
Maternal Supplementation with a Cocoa Extract during Lactation Deeply Modulates Dams’ Metabolism, Increases Adiponectin Circulating Levels and Improves the Inflammatory Profile in Obese Rat Offspring
 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cocoa Extract
2.2. Animals and Experimental Design
2.3. Adiposity Index
2.4. Body Composition Analyses
2.5. Plasma Analyses
2.6. Oral Glucose Tolerance Test (OGTT)
2.7. HOMA-IR and R-QUICKI Analyses
2.8. Indirect Calorimetry and Activity Measurements
2.9. RNA Isolation, cDNA Synthesis and Analysis of Gene Expression
2.10. Western Blot Analysis
2.11. Levels of Total Adiponectin in White Adipose Depots
2.12. Total Lipid Content in Mammary Glands
2.13. 1H NMR Analysis in Mammary Gland for Metabolite Determination
2.14. Statistical Analysis
3. Results
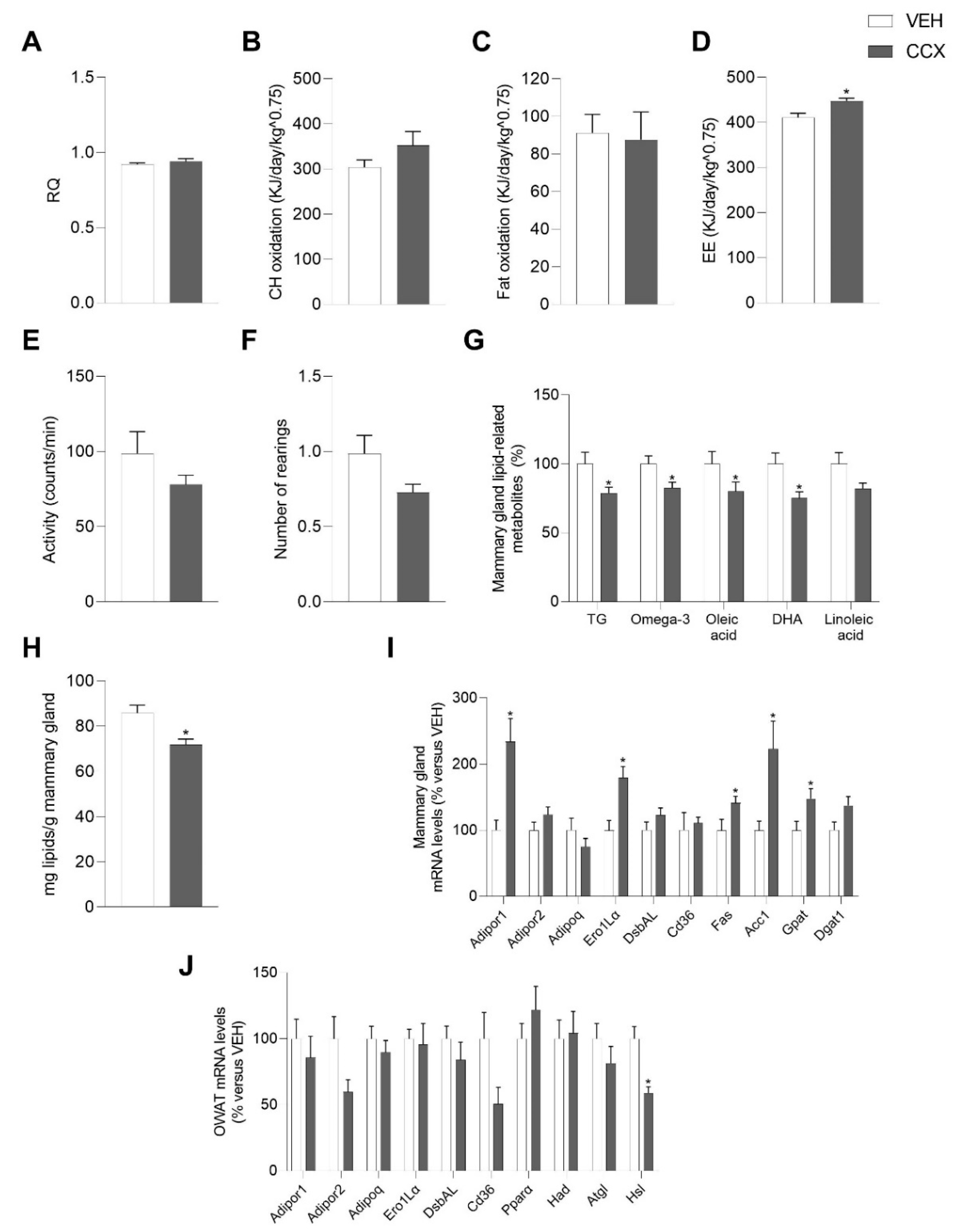
3.1. CCX Supplementation Raised the Plasma Levels of FFAs and Total Adiponectin in Lactating Rats
3.2. CCX-Treated Dams Displayed Increased Energy Expenditure
3.3. CCX Consumption during Lactation Modulated Dams’ Adiponectin Signaling and Lipid Metabolism
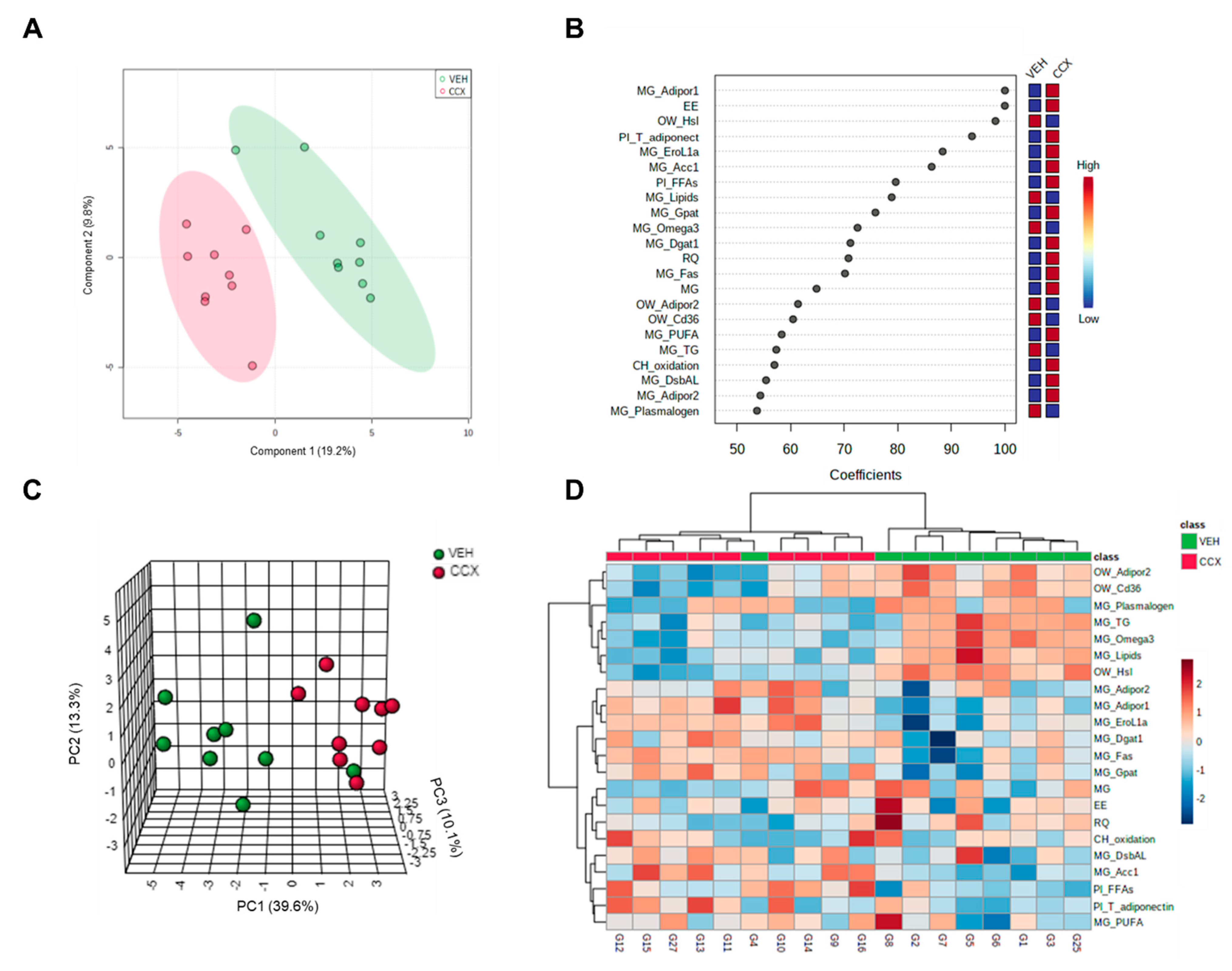
3.4. Multivariate Analysis Clearly Clustered Dams according to the Treatment Received
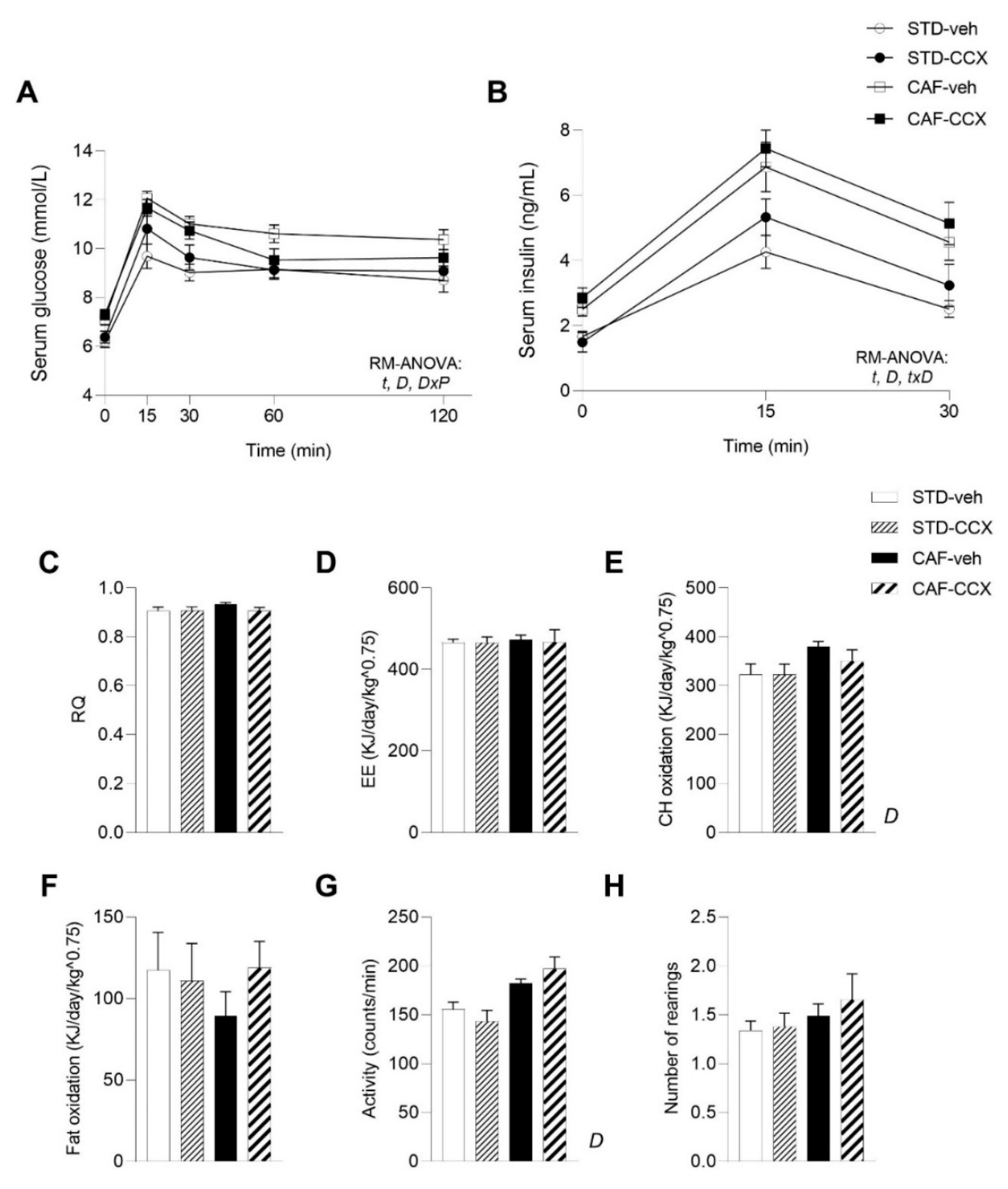
3.5. CAF Diet Intake Induced a MetS-Like Phenotype in Offspring Rats
3.6. The Offspring of Lactating Dams Supplemented with CCX Displayed Decreased Liver Weight and Lean/Fat Ratio
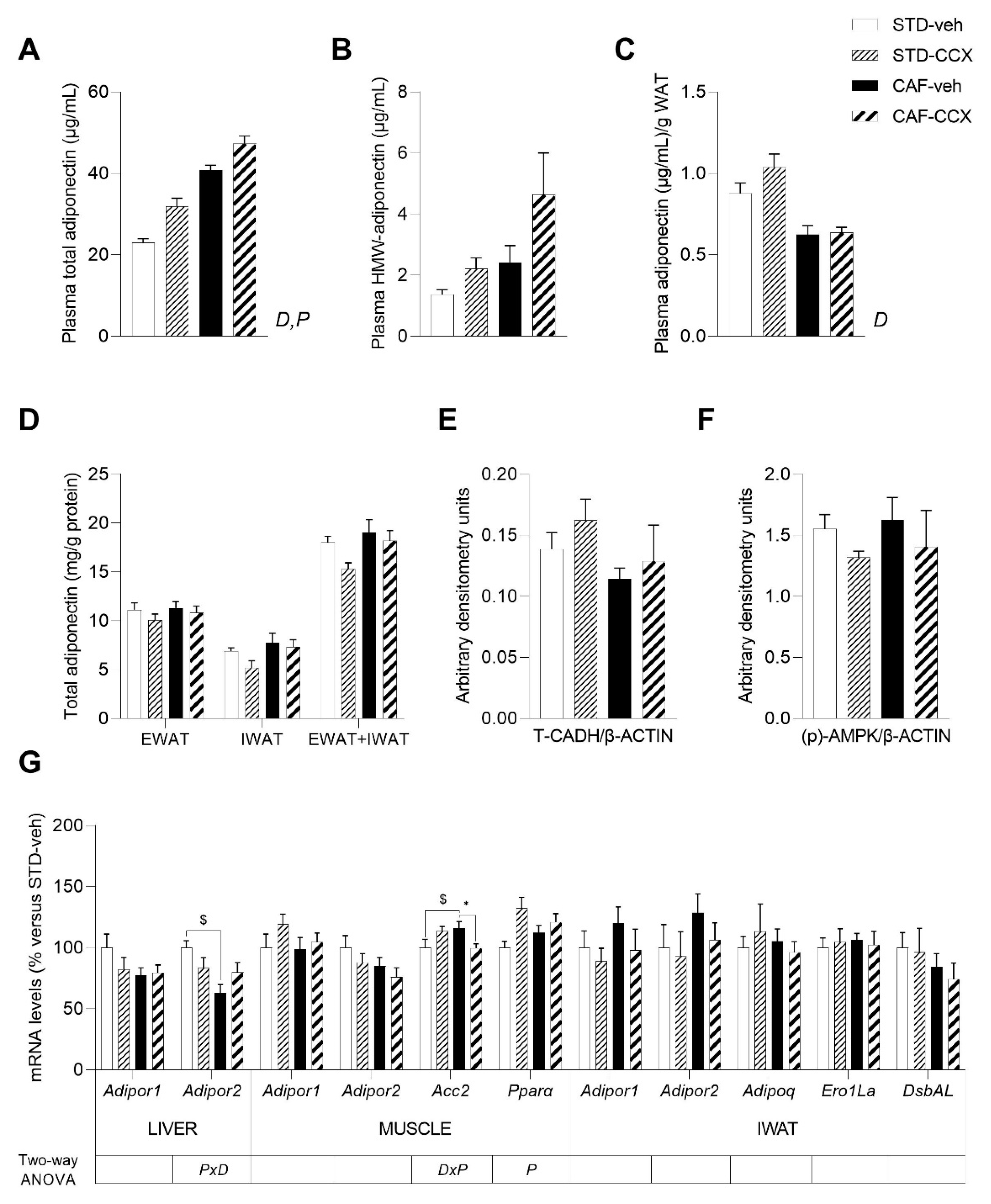
3.7. CCX Intake during Lactation Induced a Clear Metabolic Programming Effect in the Adiponectin Plasma Levels of the Offspring
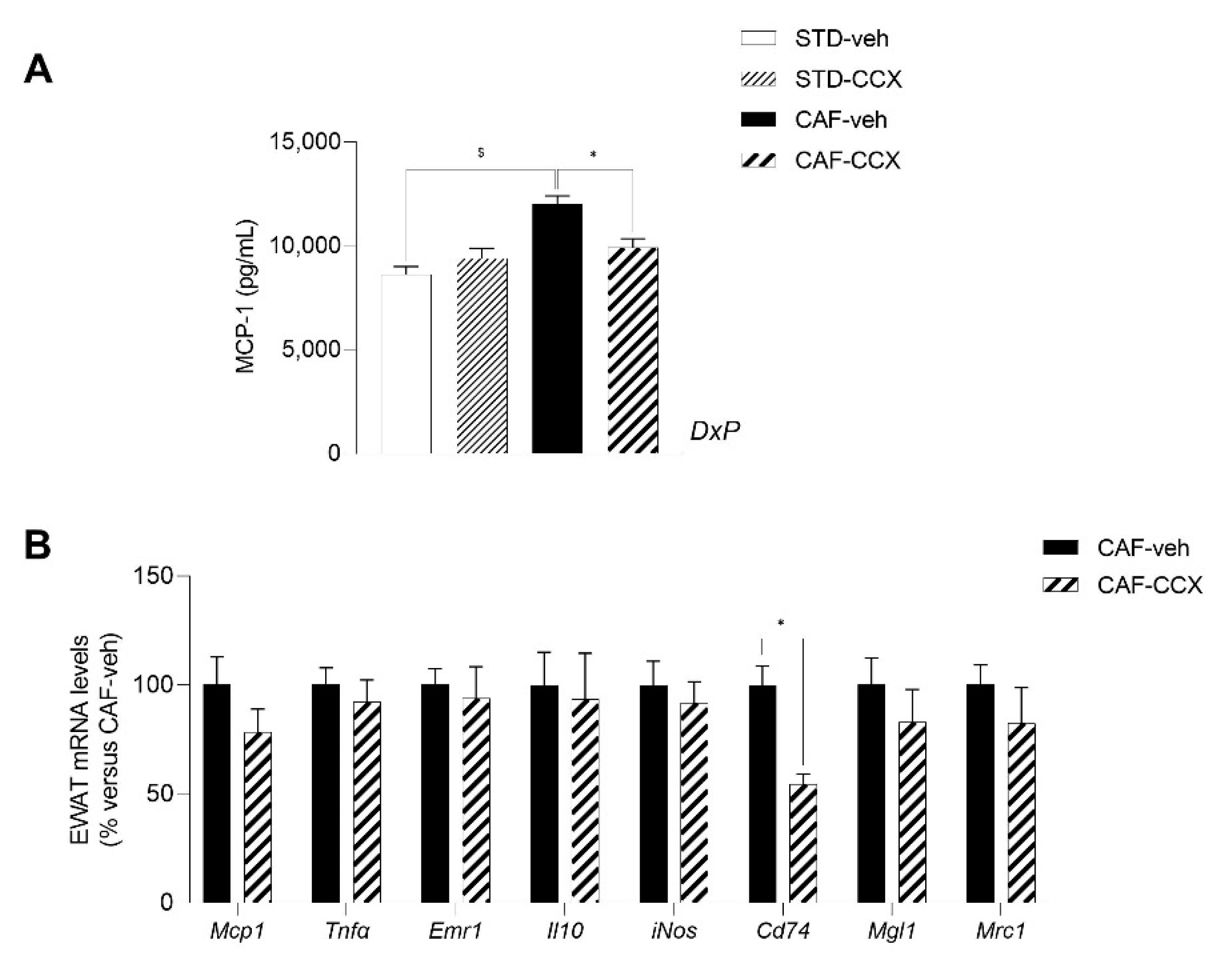
3.8. Maternal CCX Supplementation Restored the Plasma Levels of the Inflammatory Marker MCP-1 in CAF-fed Offspring
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mu, M.; Xu, L.F.; Hu, D.; Wu, J.; Bai, M.J. Dietary Patterns and Overweight/Obesity: A Review Article. Iran. J. Public Health 2017, 46, 869. [Google Scholar] [PubMed]
- Kopp, W. Development of Obesity: The Driver and the Passenger. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 4631. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 18 October 2022).
- Janssen, F.; Bardoutsos, A.; Vidra, N. Obesity Prevalence in the Long-Term Future in 18 European Countries and in the USA. Obes. Facts 2020, 13, 514–527. [Google Scholar] [CrossRef] [PubMed]
- Verduci, E.; Di Profio, E.; Fiore, G.; Zuccotti, G. Integrated Approaches to Combatting Childhood Obesity. Ann. Nutr. Metab. 2022, 78 (Suppl. 2), 8–19. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, K.M.; Reynolds, R.M.; Prescott, S.L.; Nyirenda, M.; Jaddoe, V.W.V.; Eriksson, J.G.; Broekman, B.F.P. Influence of Maternal Obesity on the Long-Term Health of Offspring. Lancet Diabetes Endocrinol. 2017, 5, 53–64. [Google Scholar] [CrossRef] [Green Version]
- North, S.; Crofts, C.; Thoma, C.; Zinn, C. The Role of Maternal Diet on Offspring Hyperinsulinaemia and Adiposity after Birth: A Systematic Review of Randomised Controlled Trials. J. Dev. Orig. Health Dis. 2022, 13, 527–540. [Google Scholar] [CrossRef]
- Schoonejans, J.M.; Ozanne, S.E. Developmental Programming by Maternal Obesity: Lessons from Animal Models. Diabet. Med. 2021, 38, e14694. [Google Scholar] [CrossRef]
- Picó, C.; Palou, M.; Priego, T.; Sánchez, J.; Palou, A. Metabolic Programming of Obesity by Energy Restriction during the Perinatal Period: Different Outcomes Depending on Gender and Period, Type and Severity of Restriction. Front. Physiol. 2012, 3, 436. [Google Scholar] [CrossRef] [Green Version]
- Agosti, M.; Tandoi, F.; Morlacchi, L.; Bossi, A. Nutritional and Metabolic Programming during the First Thousand Days of Life. Pediatr. Med. Chir. 2017, 39, 157. [Google Scholar] [CrossRef] [Green Version]
- Jean-Marie, E.; Bereau, D.; Robinson, J.C. Benefits of Polyphenols and Methylxanthines from Cocoa Beans on Dietary Metabolic Disorders. Foods 2021, 10, 2049. [Google Scholar] [CrossRef]
- Martin, M.Á.; Ramos, S. Impact of Cocoa Flavanols on Human Health. Food Chem. Toxicol. 2021, 151, 112121. [Google Scholar] [CrossRef] [PubMed]
- Goya, L.; Martín, M.Á.; Sarriá, B.; Ramos, S.; Mateos, R.; Bravo, L. Effect of Cocoa and Its Flavonoids on Biomarkers of Inflammation: Studies of Cell Culture, Animals and Humans. Nutrients 2016, 8, 212. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA) Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the Substantiation of a Health Claim Related to Cocoa Flavanols and Maintenance of Normal Endothelium-Dependent Vasodilation Pursuant to Article 13(5) of Regulation (EC) No 1924/2006. EFSA J. 2012, 10, 2809. [Google Scholar] [CrossRef] [Green Version]
- Skopiński, P.; Skopińska-Rózewska, E.; Sommer, E.; Chorostowska-Wynimko, J.; Rogala, E.; Cendrowska, I.; Chrystowska, D.; Filewska, M.; Białas-Chromiec, B.; Bany, J. Chocolate Feeding of Pregnant Mice Influences Length of Limbs of Their Progeny. Pol. J. Vet. Sci. 2003, 6, 57–59. [Google Scholar] [PubMed]
- Skopińska-Różewska, E.; Bałan, B.J.; Sommer, E.; Chorostowska-Wynimko, J.; Bany, J.; Wasiutyński, A.; Siwicki, A.K. The Influence of Chocolate Feeding of Pregnant Mice on the Immunological Response of Their Progeny. Polish J. Food Nutr. Sci. 2004, 54, 67–70. [Google Scholar]
- Patera, J.; Chorostowska-Wynimko, J.; Słodkowska, J.; Borowska, A.; Skopiński, P.; Sommer, E.; Wasiutyński, A.; Skopińska-Rózewska, E. Morphometric and Functional Abnormalities of Kidneys in the Progeny of Mice Fed Chocolate during Pregnancy and Lactation. Folia Histochem. Cytobiol. 2006, 44, 207–211. [Google Scholar]
- Skopiński, P.; Skopińska-Rózewska, E.; Sommer, E.; Chorostowska-Wynimko, J.; Rogala, E.; Cendrowska, I.; Chrystowska, D.; Filewska, M.; Białas-Chromiec, B.; Bany, J. Chocolate Feeding of Pregnant Mice Resulted in Epigallocatechin-Related Embryonic Angiogenesis Suppression and Bone Mineralization Disorder. Pol. J. Vet. Sci. 2004, 7, 131–133. [Google Scholar]
- Lewicka, A.; Szymański, Ł.; Rusiecka, K.; Kucza, A.; Jakubczyk, A.; Zdanowski, R.; Lewicki, S. Supplementation of Plants with Immunomodulatory Properties during Pregnancy and Lactation-Maternal and Offspring Health Effects. Nutrients 2019, 11, 1958. [Google Scholar] [CrossRef] [Green Version]
- Arola-Arnal, A.; Oms-Oliu, G.; Crescenti, A.; del Bas, J.M.; Ras, M.R.; Arola, L.; Caimari, A. Distribution of Grape Seed Flavanols and Their Metabolites in Pregnant Rats and Their Fetuses. Mol. Nutr. Food Res. 2013, 57, 1741–1752. [Google Scholar] [CrossRef]
- Crescenti, A.; del Bas, J.M.; Arola-Arnal, A.; Oms-Oliu, G.; Arola, L.; Caimari, A. Grape Seed Procyanidins Administered at Physiological Doses to Rats during Pregnancy and Lactation Promote Lipid Oxidation and Up-Regulate AMPK in the Muscle of Male Offspring in Adulthood. J. Nutr. Biochem. 2015, 26, 912–920. [Google Scholar] [CrossRef]
- Del Bas, J.M.; Crescenti, A.; Arola-Arnal, A.; Oms-Oliu, G.; Arola, L.; Caimari, A. Grape Seed Procyanidin Supplementation to Rats Fed a High-Fat Diet during Pregnancy and Lactation Increases the Body Fat Content and Modulates the Inflammatory Response and the Adipose Tissue Metabolism of the Male Offspring in Youth. Int. J. Obes. 2015, 39, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Del Bas, J.M.; Crescenti, A.; Arola-Arnal, A.; Oms-Oliu, G.; Arola, L.; Caimari, A. Intake of Grape Procyanidins during Gestation and Lactation Impairs Reverse Cholesterol Transport and Increases Atherogenic Risk Indexes in Adult Offspring. J. Nutr. Biochem. 2015, 26, 1670–1677. [Google Scholar] [CrossRef] [PubMed]
- Sampey, B.P.; Vanhoose, A.M.; Winfield, H.M.; Freemerman, A.J.; Muehlbauer, M.J.; Fueger, P.T.; Newgard, C.B.; Makowski, L. Cafeteria Diet Is a Robust Model of Human Metabolic Syndrome with Liver and Adipose Inflammation: Comparison to High-Fat Diet. Obesity 2011, 19, 1109–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caimari, A.; Mariné-Casadó, R.; Boqué, N.; Crescenti, A.; Arola, L.; Del Bas, J.M. Maternal Intake of Grape Seed Procyanidins during Lactation Induces Insulin Resistance and an Adiponectin Resistance-like Phenotype in Rat Offspring. Sci. Rep. 2017, 7, 12573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose Translation from Animal to Human Studies Revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, N.; Segarra, A.B.; Lara, L.; Ramírez-Sánchez, M.; Prieto, I. Diet and Oxidative Status. The Dietary Pattern and Urinary 8-Isoprostane in Healthy Spanish Women. Antioxidants 2019, 8, 271. [Google Scholar] [CrossRef] [Green Version]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis Model Assessment: Insulin Resistance and β-Cell Function from Fasting Plasma Glucose and Insulin Concentrations in Man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Perseghin, G.; Caumo, A.; Caloni, M.; Testolin, G. Incorporation of the Fasting Plasma FFA Concentration into QUICKI Improves Its Association with Insulin Sensitivity in Nonobese Individuals. J. Clin. Endocrinol. Metab. 2001, 86, 4776–4781. [Google Scholar] [CrossRef]
- MM, B. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hara, A.; Radin, N.S. Lipid Extraction of Tissues with a Low-Toxicity Solvent. Anal. Biochem. 1978, 90, 420–426. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Sureda, V.; Peinado-Onsurbe, J. A Procedure for Measuring Triacylglyceride and Cholesterol Content Using a Small Amount of Tissue. Anal. Biochem. 2005, 343, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Caimari, A.; Oliver, P.; Palou, A. Adipose Triglyceride Lipase Expression and Fasting Regulation Are Differently Affected by Cold Exposure in Adipose Tissues of Lean and Obese Zucker Rats. J. Nutr. Biochem. 2012, 23, 1041–1050. [Google Scholar] [CrossRef] [PubMed]
- Palacios-Jordan, H.; Martín-González, M.Z.; Suárez, M.; Aragonès, G.; Mugureza, B.; Rodríguez, M.A.; Bladé, C. The Disruption of Liver Metabolic Circadian Rhythms by a Cafeteria Diet Is Sex-Dependent in Fischer 344 Rats. Nutrients 2020, 12, 1085. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0-Making Metabolomics More Meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [Green Version]
- Worley, B.; Powers, R. Multivariate Analysis in Metabolomics. Curr. Metabolomics 2013, 1, 92–107. [Google Scholar] [CrossRef]
- Zhao, S.; Kusminski, C.M.; Scherer, P.E. Adiponectin, Leptin and Cardiovascular Disorders. Circ. Res. 2021, 128, 136–149. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Zhang, Q.; Aguilera, Y.; Martín-Cabrejas, M.A.; Gonzalez de Mejia, E. Relationship of the Phytochemicals from Coffee and Cocoa By-Products with their Potential to Modulate Biomarkers of Metabolic Syndrome In Vitro. Antioxidants 2019, 8, 279. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Yu, S.; Lambert, J.D. Dietary Cocoa Ameliorates Obesity-Related Inflammation in High Fat-Fed Mice. Eur. J. Nutr. 2014, 53, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Campbell, C.L.; Foegeding, E.A.; Harris, G.K. Cocoa and Whey Protein Differentially Affect Markers of Lipid and Glucose Metabolism and Satiety. J. Med. Food 2016, 19, 219–227. [Google Scholar] [CrossRef]
- Eskandari, M.; Moghadam, B.H.; Bagheri, R.; Ashtary-Larky, D.; Eskandari, E.; Nordvall, M.; Dutheil, F.; Wong, A. Effects of Interval Jump Rope Exercise Combined with Dark Chocolate Supplementation on Inflammatory Adipokine, Cytokine Concentrations, and Body Composition in Obese Adolescent Boys. Nutrients 2020, 12, 3011. [Google Scholar] [CrossRef]
- Aminuddin, F.; Ali, F.; Ismail, A.; Pei, C.P.; Hamid, M. Cocoa Polyphenol-Rich Extract Enhances the Expression Levels of PPAR-γ in the Skeletal Muscle and Adipose Tissue of Obese-Diabetic Rats Fed a High-Fat Diet. Int. J. Pharmacol. 2015, 11, 309–317. [Google Scholar] [CrossRef]
- Rosa, S.C.D.S.; Liu, M.; Sweeney, G. Adiponectin Synthesis, Secretion and Extravasation from Circulation to Interstitial Space. Physiology 2021, 36, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Du, Y.; Schwaid, A.G.; Asterholm, I.W.; Scherer, P.E.; Saghatelian, A.; Wan, Y. Maternal Adiponectin Controls Milk Composition to Prevent Neonatal Inflammation. Endocrinology 2015, 156, 1504. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.W.; Millward, C.A.; DeSantis, D.; Pisano, S.; Machova, J.; Perales, J.C.; Croniger, C.M. Reduced Milk Triglycerides in Mice Lacking Phosphoenolpyruvate Carboxykinase in Mammary Gland Adipocytes and White Adipose Tissue Contribute to the Development of Insulin Resistance in Pups. J. Nutr. 2009, 139, 2257. [Google Scholar] [CrossRef] [Green Version]
- Gessner, D.K.; Brock, C.; Hof, L.M.; Most, E.; Koch, C.; Eder, K. Effects of Supplementation of Green Tea Extract on the Milk Performance of Peripartal Dairy Cows and the Expression of Stress Response Genes in the Liver. J. Anim. Sci. Biotechnol. 2020, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Huynh, H.D.; Wan, Y. Milk Lipid Regulation at the Maternal-Offspring Interface. Semin. Cell Dev. Biol. 2018, 81, 141–148. [Google Scholar] [CrossRef]
- Argov-Argaman, N.; Cohen-Zinder, M.; Leibovich, H.; Yishay, M.; Eitam, H.; Agmon, R.; Hadaya, O.; Mesilati-Stahy, R.; Miron, J.; Shabtay, A. Dietary Pomegranate Peel Improves Milk Quality of Lactating Ewes: Emphasis on Milk Fat Globule Membrane Properties and Antioxidative Traits. Food Chem. 2020, 313, 125822. [Google Scholar] [CrossRef]
- Li, M.; Hassan, F.U.; Tang, Z.; Peng, L.; Liang, X.; Li, L.; Peng, K.; Xie, F.; Yang, C. Mulberry Leaf Flavonoids Improve Milk Production, Antioxidant, and Metabolic Status of Water Buffaloes. Front. Vet. Sci. 2020, 7, 599. [Google Scholar] [CrossRef]
- Renna, M.; Lussiana, C.; Colonna, L.; Malfatto, V.M.; Mimosi, A.; Cornale, P. Inclusion of Cocoa Bean Shell in the Diet of Dairy Goats: Effects on Milk Production Performance and Milk Fatty Acid Profile. Front. Vet. Sci. 2022, 9, 1–15. [Google Scholar] [CrossRef]
- Gessner, D.K.; Koch, C.; Romberg, F.J.; Winkler, A.; Dusel, G.; Herzog, E.; Most, E.; Eder, K. The Effect of Grape Seed and Grape Marc Meal Extract on Milk Performance and the Expression of Genes of Endoplasmic Reticulum Stress and Inflammation in the Liver of Dairy Cows in Early Lactation. J. Dairy Sci. 2015, 98, 8856–8868. [Google Scholar] [CrossRef] [Green Version]
- Neville, M.C.; Picciano, M.F. Regulation of Milk Lipid Secretion and Composition. Annu. Rev. Nutr. 1997, 17, 159–184. [Google Scholar] [CrossRef] [PubMed]
- McManaman, J.L. Lipid Transport in the Lactating Mammary Gland. J. Mammary Gland Biol. Neoplasia 2014, 19, 35–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudolph, M.C.; McManaman, J.L.; Phang, T.L.; Russell, T.; Kominsky, D.J.; Serkova, N.J.; Stein, T.; Anderson, S.M.; Neville, M.C. Metabolic Regulation in the Lactating Mammary Gland: A Lipid Synthesizing Machine. Physiol. Genom. 2007, 28, 323–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miles, J.M.; Park, Y.S.; Walewicz, D.; Russell-Lopez, C.; Windsor, S.; Isley, W.L.; Coppack, S.W.; Harris, W.S. Systemic and Forearm Triglyceride MetabolismFate of Lipoprotein Lipase-Generated Glycerol and Free Fatty Acids. Diabetes 2004, 53, 521–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shetty, S.; Ramos-Roman, M.A.; Cho, Y.R.; Brown, J.; Plutzky, J.; Muise, E.S.; Horton, J.D.; Scherer, P.E.; Parks, E.J. Enhanced Fatty Acid Flux Triggered by Adiponectin Overexpression. Endocrinology 2012, 153, 113–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker-Duffen, J.L.; Nakamura, K.; Silver, M.; Kikuchi, R.; Tigges, U.; Yoshida, S.; Denzel, M.S.; Ranscht, B.; Walsh, K. T-Cadherin Is Essential for Adiponectin-Mediated Revascularization. J. Biol. Chem. 2013, 288, 24886–24897. [Google Scholar] [CrossRef] [Green Version]
- Halberg, N.; Schraw, T.D.; Wang, Z.V.; Kim, J.Y.; Yi, J.; Hamilton, M.P.; Luby-Phelps, K.; Scherer, P.E. Systemic Fate of the Adipocyte-Derived Factor Adiponectin. Diabetes 2009, 58, 1961–1970. [Google Scholar] [CrossRef] [Green Version]
- Turer, A.T.; Scherer, P.E. Adiponectin: Mechanistic Insights and Clinical Implications. Diabetologia 2012, 55, 2319–2326. [Google Scholar] [CrossRef] [Green Version]
- Higa, T.S.; Spinola, A.V.; Fonseca-Alaniz, M.H.; Anna Evangelista, F.S. Comparison between Cafeteria and High-Fat Diets in the Induction of Metabolic Dysfunction in Mice. Int. J. Physiol. Pathophysiol. Pharmacol. 2014, 6, 47. [Google Scholar]
- Yanai, H.; Yoshida, H. Beneficial Effects of Adiponectin on Glucose and Lipid Metabolism and Atherosclerotic Progression: Mechanisms and Perspectives. Int. J. Mol. Sci. 2019, 20, 1190. [Google Scholar] [CrossRef] [Green Version]
- Dorenkott, M.R.; Griffin, L.E.; Goodrich, K.M.; Thompson-Witrick, K.A.; Fundaro, G.; Ye, L.; Stevens, J.R.; Ali, M.; O’Keefe, S.F.; Hulver, M.W.; et al. Oligomeric Cocoa Procyanidins Possess Enhanced Bioactivity Compared to Monomeric and Polymeric Cocoa Procyanidins for Preventing the Development of Obesity, Insulin Resistance, and Impaired Glucose Tolerance during High-Fat Feeding. J. Agric. Food Chem. 2014, 62, 2216–2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomaru, M.; Takano, H.; Osakabe, N.; Yasuda, A.; Inoue, K.I.; Yanagisawa, R.; Ohwatari, T.; Uematsu, H. Dietary Supplementation with Cacao Liquor Proanthocyanidins Prevents Elevation of Blood Glucose Levels in Diabetic Obese Mice. Nutrition 2007, 23, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, J.; Alves, M.G.; Oliveira, P.F.; Silva, B.M. Pharmacological Potential of Methylxanthines: Retrospective Analysis and Future Expectations. Crit. Rev. Food Sci. Nutr. 2018, 59, 2597–2625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, R.; Oñatibia-Astibia, A.; Martínez-Pinilla, E. Health Benefits of Methylxanthines in Cacao and Chocolate. Nutrients 2013, 5, 4159. [Google Scholar] [CrossRef] [PubMed]
- López, N.; Sánchez, J.; Picó, C.; Palou, A.; Serra, F. Dietary L-Leucine Supplementation of Lactating Rats Results in a Tendency to Increase Lean/Fat Ratio Associated to Lower Orexigenic Neuropeptide Expression in Hypothalamus. Peptides 2010, 31, 1361–1367. [Google Scholar] [CrossRef]
- Krause, M.P.; Milne, K.J.; Hawke, T.J. Adiponectin-Consideration for Its Role in Skeletal Muscle Health. Int. J. Mol. Sci. 2019, 20, 1528. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Kita, S.; Nishizawa, H.; Fukuda, S.; Fujishima, Y.; Obata, Y.; Nagao, H.; Masuda, S.; Nakamura, Y.; Shimizu, Y.; et al. Adiponectin Promotes Muscle Regeneration through Binding to T-Cadherin. Sci. Rep. 2019, 9, 16. [Google Scholar] [CrossRef] [Green Version]
- Inoue, A.; Cheng, X.W.; Huang, Z.; Hu, L.; Kikuchi, R.; Jiang, H.; Piao, L.; Sasaki, T.; Itakura, K.; Wu, H.; et al. Exercise Restores Muscle Stem Cell Mobilization, Regenerative Capacity and Muscle Metabolic Alterations via Adiponectin/AdipoR1 Activation in SAMP10 Mice. J. Cachexia. Sarcopenia Muscle 2017, 8, 370–385. [Google Scholar] [CrossRef]
- Ito, R.; Higa, M.; Goto, A.; Aoshima, M.; Ikuta, A.; Ohashi, K.; Yokoyama, S.; Ohno, Y.; Egawa, T.; Miyata, H.; et al. Activation of Adiponectin Receptors Has Negative Impact on Muscle Mass in C2C12 Myotubes and Fast-Type Mouse Skeletal Muscle. PLoS ONE 2018, 13, e0205645. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Tomata, Y.; Kakizaki, M.; Sugawara, Y.; Hozawa, A.; Momma, H.; Tsuji, I.; Nagatomi, R. High Circulating Adiponectin Levels Predict Decreased Muscle Strength among Older Adults Aged 70 Years and over: A Prospective Cohort Study. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 594–601. [Google Scholar] [CrossRef] [Green Version]
- Frühbeck, G.; Catalán, V.; Rodríguez, A.; Gómez-Ambrosi, J. Adiponectin-Leptin Ratio: A Promising Index to Estimate Adipose Tissue Dysfunction. Relation with Obesity-Associated Cardiometabolic Risk. Adipocyte 2018, 7, 57. [Google Scholar] [CrossRef] [PubMed]
- Frühbeck, G.; Catalán, V.; Rodríguez, A.; Ramírez, B.; Becerril, S.; Salvador, J.; Colina, I.; Gómez-Ambrosi, J. Adiponectin-Leptin Ratio Is a Functional Biomarker of Adipose Tissue Inflammation. Nutrients 2019, 11, 454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodgers, J.L.; Jones, J.; Bolleddu, S.I.; Vanthenapalli, S.; Rodgers, L.E.; Shah, K.; Karia, K.; Panguluri, S.K. Cardiovascular Risks Associated with Gender and Aging. J. Cardiovasc. Dev. Dis. 2019, 6, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez-Moreno, E.; Arias-Rico, J.; Jiménez-Sánchez, R.C.; Estrada-Luna, D.; Jiménez-Osorio, A.S.; Zafra-Rojas, Q.Y.; Ariza-Ortega, J.A.; Flores-Chávez, O.R.; Morales-Castillejos, L.; Sandoval-Gallegos, E.M. Role of Bioactive Compounds in Obesity: Metabolic Mechanism Focused on Inflammation. Foods 2022, 11, 1232. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.B.A.R.; Pinheiro-Castro, N.; Novaes, G.M.; Pascoal, G.D.F.L.; Ong, T.P. Bioactive Food Compounds, Epigenetics and Chronic Disease Prevention: Focus on Early-Life Interventions with Polyphenols. Food Res. Int. 2019, 125, 108646. [Google Scholar] [CrossRef] [PubMed]
- Villarreal-Molina, M.T.; Antuna-Puente, B. Adiponectin: Anti-Inflammatory and Cardioprotective Effects. Biochimie 2012, 94, 2143–2149. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, K.; Ouchi, N.; Matsuzawa, Y. Anti-Inflammatory and Anti-Atherogenic Properties of Adiponectin. Biochimie 2012, 94, 2137–2142. [Google Scholar] [CrossRef]
- Chawla, A.; Nguyen, K.D.; Goh, Y.P.S. Macrophage-Mediated Inflammation in Metabolic Disease. Nat. Rev. Immunol. 2011, 11, 738–749. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic Inflammation in Fat Plays a Crucial Role in the Development of Obesity-Related Insulin Resistance. J. Clin. Investig. 2003, 112, 1821. [Google Scholar] [CrossRef]
- Gordon, S. Alternative Activation of Macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef]
- Lovren, F.; Pan, Y.; Quan, A.; Szmitko, P.E.; Singh, K.K.; Shukla, P.C.; Gupta, M.; Chan, L.; Al-Omran, M.; Teoh, H.; et al. Adiponectin Primes Human Monocytes into Alternative Anti-Inflammatory M2 Macrophages. Am. J. Physiol.-Hear. Circ. Physiol. 2010, 299, H656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Liu, M. Adiponectin: A Versatile Player of Innate Immunity. J. Mol. Cell Biol. 2016, 8, 120–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holling, T.M.; Schooten, E.; Van Den Elsen, P.J. Function and Regulation of MHC Class II Molecules in T-Lymphocytes: Of Mice and Men. Hum. Immunol. 2004, 65, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Anshita, D.; Ravichandiran, V. MCP-1: Function, Regulation, and Involvement in Disease. Int. Immunopharmacol. 2021, 101, 107598. [Google Scholar] [CrossRef] [PubMed]
- Číž, M.; Dvořáková, A.; Skočková, V.; Kubala, L. The Role of Dietary Phenolic Compounds in Epigenetic Modulation Involved in Inflammatory Processes. Antioxidants 2020, 9, 691. [Google Scholar] [CrossRef]
- Alonso-Bernáldez, M.; Asensio, A.; Palou-March, A.; Sánchez, J.; Palou, A.; Serra, F.; Palou, M. Breast Milk MicroRNAs Related to Leptin and Adiponectin Function Can Be Modulated by Maternal Diet and Influence Offspring Phenotype in Rats. Int. J. Mol. Sci. 2022, 23, 7237. [Google Scholar] [CrossRef]
- Hsu, C.N.; Hou, C.Y.; Hsu, W.H.; Tain, Y.L. Early-Life Origins of Metabolic Syndrome: Mechanisms and Preventive Aspects. Int. J. Mol. Sci. 2021, 22, 11872. [Google Scholar] [CrossRef]
- Rodríguez-Daza, M.C.; Pulido-Mateos, E.C.; Lupien-Meilleur, J.; Guyonnet, D.; Desjardins, Y.; Roy, D. Polyphenol-Mediated Gut Microbiota Modulation: Toward Prebiotics and Further. Front. Nutr. 2021, 8, 689456. [Google Scholar] [CrossRef]
- Wahlig, J.L.; Bales, E.S.; Jackman, M.R.; Johnson, G.C.; McManaman, J.L.; MacLean, P.S. Impact of High-Fat Diet and Obesity on Energy Balance and Fuel Utilization during the Metabolic Challenge of Lactation. Obesity 2012, 20, 65–75. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VEH | CCX | Student’s t-Test (p) | |
|---|---|---|---|
| Biometric parameters | |||
| Initial body weight (g) | 278 ± 3 | 278 ± 6 | 0.926 |
| Final body weight (g) | 286 ± 3 | 283 ± 5 | 0.561 |
| Body weight gain (g) | 8.00 ± 3.78 | 5.11 ± 5.28 | 0.662 |
| RWAT (g) | 3.15 ± 0.41 | 3.18 ± 0.39 | 0.954 |
| MWAT (g) | 1.87 ± 0.14 | 1.67 ± 0.17 | 0.388 |
| OWAT (g) | 4.93 ± 0.64 | 4.77 ± 0.39 | 0.830 |
| Adiposity index (%) | 3.81 ± 0.45 | 3.83 ± 0.39 | 0.977 |
| Mammary gland (g) | 11.0 ± 0.3 | 11.7 ± 0.3 | 0.096 |
| Liver (g) | 10.4 ± 0.1 | 10.0 ± 0.2 | 0.129 |
| Lean mass (g) | 249 ± 4 | 243 ± 6 | 0.452 |
| Lean mass (%) | 86.3 ± 0.7 | 85.9 ± 0.7 | 0.725 |
| Fat mass (g) | 23.1 ± 1.6 | 23.0 ± 2.9 | 0.987 |
| Fat mass (%) | 8.01 ± 0.59 | 8.20 ± 1.07 | 0.885 |
| Plasma parameters | |||
| Glucose (mmol/L) | 5.93 ± 0.15 | 5.57 ± 0.24 | 0.228 |
| Insulin (mU/L) | 43.9 ± 6.6 | 57.9 ± 19.7 | 0.516 |
| FFAs (mmol/L) | 0.218 ± 0.025 | 0.348 ± 0.043 * | 0.020 |
| HOMA-IR | 11.7 ± 2.0 | 15.5 ± 5.5 | 0.530 |
| R-QUICKI | 0.338 ± 0.007 | 0.339 ± 0.022 | 0.973 |
| Total adiponectin (µg/mL) | 17.6 ± 0.7 | 23.9 ± 1.9 * | 0.010 |
| HMW adiponectin (μg/mL) | 2.99 ± 0.55 | 3.93 ± 1.03 | 0.401 |
| Total adiponectin/g WAT | 1.72 ± 0.17 | 2.31 ± 0.20 * | 0.041 |
| Triglycerides (mmol/L) | 0.375 ± 0.024 | 0.353 ± 0.029 | 0.547 |
| Phospholipids (mmol/L) | 2.11 ± 0.07 | 2.03 ± 0.08 | 0.489 |
| Total cholesterol (mmol/L) | 2.21 ± 0.11 | 1.96 ± 0.07 | 0.065 |
| Energy intake (kcal/day) | 155 ± 2 | 147 ± 5 | 0.148 |
| STD-veh | STD-CCX | CAF-veh | CAF-CCX | Two-Way ANOVA | |
|---|---|---|---|---|---|
| Biometric parameters | |||||
| Initial body weight (g) | 39.7 ± 1.9 | 41.3 ± 1.3 | 41.0 ± 1.4 | 40.8 ± 1.1 | - |
| Final body weight (g) | 373 ± 9 | 371 ± 4 | 446 ± 12 | 433 ± 14 | D |
| Body weight gain (g) | 333 ± 8 | 330 ± 4 | 405 ± 12 | 392 ± 14 | D |
| Liver (g) | 12.0 ± 0.4 | 10.9 ± 0.3 | 14.3 ± 0.5 | 13.2 ± 0.4 | D, P |
| RWAT (g) | 6.47 ± 0.45 | 7.87 ± 0.61 | 18.6 ± 1.7 | 19.0 ± 1.4 | D |
| MWAT (g) | 4.28 ± 0.27 | 4.75 ± 0.33 | 8.46 ± 0.57 | 9.21 ± 0.80 | D |
| EWAT (g) | 5.67 ± 0.48 | 6.86 ± 0.57 | 14.9 ± 1.3 | 15.8 ± 1.5 | D |
| IWAT (g) | 11.9 ± 1.0 | 12.4 ± 1.0 | 29.2 ± 3.0 | 30.0 ± 2.4 | D |
| Adiposity index (%) | 7.75 ± 0.54 | 8.69 ± 0.61 | 16.1 ± 1.0 | 17.2 ± 0.9 | D |
| Soleus muscle (g) | 0.147 ± 0.006 | 0.135 ± 0.005 | 0.142 ± 0.004 | 0.134 ± 0.002 | P |
| Gastrocnemius muscle (g) | 1.74 ± 0.05 | 1.76 ± 0.03 | 1.86 ± 0.02 | 1.79 ± 0.03 | D |
| Lean mass (%) | 86.2 ± 0.8 | 85.0 ± 1.0 | 74.8 ± 1.4 | 73.2 ± 1.2 | D |
| Fat mass (%) | 8.64 ± 0.76 | 9.86 ± 1.04 | 20.5 ± 1.4 | 22.4 ± 1.3 | D |
| Lean/fat ratio | 10.8 ± 0.9 | 8.62 ± 0.81 | 3.94 ± 0.35 | 3.46 ± 0.29 | D, P |
| Plasma parameters | |||||
| Glucose (mmol/L) | 6.00 ± 0.34 | 6.09 ± 0.25 | 7.41 ± 0.34 | 7.72 ± 0.13 | D |
| Insulin (mU/L) | 77.2 ± 8.4 | 94.9 ± 10.3 | 127 ± 10 | 141 ± 8 | D |
| HOMA-IR | 20.4 ± 2.5 | 26.3 ± 3.8 | 43.0 ± 5.2 | 48.4 ± 3.0 | D |
| R-QUICKI | 0.306 ± 0.008 | 0.291 ± 0.004 | 0.269 ± 0.004 | 0.270 ± 0.003 | D |
| Total cholesterol (mmol/L) | 1.65 ± 0.11 | 1.68 ± 0.07 | 2.20 ± 0.09 | 2.09 ± 0.08 | D |
| Triglycerides (mmol/L) | 1.75 ± 0.32 | 1.15 ± 0.22 | 3.67 ± 0.39 | 3.17 ± 0.25 | D |
| Free fatty acids (mmol/L) | 0.279 ± 0.035 | 0.272 ± 0.030 | 0.336 ± 0.022 | 0.280 ± 0.022 | - |
| Phospholipids (mmol/L) | 1.77 ± 0.09 | 1.72 ± 0.08 | 2.38 ± 0.12 | 2.51 ± 0.11 | D |
| Leptin (ng/mL) | 9.33 ± 0.63 | 9.89 ± 0.77 | 39.6 ± 3.6 | 45.4 ± 3.1 | D |
| Adiponectin/leptin ratio | 2.65 ± 0.22 | 3.40 ± 0.31 * | 1.18 ± 0.16 $ | 1.09 ± 0.07 | D, P, DxP |
| Energy intake (kcal/day) | 52.8 ± 1.5 | 55.3 ± 1.3 | 112 ± 3 | 113 ± 3 | D |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mariné-Casadó, R.; Domenech-Coca, C.; Crescenti, A.; Rodríguez Gómez, M.Á.; Del Bas, J.M.; Arola, L.; Boqué, N.; Caimari, A. Maternal Supplementation with a Cocoa Extract during Lactation Deeply Modulates Dams’ Metabolism, Increases Adiponectin Circulating Levels and Improves the Inflammatory Profile in Obese Rat Offspring. Nutrients 2022, 14, 5134. https://doi.org/10.3390/nu14235134
Mariné-Casadó R, Domenech-Coca C, Crescenti A, Rodríguez Gómez MÁ, Del Bas JM, Arola L, Boqué N, Caimari A. Maternal Supplementation with a Cocoa Extract during Lactation Deeply Modulates Dams’ Metabolism, Increases Adiponectin Circulating Levels and Improves the Inflammatory Profile in Obese Rat Offspring. Nutrients. 2022; 14(23):5134. https://doi.org/10.3390/nu14235134
Chicago/Turabian StyleMariné-Casadó, Roger, Cristina Domenech-Coca, Anna Crescenti, Miguel Ángel Rodríguez Gómez, Josep Maria Del Bas, Lluís Arola, Noemí Boqué, and Antoni Caimari. 2022. "Maternal Supplementation with a Cocoa Extract during Lactation Deeply Modulates Dams’ Metabolism, Increases Adiponectin Circulating Levels and Improves the Inflammatory Profile in Obese Rat Offspring" Nutrients 14, no. 23: 5134. https://doi.org/10.3390/nu14235134
APA StyleMariné-Casadó, R., Domenech-Coca, C., Crescenti, A., Rodríguez Gómez, M. Á., Del Bas, J. M., Arola, L., Boqué, N., & Caimari, A. (2022). Maternal Supplementation with a Cocoa Extract during Lactation Deeply Modulates Dams’ Metabolism, Increases Adiponectin Circulating Levels and Improves the Inflammatory Profile in Obese Rat Offspring. Nutrients, 14(23), 5134. https://doi.org/10.3390/nu14235134