 inflammasome; Positive effect on liver injury by inhibiting the activity of the autophagy-inflammasome (rats with short bowel syndrome) | [45,84] |
| Animal study/2022 | Oral supplementation of MFGM | Intestinal barrier differentiation, increase expression of tight junctions proteins (rats) | [31] |
| Animal study/2022 | MFGM-enriched formula | Modification of the hippocampus lipidome (piglets) | [85] |
| Animal study/2022 | MFGM supplementation in basal and high fat diet | Neuroinflammation reduction by decrease of lipopolysaccharides and pro-inflammatory cytokines in the circulation and brain, inhibition the activation of microglia, downregulation of pro-inflammatory bacteria in the gut (obese rats) | [86] |
| Animal study/2022 | MFGM supplementation | Reduction of colitis and hepatic injury, positive effect on the mucosal barrier and bacterial community, oxidative stress inhibition (mice) | [87] |
| Animal study/2022 | MFGM-enriched formula | reduction of induced visceral hypersensitivity, better cognitive performance (maternal separation in rats) | [88] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavaletto, M.; Givonetti, A.; Cattaneo, C. The Immunological Role of Milk Fat Globule Membrane. Nutrients 2022, 14, 4574. https://doi.org/10.3390/nu14214574
Cavaletto M, Givonetti A, Cattaneo C. The Immunological Role of Milk Fat Globule Membrane. Nutrients. 2022; 14(21):4574. https://doi.org/10.3390/nu14214574
Chicago/Turabian StyleCavaletto, Maria, Annalisa Givonetti, and Chiara Cattaneo. 2022. "The Immunological Role of Milk Fat Globule Membrane" Nutrients 14, no. 21: 4574. https://doi.org/10.3390/nu14214574
APA StyleCavaletto, M., Givonetti, A., & Cattaneo, C. (2022). The Immunological Role of Milk Fat Globule Membrane. Nutrients, 14(21), 4574. https://doi.org/10.3390/nu14214574