
Combination Treatment of Withalongolide a Triacetate with Cisplatin Induces Apoptosis by Targeting Translational Initiation, Migration, and Epithelial to Mesenchymal Transition in Head and Neck Squamous Cell Carcinoma

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Viability Assay
2.2. Western Blot Analysis
2.3. Boyden Chamber Assay for Migration and Invasion
2.4. Statistical Analysis
3. Results
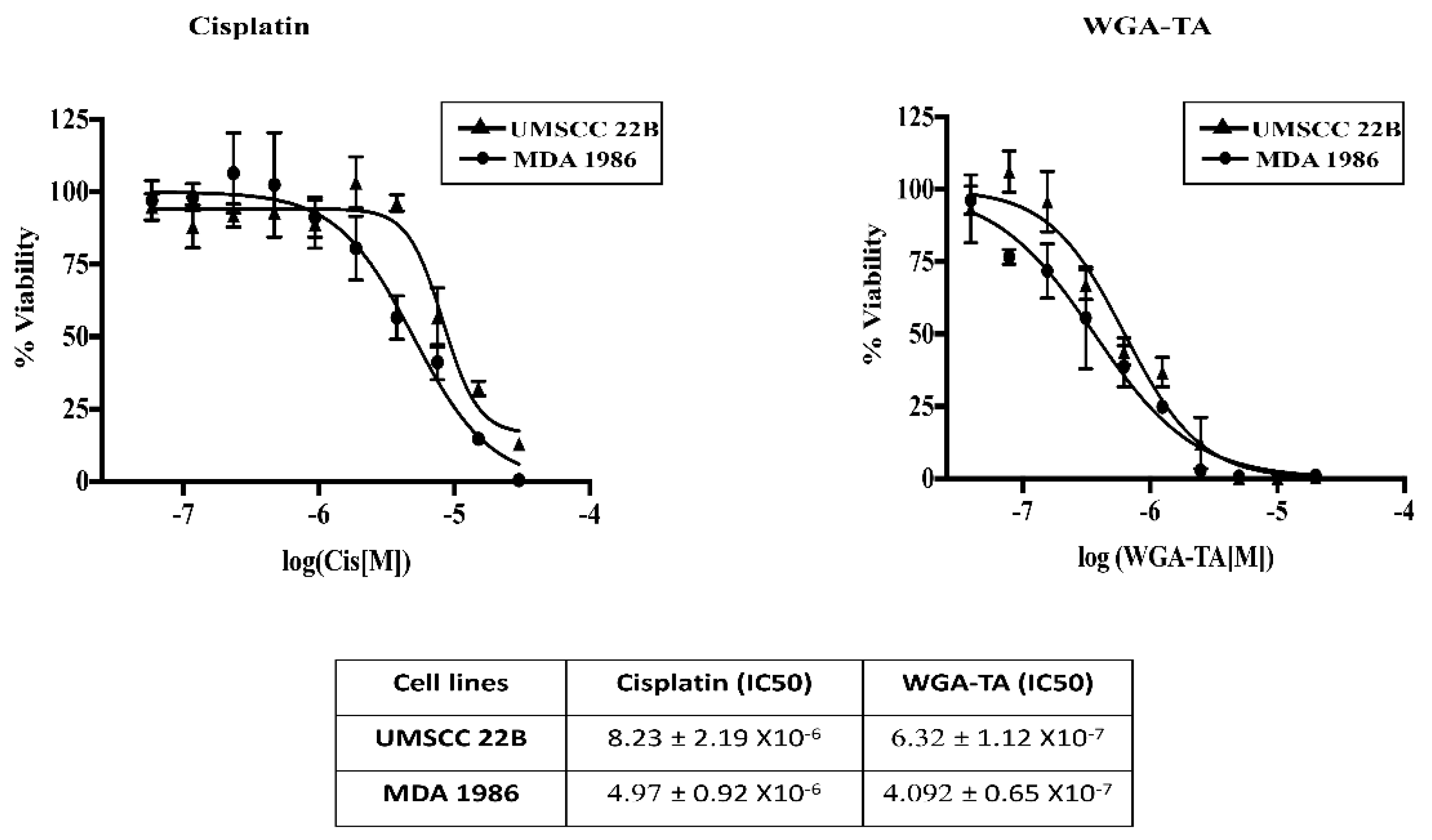
3.1. Determination of IC50 Values for Single Drug Treatment
3.2. Synergistic Effect of Combination Treatment
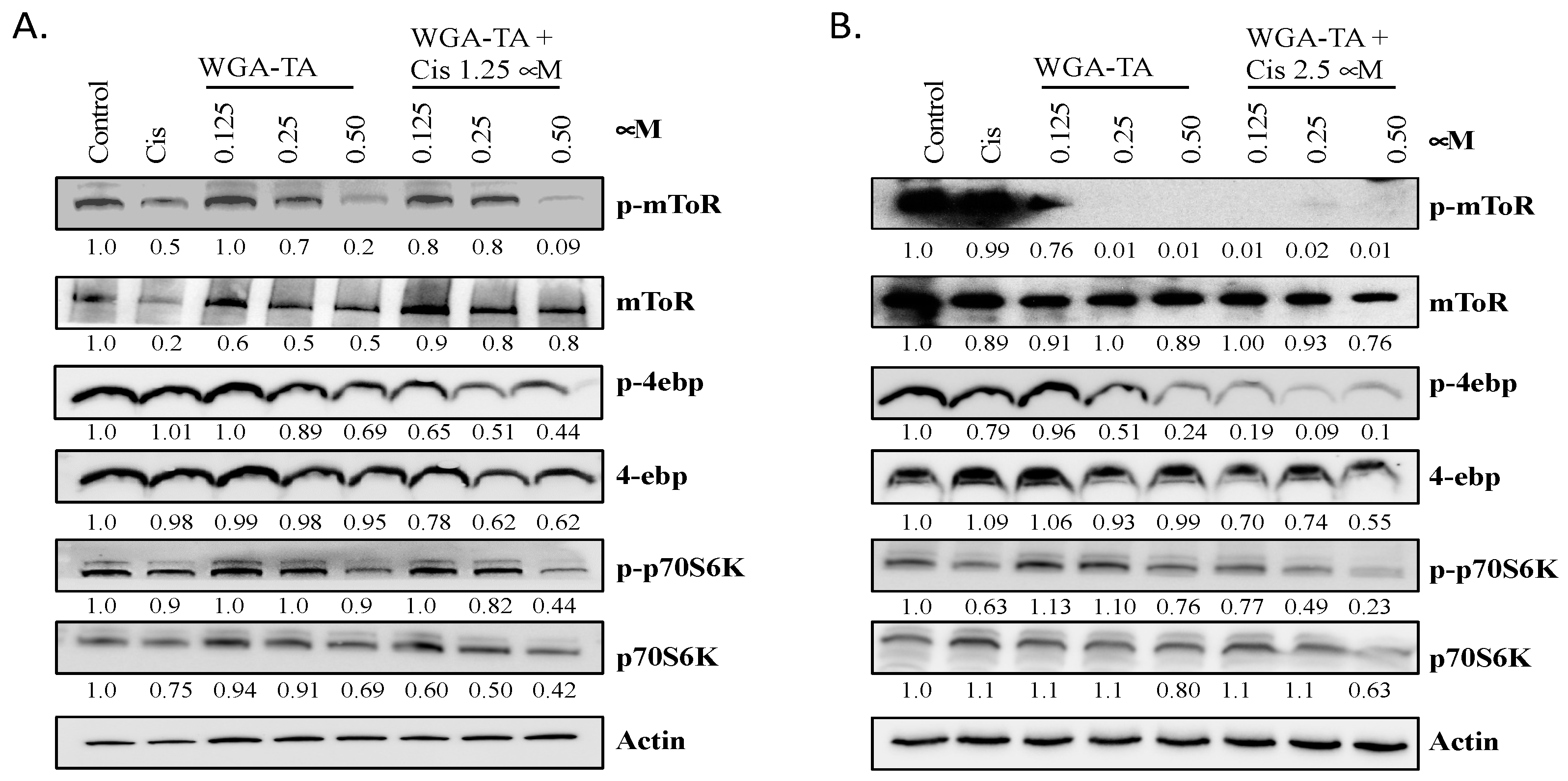
3.3. Effect of WGA-TA and Combination Treatment on the mTOR Pathway and Its Substrates
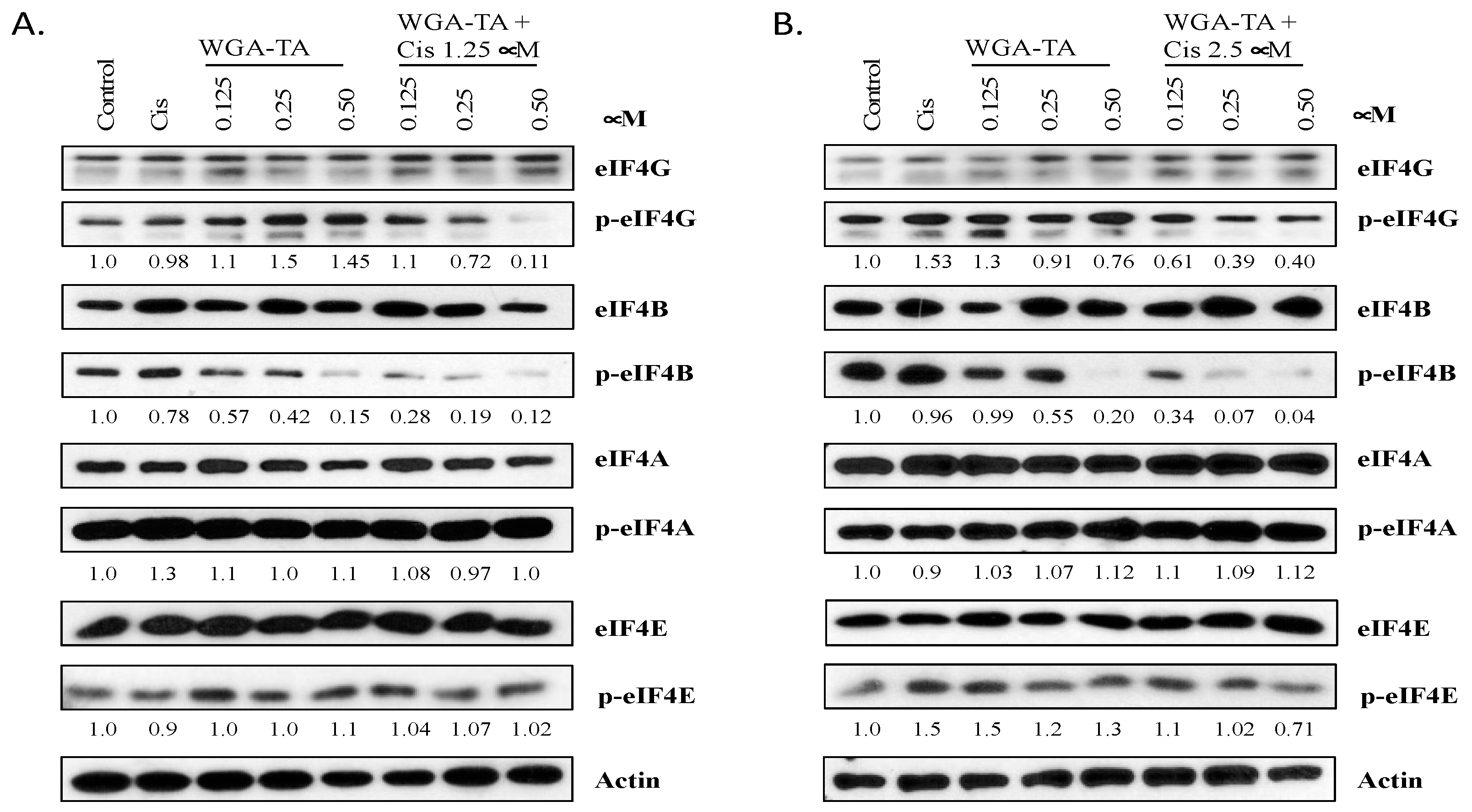
3.4. Effect of WGA-TA and Combination Treatment on Translation Initiation
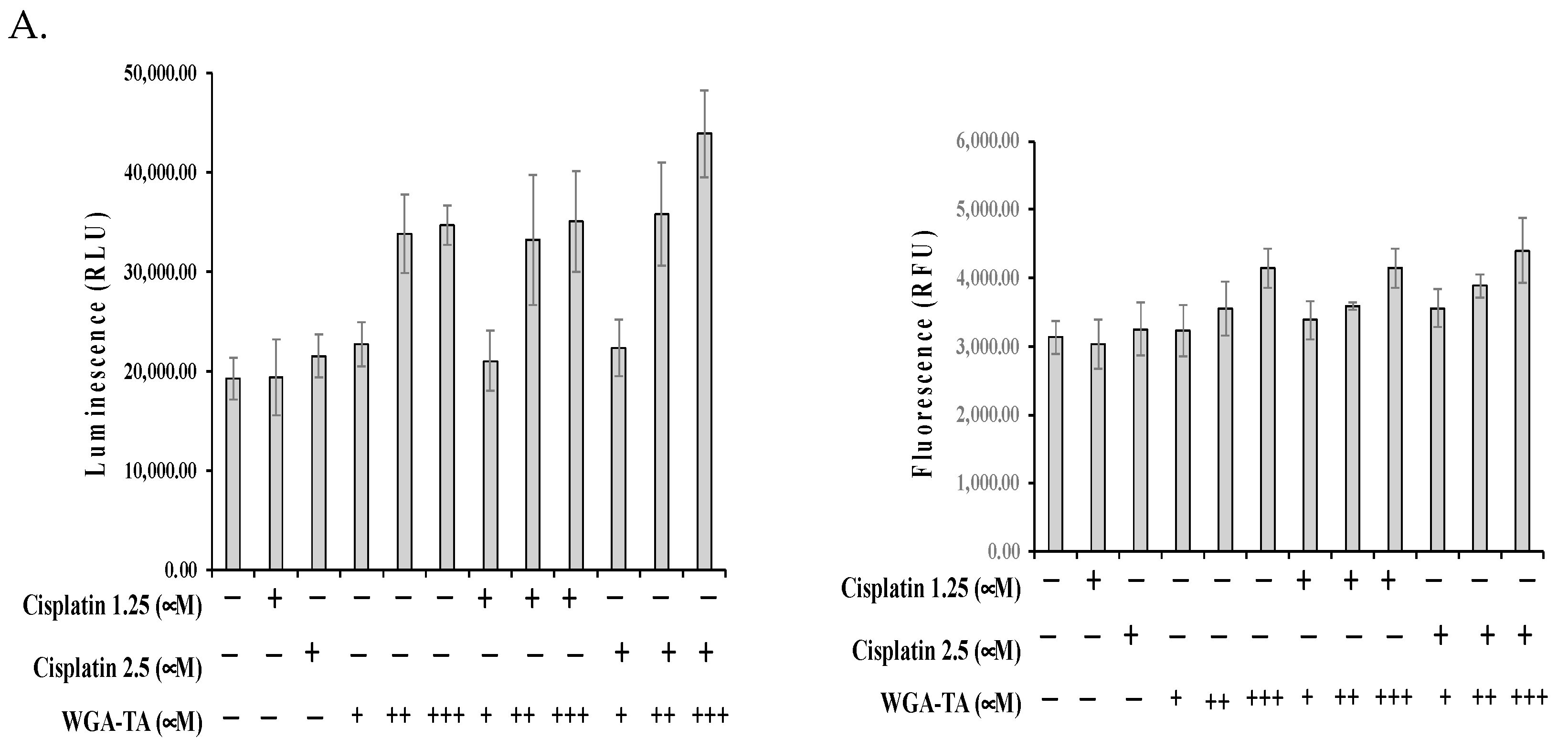
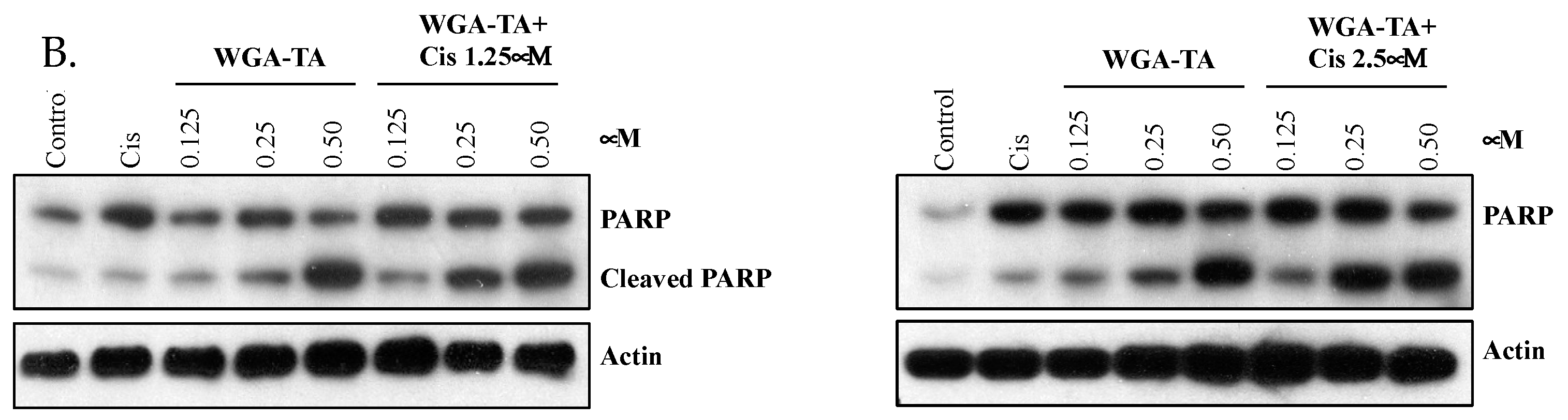
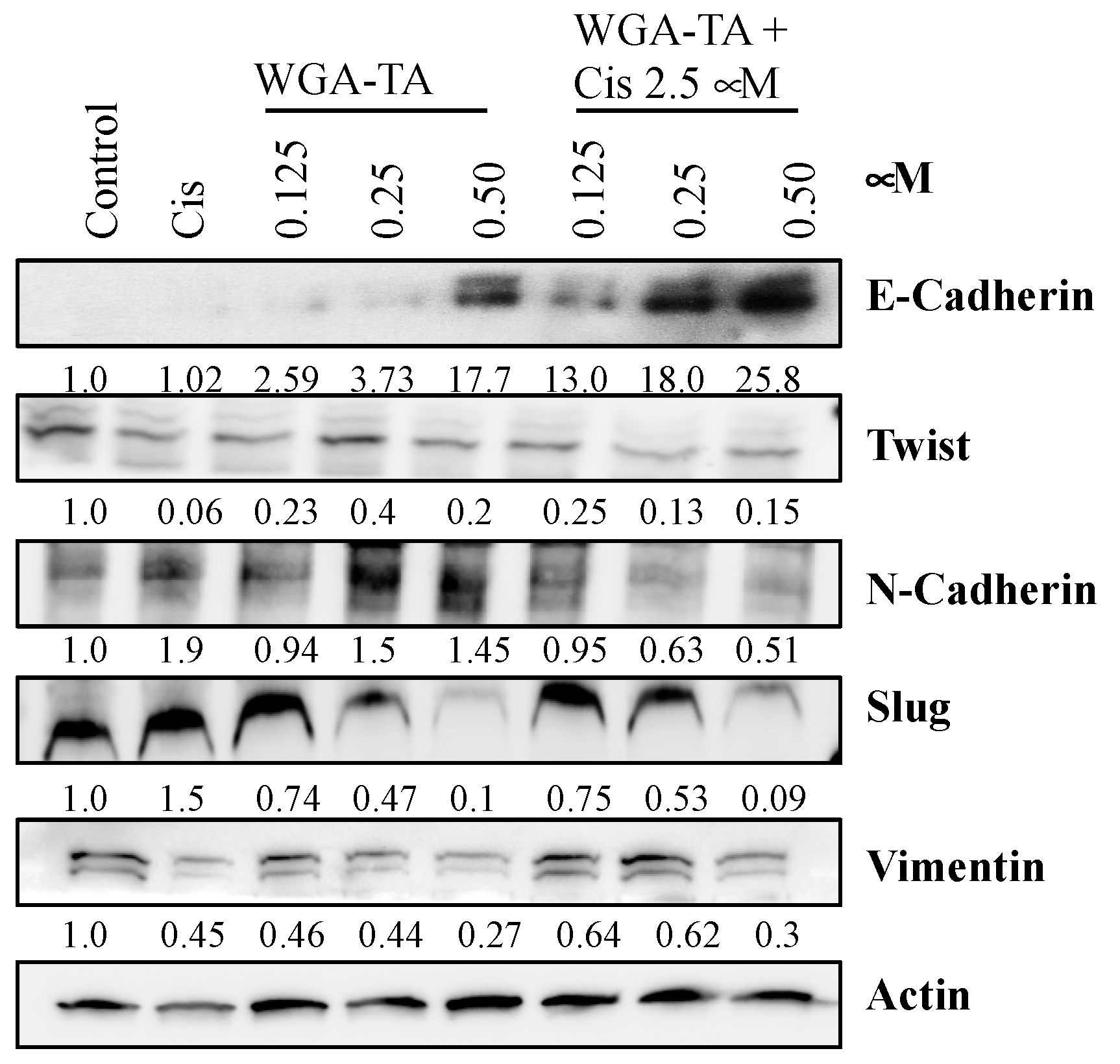
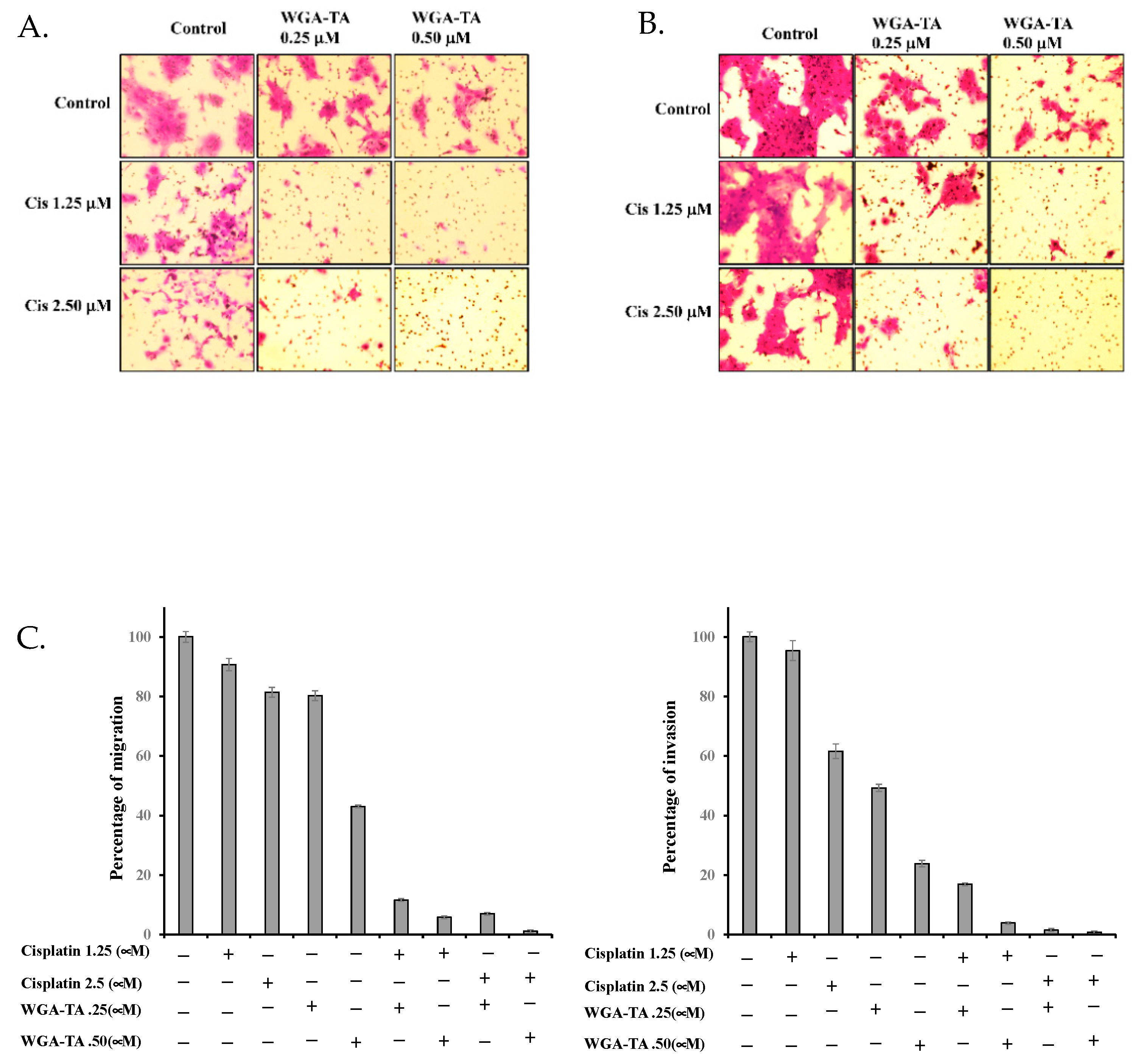
3.5. Effect of WGA-TA and Combination Treatment on the EMT, Apoptosis, Migration and Invasion
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Ang, K.K.; Harris, J.; Wheeler, R.; Weber, R.; Rosenthal, D.I.; Nguyen-Tan, P.F.; Westra, W.H.; Chung, C.H.; Jordan, R.C.; Lu, C.; et al. Human papillomavirus and survival of patients with oropharyngeal cancer. N. Engl. J. Med. 2010, 363, 24–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, K.D.; Siegel, R.L.; Lin, C.C.; Mariotto, A.B.; Kramer, J.L.; Rowland, J.H.; Stein, K.D.; Alteri, R.; Jemal, A. Cancer treatment and survivorship statistics, 2016. CA Cancer J. Clin. 2016, 66, 271–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsel, S.M.; Grandis, J.R.; Thomas, S.M. Nucleic acid targeting: Towards personalized therapy for head and neck cancer. Oncogene 2016, 35, 3217–3226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelland, L. The resurgence of platinum-based cancer chemotherapy. Nat. Rev. Cancer 2007, 7, 573–584. [Google Scholar] [CrossRef]
- Sacco, A.G.; Cohen, E.E. Current Treatment Options for Recurrent or Metastatic Head and Neck Squamous Cell Carcinoma. J. Clin. Oncol. 2015, 33, 3305–3313. [Google Scholar] [CrossRef]
- Forastiere, A.A. Chemotherapy in the treatment of locally advanced head and neck cancer. J. Surg. Oncol. 2008, 97, 701–707. [Google Scholar] [CrossRef]
- Jhaveri, K.; Ochiana, S.O.; Dunphy, M.P.; Gerecitano, J.F.; Corben, A.D.; Peter, R.I.; Janjigian, Y.Y.; Gomes-DaGama, E.M.; Koren, J., 3rd; Modi, S.; et al. Heat shock protein 90 inhibitors in the treatment of cancer: Current status and future directions. Expert Opin. Investig. Drugs 2014, 23, 611–628. [Google Scholar] [CrossRef] [Green Version]
- Mahalingam, D.; Swords, R.; Carew, J.S.; Nawrocki, S.T.; Bhalla, K.; Giles, F.J. Targeting HSP90 for cancer therapy. Br. J. Cancer 2009, 100, 1523–1529. [Google Scholar] [CrossRef] [Green Version]
- Sidera, K.; Patsavoudi, E. HSP90 inhibitors: Current development and potential in cancer therapy. Recent Pattents Anticancer Drug Discov. 2014, 9, 1–20. [Google Scholar] [CrossRef]
- Cohen, S.M.; Mukerji, R.; Samadi, A.K.; Zhang, X.; Zhao, H.; Blagg, B.S.; Cohen, M.S. Novel C-terminal Hsp90 inhibitor for head and neck squamous cell cancer (HNSCC) with in vivo efficacy and improved toxicity profiles compared with standard agents. Ann. Surg. Oncol. 2012, 19 (Suppl. 3), S483–S490. [Google Scholar] [CrossRef]
- Donnelly, A.; Blagg, B.S. Novobiocin and additional inhibitors of the Hsp90 C-terminal nucleotide-binding pocket. Curr. Med. Chem. 2008, 15, 2702–2717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birkeland, A.C.; Brenner, J.C. Personalizing Medicine in Head and Neck Squamous Cell Carcinoma: The Rationale for Combination Therapies. Med. Res. Arch. 2015, 3. [Google Scholar] [CrossRef]
- White, P.T.; Subramanian, C.; Motiwala, H.F.; Cohen, M.S. Natural Withanolides in the Treatment of Chronic Diseases. Adv. Exp. Med. Biol. 2016, 928, 329–373. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Motiwala, H.; Samadi, A.; Day, V.; Aube, J.; Cohen, M.; Kindscher, K.; Gollapudi, R.; Timmermann, B. Minor withanolides of Physalis longifolia: Structure and cytotoxicity. Chem. Pharm. Bull. 2012, 60, 1234–1239. [Google Scholar] [CrossRef] [Green Version]
- Samadi, A.K.; Tong, X.; Mukerji, R.; Zhang, H.; Timmermann, B.N.; Cohen, M.S. Withaferin A, a cytotoxic steroid from Vassobia breviflora, induces apoptosis in human head and neck squamous cell carcinoma. J. Nat. Prod. 2010, 73, 1476–1481. [Google Scholar] [CrossRef] [Green Version]
- Livingstone, M.; Sikstrom, K.; Robert, P.A.; Uze, G.; Larsson, O.; Pellegrini, S. Assessment of mTOR-Dependent Translational Regulation of Interferon Stimulated Genes. PLoS ONE 2015, 10, e0133482. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.C.; Talalay, P. Quantitative analysis of dose-effect relationships: The combined effects of multiple drugs or enzyme inhibitors. Adv. Enzyme Regul. 1984, 22, 27–55. [Google Scholar] [CrossRef]
- Pearl, L.H.; Prodromou, C.; Workman, P. The Hsp90 molecular chaperone: An open and shut case for treatment. Biochem. J. 2008, 410, 439–453. [Google Scholar] [CrossRef] [Green Version]
- Grzmil, M.; Hemmings, B.A. Translation regulation as a therapeutic target in cancer. Cancer Res. 2012, 72, 3891–3900. [Google Scholar] [CrossRef] [Green Version]
- Furic, L.; Rong, L.; Larsson, O.; Koumakpayi, I.H.; Yoshida, K.; Brueschke, A.; Petroulakis, E.; Robichaud, N.; Pollak, M.; Gaboury, L.A.; et al. eIF4E phosphorylation promotes tumorigenesis and is associated with prostate cancer progression. Proc. Natl. Acad. Sci. USA 2010, 107, 14134–14139. [Google Scholar] [CrossRef]
- Liu, T.; Li, R.; Zhao, H.; Deng, J.; Long, Y.; Shuai, M.T.; Li, Q.; Gu, H.; Chen, Y.Q.; Leng, A.M. eIF4E promotes tumorigenesis and modulates chemosensitivity to cisplatin in esophageal squamous cell carcinoma. Oncotarget 2016, 7, 66851–66864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topisirovic, I.; Guzman, M.L.; McConnell, M.J.; Licht, J.D.; Culjkovic, B.; Neering, S.J.; Jordan, C.T.; Borden, K.L. Aberrant eukaryotic translation initiation factor 4E-dependent mRNA transport impedes hematopoietic differentiation and contributes to leukemogenesis. Mol. Cell Biol. 2003, 23, 8992–9002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culjkovic-Kraljacic, B.; Baguet, A.; Volpon, L.; Amri, A.; Borden, K.L. The oncogene eIF4E reprograms the nuclear pore complex to promote mRNA export and oncogenic transformation. Cell Rep. 2012, 2, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Comtesse, N.; Keller, A.; Diesinger, I.; Bauer, C.; Kayser, K.; Huwer, H.; Lenhof, H.P.; Meese, E. Frequent overexpression of the genes FXR1, CLAPM1 and EIF4G located on amplicon 3q26-27 in squamous cell carcinoma of the lung. Int. J. Cancer 2007, 120, 2538–2544. [Google Scholar] [CrossRef]
- Silvera, D.; Arju, R.; Darvishian, F.; Levine, P.H.; Zolfaghari, L.; Goldberg, J.; Hochman, T.; Formenti, S.C.; Schneider, R.J. Essential role for eIF4GI overexpression in the pathogenesis of inflammatory breast cancer. Nat. Cell Biol. 2009, 11, 903–908. [Google Scholar] [CrossRef]
- El-Naggar, A.M.; Sorensen, P.H. Translational control of aberrant stress responses as a hallmark of cancer. J. Pathol. 2018, 244, 650–666. [Google Scholar] [CrossRef] [Green Version]
- Truitt, M.L.; Ruggero, D. New frontiers in translational control of the cancer genome. Nat. Rev. Cancer 2016, 16, 288–304. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Wang, J.; Chen, K.; Guo, G.; Xi, R.; Rothman, P.B.; Whitten, D.; Zhang, L.; Huang, S.; Chen, J.L. eIF4B phosphorylation by pim kinases plays a critical role in cellular transformation by Abl oncogenes. Cancer Res. 2013, 73, 4898–4908. [Google Scholar] [CrossRef] [Green Version]
- Dauphin, M.; Barbe, C.; Lemaire, S.; Nawrocki-Raby, B.; Lagonotte, E.; Delepine, G.; Birembaut, P.; Gilles, C.; Polette, M. Vimentin expression predicts the occurrence of metastases in non small cell lung carcinomas. Lung Cancer 2013, 81, 117–122. [Google Scholar] [CrossRef]
- Oliver, F.J.; de la Rubia, G.; Rolli, V.; Ruiz-Ruiz, M.C.; de Murcia, G.; Murcia, J.M. Importance of poly(ADP-ribose) polymerase and its cleavage in apoptosis. Lesson from an uncleavable mutant. J. Biol. Chem. 1998, 273, 33533–33539. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | ||||
| Cisplatin Dose (μM) | WGA-TA Dose (μM) | Combination Effect | CI Value | Cell Lines |
| 1.25 | 2.50 | 0.017 | 0.04 | UMSCC-22B |
| 1.25 | 1.25 | 0.078 | 0.07 | |
| 1.25 | 0.63 | 0.178 | 0.07 | |
| 1.25 | 0.31 | 0.473 | 0.12 | |
| 1.25 | 0.16 | 0.864 | 0.31 | |
| 1.25 | 0.08 | 0.972 | 0.69 | |
| 2.50 | 2.50 | 0.017 | 0.04 | |
| 2.50 | 1.25 | 0.064 | 0.06 | |
| 2.50 | 0.63 | 0.146 | 0.06 | |
| 2.50 | 0.31 | 0.490 | 0.13 | |
| 2.50 | 0.16 | 0.795 | 0.21 | |
| 1.25 | 2.50 | 0.020 | 0.42 | MDA-1986 |
| 1.25 | 1.25 | 0.130 | 0.72 | |
| 1.25 | 0.63 | 0.140 | 0.39 | |
| 1.25 | 0.31 | 0.410 | 0.44 | |
| 1.25 | 0.16 | 0.760 | 0.54 | |
| 1.25 | 0.08 | 0.920 | 0.66 | |
| 2.50 | 2.50 | 0.010 | 0.30 | |
| 2.50 | 1.25 | 0.080 | 0.55 | |
| 2.50 | 0.63 | 0.140 | 0.38 | |
| 2.50 | 0.31 | 0.600 | 0.36 | |
| 2.50 | 0.16 | 0.740 | 0.28 | |
| B | ||||
| Cisplatin Dose (μM) | WGA-TA Dose (μM) | Combination Effect | CI Value | Cell Lines |
| 1.25 | 5.000 | 0.016 | 0.002 | |
| 1.25 | 2.500 | 0.212 | 0.010 | UMSCC-22B |
| 2.50 | 5.000 | 0.021 | 0.003 | |
| 2.50 | 2.500 | 0.239 | 0.012 | |
| 2.50 | 1.250 | 0.479 | 0.014 | |
| 1.25 | 5.000 | 0.008 | 0.556 | |
| 1.25 | 0.625 | 0.520 | 0.710 | MDA-1986 |
| 2.50 | 5.000 | 0.003 | 0.375 | |
| 2.50 | 1.250 | 0.310 | 0.953 | |
| 2.50 | 0.313 | 0.710 | 0.523 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subramanian, C.; Spielbauer, K.K.; Pearce, R.; Kovatch, K.J.; Prince, M.E.; Timmermann, B.N.; Cohen, M.S. Combination Treatment of Withalongolide a Triacetate with Cisplatin Induces Apoptosis by Targeting Translational Initiation, Migration, and Epithelial to Mesenchymal Transition in Head and Neck Squamous Cell Carcinoma. Nutrients 2022, 14, 5398. https://doi.org/10.3390/nu14245398
Subramanian C, Spielbauer KK, Pearce R, Kovatch KJ, Prince ME, Timmermann BN, Cohen MS. Combination Treatment of Withalongolide a Triacetate with Cisplatin Induces Apoptosis by Targeting Translational Initiation, Migration, and Epithelial to Mesenchymal Transition in Head and Neck Squamous Cell Carcinoma. Nutrients. 2022; 14(24):5398. https://doi.org/10.3390/nu14245398
Chicago/Turabian StyleSubramanian, Chitra, Katie K. Spielbauer, Robin Pearce, Kevin J. Kovatch, Mark E. Prince, Barbara N. Timmermann, and Mark S. Cohen. 2022. "Combination Treatment of Withalongolide a Triacetate with Cisplatin Induces Apoptosis by Targeting Translational Initiation, Migration, and Epithelial to Mesenchymal Transition in Head and Neck Squamous Cell Carcinoma" Nutrients 14, no. 24: 5398. https://doi.org/10.3390/nu14245398
APA StyleSubramanian, C., Spielbauer, K. K., Pearce, R., Kovatch, K. J., Prince, M. E., Timmermann, B. N., & Cohen, M. S. (2022). Combination Treatment of Withalongolide a Triacetate with Cisplatin Induces Apoptosis by Targeting Translational Initiation, Migration, and Epithelial to Mesenchymal Transition in Head and Neck Squamous Cell Carcinoma. Nutrients, 14(24), 5398. https://doi.org/10.3390/nu14245398