

On the Centennial of Vitamin D—Vitamin D, Inflammation, and Autoimmune Thyroiditis: A Web of Links and Implications


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
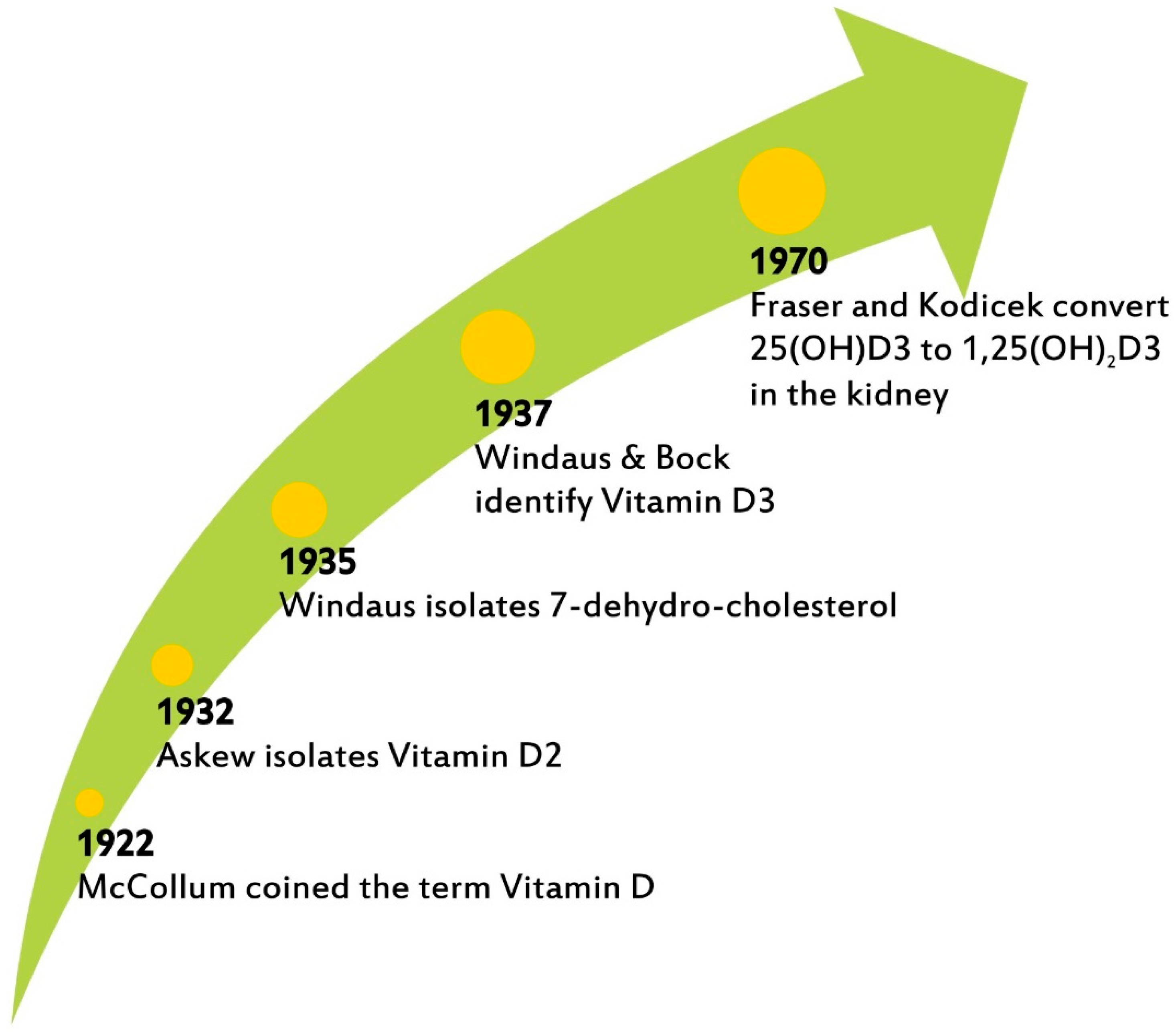
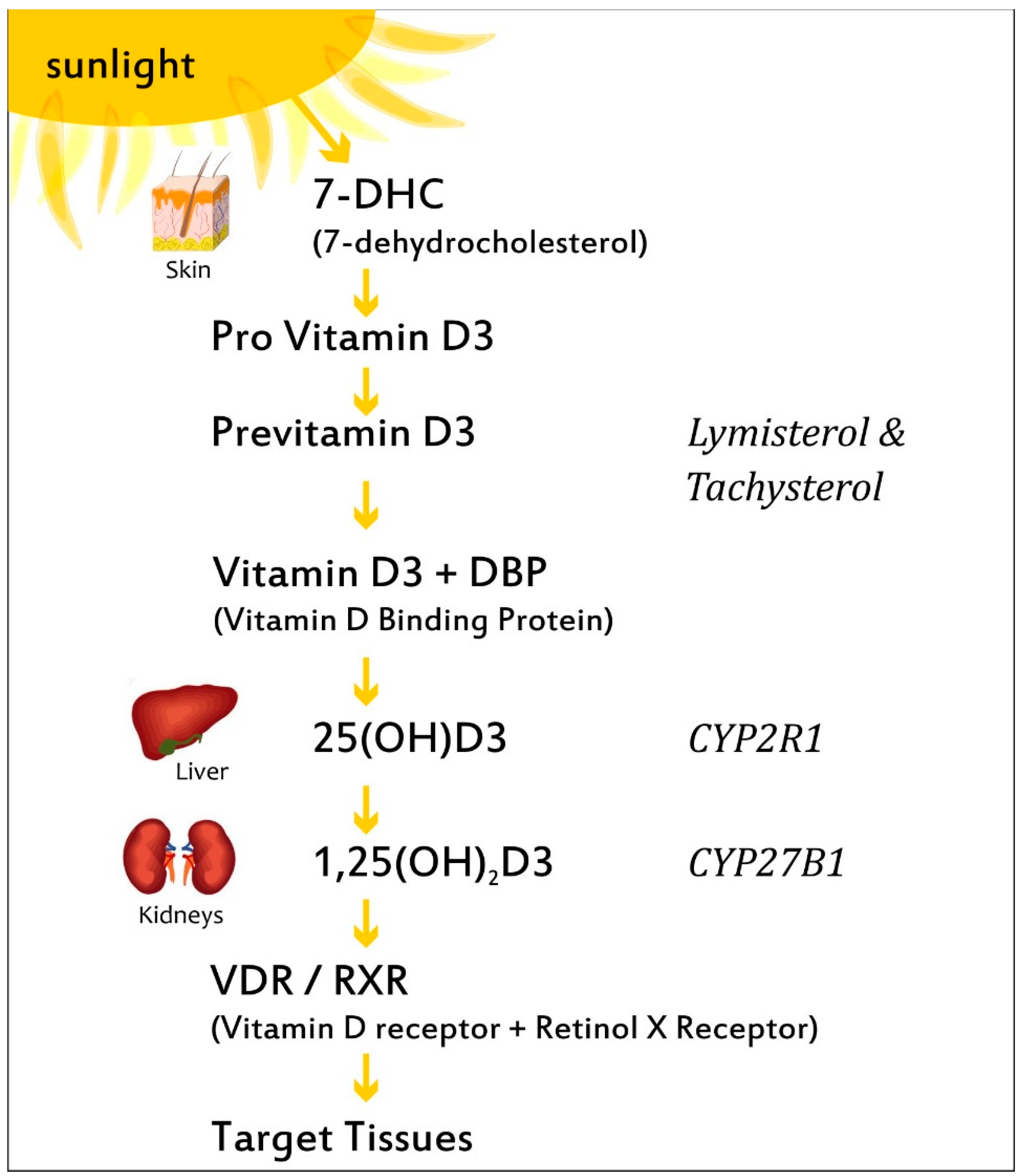
3. VitD Biosynthesis and Metabolism
4. VitD3 and Autoimmune Thyroiditis
4.1. VitD3 and the Epidemiology of Autoimmune Thyroiditis
4.2. Effects of VitD3 Treatment on Hashimoto’s Thyroiditis and Graves’ Disease
4.3. VitD3 and Pregnancy
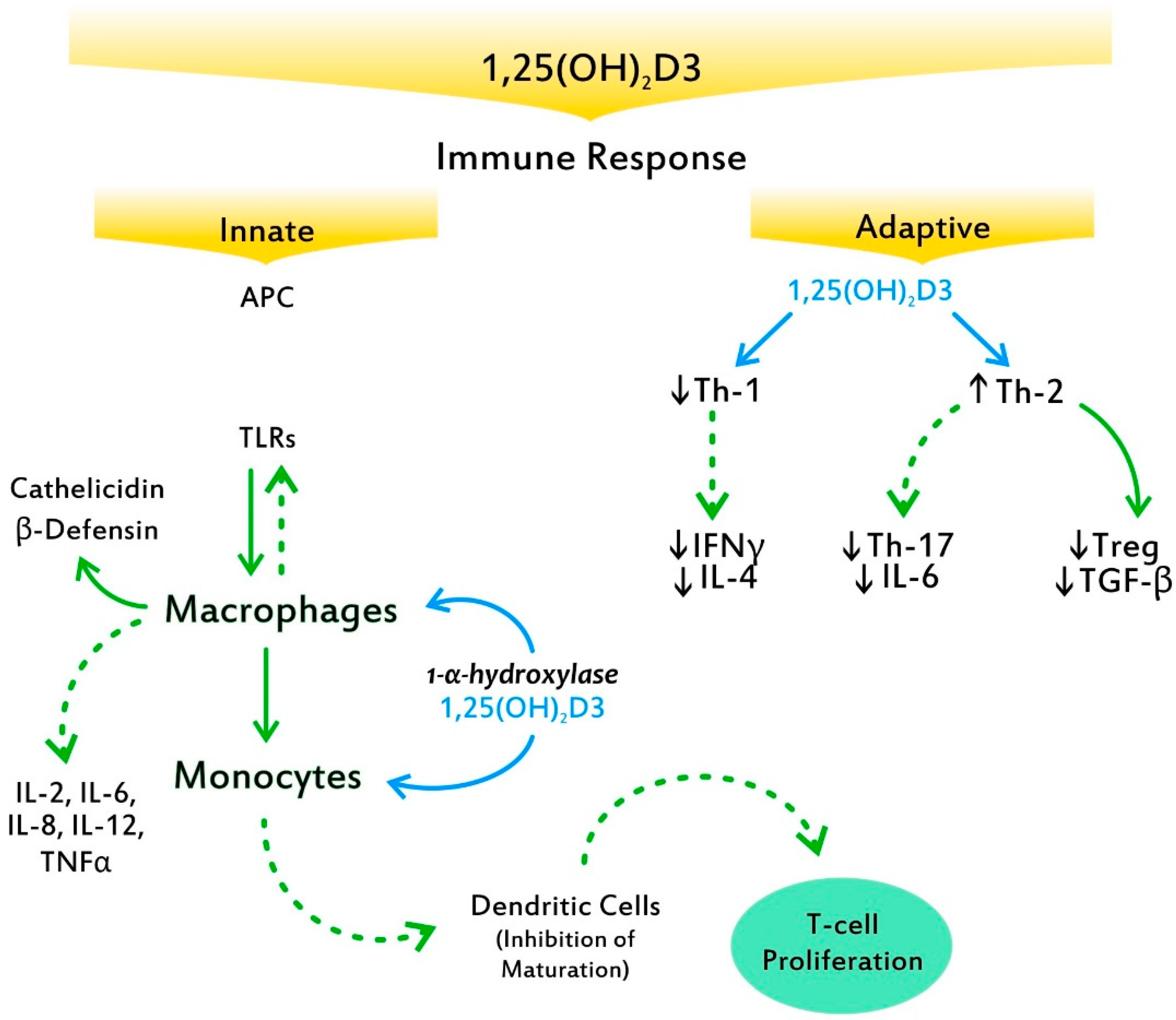
5. VitD3 and Mechanisms of Autoimmunity
6. Discrepancies and Limitations
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AID | autoimmune disease |
| AIT | autoimmune thyroiditis |
| AITD | autoimmune thyroid disease |
| DBD | vitamin D binding protein |
| CYP27B1 | (1-α-hydroxylase) |
| GWAS | genome-wide association studies |
| GD | Graves’ disease |
| GO | Graves’ opthalmopathy |
| HT | Hashimoto’s thyroiditis |
| IL-6 | interleukin 6 |
| SNP | single-nucleotide polymorphism |
| TgAB | antithyroglobulin antibodies |
| TNFα | tumor necrotic factor α |
| TPOAb | antithyroid peroxidase antibodies |
| TRAb | TSH receptor antibodies |
| Tregs | T regulatory cells |
| TSH | thyroid-stimulating hormone |
| VDR | vitamin D receptor |
References
- McCollum, E.V.; Simmonds, N.; Becker, J.E.; Shipley, P.G. An experimental demonstration of the existence of a vitamin which promotes calcium deposition. J. Biol. Chem. 1922, 53, 293–312. [Google Scholar] [CrossRef]
- Askew, F.A.; Bourdillon, R.B.; Bruce, R.G.C.; Webster, T. A The distillation of vitamin D. Proc. R. Soc. B 1931, 107, 76–90. [Google Scholar]
- Windaus, A.; Bock, F. Uber das Provitamin aus dem Sterin der Schweineschwarte. Z. Physiol. Chem. 1937, 245, 168–170. [Google Scholar] [CrossRef]
- Fraser, D.R.; Kodicek, E. Unique biosynthesis by kidney of a biologically active vitamin D metabolite. Nature 1970, 228, 764–766. [Google Scholar] [CrossRef] [PubMed]
- Bikle, D.D. Vitamin D metabolism, mechanism of action, and clinical applications. Chem. Biol. 2014, 21, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Cheung, M.M.; Dall, R.D.; Shewokis, P.A.; Altasan, A.; Volpe, S.L.; Amori, R.; Singh, H.; Sukumar, D. The effect of combined magnesium and vitamin D supplementation on vitamin D status, systemic inflammation, and blood pressure: A randomized double-blinded controlled trial. Nutrition 2022, 99–100, 111674. [Google Scholar] [CrossRef]
- Janjusevic, M.; Gagno, G.; Fluca, A.L.; Padoan, L.; Beltrami, P.A.; Sinagra, G.; Moretti, G.; Aleksova, A. The peculiar role of vitamin D in the pathophysiology of cardiovascular and neurodegenerative diseases. Life Sci. 2022, 289, 120193. [Google Scholar] [CrossRef]
- Sassi, F.; Tamone, C.; D’Amelio, P. Vitamin D: Nutrient, Hormone, and Immunomodulator. Nutrients 2018, 10, 1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, E.; Jørgensen, T.N. Immunological effects of vitamin D and their relations to autoimmunity. J. Autoimmun. 2019, 100, 7–16. [Google Scholar] [CrossRef]
- Cashman, K.D. Global differences in vitamin D status and dietary intake: A review of the data. Endocr. Connect. 2022, 11, e210282. [Google Scholar] [CrossRef]
- Munns, C.F.; Shaw, N.; Kiely, M.; Specker, B.L.; Thacher, T.D.; Ozono, K.; Michigami, T.; Tiosano, D.; Mughal, M.Z.; Mäkitie, O.; et al. Global consensus recommendations on prevention and management of nutritional rickets. J. Clin. Endocrinol. Metab. 2016, 101, 394–415. [Google Scholar] [CrossRef]
- Kim, Y.A.; Yoon, J.W.; Lee, Y.; Choi, H.J.; Yun, J.W.; Bae, E.; Kwon, S.H.; Ahn, S.E.; Do, A.R.; Jin, H.; et al. Unveiling genetic variants underlying vitamin D deficiency in multiple Korean Cohorts by an association study. Endocrinol. Metab. 2021, 36, 1189–1200. [Google Scholar] [CrossRef] [PubMed]
- Milan, Κ.L.; Jayasuriya, R.; Harithpriya, K.; Anuradha, M.; Sarada, D.V.L.; Rahayu, N.S.; Ramkumar, N.S. Vitamin D resistant genes—promising therapeutic targets of chronic diseases. Food Funct. 2022, 13, 7984–7998. [Google Scholar] [CrossRef]
- Ma, J.; Wu, D.; Li, C.; Fan, C.; Chao, J.; Liu, Y.; Li, R.; Wang, R.; Miao, W.; Guan, H.; et al. Lower serum 25-hydroxyvitamin D level is associated with 3 types of autoimmune thyroid diseases. Medicine 2015, 94, e1639. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Chen, X.; Qian, X.; Shao, S. Effects of vitamin D treatment on thyroid function and autoimmunity markers in patients with Hashimoto’s thyroiditis-A meta-analysis of randomized controlled trials. J. Clin. Pharm. Ther. 2022, 47, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Vieira, I.H.; Rodrigues, D.; Paiva, I. Vitamin D and autoimmune thyroid disease-cause, consequence, or a vicious cycle? Nutrients 2020, 12, 2791. [Google Scholar] [CrossRef]
- Windhaus, A.; Lettre, H.; Schenck, F. 7-dehydrocholesterol. Ann. Chem. 1945, 520, 98–107. [Google Scholar]
- Holick, M.F.; Smith, E.; Pincus, S. Skin as the site of vitamin D synthesis and target tissue for 1,25-dihydroxyvitamin D3. Use of calcitriol (1,25-dihydroxyvitamin D3) for treatment of psoriasis. Arch. Dermatol. 1987, 123, 1677–1683a. [Google Scholar] [CrossRef]
- Chen, T.C.; Persons, K.S.; Lu, Z.; Mathieu, J.S.; Holick, M.F. An evaluation of the biologic activity and vitamin D receptor binding affinity of the photoisomers of vitamin D3 and previtamin D3. J. Nutr. Biochem. 2000, 11, 267–272. [Google Scholar] [CrossRef]
- Baeke, F.; Takiishi, T.; Korf, H.; Gysemans, C.; Mathieu, C. Vitamin D: Modulator of the immune system. Curr. Opin. Pharmacol. 2010, 10, 482–496. [Google Scholar] [CrossRef]
- Kivity, S.; Agmon-Levin, N.; Zisappl, M.; Shapira, Y.; Nagy, E.V.; Dankó, K.; Szekanecz, Z.; Langevitz, P.; Shoenfeld, Y. Vitamin D, and autoimmune thyroid diseases. Cell. Mol. Immunol. 2011, 8, 243–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Effraimidis, G.; Badenhoop, K.; Tijssen, J.G.; Wiersinga, W.M. Vitamin D deficiency is not associated with early stages of thyroid autoimmunity. Eur. J. Endocrinol. 2012, 167, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Turashvili, N.; Javashvili, L.; Giorgadze, E. Vitamin D deficiency is more common in women with autoimmune thyroiditis: A retrospective Study. Int. J. Endocrinol. 2021, 2021, 4465563. [Google Scholar] [CrossRef]
- Choi, Y.M.; Kim, W.G.; Kim, T.Y.; Bae, S.J.; Kim, H.K.; Jang, E.K.; Jeon, M.J.; Han, J.M.; Lee, S.H.; Baek, J.H.; et al. Low levels of serum vitamin D3 are associated with autoimmune thyroid disease in pre-menopausal women. Thyroid 2014, 24, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Chai, Y.; Wei, H.; Wang, K.; Tan, L.; Zhang, W.; Fan, Y.; Li, F.; Shan, Z.; Zhu, M. Vitamin D deficiency is associated with thyroid autoimmunity: Results from an epidemiological survey in Tianjin, China. Endocrine 2021, 73, 447–454. [Google Scholar] [CrossRef]
- Chao, G.; Zhu, Y.; Fang, L. Correlation between Hashimoto’s Thyroiditis-related thyroid hormone levels and 25-hydroxyvitamin D. Front. Endocrinol. 2020, 14, 32117049. [Google Scholar] [CrossRef] [Green Version]
- De Pergola, G.; Triggiani, V.; Bartolomeo, N.; Giagulli, V.A.; Anelli, M.; Masiello, M.; Candita, V.; De Bellis, D.; Silvestris, F. Low 25 hydroxyvitamin D levels are independently associated with autoimmune thyroiditis in a cohort of apparently healthy overweight and obese subjects. Endocr. Metab. Immune Disords-Drug Targets. 2018, 18, 646–652. [Google Scholar] [CrossRef]
- Bozkurt, N.C.; Karbek, B.; Ucan, B.; Sahin, M.; Cakal, E.; Ozbek, M.; Delibasi, T. The association between severity of vitamin D deficiency and Hashimoto’s thyroiditis. Endocr. Pract. 2013, 19, 479–484. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Han, B.; Zhu, C.; Li, Q.; Chen, C.; Zhai, H.; Wang, N.; Chen, Y.; Lu, Y. Bidirectional mendelian randomization analysis for vitamin D and thyroid peroxidase antibody. Int. J. Endocrinol. 2022, 2022, 2260388. [Google Scholar] [CrossRef]
- Cvek, M.; Kaličanin, D.; Barić, A.; Vuletić, M.; Gunjača, I.; Torlak Lovrić, V.; Škrabić, V.; Punda, A.; Boraska Perica, V. Vitamin D and Hashimoto’s thyroiditis: Observations from CROHT biobank. Nutrients 2021, 13, 2793. [Google Scholar] [CrossRef]
- Misharin, A.; Hewison, M.; Chen, C.R.; Lagishetty, V.; Aliesky, H.A.; Mizutori, Y.; Rapoport, B.; McLachlan, S.M. Vitamin D deficiency modulates Graves’ hyperthyroidism induced in BALB/c mice by thyrotropin receptor immunization. Endocrinology 2009, 150, 1051–1060. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, T.; Okamoto, Y.; Hamada, N. Serum vitamin D levels are decreased in patients without remission of Graves’ disease. Endocrine 2013, 43, 230–232. [Google Scholar] [CrossRef]
- Planck, T.; Shahida, B.; Malm, J.; Manjer, J. Vitamin D in Graves’ disease: Levels, correlation with laboratory and clinical parameters, and genetics. Eur. Thyroid J. 2018, 7, 27–33. [Google Scholar] [CrossRef]
- Rotondi, M.; Chiovato, L. Vitamin D deficiency in patients with Graves’ disease: Probably something more than a casual association. Endocrine 2013, 43, 3–5. [Google Scholar] [CrossRef] [Green Version]
- Mazokopakis, E.E.; Papadomanolaki, M.G.; Tsekouras, K.C.; Evangelopoulos, A.D.; Kotsiris, D.A.; Tzortzinis, A.A. Is vitamin D related to pathogenesis and treatment of Hashimoto’s thyroiditis? Hell. J. Nucl. Med. 2015, 18, 222–227. [Google Scholar]
- Vahabi Anaraki, P.; Aminorroaya, A.; Amini, M.; Momeni, F.; Feizi, A.; Iraj, B.; Tabatabaei, A. Effect of Vitamin D deficiency treatment on thyroid function and autoimmunity markers in Hashimoto’s thyroiditis: A double-blind randomized placebo-controlled clinical trial. J. Res. Med. Sci. 2017, 26, 29026419. [Google Scholar] [CrossRef]
- Chahardoli, R.; Saboor-Yaraghi, A.A.; Amouzegar, A.; Khalili, D.; Vakili, A.Z.; Azizi, F. Can supplementation with vitamin D modify thyroid autoantibodies (Anti-TPO Ab, Anti-Tg Ab) and thyroid profile (T3, T4, TSH) in Hashimoto’s thyroiditis? A double-blind, randomized clinical trial. Horm. Metab. Res. 2019, 51, 296–301. [Google Scholar] [CrossRef]
- Grove-Laugesen, D.; Cramon, P.K.; Malmstroem, S.; Ebbehoj, E.; Watt, T.; Hansen, K.W.; Rejnmark, L. Effects of supplemental vitamin D on muscle performance and quality of life in Graves’ Disease: A randomized clinical trial. Thyroid 2020, 30, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Khozam, S.A.; Sumaili, A.M.; Alflan, M.A.; Shawabkeh, R.A.S. Association between vitamin D deficiency and autoimmune thyroid disorder: A systematic review. Cureus 2022, 14, e25869. [Google Scholar] [CrossRef] [PubMed]
- Botelho, I.M.B.; Moura Neto, A.; Silva, C.A.; Tambascia, M.A.; Alegre, S.M.; Zantut-Wittmann, D.E. Vitamin D in Hashimoto’s thyroiditis and its relationship with thyroid function and inflammatory status. Endocr. J. 2018, 65, 1029–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilezikian, J.P.; Formenti, A.M.; Adler, R.A.; Binkley, N.; Bouillon, R.; Lazaretti-Castro, M.; Marcocci, C.; Napoli, N.; Rizzoli, R.; Giustina, A. Vitamin D: Dosing, levels, form, and route of administration: Does one approach fit all? Rev. Endocr. Metab. Disord. 2021, 22, 1201–1218. [Google Scholar] [CrossRef]
- Duntas, L.H. Back to the drawing board? Effects of high-dose vitamin D supplementation in Graves’ disease on muscle strength, lean mass gain, and quality of life. Thyroid 2020, 30, 645–647. [Google Scholar] [CrossRef]
- Ota, K.; Dambaeva, S.; Han, A.R.; Beaman, K.; Gilman-Sachs, A.; Kwak-Kim, J. Vitamin D deficiency may be a risk factor for recurrent pregnancy losses by increasing cellular immunity and autoimmunity. Hum. Reprod. 2014, 29, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Wang, L.; Bu, L.; Song, Y.; Huang, X.; Zhao, J. Immunopharmacological properties of VitD3: 1, 25VitD3 modulates regulatory T Cells and Th17 Cells and the cytokine balance in PBMCs from Women with unexplained recurrent spontaneous abortion (URSA). Curr. Mol. Pharmacol. 2022, 15, 779–793. [Google Scholar] [CrossRef]
- Zhao, Y.; Miao, W.; Li, C.; Yu, X.; Shan, Z.; Guan, H.; Teng, W. Dynamic changes in serum 25-hydroxyvitamin D during pregnancy. PLoS ONE 2014, 9, e90161. [Google Scholar] [CrossRef]
- Christakos, S.; Ajibade, D.V.; Dhawan, P.; Fechner, A.J.; Mady, L.J. Vitamin D metabolism. Endocrinol. Metab. Clin. N. Am. 2010, 39, 243–253. [Google Scholar] [CrossRef]
- Mele, C.; Caputo, M.; Bisceglia, A.; Samà, M.T.; Zavattaro, M.; Aimaretti, G.; Pagano, L.; Prodam, F.; Marzullo, P. Immunomodulatory effects of vitamin D in thyroid diseases. Nutrients 2020, 12, 1444. [Google Scholar] [CrossRef] [PubMed]
- Kreutz, M.; Andressen, R. Induction of human monocyte to macrophage maturation in vitro by 1,25-dihydroxyvitamin D3. Blood 1990, 76, 2457–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokuda, N.; Levy, R.B. 1,25-dihydroxyvitamin D3 stimulates phagocytosis but suppresses HLA-DR and CD13 antigen expression in human mononuclear phagocytes. Proc. Soc. Exp. Biol. Med. 1996, 211, 244–250. [Google Scholar] [CrossRef]
- Bikle, D.D. Vitamin D regulation of immune function. Curr. Osteoporos. Rep. 2022, 20, 186–193. [Google Scholar] [CrossRef]
- Yuk, J.M.; Shin, D.M.; Lee, H.M.; Yang, C.S.; Jin, H.S.; Kim, K.K.; Lee, Z.W.; Lee, S.H.; Kim, J.M.; Jo, E.K. Vitamin D3 induces autophagy in human monocytes/macrophages via cathelicidin. Cell Host Microbe 2009, 6, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Gombart, A.F.; Borregaard, N.; Koeffler, H.P. Human cathelicidin antimicrobial peptide (CAMP) gene is a direct target of the vitamin D receptor and is strongly upregulated in myeloid cells by 1,25-dihydroxyvitamin D3. FASEB J. 2005, 19, 1067–1077. [Google Scholar] [CrossRef] [Green Version]
- Bilezikian, J.P.; Bikle, D.; Hewison, M.; Lazaretti-Castro, M.; Formenti, A.M.; Gupta, A.; Madhavan, M.V.; Nair, N.; Babalyan, V.; Hutchings, N.; et al. Vitamin D and COVID-19. Eur. J. Endocrinol. 2020, 183, R133–R147. [Google Scholar] [CrossRef]
- Boonstra, A.; Barrat, F.J.; Crain, C.; Heath, V.L.; Savelkoul, H.F.; O’Garra, A. 1alpha,25-Dihydroxyvitamin d3 has a direct effect on naive CD4(+) T cells to enhance the development of Th2 cells. J. Immunol. 2001, 167, 4974–4980. [Google Scholar] [CrossRef] [Green Version]
- Albergamo, A.; Apprato, G.; Silvagno, F. The role of Vitamin D in supporting health in the COVID-19 era. Int. J. Mol. Sci. 2022, 23, 3621. [Google Scholar] [CrossRef]
- Honardoost, M.; Ghavideldarestani, M.; Khamseh, M.E. Role of vitamin D in pathogenesis and severity of COVID-19 infection. Arch. Physiol. Biochem. 2020, 30, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Rathi, H.; Haq, A.; Wimalawansa, S.J.; Sharma, A. Putative roles of vitamin D in modulating immune response and immunopathology associated with COVID-19. Virus Res. 2021, 292, 198235. [Google Scholar] [CrossRef] [PubMed]
- Aranow, C. Vitamin D and the immune system. J. Investig. Med. 2011, 59, 881–886. [Google Scholar] [CrossRef] [Green Version]
- Saul, L.; Mair, I.; Ivens, A.; Brown, P.; Samuel, K.; Campbell, J.D.M.; Soong, D.Y.; Kamenjarin, N.; Mellanby, R.J. 1,25-dihydroxyvitamin D3 restrains CD4 + T cell priming ability of CD11c + dendritic cells by upregulating expression of CD31. Front. Immunol. 2019, 10, 600. [Google Scholar] [CrossRef] [Green Version]
- Roehlen, N.; Doering, C.; Hansmann, M.L.; Gruenwald, F.; Vorlaender, C.; Bechstein, W.O.; Holzer, K.; Badenhoop, K.; Penna-Martinez, M. Vitamin D, FOXO3a, and Sirtuin1 in Hashimoto’s Thyroiditis and Differentiated Thyroid Cancer. Front. Endocrinol. 2018, 9, 527. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; He, S.T.; Jiang, W.J.; Xiao, L.; Li, D.F.; Xu, J.; Shi, X.H.; Zhang, J.A. Genetic susceptibility to autoimmune thyroid diseases in a Chinese Han population: Role of vitamin D receptor gene polymorphisms. Ann. D’endocrinologie 2015, 76, 684–689. [Google Scholar] [CrossRef]
- Wang, T.J.; Zhang, F.; Richards, J.B.; Kestenbaum, B.; van Meurs, J.B.; Berry, D.; Kiel, D.P.; Streeten, E.A.; Ohlsson, C.; Koller, D.L.; et al. Common genetic determinants of vitamin D insufficiency: A genome-wide association study. Lancet 2010, 376, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Bizzaro, G.; Shoenfeld, Y. Vitamin D and autoimmune thyroid diseases: Facts and unresolved questions. Immunol. Res. 2015, 61, 46–52. [Google Scholar] [CrossRef]
- Ding, Y.; Liao, W.; He, X.J.; Xiang, W. Effects of 1,25(OH)2D3 and vitamin D receptor on peripheral CD4+/CD8+ double-positive T lymphocytes in a mouse model of systemic lupus erythematosus. J. Cell. Mol. Med. 2017, 21, 975–985. [Google Scholar] [CrossRef]
- Skaaby, T.; Husemoen, L.L.N.; Thuesen, B.H.; Linneberg, A. Prospective population-based study of the association between vitamin D status and incidence of autoimmune disease. Endocrine 2015, 50, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Scragg, R. Limitations of vitamin D supplementation trials: Why observational studies will continue to help determine the role of vitamin D in health. J. Steroid Biochem. Mol. Biol. 2018, 177, 6–9. [Google Scholar] [CrossRef]
- Hahn, J.; Cook, N.R.; Alexander, E.K.; Friedman, S.; Walter, J.; Bubes, V.; Kotler, G.; Lee, I.M.; Manson, J.E.; Costenbader, K.H. Vitamin D and marine omega 3 fatty acid supplementation and incident autoimmune disease: VITAL randomized controlled trial. BMJ 2022, 376, e066452. [Google Scholar] [CrossRef] [PubMed]
- Baeke, F.; Etten, E.V.; Overbergh, L.; Mathieu, C. Vitamin D3 and the immune system: Maintaining the balance in health and disease. Nutr. Res. Rev. 2007, 20, 106–118. [Google Scholar] [CrossRef] [Green Version]
- Manson, J.E.; Cook, N.R.; Lee, I.M.; Christen, W.; Bassuk, S.S.; Mora, S.; Gibson, H.; Gordon, D.; Copeland, T.; D’Agostino, D.; et al. Vitamin D Supplements and Prevention of Cancer and Cardiovascular Disease. N. Engl. J. Med. 2019, 380, 33–44. [Google Scholar] [CrossRef]
- Giustina, A.; Adler, R.A.; Binkley, N.; Bollerslev, J.; Bouillon, R.; Dawson-Hughes, B.; Ebeling, P.R.; Feldman, D.; Formenti, A.M.; Lazaretti-Castro, M.; et al. Consensus statement from 2nd International Conference on Controversies in Vitamin, D. Rev. Endocr. Metab. Disord. 2020, 21, 89–116. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duntas, L.H.; Alexandraki, K.I. On the Centennial of Vitamin D—Vitamin D, Inflammation, and Autoimmune Thyroiditis: A Web of Links and Implications. Nutrients 2022, 14, 5032. https://doi.org/10.3390/nu14235032
Duntas LH, Alexandraki KI. On the Centennial of Vitamin D—Vitamin D, Inflammation, and Autoimmune Thyroiditis: A Web of Links and Implications. Nutrients. 2022; 14(23):5032. https://doi.org/10.3390/nu14235032
Chicago/Turabian StyleDuntas, Leonidas H., and Krystallenia I. Alexandraki. 2022. "On the Centennial of Vitamin D—Vitamin D, Inflammation, and Autoimmune Thyroiditis: A Web of Links and Implications" Nutrients 14, no. 23: 5032. https://doi.org/10.3390/nu14235032
APA StyleDuntas, L. H., & Alexandraki, K. I. (2022). On the Centennial of Vitamin D—Vitamin D, Inflammation, and Autoimmune Thyroiditis: A Web of Links and Implications. Nutrients, 14(23), 5032. https://doi.org/10.3390/nu14235032