
Association between FADS Gene Expression and Polyunsaturated Fatty Acids in Breast Milk


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Breast Milk Collection and Questionnaires
2.3. Fatty Acids Analysis
2.4. RNA Extraction
2.5. Quantitative Real-Time PCR
2.6. Statistical Analysis
3. Results
3.1. Maternal Demographics of Participants
3.2. PUFA Profile of Breast Milk
3.3. Groups Divided by the mRNA Expression of FADS Gene
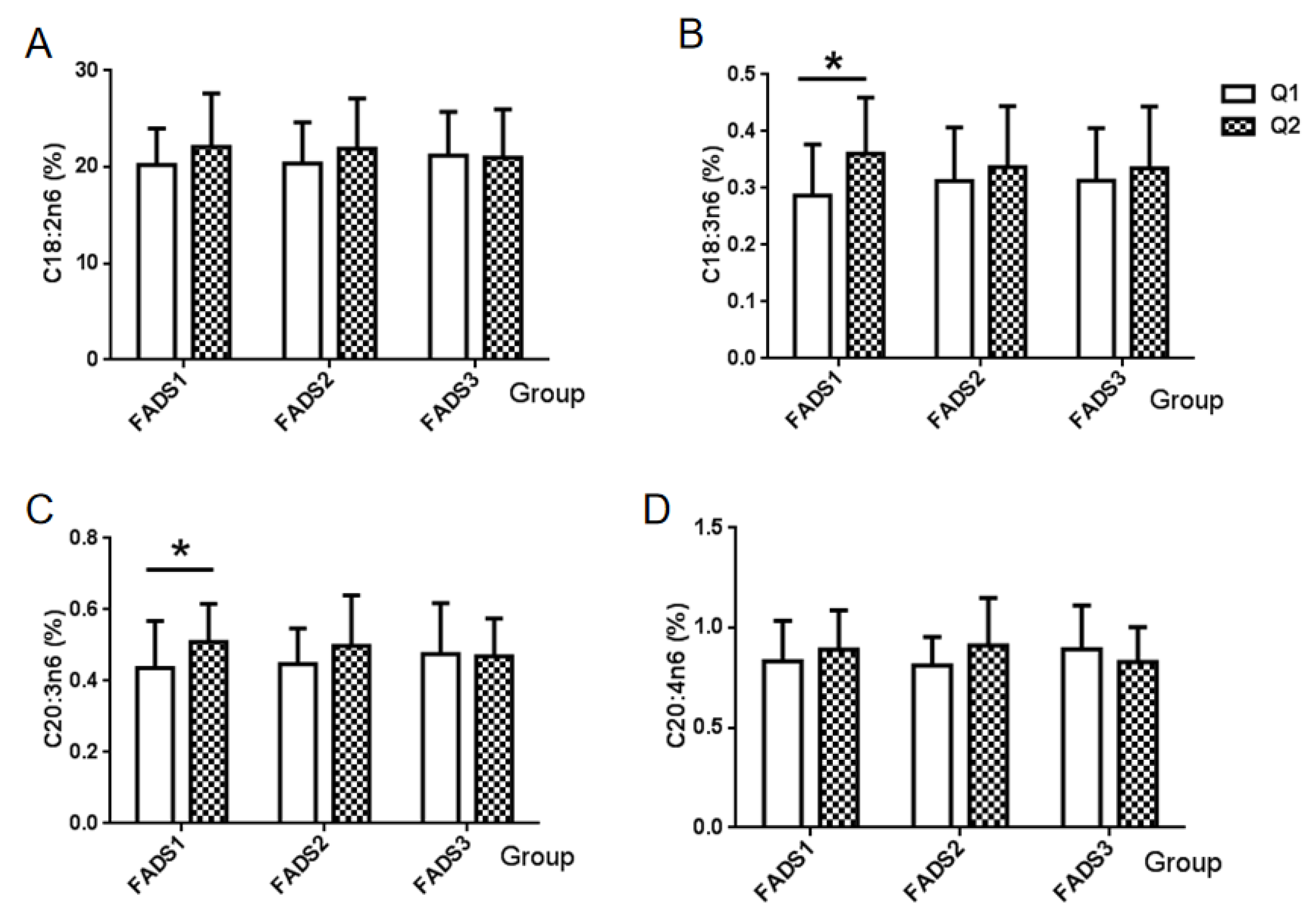
3.4. n-6 PUFA and the mRNA Expression of FADS Gene
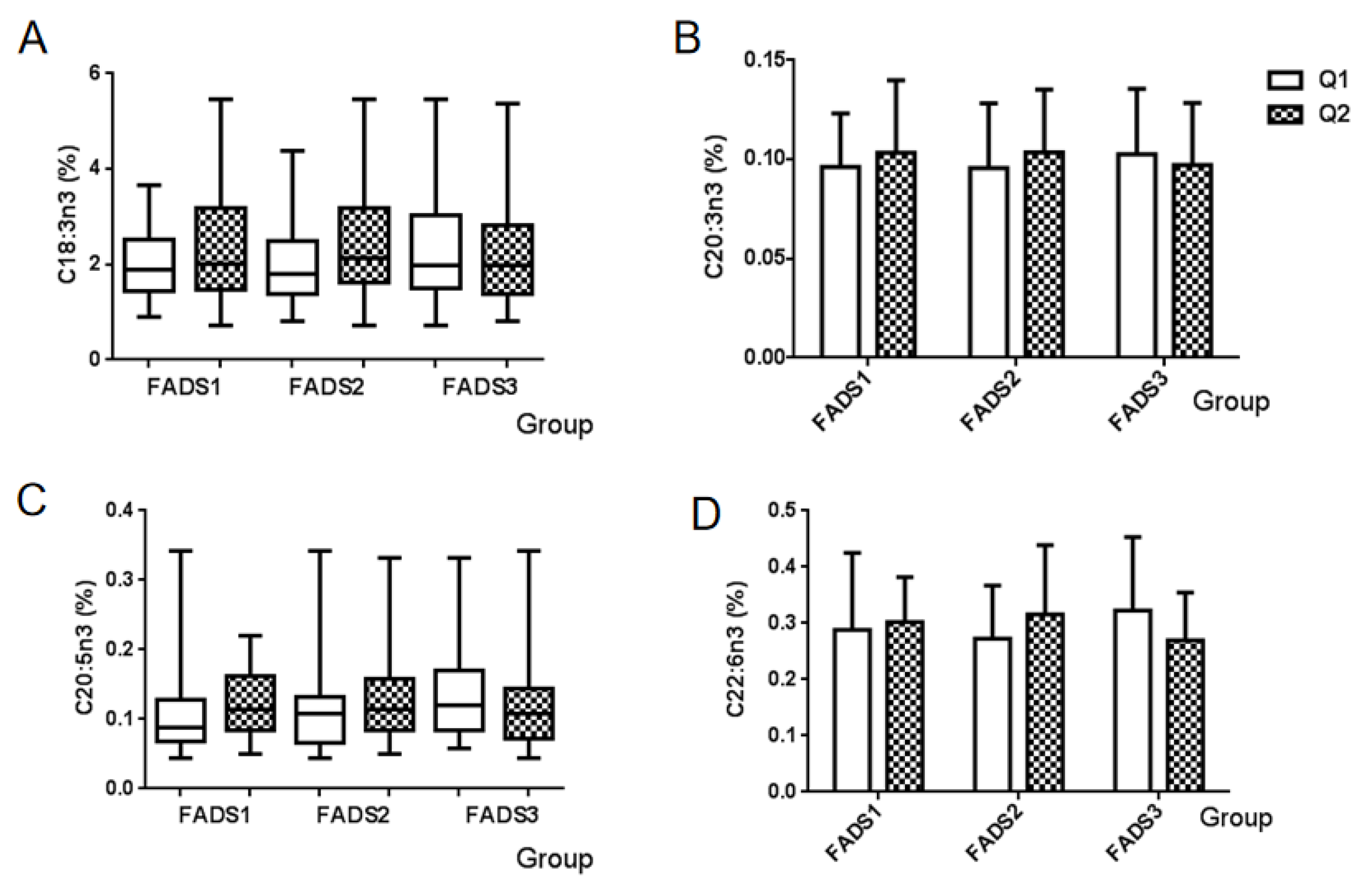
3.5. n-3 PUFA and the mRNA Expression of FADS Gene
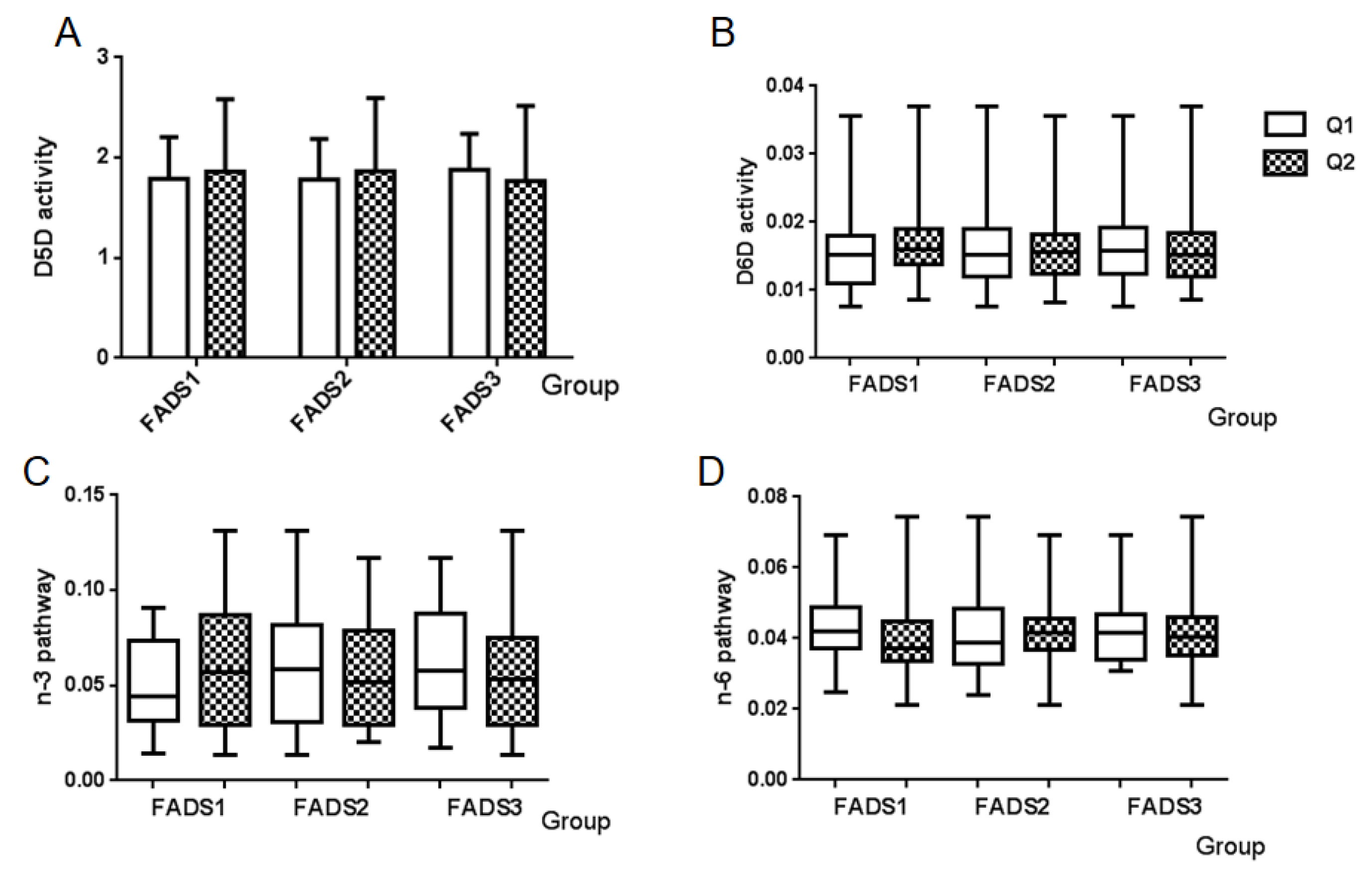
3.6. Enzyme Activity and PUFA Pathway Activity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lyons, K.E.; Ryan, C.A.; Dempsey, E.M.; Ross, R.P.; Stanton, C. Breast milk, a source of beneficial microbes and associated benefits for infant health. Nutrients 2020, 12, 1039. [Google Scholar] [CrossRef]
- Miliku, K.; Duan, Q.L.; Moraes, T.J.; Becker, A.B.; Mandhane, P.J.; Turvey, S.E.; Lefebvre, D.L.; Sears, M.R.; Subbarao, P.; Field, C.J.; et al. Human milk fatty acid composition is associated with dietary, genetic, sociodemographic, and environmental factors in the CHILD Cohort Study. Am. J. Clin. Nutr. 2019, 110, 1370–1383. [Google Scholar] [CrossRef]
- Keikha, M.; Bahreynian, M.; Saleki, M.; Kelishadi, R. Macro- and micronutrients of human milk composition: Are they related to maternal diet? A comprehensive systematic review. Breastfeed. Med. 2017, 12, 517–527. [Google Scholar] [CrossRef]
- Bravi, F.; Wiens, F.; Decarli, A.; Dal Pont, A.; Agostoni, C.; Ferraroni, M. Impact of maternal nutrition on breast-milk composition: A systematic review. Am. J. Clin. Nutr. 2016, 104, 646–662. [Google Scholar] [CrossRef] [Green Version]
- Innis, S.M. Impact of maternal diet on human milk composition and neurological development of infants. Am. J. Clin. Nutr. 2014, 99, 734S–741S. [Google Scholar] [CrossRef] [Green Version]
- Andreas, N.J.; Kampmann, B.; Mehring, L.K. Human breast milk: A review on its composition and bioactivity. Early Hum. Dev. 2015, 91, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Koska, J.; Ozias, M.K.; Deer, J.; Kurtz, J.; Salbe, A.D.; Harman, S.M.; Reaven, P.D. A human model of dietary saturated fatty acid induced insulin resistance. Metabolism 2016, 65, 1621–1628. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, M.M.; Jew, S.; Jones, P.J. Health benefits and evaluation of healthcare cost savings if oils rich in monounsaturated fatty acids were substituted for conventional dietary oils in the United States. Nutr. Rev. 2017, 75, 163–174. [Google Scholar] [CrossRef]
- Mennitti, L.V.; Oliveira, J.L.; Morais, C.A.; Estadella, D.; Oyama, L.M.; Oller, D.N.C.; Pisani, L.P. Type of fatty acids in maternal diets during pregnancy and/or lactation and metabolic consequences of the offspring. J. Nutr. Biochem. 2015, 26, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Mun, J.G.; Legette, L.L.; Ikonte, C.J.; Mitmesser, S.H. Choline and DHA in maternal and infant nutrition: Synergistic implications in brain and eye health. Nutrients 2019, 11, 1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, C.; Calder, P.C. Docosahexaenoic acid. Adv. Nutr. 2016, 7, 1139–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calder, P.C. Docosahexaenoic acid. Ann. Nutr. Metab. 2016, 69, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Eicosapentaenoic and docosahexaenoic acid derived specialised pro-resolving mediators: Concentrations in humans and the effects of age, sex, disease and increased omega-3 fatty acid intake. Biochimie 2020, 178, 105–123. [Google Scholar] [CrossRef] [PubMed]
- Zielinska, M.A.; Hamulka, J.; Grabowicz-Chadrzynska, I.; Brys, J.; Wesolowska, A. Association between breastmilk LC PUFA, carotenoids and psychomotor development of exclusively breastfed infants. Int. J. Environ. Res. Public Health 2019, 16, 1144. [Google Scholar] [CrossRef] [Green Version]
- Shulkin, M.; Pimpin, L.; Bellinger, D.; Kranz, S.; Fawzi, W.; Duggan, C.; Mozaffarian, D. n-3 fatty acid supplementation in mothers, preterm infants, and term infants and childhood psychomotor and visual development: A systematic review and meta-analysis. J. Nutr. 2018, 148, 409–418. [Google Scholar] [CrossRef]
- Miles, E.A.; Calder, P.C. Can early omega-3 fatty acid exposure reduce risk of childhood allergic disease? Nutrients 2017, 9, 784. [Google Scholar] [CrossRef] [Green Version]
- Silva-Zolezzi, I.; Samuel, T.M.; Spieldenner, J. Maternal nutrition: Opportunities in the prevention of gestational diabetes. Nutr. Rev. 2017, 75, 32–50. [Google Scholar] [CrossRef]
- World Health Organization. Infant and Young Child Feeding; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- Lauritzen, L.; Hansen, H.S.; Jorgensen, M.H.; Michaelsen, K.F. The essentiality of long chain n-3 fatty acids in relation to development and function of the brain and retina. Prog. Lipid Res. 2001, 40, 1–94. [Google Scholar] [CrossRef]
- Bahreynian, M.; Feizi, A.; Kelishadi, R. Is fatty acid composition of breast milk different in various populations? A systematic review and meta-analysis. Int. J. Food Sci. Nutr. 2020, 71, 909–920. [Google Scholar] [CrossRef]
- Liu, M.J.; Li, H.T.; Yu, L.X.; Xu, G.S.; Ge, H.; Wang, L.L.; Zhang, Y.L.; Zhou, Y.B.; Li, Y.; Bai, M.X.; et al. A correlation study of DHA dietary intake and plasma, erythrocyte and breast milk DHA concentrations in lactating women from Coastland, Lakeland, and Inland areas of China. Nutrients 2016, 8, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Casanova, I.; Rzehak, P.; Stein, A.D.; Garcia, F.R.; Rivera, D.J.; Barraza-Villarreal, A.; Demmelmair, H.; Romieu, I.; Villalpando, S.; Martorell, R.; et al. Maternal single nucleotide polymorphisms in the fatty acid desaturase 1 and 2 coding regions modify the impact of prenatal supplementation with DHA on birth weight. Am. J. Clin. Nutr. 2016, 103, 1171–1178. [Google Scholar] [CrossRef] [Green Version]
- Blanchard, H.; Legrand, P.; Pedrono, F. Fatty acid desaturase 3 (Fads3) is a singular member of the Fads cluster. Biochimie 2011, 93, 87–90. [Google Scholar] [CrossRef]
- Sosa-Castillo, E.; Rodriguez-Cruz, M.; Molto-Puigmarti, C. Genomics of lactation: Role of nutrigenomics and nutrigenetics in the fatty acid composition of human milk. Br. J. Nutr. 2017, 118, 161–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, L.M.; Howard, T.D.; Ruczinski, I.; Kanchan, K.; Seeds, M.C.; Mathias, R.A.; Chilton, F.H. Tissue-specific impact of FADS cluster variants on FADS1 and FADS2 gene expression. PLoS ONE 2018, 13, e194610. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Zhao, J.; Kothapalli, K.; Li, X.; Li, H.; Han, Y.; Mi, S.; Zhao, W.; Li, Q.; Zhang, H.; et al. A regulatory insertion-deletion polymorphism in the FADS gene cluster influences PUFA and lipid profiles among Chinese adults: A population-based study. Am. J. Clin. Nutr. 2018, 107, 867–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vass, R.A.; Kemeny, A.; Dergez, T.; Ertl, T.; Reglodi, D.; Jungling, A.; Tamas, A. Distribution of bioactive factors in human milk samples. Int. Breastfeed. J. 2019, 14, 9. [Google Scholar] [CrossRef] [Green Version]
- Ding, Z.; Liu, G.L.; Li, X.; Chen, X.Y.; Wu, Y.X.; Cui, C.C.; Zhang, X.; Yang, G.; Xie, L. Association of polyunsaturated fatty acids in breast milk with fatty acid desaturase gene polymorphisms among Chinese lactating mothers. Prostaglandins Leukot. Essent. Fat. Acids 2016, 109, 66–71. [Google Scholar] [CrossRef]
- Tian, H.M.; Wu, Y.X.; Lin, Y.Q.; Chen, X.Y.; Yu, M.; Lu, T.; Xie, L. Dietary patterns affect maternal macronutrient intake levels and the fatty acid profile of breast milk in lactating Chinese mothers. Nutrition 2019, 58, 83–88. [Google Scholar] [CrossRef]
- Alsaweed, M.; Hepworth, A.R.; Lefevre, C.; Hartmann, P.E.; Geddes, D.T.; Hassiotou, F. Human milk microRNA and total RNA differ depending on milk fractionation. J. Cell. Biochem. 2015, 116, 2397–2407. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.C.; Chang, W.T.; Chang, H.Y.; Chung, H.F.; Chen, F.P.; Huang, Y.F.; Hsu, C.C.; Hwang, S.J. FADS gene polymorphisms, fatty acid desaturase activities, and HDL-C in type 2 diabetes. Int. J. Environ. Res. Public Health 2017, 14, 572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juber, B.A.; Jackson, K.H.; Johnson, K.B.; Harris, W.S.; Baack, M.L. Breast milk DHA levels may increase after informing women: A community-based cohort study from South Dakota USA. Int. Breastfeed. J. 2016, 12, 7. [Google Scholar] [CrossRef] [Green Version]
- Aumeistere, L.; Ciprovica, I.; Zavadska, D.; Volkovs, V. Fish intake reflects on DHA level in breast milk among lactating women in Latvia. Int. Breastfeed. J. 2018, 13, 33. [Google Scholar] [CrossRef]
- Kim, H.; Kang, S.; Jung, B.M.; Yi, H.; Jung, J.A.; Chang, N. Breast milk fatty acid composition and fatty acid intake of lactating mothers in South Korea. Br. J. Nutr. 2017, 117, 556–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meldrum, S.J.; Li, Y.; Zhang, G.; Heaton, A.; D’Vaz, N.; Manz, J.; Reischl, E.; Koletzko, B.V.; Prescott, S.L.; Simmer, K. Can polymorphisms in the fatty acid desaturase (FADS) gene cluster alter the effects of fish oil supplementation on plasma and erythrocyte fatty acid profiles? An exploratory study. Eur. J. Nutr. 2018, 57, 2583–2594. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.L.; Blomquist, S.A.; Hallmark, B.; Chilton, F.H. Omega-3 supplementation and heart disease: A population-based diet by gene analysis of clinical trial outcomes. Nutrients 2021, 13, 2154. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Balter, A.; Vodsky, V.; Odetallh, Y.; Ben-Dror, G.; Zhang, Y.; Zhao, A. Chinese breast milk fat composition and its associated dietary factors: A pilot study on lactating mothers in Beijing. Front. Nutr. 2021, 8, 606950. [Google Scholar] [CrossRef]
- Chilton, F.H.; Dutta, R.; Reynolds, L.M.; Sergeant, S.; Mathias, R.A.; Seeds, M.C. Precision nutrition and omega-3 polyunsaturated fatty acids: A case for personalized supplementation approaches for the prevention and management of human diseases. Nutrients 2017, 9, 1165. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, X.; Zhou, B.; Jiang, A.C.; Chai, L. An updated review of worldwide levels of docosahexaenoic and arachidonic acid in human breast milk by region. Public Health Nutr. 2016, 19, 2675–2687. [Google Scholar] [CrossRef] [Green Version]
- Al-Hilal, M.; Alsaleh, A.; Maniou, Z.; Lewis, F.J.; Hall, W.L.; Sanders, T.A.; O’Dell, S.D. Genetic variation at the FADS1-FADS2 gene locus influences delta-5 desaturase activity and LC-PUFA proportions after fish oil supplement. J. Lipid Res. 2013, 54, 542–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Ding, Z.; Li, X.; Chen, X.; Wu, Y.; Xie, L. Relationship between polyunsaturated fatty acid levels in maternal diets and human milk in the first month post-partum. J. Hum. Nutr. Diet. 2016, 29, 405–410. [Google Scholar] [CrossRef]
- Chinese Nutrition Society. Chinese Dietary Reference Intakes; China Science Publishing: Beijing, China, 2014; pp. 77–161. [Google Scholar]
- Lu, Z.Q.; Ren, Y.; Zhou, X.H.; Yu, X.F.; Huang, J.; Yu, D.Y.; Wang, X.X.; Wang, Y.Z. Maternal dietary linoleic acid supplementation promotes muscle fibre type transformation in suckling piglets. J. Anim. Physiol. Anim. Nutr. 2017, 101, 1130–1136. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Cruz, M.; Sanchez, R.; Sanchez, A.M.; Kelleher, S.L.; Sanchez-Munoz, F.; Maldonado, J.; Lopez-Alarcon, M. Participation of mammary gland in long-chain polyunsaturated fatty acid synthesis during pregnancy and lactation in rats. Biochim. Biophys. Acta 2011, 1811, 284–293. [Google Scholar] [CrossRef]
- Mathias, R.A.; Pani, V.; Chilton, F.H. Genetic variants in the FADS gene: Implications for dietary recommendations for fatty acid intake. Curr. Nutr. Rep. 2014, 3, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Koletzko, B.; Reischl, E.; Tanjung, C.; Gonzalez-Casanova, I.; Ramakrishnan, U.; Meldrum, S.; Simmer, K.; Heinrich, J.; Demmelmair, H. FADS1 and FADS2 polymorphisms modulate fatty acid metabolism and dietary impact on health. Annu. Rev. Nutr. 2019, 39, 21–44. [Google Scholar] [CrossRef]
- Yang, H.T.; Wang, R.Y.; Huang, S.Y.; Huang, C.L.; Su, K.P. Genetic polymorphisms of FADS1, FADS2, and FADS3 and fatty acid profiles in subjects received methadone maintenance therapy. Prostaglandins Leukot. Essent. Fatty Acids 2018, 136, 117–121. [Google Scholar] [CrossRef]
- Bernard, J.Y.; Pan, H.; Aris, I.M.; Moreno-Betancur, M.; Soh, S.; Yap, F.; Tan, K.H.; Shek, L.P.; Chong, Y.; Gluckman, P.D.; et al. Long-chain polyunsaturated fatty acids, gestation duration, and birth size: A Mendelian randomization study using fatty acid desaturase variants. Am. J. Clin. Nutr. 2018, 108, 92–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuchardt, J.P.; Kobe, T.; Witte, V.; Willers, J.; Gingrich, A.; Tesky, V.; Pantel, J.; Rujescu, D.; Illig, T.; Floel, A.; et al. Genetic variants of the FADS gene cluster are associated with erythrocyte membrane LC PUFA levels in patients with mild cognitive impairment. J. Nutr. Health Aging 2016, 20, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Panda, C.; Varadharaj, S.; Voruganti, V.S. PUFA, genotypes and risk for cardiovascular disease. Prostaglandins Leukot. Essent. Fatty Acids 2021, 176, 102377. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Innis, S.M. Genetic variants of the FADS1 FADS2 gene cluster are associated with altered (n-6) and (n-3) essential fatty acids in plasma and erythrocyte phospholipids in women during pregnancy and in breast milk during lactation. J. Nutr. 2008, 138, 2222–2228. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Athinarayanan, S.; Jiang, G.; Chalasani, N.; Zhang, M.; Liu, W. Fatty acid desaturase 1 gene polymorphisms control human hepatic lipid composition. Hepatology 2015, 61, 119–128. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence 5′→3′ |
|---|---|---|
| FADS1 | Forward | GTGGC TAGTG ATCGA CCGTA A |
| Reverse | ATTCT TGGTG GGCTC AAAGC | |
| FADS2 | Forward | AACTG GTGGA ATCAT CGCCA |
| Reverse | ATTCG CCCAG AACAA ACACG | |
| FADS3 | Forward | TCGTG ATGGG GCAGC TAAAG |
| Reverse | CTTCT TGCCA TACTC GACGG A |
| Characteristics | Mean ± SD/N (%) |
|---|---|
| Maternal age (year) | 29.58 ± 2.42 |
| Preconception BMI | 20.68 ± 2.85 |
| Gestational weight gain (kg) | 16.23 ± 5.53 |
| Gestational age (weeks) | 39.58 ± 0.93 |
| Birth weight (kg) | 3.45 ± 0.49 |
| Birth length (cm) | 50.59 ± 1.26 |
| Family income (yuan/month) | |
| <5000 | 9 (18.00) |
| 5000–9999 | 26 (52.00) |
| ≥10,000 | 15 (30.00) |
| Mode of delivery | |
| Vaginal | 26 (52.00) |
| Cesarean | 24 (48.00) |
| Feeding patterns | |
| Breastfeeding | 36 (72.00) |
| Mixed feeding | 14 (28.00) |
| Series | PUFA | Percentage of Total FA (%) (Mean ± SD/Median (P25, P75)) | |
|---|---|---|---|
| n-6 | Linoleic acid (LA) | C18:2n6 | 21.0886 ± 4.7437 |
| γ-Linolenic acid | C18:3n6 | 0.3245 ± 0.1008 | |
| Eicosatrienoic acid | C20:3n6 | 0.4726 ± 0.1245 | |
| Arachidonic acid (AA) | C20:4n6 | 0.8616 ± 0.1996 | |
| n-3 | α-Linolenic acid (ALA) | C18:3n3 | 1.9504 (1.4444, 2.7370) |
| Eicosatrienoic acid | C20:3n3 | 0.0999 ± 0.0320 | |
| Eicosapentaenoic acid (EPA) | C20:5n3 | 0.1097 (0.0787, 0.1499) | |
| Docosahexaenoic acid (DHA) | C22:6n3 | 0.2952 ± 0.1114 | |
| Gene | Relative Level of mRNA Median (P25, P75) | Group | p | |
|---|---|---|---|---|
| mRNA Q1 | mRNA Q2 | |||
| FADS1 | −0.1302 (−0.1319, −0.1255) | ≤−0.1302 | >−0.1302 | <0.001 |
| FADS2 | −0.2619 (−0.3497, −0.1359) | ≤−0.2619 | >−0.2619 | <0.001 |
| FADS3 | −0.5039 (−0.8220, −0.0132) | ≤−0.5039 | >−0.5039 | <0.001 |
| Index | Calculation Formula | Mean ± SD/Median (P25, P75) |
|---|---|---|
| D5D * | C20:4n6/C20:3n6 | 1.8482 ± 0.5725 |
| D6D * | C18:3n6/C18:2n6 | 0.0154 (0.0127, 0.0190) |
| n-3 pathway | C20:5n3/C18:3n3 | 0.0554 (0.0303, 0.0798) |
| n-6 pathway | C20:4n6/C18:2n6 | 0.0418 (0.0354, 0.0481) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, H.; Yu, H.; Lin, Y.; Li, Y.; Xu, W.; Chen, Y.; Liu, G.; Xie, L. Association between FADS Gene Expression and Polyunsaturated Fatty Acids in Breast Milk. Nutrients 2022, 14, 457. https://doi.org/10.3390/nu14030457
Tian H, Yu H, Lin Y, Li Y, Xu W, Chen Y, Liu G, Xie L. Association between FADS Gene Expression and Polyunsaturated Fatty Acids in Breast Milk. Nutrients. 2022; 14(3):457. https://doi.org/10.3390/nu14030457
Chicago/Turabian StyleTian, Huimin, Haitao Yu, Yiqi Lin, Yueting Li, Wenhui Xu, Yiru Chen, Guoliang Liu, and Lin Xie. 2022. "Association between FADS Gene Expression and Polyunsaturated Fatty Acids in Breast Milk" Nutrients 14, no. 3: 457. https://doi.org/10.3390/nu14030457
APA StyleTian, H., Yu, H., Lin, Y., Li, Y., Xu, W., Chen, Y., Liu, G., & Xie, L. (2022). Association between FADS Gene Expression and Polyunsaturated Fatty Acids in Breast Milk. Nutrients, 14(3), 457. https://doi.org/10.3390/nu14030457