
Dietary Effects of Chromium Picolinate and Chromium Nanoparticles in Wistar Rats Fed with a High-Fat, Low-Fiber Diet: The Role of Fat Normalization

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Drugs and Chemicals
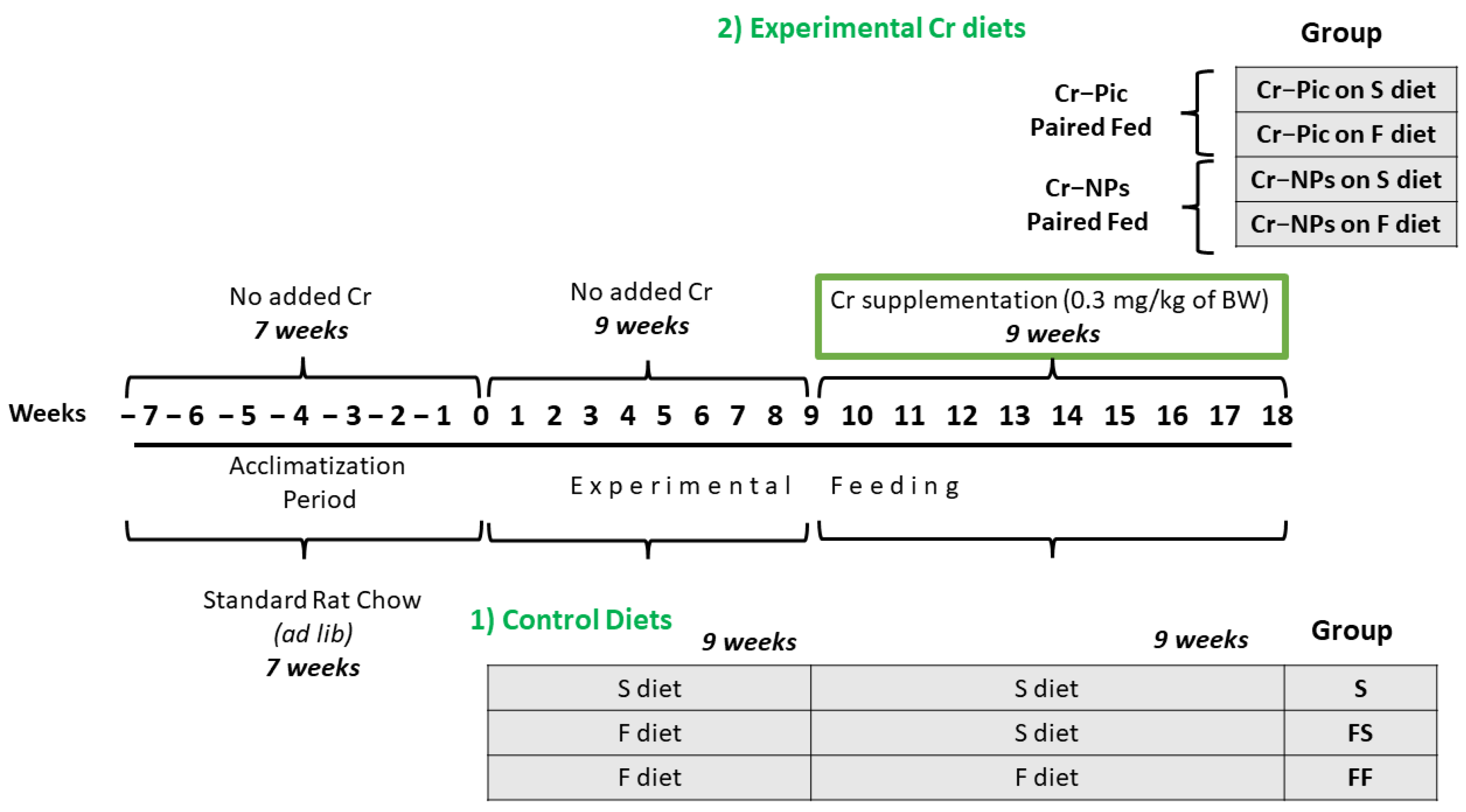
2.2. Animal Protocol and Dietary Treatment
- (A)
- Control, which was further subdivided into:
- 1.
- Control S (the S diet and the S diet)
- 2.
- Control FS (the F diet and the S diet)
- 3.
- Control FF (the F diet and the F diet)
- (B)
- Supplemented, which was further subdivided into:
- 1.
- Cr–Pic FS (the FS diet supplemented with Cr–Pic)
- 2.
- Cr–NPs FS (the FS diet supplemented with Cr–NPs)
- 3.
- Cr–Pic FF (the FF diet supplemented with Cr–Pic)
- 4.
- Cr–NPs FF (the FF diet supplemented with Cr–NPs)
2.3. Experimental Procedures
2.4. Blood Pressure
2.5. The Antioxidant Capacity of Blood Plasma
2.6. Markers of Antioxidant Status in the Heart
2.7. Vascular Reactivity Studies
2.8. Biochemical Studies of Aortic Rings
2.8.1. TXA2 and PGI2 Production
2.8.2. Detection of Superoxide Anion
2.8.3. Detection of Hydrogen Peroxide
2.9. Data Analysis and Statistics
3. Results
3.1. General Characteristics
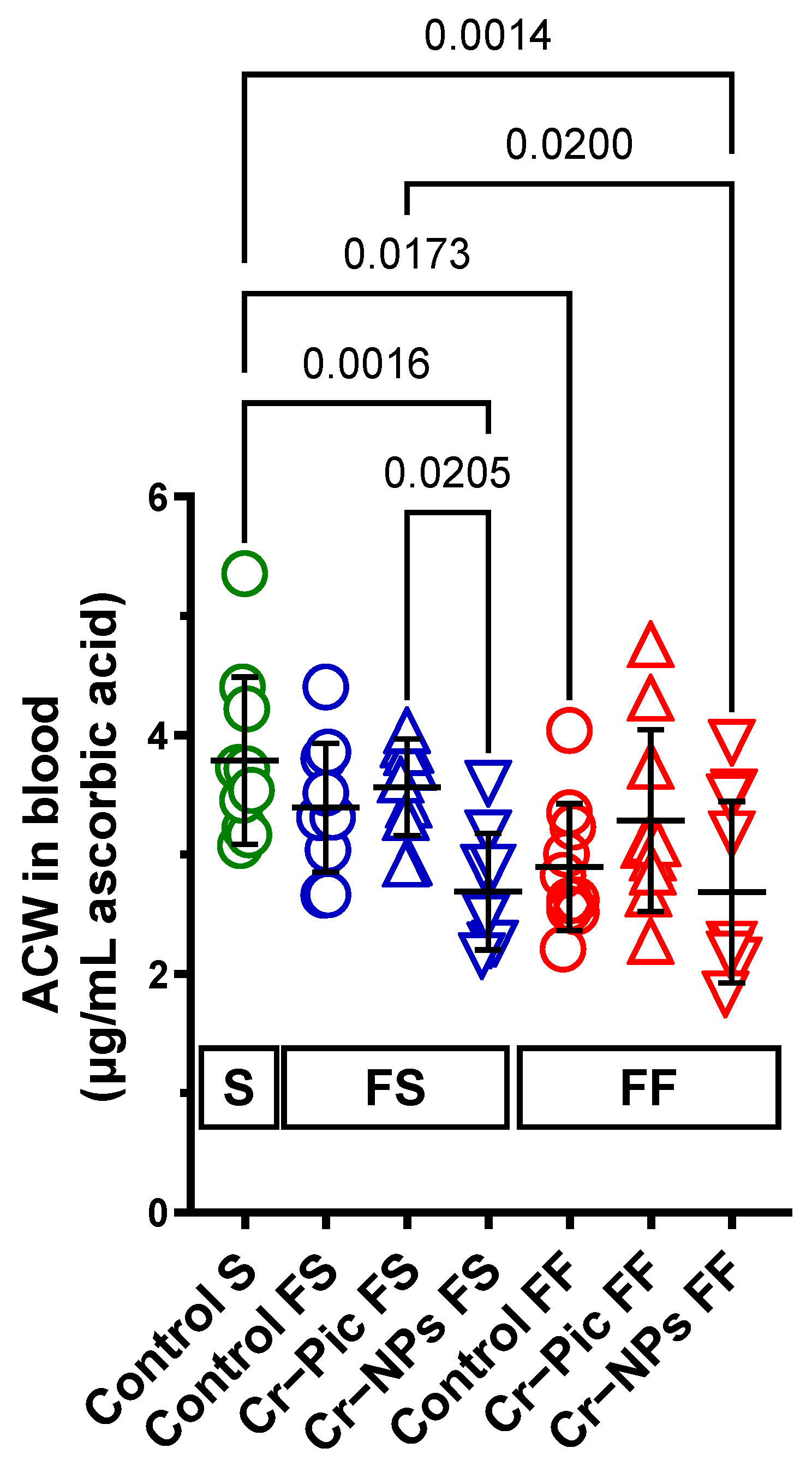
3.2. The Antioxidant Capacity of Blood Plasma
3.2.1. Blood Plasma ACW
3.2.2. Blood Plasma ACL
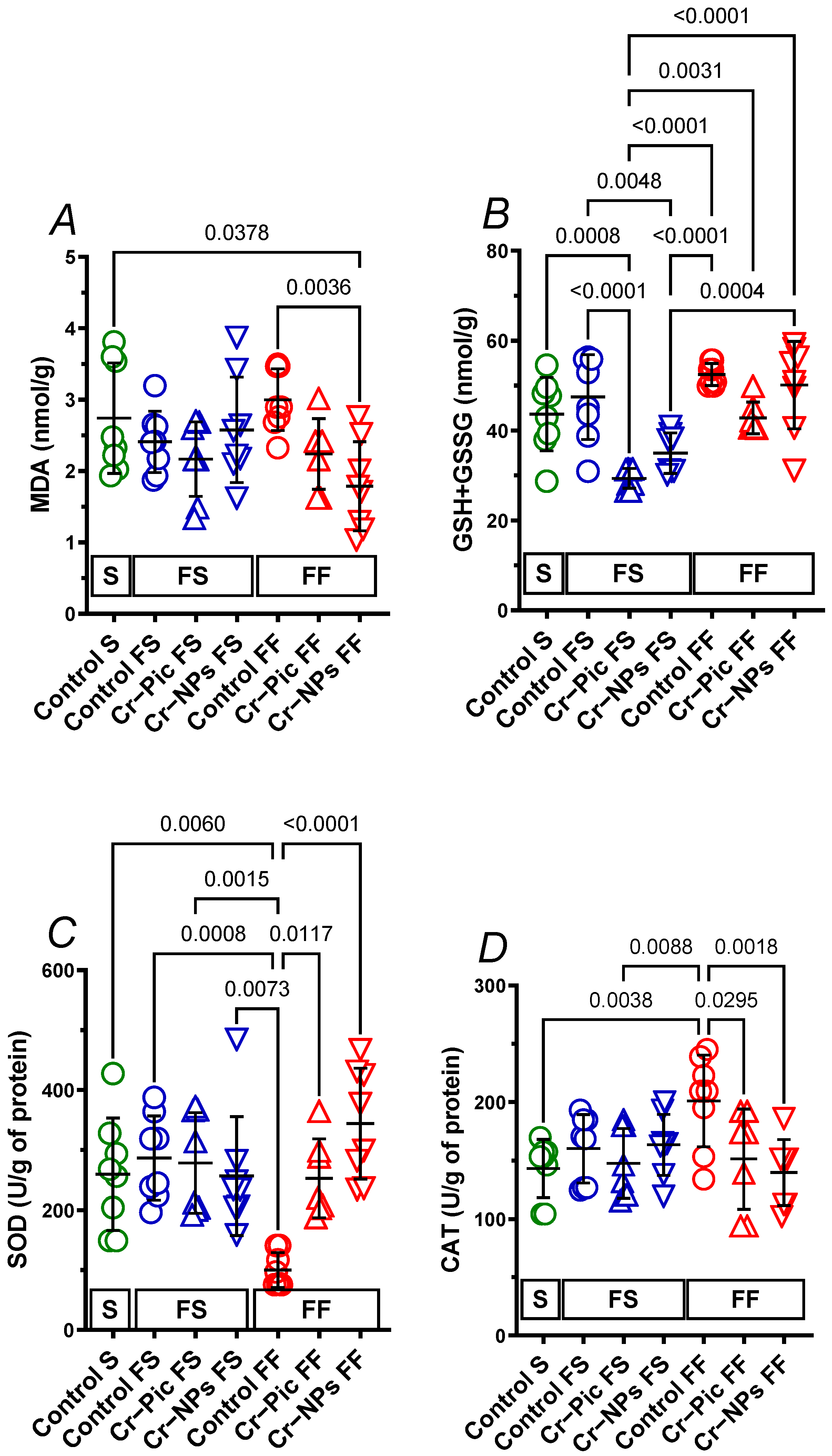
3.3. Markers of Antioxidant Status in the Heart
3.3.1. Heart Malondialdehyde
3.3.2. Heart GSH+GSSG
3.3.3. Heart SOD
3.3.4. Heart CAT
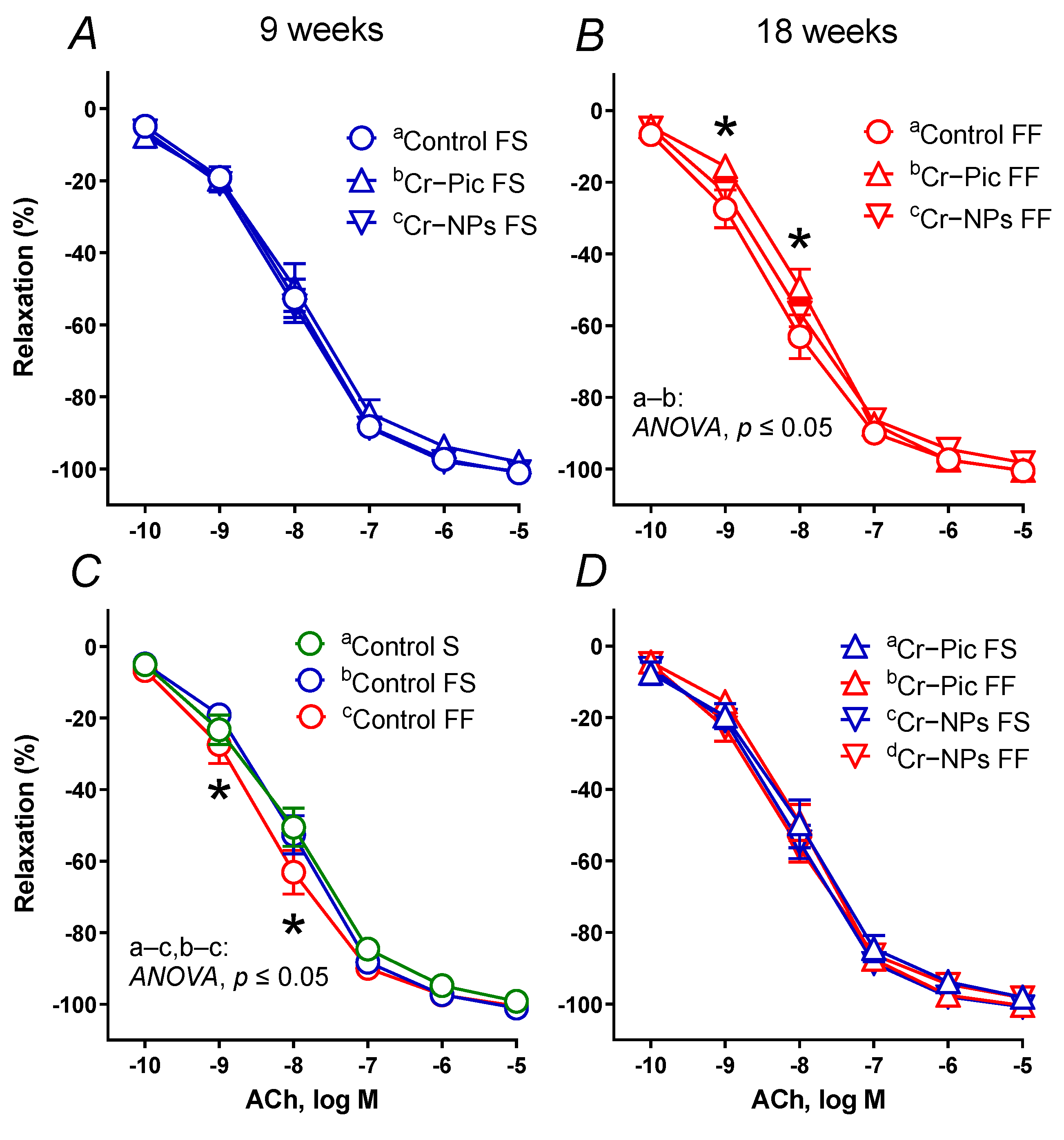
3.4. Vascular Studies
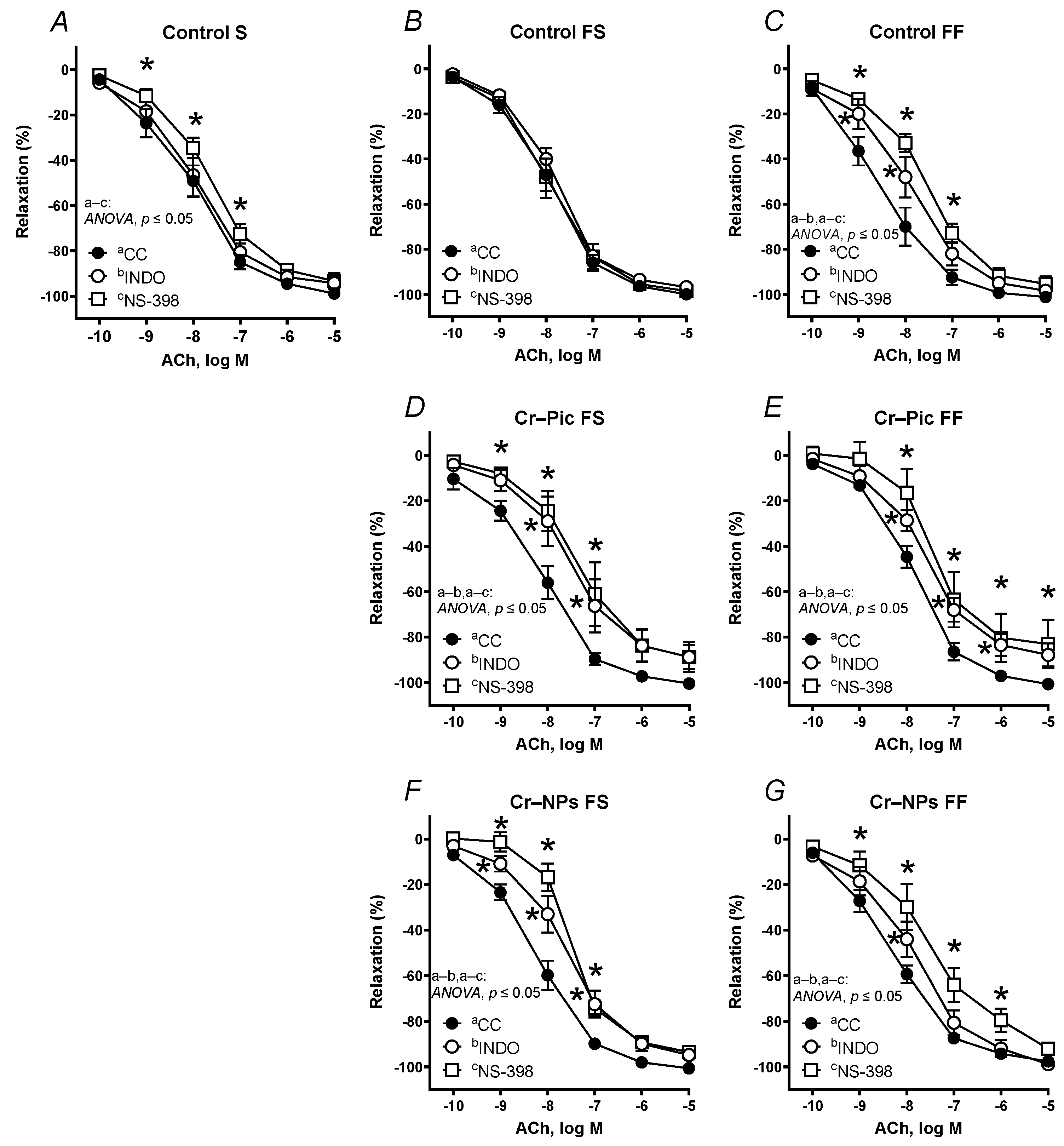
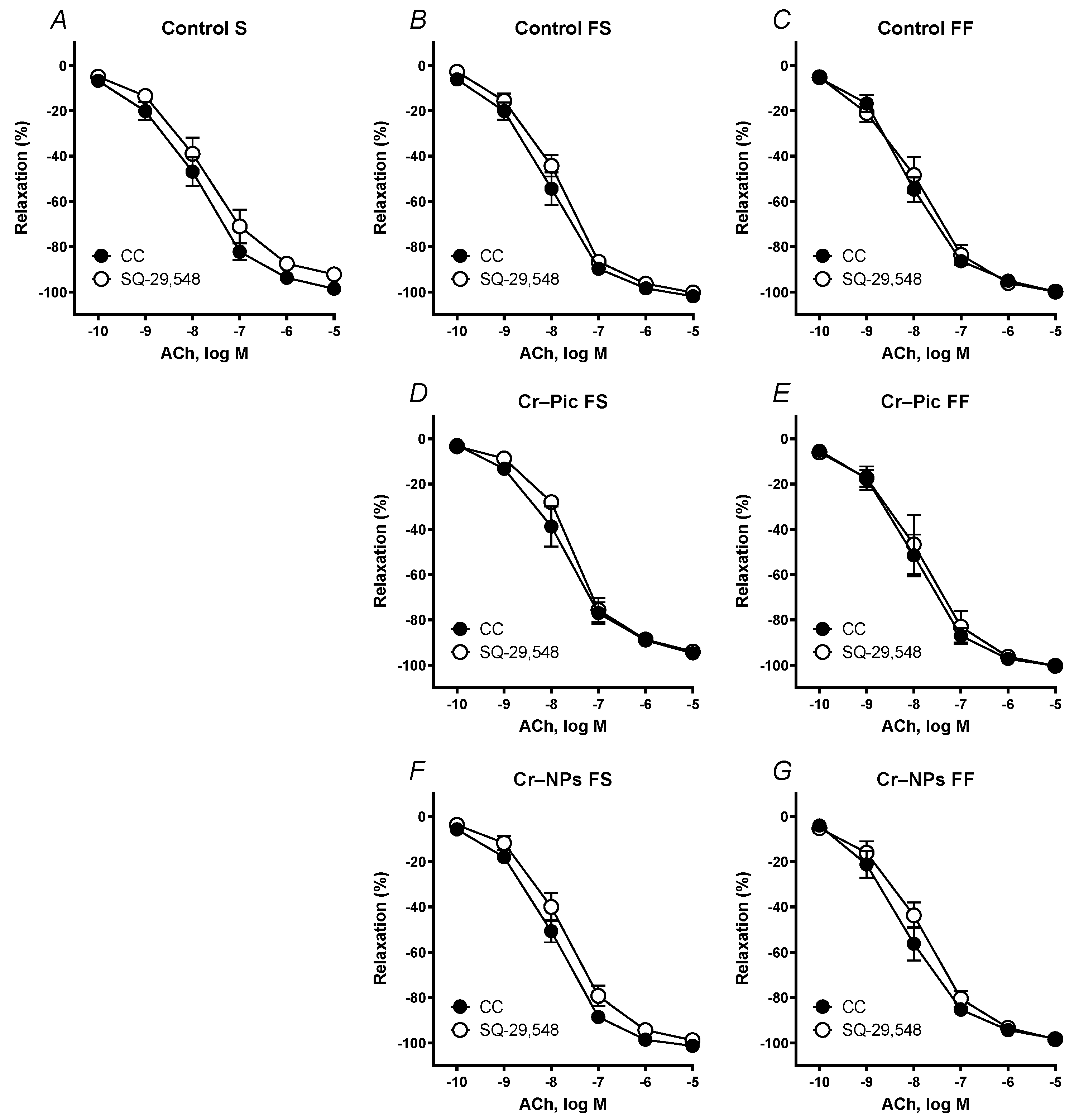
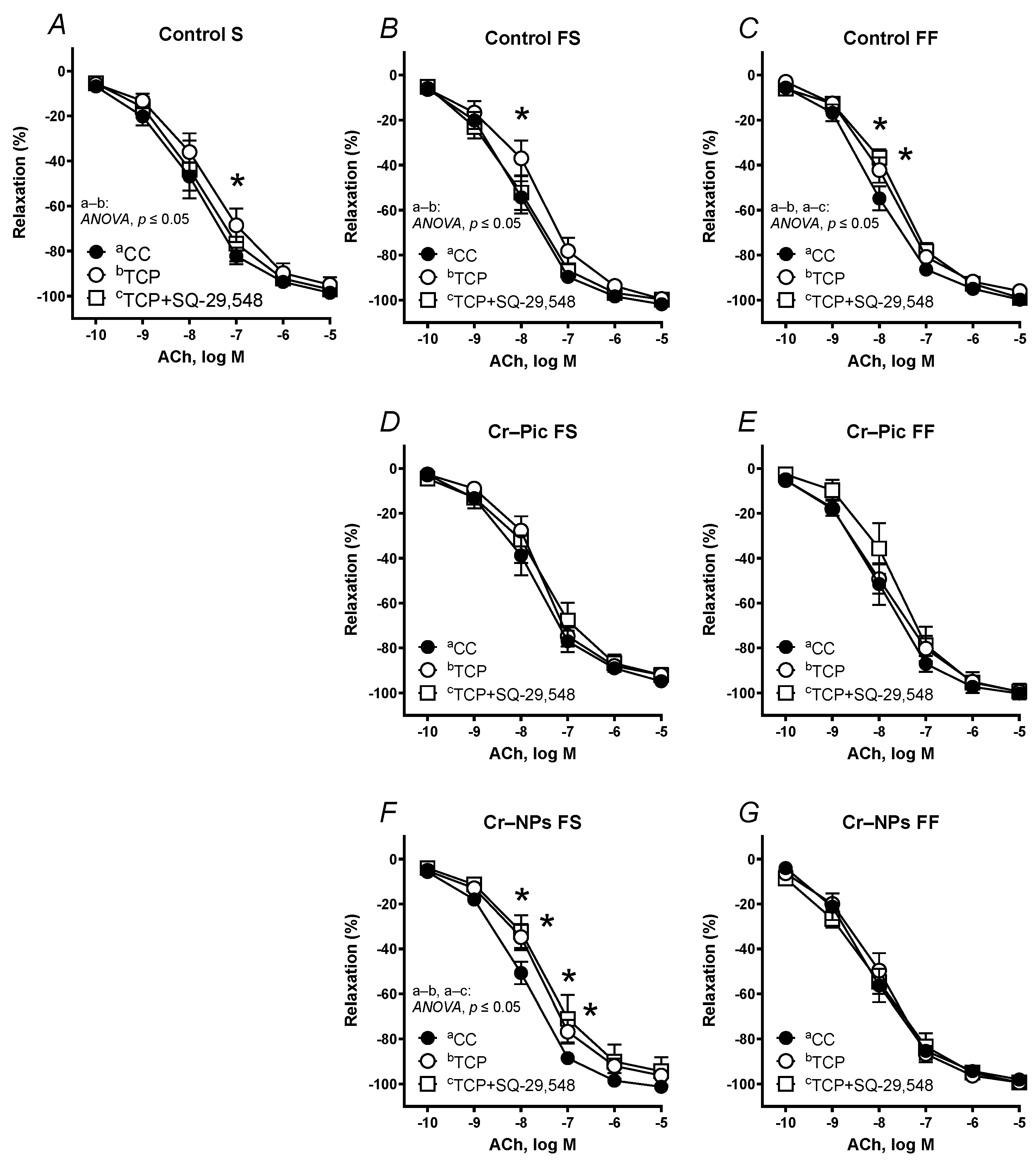
3.4.1. Vascular Reactivity Studies
3.4.2. TXA2 and PGI2 Production
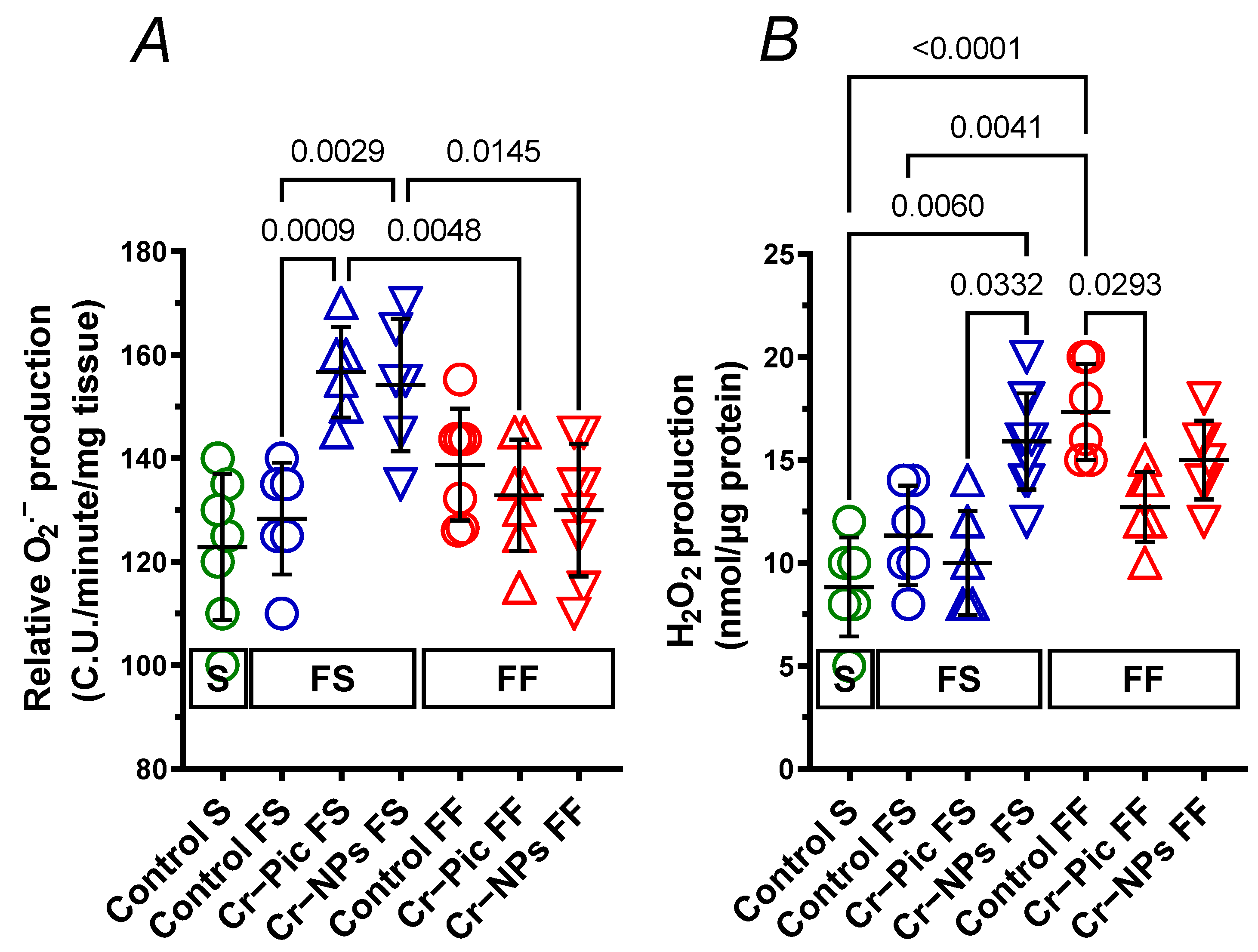
3.4.3. Detection of Superoxide Anion
3.4.4. Detection of Hydrogen Peroxide
4. Discussion
4.1. Vascular Effects of Experimental Feeding
4.2. Cardiac Effects of Experimental Feeding
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ognik, K.; Dworzański, W.; Sembratowicz, I.; Fotschki, B.; Cholewińska, E.; Listos, P.; Juśkiewicz, J. The effect of the a high-fat diet supplemented with various forms of chromium on rats’ body composition, liver metabolism and organ histology Cr in liver metabolism and histology of selected organs. J. Trace Elem. Med. Biol. 2021, 64, 126705. [Google Scholar] [CrossRef]
- Majewski, M.; Lis, B.; Juśkiewicz, J.; Ognik, K.; Jedrejek, D.; Stochmal, A.; Olas, B. The composition and vascular/antioxidant properties of Taraxacum officinale flower water syrup in a normal-fat diet using an obese rat model. J. Ethnopharmacol. 2021, 265, 113393. [Google Scholar] [CrossRef]
- Żary-Sikorska, E.; Fotschki, B.; Jurgoński, A.; Kosmala, M.; Milala, J.; Kołodziejczyk, K.; Majewski, M.; Ognik, K.; Juśkiewicz, J. Protective Effects of a Strawberry Ellagitannin-Rich Extract against Pro-Oxidative and Pro-Inflammatory Dysfunctions Induced by a high-fat Diet in a Rat Model. Molecules 2020, 25, 5874. [Google Scholar] [CrossRef] [PubMed]
- Żary-Sikorska, E.; Fotschki, B.; Kołodziejczyk, K.; Jurgoński, A.; Kosmala, M.; Milala, J.; Majewski, M.; Ognik, K.; Juśkiewicz, J. Strawberry phenolic extracts effectively mitigated metabolic disturbances associated with a high-fat ingestion in rats depending on the ellagitannin polymerization degree. Food Funct. 2021, 12, 5779–5792. [Google Scholar] [CrossRef]
- Dworzański, W.; Cholewińska, E.; Fotschki, B.; Juśkiewicz, J.; Listos, P.; Ognik, K. Assessment of DNA Methylation and Oxidative Changes in the Heart and Brain of Rats Receiving a high-fat Diet Supplemented with Various Forms of Chromium. Animals 2020, 10, 1470. [Google Scholar] [CrossRef]
- Majewski, M.; Jurgoński, A.; Fotschki, B.; Juśkiewicz, J. The toxic effects of monosodium glutamate (MSG) - The involvement of nitric oxide, prostanoids and potassium channels in the reactivity of thoracic arteries in MSG-obese rats. Toxicol. Appl. Pharmacol. 2018, 359, 62–69. [Google Scholar] [CrossRef]
- Liu, B.; Zhan, M.; Zhang, Y.; Li, H.; Wu, X.; Zhuang, F.; Luo, W.; Zhou, Y. Increased role of E prostanoid receptor-3 in prostacyclin-evoked contractile activity of spontaneously hypertensive rat mesenteric resistance arteries. Sci. Rep. 2017, 7, 8927. [Google Scholar] [CrossRef] [Green Version]
- Turck, D.; Bresson, J.L.; Burlingame, B.; Dean, T.; Fairweather-Tait, S.; Heinonen, M.; McArdle, H.J. EFSA NDA Panel (EFSA Panel on Dietetic Products, Nutrition and Allergies), Scientific Opinion on Dietary Reference Values for chromium. EFSA J. 2014, 12, 3845. [Google Scholar]
- Stępniowska, A.; Tutaj, K.; Juśkiewicz, J.; Ognik, K. Effect of a high-fat diet and chromium on hormones level and Cr retention in rats. J. Endocrinol. Investig. 2022, 45, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Dworzański, W.; Cholewińska, E.; Fotschki, B.; Juśkiewicz, J.; Ognik, K. Oxidative, epigenetic changes and fermentation processes in the intestine of rats fed a high-fat diets supplemented with various chromium forms. Sci. Rep. 2022, 12, 9817. [Google Scholar] [CrossRef] [PubMed]
- Dworzański, W.; Sembratowicz, I.; Cholewińska, E.; Tutaj, K.; Fotschki, B.; Juśkiewicz, J.; Ognik, K. Effects of Different Chromium Compounds on Hematology and Inflammatory Cytokines in Rats Fed A high-fat Diet. Front. Immunol. 2021, 12, 614000. [Google Scholar] [CrossRef] [PubMed]
- Majewski, M.; Kasica, N.; Jakimiuk, A.; Podlasz, P. Toxicity and cardiac effects of acute exposure to tryptophan metabolites on the kynurenine pathway in early developing zebrafish (Danio rerio) embryos. Toxicol. Appl. Pharmacol. 2018, 341, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Majewski, M.; Kozlowska, A.; Thoene, M.; Lepiarczyk, E.; Grzegorzewski, W.J. Overview of the role of vitamins and minerals on the kynurenine pathway in health and disease. J. Physiol. Pharmacol. 2016, 67, 3–19. [Google Scholar]
- Majewski, M.; Juśkiewicz, J.; Krajewska-Włodarczyk, M.; Gromadziński, L.; Socha, K.; Cholewińska, E.; Ognik, K. The Role of 20-HETE, COX, Thromboxane Receptors, and Blood Plasma Antioxidant Status in Vascular Relaxation of Copper-Nanoparticle-Fed WKY Rats. Nutrients 2021, 13, 3793. [Google Scholar] [CrossRef] [PubMed]
- Cholewińska, E.; Juśkiewicz, J.; Majewski, M.; Smagieł, R.; Listos, P.; Fotschki, B.; Godycka-Kłos, I.; Ognik, K. Effect of Copper Nanoparticles in the Diet of WKY and SHR Rats on the Redox Profile and Histology of the Heart, Liver, Kidney, and Small Intestine. Antioxidants 2022, 11, 910. [Google Scholar] [CrossRef]
- Majewski, M.; Lis, B.; Olas, B.; Ognik, K.; Juśkiewicz, J. Dietary supplementation with copper nanoparticles influences the markers of oxidative stress and modulates vasodilation of thoracic arteries in young Wistar rats. PLoS ONE 2020, 15, e0229282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majewski, M.; Ognik, K.; Juśkiewicz, J. Copper nanoparticles modify the blood plasma antioxidant status and modulate the vascular mechanisms with nitric oxide and prostanoids involved in Wistar rats. Pharmacol. Rep. 2019, 71, 509–516. [Google Scholar] [CrossRef]
- Majewski, M.; Ognik, K.; Juśkiewicz, J. Copper nanoparticles enhance vascular contraction induced by prostaglandin F2-alpha and decrease the blood plasma cu-zn ratio in wistar rats. J. Elem. 2019, 24, 911–922. [Google Scholar] [CrossRef]
- Majewski, M.; Ognik, K.; Zdunczyk, P.; Juskiewicz, J. Effect of dietary copper nanoparticles versus one copper (II) salt: Analysis of vasoreactivity in a rat model. Pharmacol. Rep. 2017, 69, 1282–1288. [Google Scholar] [CrossRef]
- Majewski, M.; Klett-Mingo, M.; Verdasco-Martín, C.M.; Otero, C.; Ferrer, M. Spirulina extract improves age-induced vascular dysfunction. Pharm. Biol. 2022, 60, 627–637. [Google Scholar] [CrossRef]
- Majewski, M.; Jurgoński, A. The Effect of Hemp (Cannabis sativa L.) Seeds and Hemp Seed Oil on Vascular Dysfunction in Obese Male Zucker Rats. Nutrients 2021, 3, 2575. [Google Scholar] [CrossRef] [PubMed]
- Abebe, W.; Liu, J.Y.; Wimborne, H.; Mozaffari, M.S. Effects of chromium picolinate on vascular reactivity and cardiac ischemia-reperfusion injury in spontaneously hypertensive rats. Pharmacol. Rep. 2010, 62, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Kopilas, M.A.; Dang, L.N.; Anderson, H.D. Effect of dietary chromium on resistance artery function and nitric oxide signaling in the sucrose-fed spontaneously hypertensive rat. J. Vasc. Res. 2007, 44, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Gluais, P.; Lonchampt, M.; Morrow, J.D.; Vanhoutte, P.M.; Feletou, M. Acetylcholine-induced endothelium-dependent contractions in the SHR aorta: The Janus face of prostacyclin. Br. J. Pharmacol. 2005, 146, 834–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martorell, A.; Blanco-Rivero, J.; Aras-López, R.; Sagredo, A.; Balfagón, G.; Ferrer, M. Orchidectomy increases the formation of prostanoids and modulates their role in the acetylcholine-induced relaxation in the rat aorta. Cardiovasc. Res. 2008, 77, 590–599. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Rivero, J.; Aller, M.A.; Arias, J.; Ferrer, M.; Balfagón, G. Long-term portal hypertension increases the vasodilator response to acetylcholine in rat aorta: Role of prostaglandin I2. Clin. Sci. 2009, 117, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Noeman, S.A.; Hamooda, H.E.; Baalash, A.A. Biochemical study of oxidative stress markers in the liver, kidney and heart of high fat diet induced obesity in rats. Diabetol. Metab. Syndr. 2011, 3, 17. [Google Scholar] [CrossRef] [Green Version]
- Llévenes, P.; Rodrigues-Díez, R.; Cros-Brunsó, L.; Prieto, M.I.; Casaní, L.; Balfagón, G.; Blanco-Rivero, J. Beneficial Effect of a Multistrain Synbiotic Prodefen® Plus on the Systemic and Vascular Alterations Associated with Metabolic Syndrome in Rats: The Role of the Neuronal Nitric Oxide Synthase and Protein Kinase A. Nutrients 2020, 12, 117. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.Y.; Chen, C.J.; Liu, C.H.; Mao, F.C. Chromium attenuates a high-fat diet-induced nonalcoholic fatty liver disease in KK/HlJ mice. Biochem. Biophys. Res. Commun. 2010, 397, 459–464. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
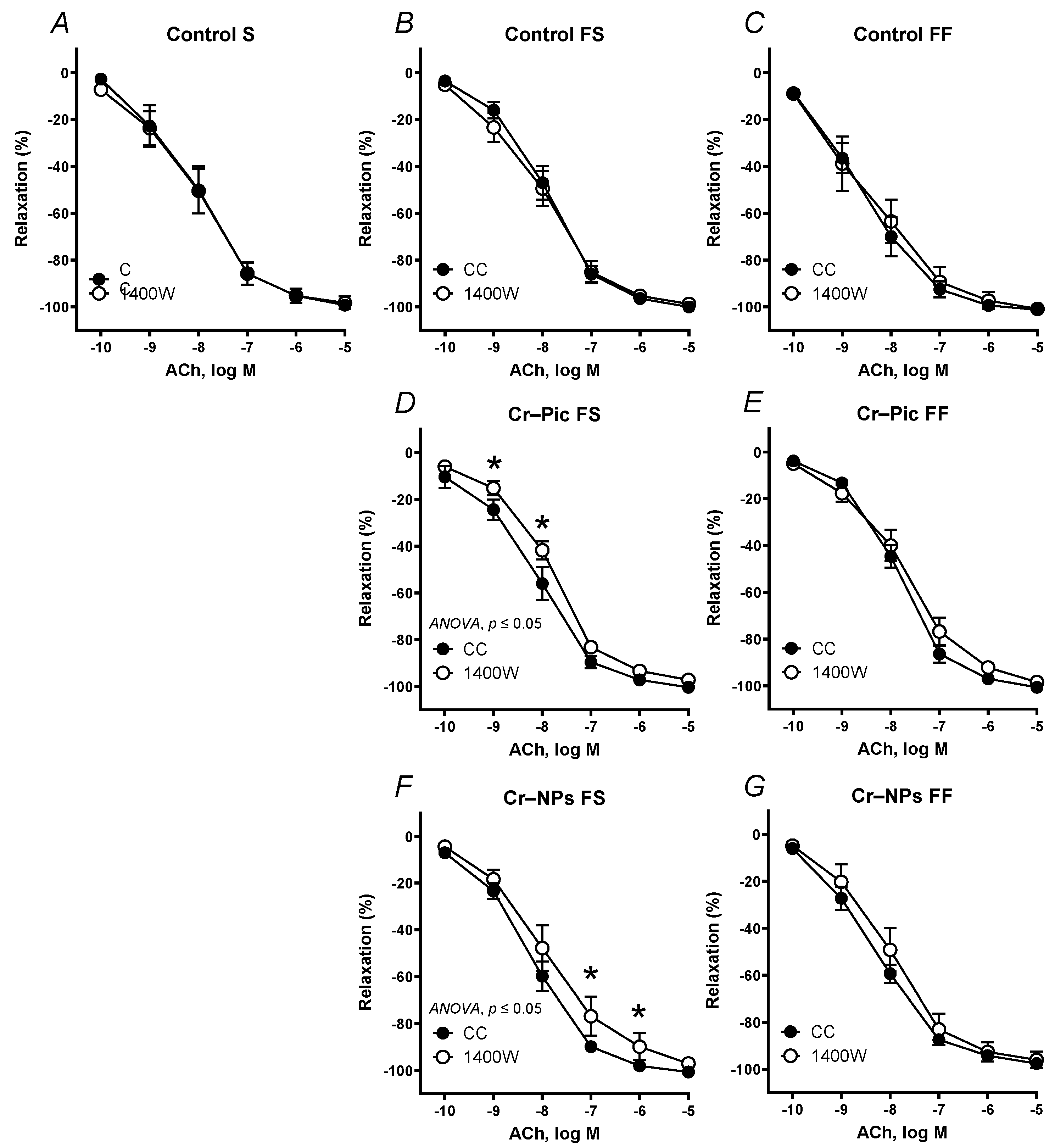{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient/Group | S | F |
|---|---|---|
| Casein | 14.8 | 14.8 |
| DL-methionine | 0.2 | 0.2 |
| Cellulose | 8.0 | 3.0 |
| Choline chloride | 0.2 | 0.2 |
| Cholesterol | 0.3 | 0.3 |
| Vitamin mix | 1.0 | 1.0 |
| Mineral mix | 3.5 | 3.5 |
| Maize starch | 64 | 52 |
| Rapeseed oil | 8.0 | 8.0 |
| Lard | - | 17.0 |
| Control Conditions * | 1400 W | Indomethacin | NS-398 | SQ-29,548 | Tranylcypromine | TCP plus SQ-29,548 | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AUC | Emax (%) | pEC50 | AUC | Emax (%) | pEC50 | AUC | Emax (%) | pEC50 | AUC | Emax (%) | pEC50 | AUC | Emax (%) | pEC50 | AUC | Emax (%) | pEC50 | AUC | Emax (%) | pEC50 | ||
| S | Control | 305.3 | 96.16 | 7.967 | 308.0 | 96.38 | 7.950 | 287.2 | 92.37 | 7.921 | 254.9 * | 90.84 | 7.675 | 259.6 | 88.87 | 7.743 | 257.9 * | −91.97 | 7.566 * | 280.0 | 93.47 | 7.815 |
| FS | Control | 310.4 | 98.78 | 8.002 | 305.2 | 96.47 | 7.943 | 278.0 * | 95.75 | 7.816 | 290.9 | 96.26 | 7.939 | 294.5 | 98.61 | 7.866 | 278.3 * | −97.01 | 7.632 * | 311.3 | 97.54 | 7.996 |
| Cr–Pic | 300.4 | 95.60 | 7.940 | 285.2 * | 95.81 | 7.805 | 236.5 * | 86.90 * | 7.530 * | 223.0 * | 87.51 * | 7.384 * | 249.7 | 92.89 | 7.589 | 246.7 | −91.48 | 7.597 | 247.5 | 89.71 | 7.536 | |
| Cr–NPs | 315.6 | 98.77 | 8.032 | 283.5 * | 91.61 | 7.938 | 254.9 * | 92.52 * | 7.616 * | 228.2 * | 94.09 | 7.486 * | 276.5 | 96.21 | 7.759 | 267.3 * | −94.70 * | 7.641 * | 253.8 * | 92.49 * | 7.571 * | |
| FF | Control | 331.5 | 97.63 | 8.238 | 343.9 | 96.77 | 8.285 | 298.6 * | 95.93 | 7.867 * | 261.2 * | 94.13 | 7.562 * | 301.5 | 97.05 | 7.904 | 276.9 | −93.83 | 7.855 | 273.0 * | 96.29 | 7.661 * |
| Cr–Pic | 302.3 | 98.81 | 7.943 | 278.3 | 94.88 | 7.710 | 233.9 * | 86.18 * | 7.605 * | 203.1 * | 83.53 * | 7.471 * | 296.5 | 97.64 | 7.858 | 294.9 | −95.57 | 7.924 | 270.3 | 97.58 | 7.676 | |
| Cr–NPs | 311.7 | 95.31 | 8.124 | 295.4 | 93.71 | 7.977 | 288.2 * | 95.10 | 7.805 * | 232.4 * | 86.34 | 7.503 * | 285.3 | 95.27 | 7.826 | 305.5 | −97.65 | 7.937 | 313.7 | 95.52 | 8.017 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majewski, M.; Gromadziński, L.; Cholewińska, E.; Ognik, K.; Fotschki, B.; Juśkiewicz, J. Dietary Effects of Chromium Picolinate and Chromium Nanoparticles in Wistar Rats Fed with a High-Fat, Low-Fiber Diet: The Role of Fat Normalization. Nutrients 2022, 14, 5138. https://doi.org/10.3390/nu14235138
Majewski M, Gromadziński L, Cholewińska E, Ognik K, Fotschki B, Juśkiewicz J. Dietary Effects of Chromium Picolinate and Chromium Nanoparticles in Wistar Rats Fed with a High-Fat, Low-Fiber Diet: The Role of Fat Normalization. Nutrients. 2022; 14(23):5138. https://doi.org/10.3390/nu14235138
Chicago/Turabian StyleMajewski, Michał, Leszek Gromadziński, Ewelina Cholewińska, Katarzyna Ognik, Bartosz Fotschki, and Jerzy Juśkiewicz. 2022. "Dietary Effects of Chromium Picolinate and Chromium Nanoparticles in Wistar Rats Fed with a High-Fat, Low-Fiber Diet: The Role of Fat Normalization" Nutrients 14, no. 23: 5138. https://doi.org/10.3390/nu14235138
APA StyleMajewski, M., Gromadziński, L., Cholewińska, E., Ognik, K., Fotschki, B., & Juśkiewicz, J. (2022). Dietary Effects of Chromium Picolinate and Chromium Nanoparticles in Wistar Rats Fed with a High-Fat, Low-Fiber Diet: The Role of Fat Normalization. Nutrients, 14(23), 5138. https://doi.org/10.3390/nu14235138