
Urocortin Neuropeptide Levels Are Impaired in the PBMCs of Overweight Children

 , ,
, ,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population Characteristics and Anthropometric Measurements
2.2. Blood Sample Collection and Analysis
2.3. Cell Culture and Treatment
2.4. RNA Extraction and Gene Expression by Real-Time Quantitative PCR
2.5. Protein Extraction and Analysis by Western Blotting
2.6. Statistical Analysis
3. Results
3.1. Study Population Characteristics
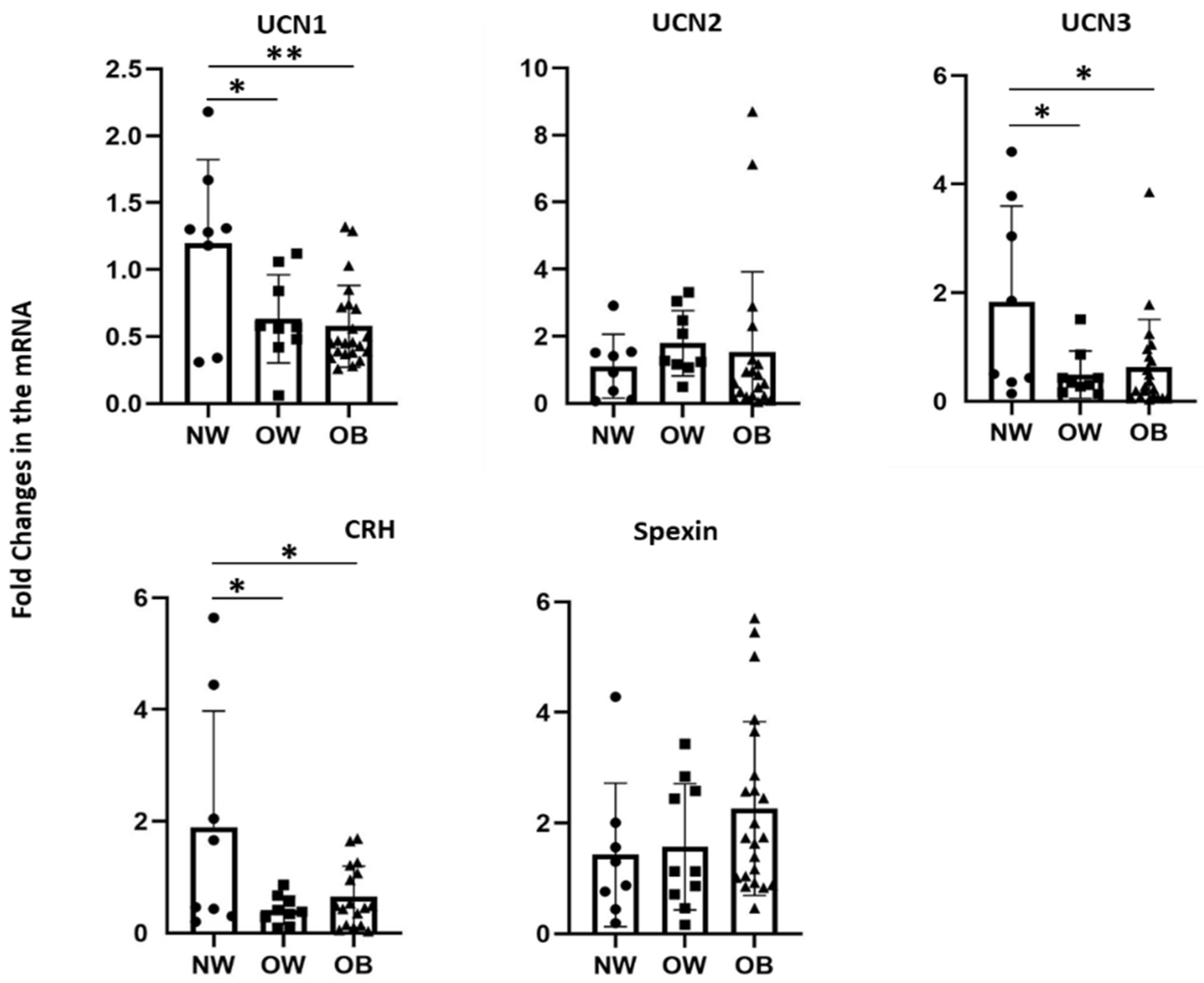
3.2. Effects of Bodyweight on Transcript Levels of CRF Peptides in PBMCs
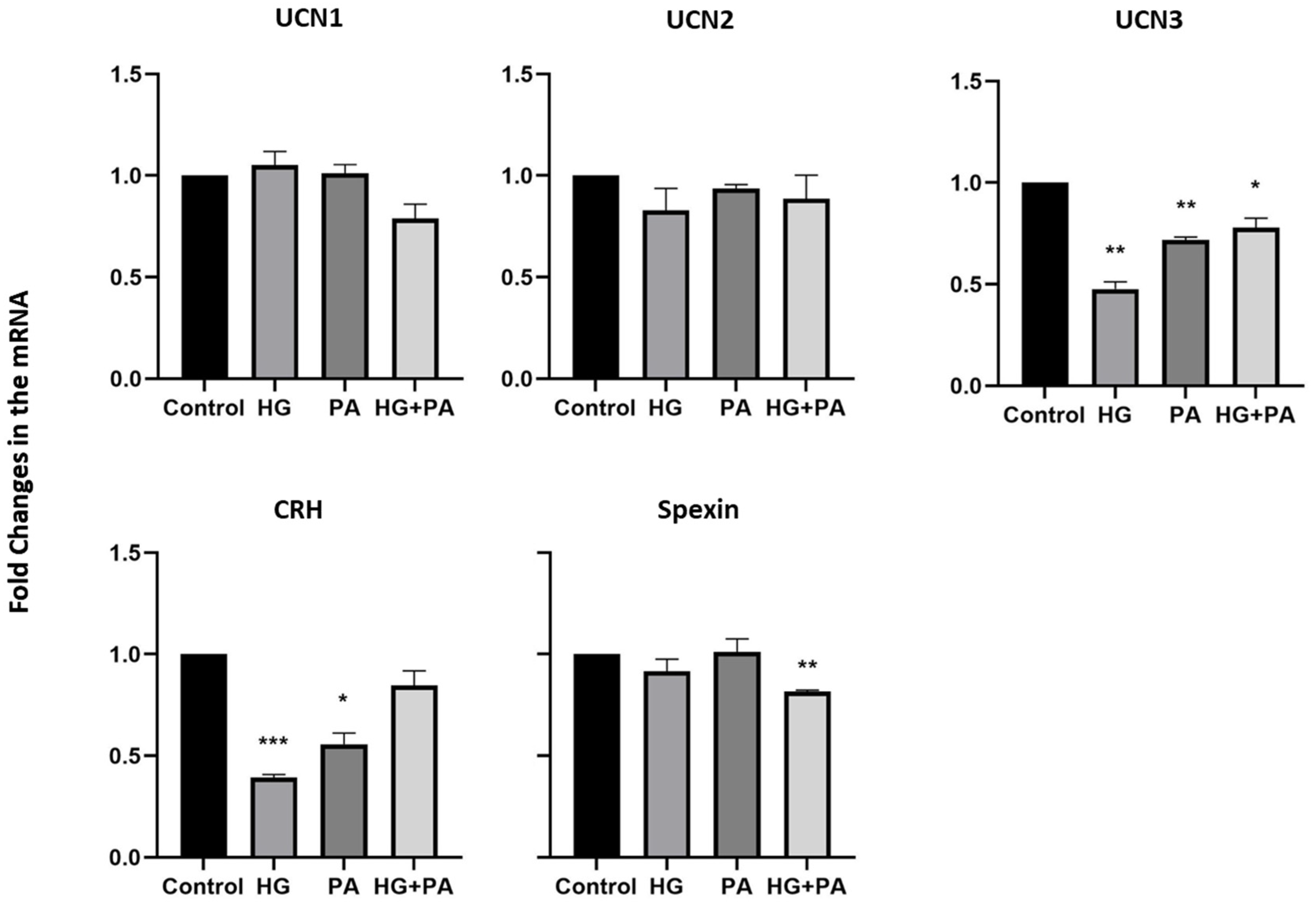
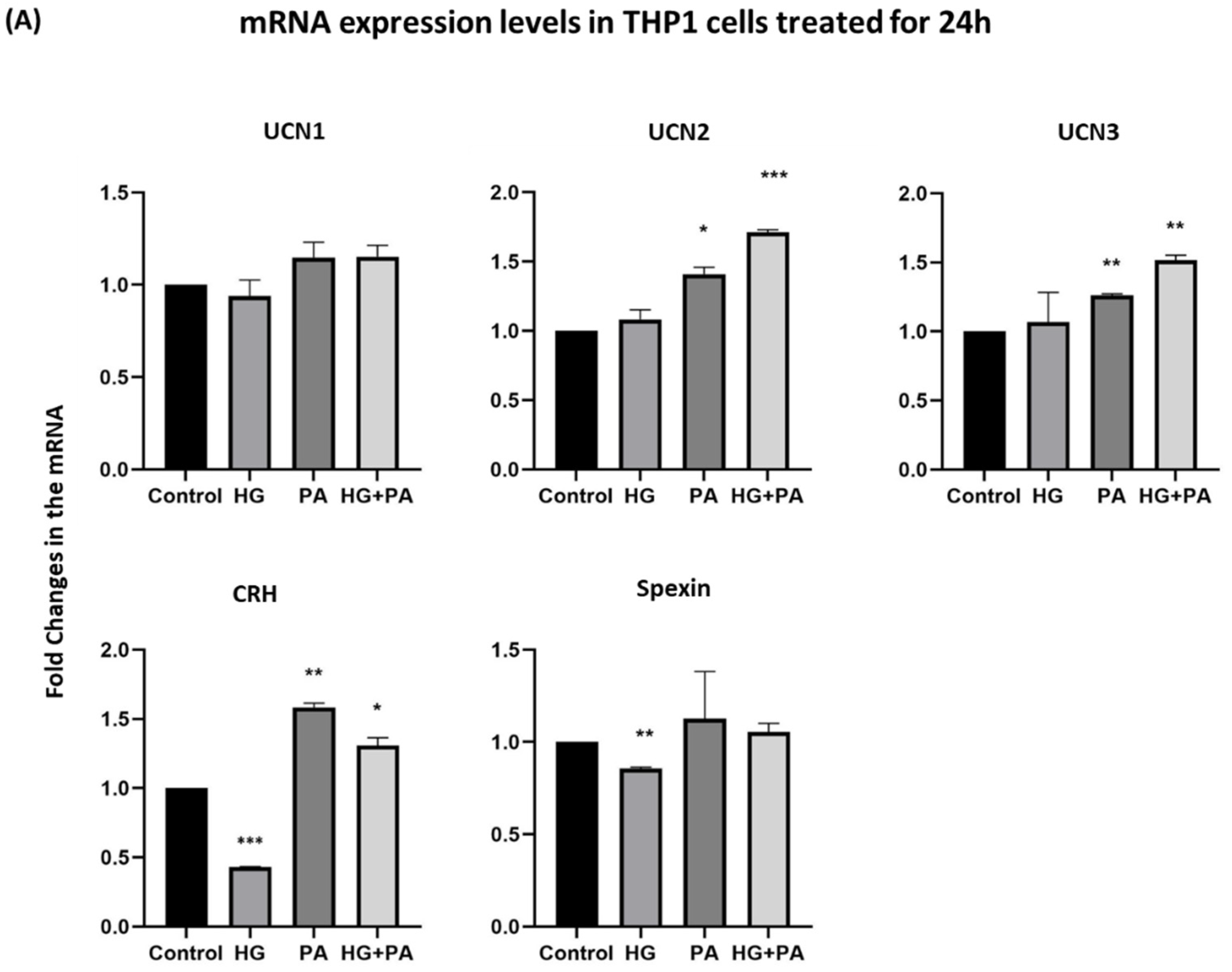
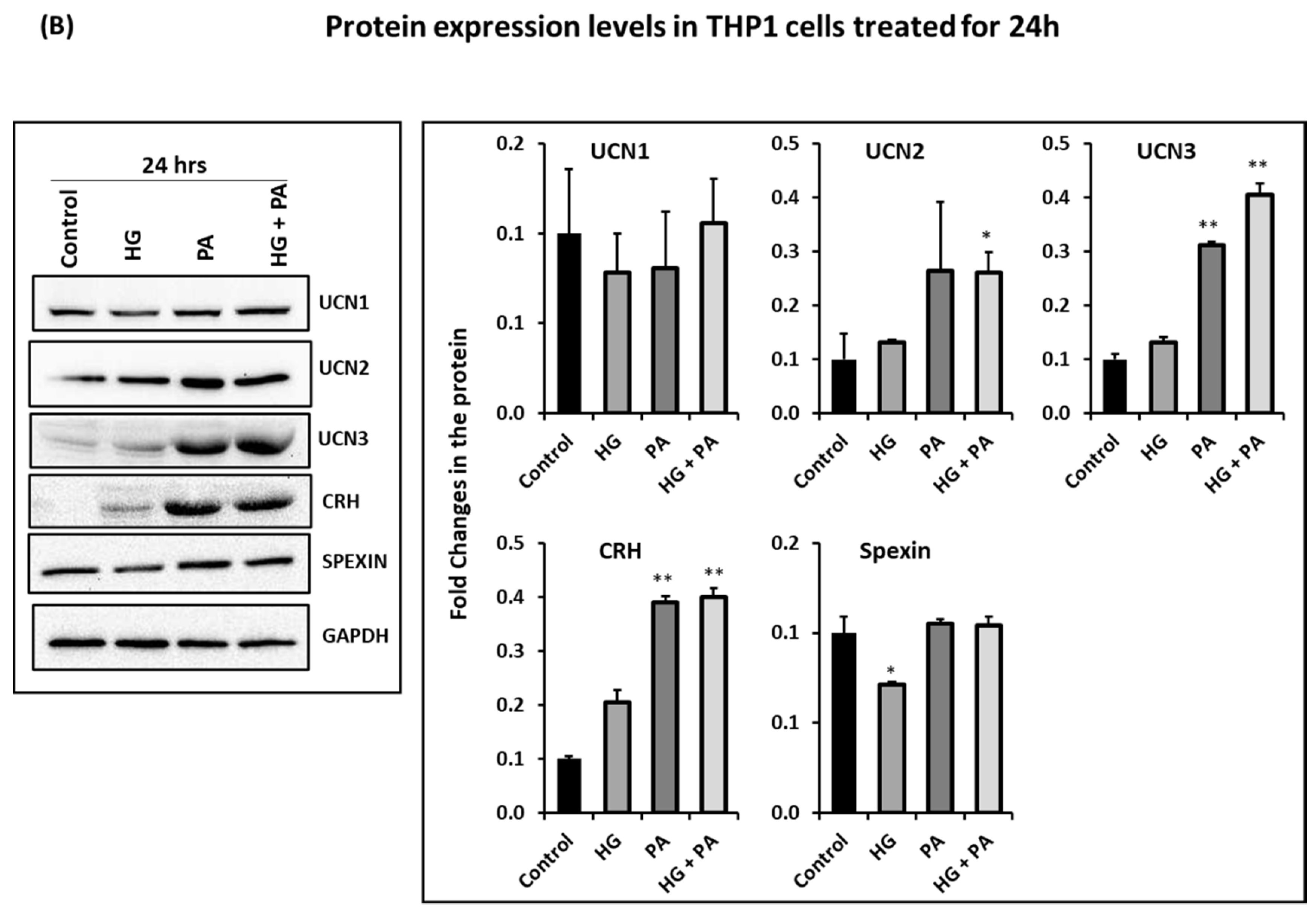
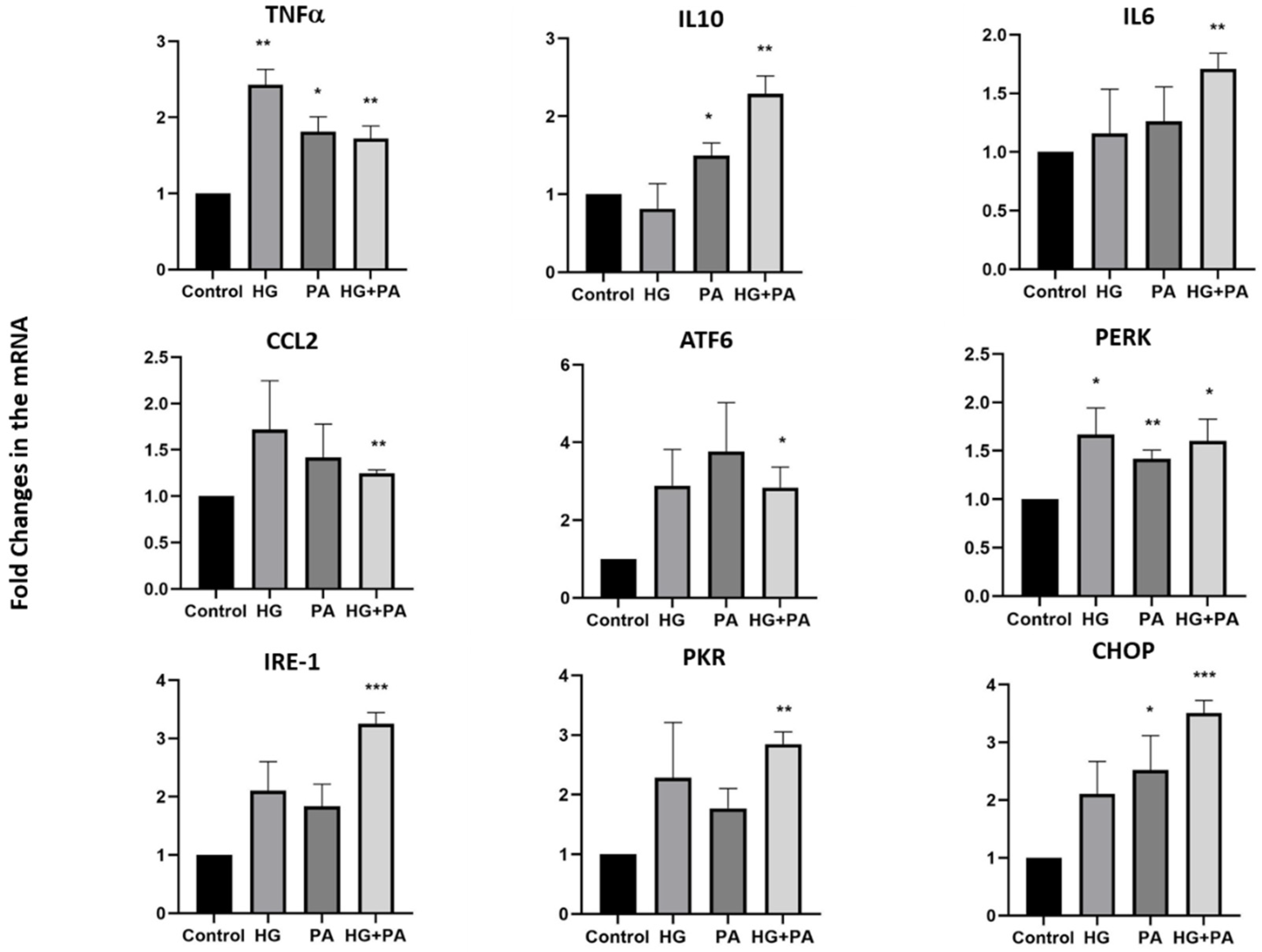
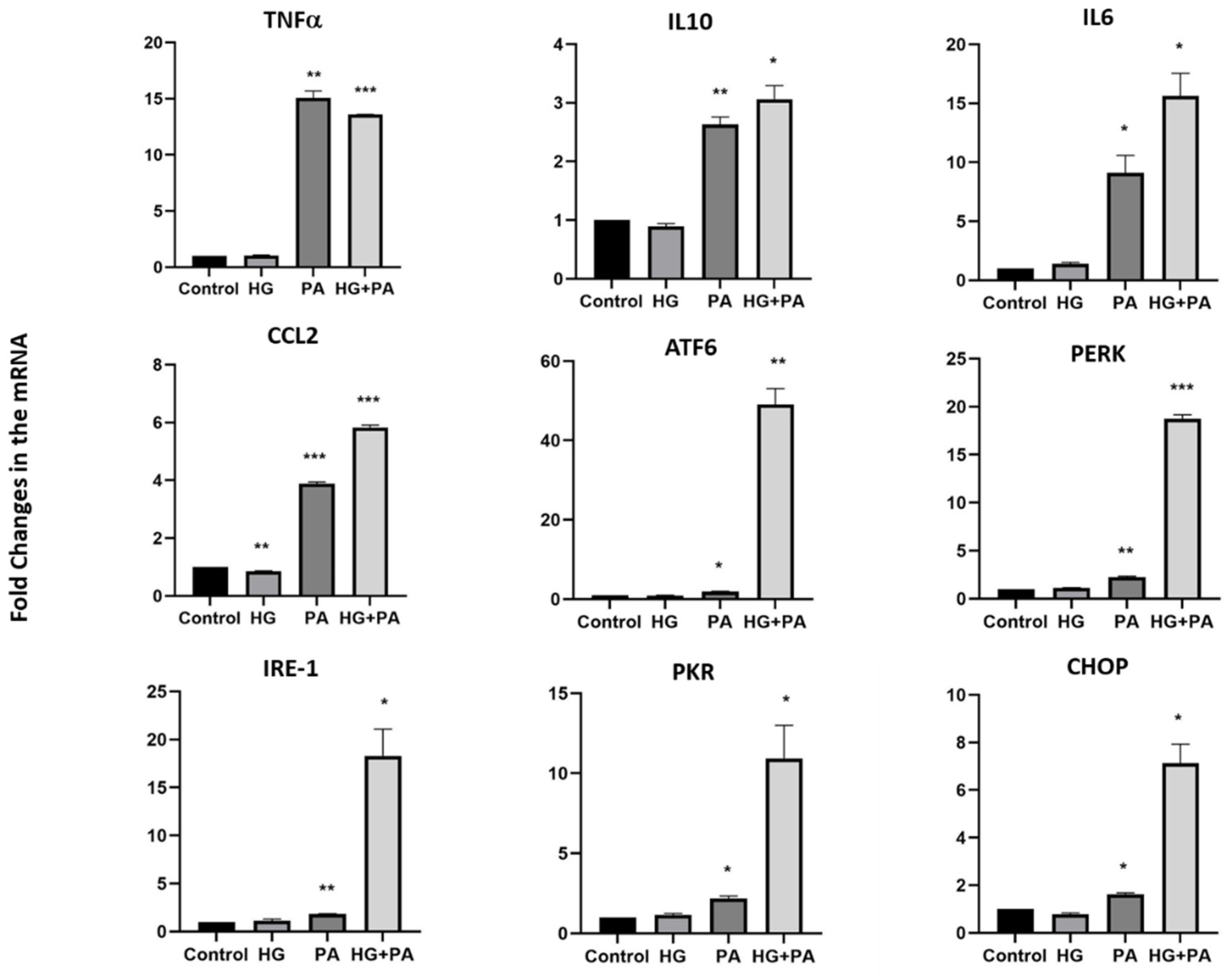
3.3. Metabolic Stress Affects the Expression of UCN1,2,3, CRH, and Spexin in THP1 Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ogden, C.L.; Carroll, M.D.; Lawman, H.G.; Fryar, C.D.; Kruszon-Moran, D.; Kit, B.K.; Flegal, K.M. Trends in Obesity Prevalence Among Children and Adolescents in the United States, 1988–1994 Through 2013–2014. JAMA 2016, 315, 2292–2299. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Tripathi, Y.; Pandey, V. Obesity and endoplasmic reticulum (ER) stresses. Front. Immunol. 2012, 3, 240. [Google Scholar] [CrossRef] [Green Version]
- Fekete, E.M.; Zorrilla, E.P. Physiology, pharmacology, and therapeutic relevance of urocortins in mammals: Ancient CRF paralogs. Front. Neuroendocrinol. 2007, 28, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Charmandari, E.; Tsigos, C.; Chrousos, G. Endocrinology of the stress response. Annu. Rev. Physiol. 2005, 67, 259–284. [Google Scholar] [CrossRef]
- Roustit, M.M.; Vaughan, J.M.; Jamieson, P.M.; Cleasby, M.E. Urocortin 3 activates AMPK and AKT pathways and enhances glucose disposal in rat skeletal muscle. J. Endocrinol. 2014, 223, 143–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dermitzaki, E.; Liapakis, G.; Androulidaki, A.; Venihaki, M.; Melissas, J.; Tsatsanis, C.; Margioris, A.N. Corticotrophin-Releasing Factor (CRF) and the urocortins are potent regulators of the inflammatory phenotype of human and mouse white adipocytes and the differentiation of mouse 3T3L1 pre-adipocytes. PLoS ONE 2014, 9, e97060. [Google Scholar] [CrossRef] [Green Version]
- Van der Meulen, T.; Donaldson, C.J.; Caceres, E.; Hunter, A.E.; Cowing-Zitron, C.; Pound, L.D.; Adams, M.W.; Zembrzycki, A.; Grove, K.L.; Huising, M.O. Urocortin3 mediates somatostatin-dependent negative feedback control of insulin secretion. Nat. Med. 2015, 21, 769–776. [Google Scholar] [CrossRef] [Green Version]
- Brar, B.K.; Railson, J.; Stephanou, A.; Knight, R.A.; Latchman, D.S. Urocortin increases the expression of heat shock protein 90 in rat cardiac myocytes in a MEK1/2-dependent manner. J. Endocrinol. 2002, 172, 283–293. [Google Scholar] [CrossRef] [Green Version]
- Brar, B.K.; Jonassen, A.K.; Egorina, E.M.; Chen, A.; Negro, A.; Perrin, M.H.; Mjos, O.D.; Latchman, D.S.; Lee, K.F.; Vale, W. Urocortin-II and urocortin-III are cardioprotective against ischemia reperfusion injury: An essential endogenous cardioprotective role for corticotropin releasing factor receptor type 2 in the murine heart. Endocrinology 2004, 145, 24–35, discussion 21–23. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, C.; Asakawa, A.; Ushikai, M.; Sakoguchi, T.; Amitani, H.; Terashi, M.; Cheng, K.; Chaolu, H.; Nakamura, N.; Inui, A. Comparison of the anorexigenic activity of CRF family peptides. Biochem. Biophys. Res. Commun. 2009, 390, 887–891. [Google Scholar] [CrossRef] [PubMed]
- Kuperman, Y.; Issler, O.; Regev, L.; Musseri, I.; Navon, I.; Neufeld-Cohen, A.; Gil, S.; Chen, A. Perifornical Urocortin-3 mediates the link between stress-induced anxiety and energy homeostasis. Proc. Natl. Acad. Sci. USA 2010, 107, 8393–8398. [Google Scholar] [CrossRef] [Green Version]
- Chao, H.; Digruccio, M.; Chen, P.; Li, C. Type 2 corticotropin-releasing factor receptor in the ventromedial nucleus of hypothalamus is critical in regulating feeding and lipid metabolism in white adipose tissue. Endocrinology 2012, 153, 166–176. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.; Brar, B.; Choi, C.S.; Rousso, D.; Vaughan, J.; Kuperman, Y.; Kim, S.N.; Donaldson, C.; Smith, S.M.; Jamieson, P.; et al. Urocortin 2 modulates glucose utilization and insulin sensitivity in skeletal muscle. Proc. Natl. Acad. Sci. USA 2006, 103, 16580–16585. [Google Scholar] [CrossRef] [Green Version]
- Kubat, E.; Mahajan, S.; Liao, M.; Ackerman, L.; Ohara, P.T.; Grady, E.F.; Bhargava, A. Corticotropin-releasing Factor Receptor 2 Mediates Sex-Specific Cellular Stress Responses. Mol. Med. 2013, 19, 212–222. [Google Scholar] [CrossRef]
- Gozal, D.; Jortani, S.; Snow, A.B.; Kheirandish-Gozal, L.; Bhattacharjee, R.; Kim, J.; Capdevila, O.S. Two-dimensional differential in-gel electrophoresis proteomic approaches reveal urine candidate biomarkers in pediatric obstructive sleep apnea. Am. J. Respir. Crit Care Med. 2009, 180, 1253–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavalakatt, S.; Khadir, A.; Madhu, D.; Hammad, M.; Devarajan, S.; Abubaker, J.; Al-Mulla, F.; Tuomilehto, J.; Tiss, A. Urocortin 3 Levels Are Impaired in Overweight Humans With and Without Type 2 Diabetes and Modulated by Exercise. Front. Endocrinol. 2019, 10, 762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.; Michal, J.J.; Williams, G.A.; Daniels, T.F.; Kunej, T. Cross species association examination of UCN3 and CRHR2 as potential pharmacological targets for antiobesity drugs. PLoS ONE 2006, 1, e80. [Google Scholar] [CrossRef]
- Wong, M.K.; Sze, K.H.; Chen, T.; Cho, C.K.; Law, H.C.; Chu, I.K.; Wong, A.O. Goldfish spexin: Solution structure and novel function as a satiety factor in feeding control. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E348–E366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walewski, J.L.; Ge, F.; Lobdell, H.t.; Levin, N.; Schwartz, G.J.; Vasselli, J.R.; Pomp, A.; Dakin, G.; Berk, P.D. Spexin is a novel human peptide that reduces adipocyte uptake of long chain fatty acids and causes weight loss in rodents with diet-induced obesity. Obesity 2014, 22, 1643–1652. [Google Scholar] [CrossRef] [Green Version]
- Lv, S.Y.; Zhou, Y.C.; Zhang, X.M.; Chen, W.D.; Wang, Y.D. Emerging Roles of NPQ/Spexin in Physiology and Pathology. Front. Pharmacol. 2019, 10, 457. [Google Scholar] [CrossRef] [Green Version]
- Hodges, S.K.; Teague, A.M.; Dasari, P.S.; Short, K.R. Effect of obesity and type 2 diabetes, and glucose ingestion on circulating spexin concentration in adolescents. Pediatr. Diabetes 2018, 19, 212–216. [Google Scholar] [CrossRef]
- Khadir, A.; Kavalakatt, S.; Madhu, D.; Devarajan, S.; Abubaker, J.; Al-Mulla, F.; Tiss, A. Spexin as an indicator of beneficial effects of exercise in human obesity and diabetes. Sci. Rep. 2020, 10, 10635. [Google Scholar] [CrossRef] [PubMed]
- Priego, T.; Sánchez, J.; Picó, C.; Ahrens, W.; De Henauw, S.; Kourides, Y.; Lissner, L.; Molnár, D.; Moreno, L.A.; Russo, P.; et al. TAS1R3 and UCN2 Transcript Levels in Blood Cells Are Associated With Sugary and Fatty Food Consumption in Children. J. Clin. Endocrinol. Metab. 2015, 100, 3556–3564. [Google Scholar] [CrossRef] [Green Version]
- Suda, T.; Kageyama, K.; Sakihara, S.; Nigawara, T. Physiological roles of urocortins, human homologues of fish urotensin I, and their receptors. Peptides 2004, 25, 1689–1701. [Google Scholar] [CrossRef] [PubMed]
- Kuczmarski, R.J.; Ogden, C.L.; Guo, S.S.; Grummer-Strawn, L.M.; Flegal, K.M.; Mei, Z.; Wei, R.; Curtin, L.R.; Roche, A.F.; Johnson, C.L. 2000 CDC Growth Charts for the United States: Methods and Development; Vital Health Stat Series 11, Data from the National Health survey; DHSS Publication No. (PHS)2002-1696; DHSS: Hyattsville, MD, USA, 2002; pp. 1–190. [Google Scholar]
- Tillinger, A.; Nostramo, R.; Kvetnansky, R.; Serova, L.; Sabban, E.L. Stress-induced changes in gene expression of urocortin 2 and other CRH peptides in rat adrenal medulla: Involvement of glucocorticoids. J. Neurochem. 2013, 125, 185–192. [Google Scholar] [CrossRef]
- Huising, M.O.; Pilbrow, A.P.; Matsumoto, M.; van der Meulen, T.; Park, H.; Vaughan, J.M.; Lee, S.; Vale, W.W. Glucocorticoids differentially regulate the expression of CRFR1 and CRFR2α in MIN6 insulinoma cells and rodent islets. Endocrinology 2011, 152, 138–150. [Google Scholar] [CrossRef]
- Kageyama, K.; Bradbury, M.J.; Zhao, L.; Blount, A.L.; Vale, W.W. Urocortin messenger ribonucleic acid: Tissue distribution in the rat and regulation in thymus by lipopolysaccharide and glucocorticoids. Endocrinology 1999, 140, 5651–5658. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Hover, C.V.; Lindberg, D.; Li, C. Central urocortin 3 and type 2 corticotropin-releasing factor receptor in the regulation of energy homeostasis: Critical involvement of the ventromedial hypothalamus. Front. Endocrinol. 2012, 3, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livingstone, D.E.W.; Jones, G.C.; Smith, K.; Jamieson, P.M.; Andrew, R.; Kenyon, C.J.; Walker, B.R. Understanding the Role of Glucocorticoids in Obesity: Tissue-Specific Alterations of Corticosterone Metabolism in Obese Zucker Rats1. Endocrinology 2000, 141, 560–563. [Google Scholar] [CrossRef] [PubMed]
- Rizza, R.A.; Mandarino, L.J.; Gerich, J.E. Cortisol-induced insulin resistance in man: Impaired suppression of glucose production and stimulation of glucose utilization due to a postreceptor detect of insulin action. J. Clin. Endocrinol. Metab. 1982, 54, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Alsiö, J.; Roman, E.; Olszewski, P.K.; Jonsson, P.; Fredriksson, R.; Levine, A.S.; Meyerson, B.J.; Hulting, A.L.; Lindblom, J.; Schiöth, H.B. Inverse association of high-fat diet preference and anxiety-like behavior: A putative role for urocortin 2. Genes Brain Behav. 2009, 8, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Cottone, P.; Sabino, V.; Nagy, T.R.; Coscina, D.V.; Levin, B.E.; Zorrilla, E.P. Centrally administered urocortin 2 decreases gorging on high-fat diet in both diet-induced obesity-prone and -resistant rats. Int. J. Obes. 2013, 37, 1515–1523. [Google Scholar] [CrossRef] [Green Version]
- Saruta, M.; Takahashi, K.; Suzuki, T.; Torii, A.; Kawakami, M.; Sasano, H. Urocortin 1 in colonic mucosa in patients with ulcerative colitis. J. Clin. Endocrinol. Metab. 2004, 89, 5352–5361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uzuki, M.; Sasano, H.; Muramatsu, Y.; Totsune, K.; Takahashi, K.; Oki, Y.; Iino, K.; Sawai, T. Urocortin in the synovial tissue of patients with rheumatoid arthritis. Clin. Sci. 2001, 100, 577–589. [Google Scholar] [CrossRef]
- Kohno, M.; Kawahito, Y.; Tsubouchi, Y.; Hashiramoto, A.; Yamada, R.; Inoue, K.I.; Kusaka, Y.; Kubo, T.; Elenkov, I.J.; Chrousos, G.P.; et al. Urocortin expression in synovium of patients with rheumatoid arthritis and osteoarthritis: Relation to inflammatory activity. J. Clin. Endocrinol. Metab. 2001, 86, 4344–4352. [Google Scholar] [CrossRef]
- Chatzaki, E.; Charalampopoulos, I.; Leontidis, C.; Mouzas, I.A.; Tzardi, M.; Tsatsanis, C.; Margioris, A.N.; Gravanis, A. Urocortin in human gastric mucosa: Relationship to inflammatory activity. J. Clin. Endocrinol. Metab. 2003, 88, 478–483. [Google Scholar] [CrossRef] [Green Version]
- Tsatsanis, C.; Androulidaki, A.; Dermitzaki, E.; Charalampopoulos, I.; Spiess, J.; Gravanis, A.; Margioris, A.N. Urocortin 1 and Urocortin 2 induce macrophage apoptosis via CRFR2. FEBS Lett. 2005, 579, 4259–4264. [Google Scholar] [CrossRef] [Green Version]
- Simpson, S.J.S.; Smith, L.I.F.; Jones, P.M.; Bowe, J.E. UCN2: A new candidate influencing pancreatic β-cell adaptations in pregnancy. J. Endocrinol. 2020, 245, 247–257. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Mean ± SD | p Value | Overall p | ||||
|---|---|---|---|---|---|---|---|
| Normal weight, NW (n = 8) | Overweight, OW (n = 10) | Obese (OB) (n = 22) | NW vs. OW | NW vs. OB | OW vs. OB | All Children | |
| Physical and clinical characteristics | |||||||
| Sex (M/F) | 5/3 | 3/7 | 14/8 | 0.188 | 0.956 | 0.082 | 0.196 |
| Age (years) | 10.5 ± 4.3 | 12.8 ± 3.4 | 11.3 ± 2.1 | 0.260 | 0.788 | 0.429 | 0.265 |
| Weight (Kg) | 38.2 ± 18.1 | 58.5 ± 14.6 | 83.6 ± 30.9 | 0.230 | <0.001 | 0.037 | <0.001 |
| Height (m) | 1.44 ± 0.25 | 1.54 ± 0.12 | 1.52 ± 0.17 | 0.482 | 0.511 | 0.969 | 0.460 |
| BMI (Kg/m2) | 17.16 ± 2.59 | 24.22 ± 2.97 | 35.05 ± 6.95 | 0.029 | <0.001 | <0.001 | <0.001 |
| Percentile # | 41.0 ± 18.8 | 91.8 ± 1.8 | 98.8 ± 0.6 | <0.001 | <0.001 | 0.078 | <0.001 |
| Body fat (%) | 6.1 ± 3.6 | 16.7 ± 6.8 | 32.9 ± 15.3 | 0.003 | <0.001 | 0.005 | <0.001 |
| Metabolic and Hormonal markers | |||||||
| Cholesterol (mmol/L) | 4.09 ± 0.62 | 4.57 ± 0.51 | 4.11 ± 0.96 | 0.451 | 0.997 | 0.357 | 0.344 |
| HDL (mmol/L) | 1.43 ± 0.31 | 1.34 ± 0.15 | 1.10 ± 0.28 | 0.748 | 0.013 | 0.074 | 0.008 |
| LDL (mmol/L) | 2.33 ± 0.59 | 2.82 ± 0.44 | 2.57 ± 0.82 | 0.323 | 0.692 | 0.639 | 0.355 |
| TGL (mmol/L) | 0.71 ± 0.36 | 0.99 ± 0.51 | 1.00 ± 0.45 | 0.410 | 0.295 | 0.999 | 0.297 |
| GLU (mmol/L) | 4.5 ± 0.4 | 5.6 ± 2.0 | 5.0 ± 0.4 | 0.120 | 0.633 | 0.326 | 0.132 |
| HbA1c (%) | 5.2 ± 0.3 | 5.7 ± 2.0 | 5.2 ± 0.7 | 0.670 | 0.998 | 0.525 | 0.528 |
| Insulin (mIU/L) | 3.6 ± 1.8 | 9.4 ± 4.4 | 17.15 ± 6.8 | 0.003 | 0.016 | 0.08 | 0.012 |
| HOMA IR | 0.74 ± 0.39 | 2.59 ± 2.34 | 5.54 ± 5.43 | 0.043 | 0.001 | 0.010 | 0.001 |
| Adiponectin (µg/mL) | 9.8 ± 1.1 | 8.6 ± 1.3 | 7.6 ± 2.7 | 0.496 | 0.057 | 0.473 | 0.065 |
| TNFα (ng/mL) | 1.21 ± 0.59 | 31.79 ± 9.40 | 25.86 ± 34.60 | 0.046 | 0.073 | 0.826 | 0.344 |
| Other markers | |||||||
| Leptin (ng/mL) | 7.78 ± 2.88 | 13.12 ± 6.67 | 11.17 ± 4.04 | 0.053 | 0.197 | 0.521 | 0.041 |
| C-pep (ng/mL) | 366 ± 219 | 187 ± 75 | 112 ± 41 | 0.006 | <0.001 | 0.221 | 0.064 |
| Glucagon (ng/mL) | 4.04 ± 1.81 | 1.62 ± 0.51 | 2.38 ± 1.14 | <0.001 | 0.005 | 0.236 | <0.001 |
| NGAL (ng/mL) | 24.71 ± 8.59 | 36.57 ± 17.87 | 24.31 ± 5.02 | 0.058 | 0.996 | 0.013 | <0.001 |
| RBP4 (µg/mL) | 19.45 ± 2.92 | 54.97 ± 19.74 | 19.10 ± 5.75 | <0.001 | 0.997 | <0.001 | 0.013 |
| sICAM-1 (ng/mL) | 132 ± 31 | 284 ± 233 | 157 ± 83 | 0.057 | 0.890 | 0.057 | <0.001 |
| ZAG (µg/mL) | 1.60 ± 0.22 | 5.02 ± 0.71 | 1.70 ± 0.37 | <0.001 | 0.847 | <0.001 | 0.037 |
| Resistin (ng/mL) | 5.16 ± 2.98 | 4.76 ± 1.46 | 6.75 ± 2.92 | 0.950 | 0.337 | 0.166 | <0.001 |
| Circulating CRH (pg/mL) | 76.80 ± 103.89 | 136.34 ± 105.24 | 23.38 ± 44.55 | 0.242 | 0.222 | 0.001 | 0.132 |
| Circulating UCN1 (pg/mL) | 43.38 ± 42.12 | 15.35 ± 27.67 | 32.30 ± 32.41 | 0.195 | 0.704 | 0.388 | 0.002 |
| Circulating UCN2 (ng/mL) | 1297 ± 362 | 1832 ± 400 | 1397 ± 374 | 0.014 | 0.800 | 0.013 | 0.206 |
| Circulating UCN3 (ng/mL) | 2.94 ± 0.99 | 33.28 ± 37.81 | 2.33 ± 0.32 | 0.006 | 0.997 | 0.001 | 0.007 |
| Circulating Spexin (ng/mL) | 14.67 ± 1.59 | 13.14 ± 2.13 | 18.04 ± 4.38 | 0.648 | 0.071 | 0.004 | 0.001 |
| All Children | FC CRH | FC UCN1 | FC UCN2 | FC UCN3 | FC SPX |
|---|---|---|---|---|---|
| Sex (M/F) | −0.045 | −0.023 | 0.063 | −0.089 | −0.039 |
| Age (years) | −0.323 | −0.169 | −0.186 | −0.069 | −0.186 |
| Weight (Kg) | −0.091 | −0.352 * | 0.026 | −0.129 | 0.093 |
| Height (m) | −0.030 | −0.240 | −0.030 | −0.033 | −0.085 |
| BMI (Kg/m *) | −0.141 | −0.362 * | −0.295 | −0.308 | 0.291 |
| Percentiles | −0.132 | −0.351 * | −0.143 | −0.234 | 0.196 |
| Body fat (%) | −0.019 | −0.423 * | 0.087 | −0.051 | 0.059 |
| Cholesterol (mmol/L) | −0.101 | −0.427 ** | 0.031 | −0.164 | 0.120 |
| HDL (mmol/L) | 0.145 | 0.071 | 0.011 | 0.094 | 0.125 |
| LDL (mmol/L) | 0.058 | 0.261 | −0.130 | 0.140 | −0.232 |
| TGL (mmol/L) | 0.144 | −0.034 | 0.058 | 0.037 | 0.252 |
| GLU (mmol/L) | −0.043 | 0.047 | 0.127 | −0.039 | 0.208 |
| HBA1C (%) | −0.203 | −0.039 | 0.028 | −0.230 | −0.032 |
| Insulin (mIU/L) | −0.040 | −0.047 | 0.182 | 0.075 | −0.244 |
| HOMA IR | −0.239 | −0.333 | −0.256 | −0.317 | 0.048 |
| Adiponectin (µg/mL) | 0.086 | 0.254 | −0.139 | 0.003 | −0.019 |
| TNFα (ng/mL) | −0.387 * | −0.050 | −0.208 | −0.388 * | 0.083 |
| Leptin (ng/mL) | −0.197 | −0.005 | 0.176 | −0.212 | 0.289 |
| C−pep (pg/mL) | 0.069 | 0.257 | 0.193 | 0.219 | −0.121 |
| glucagon Final (ng/mL) | 0.269 | 0.119 | −0.025 | 0.221 | −0.081 |
| NGAL (ng/mL) | −0.220 | 0.256 | 0.066 | −0.343 * | 0.206 |
| RBP4 (µg/mL) | −0.415 * | −0.027 | −0.056 | −0.328 | −0.175 |
| sICAM−1 (ng/mL) | −0.323 | 0.067 | 0.174 | −0.122 | −0.136 |
| ZAG (µg/mL) | −0.428 * | −0.082 | 0.054 | −0.224 | −0.190 |
| Resistin (ng/mL) | −0.068 | −0.235 | −0.078 | −0.205 | 0.057 |
| Circulating CRH (pg/mL) | −0.327 | 0.091 | 0.058 | −0.081 | −0.228 |
| Circulating UCN1 (pg/mL) | 0.097 | −0.209 | −0.121 | −0.090 | 0.114 |
| Circulating UCN2 (ng/mL) | −0.353 * | −0.274 | −0.215 | −0.209 | −0.014 |
| Circulating UCN3 (ng/mL) | 0.050 | 0.298 | 0.545 ** | 0.331 * | −0.083 |
| Circulating Spexin (ng/mL) | 0.106 | −0.304 | −0.439 * | −0.263 | 0.307 |
| FC CRH | − | 0.293 | 0.663 ** | 0.668 ** | 0.258 |
| FC UCN1 | 0.293 | − | 0.377 | 0.409 * | 0.071 |
| FC UCN2 | 0.663 ** | 0.377 | − | 0.887 ** | 0.163 |
| FC UCN3 | 0.668 ** | 0.409 * | 0.887 ** | − | −0.056 |
| FC SPX | 0.258 | 0.071 | 0.163 | −0.056 | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kavalakatt, S.; Khadir, A.; Kochumon, S.; Madhu, D.; Devarajan, S.; Hammad, M.; Alam-Eldin, N.; Warsame, S.; Al-Kandari, H.; AlMahdi, M.; et al. Urocortin Neuropeptide Levels Are Impaired in the PBMCs of Overweight Children. Nutrients 2022, 14, 429. https://doi.org/10.3390/nu14030429
Kavalakatt S, Khadir A, Kochumon S, Madhu D, Devarajan S, Hammad M, Alam-Eldin N, Warsame S, Al-Kandari H, AlMahdi M, et al. Urocortin Neuropeptide Levels Are Impaired in the PBMCs of Overweight Children. Nutrients. 2022; 14(3):429. https://doi.org/10.3390/nu14030429
Chicago/Turabian StyleKavalakatt, Sina, Abdelkrim Khadir, Shihab Kochumon, Dhanya Madhu, Sriraman Devarajan, Maha Hammad, Nada Alam-Eldin, Samia Warsame, Hessa Al-Kandari, Maria AlMahdi, and et al. 2022. "Urocortin Neuropeptide Levels Are Impaired in the PBMCs of Overweight Children" Nutrients 14, no. 3: 429. https://doi.org/10.3390/nu14030429
APA StyleKavalakatt, S., Khadir, A., Kochumon, S., Madhu, D., Devarajan, S., Hammad, M., Alam-Eldin, N., Warsame, S., Al-Kandari, H., AlMahdi, M., Ahmad, R., Koistinen, H. A., Tuomilehto, J., Al-Mulla, F., Abubaker, J., & Tiss, A. (2022). Urocortin Neuropeptide Levels Are Impaired in the PBMCs of Overweight Children. Nutrients, 14(3), 429. https://doi.org/10.3390/nu14030429