
Omega-3 Fatty Acids and Their Interaction with the Gut Microbiome in the Prevention and Amelioration of Type-2 Diabetes

 , , , ,
, , , ,  and
and 
Abstract
:
1. Introduction
2. The Role of Omega-3 Fatty Acids in Type-2 Diabetes Mellitus (T2DM)
3. The Role of Gut Microbiota in the Pathophysiology of Type-2 Diabetes
4. The Intricate Interaction between Omega-3 Fatty Acids and the Gut Microbiome
5. Role of Gut Microbiota in Alleviating the Inflammatory Responses in T2DM
6. Combinatorial Strategy to Deal with Type 2 Diabetes
7. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Silva, K.; Demmer, R.T.; Jonsson, D.; Mousa, A.; Forbes, A.; Enticott, J. A data-driven biocomputing pipeline with meta-analysis on high throughput transcriptomics to identify genome-wide miRNA markers associated with type 2 diabetes. Heliyon 2022, 8, e08886. [Google Scholar] [CrossRef] [PubMed]
- Valder, S.; Brinkmann, C. Exercise for the Diabetic Gut—Potential Health Effects and Underlying Mechanisms. Nutrients 2022, 14, 813. [Google Scholar] [CrossRef] [PubMed]
- McCrimmon, R.J.; Ryan, C.M.; Frier, B.M. Diabetes and cognitive dysfunction. Lancet 2012, 379, 2291–2299. [Google Scholar] [CrossRef]
- Pantalone, K.M.; Hobbs, T.M.; Wells, B.J.; Kong, S.X.; Kattan, M.W.; Bouchard, J.; Yu, C.; Sakurada, B.; Milinovich, A.; Weng, W.; et al. Clinical characteristics, complications, comorbidities and treatment patterns among patients with type 2 diabetes mellitus in a large integrated health system. BMJ Open Diabetes Res. Care 2015, 3, e000093. [Google Scholar] [CrossRef] [Green Version]
- Domingueti, C.P.; Dusse, L.M.S.A.; das Graças Carvalho, M.; de Sousa, L.P.; Gomes, K.B.; Fernandes, A.P. Diabetes mellitus: The linkage between oxidative stress, inflammation, hypercoagulability and vascular complications. J. Diabetes Its Complicat. 2016, 30, 738–745. [Google Scholar] [CrossRef]
- Bagchi, D.; Nair, S. Nutritional and Therapeutic Interventions for Diabetes and Metabolic Syndrome; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Hall, R.M.; Strong, A.P.; Krebs, J.D. Importance of low carbohydrate diets in diabetes management. Nutr. Diet. Suppl. 2016, 8, 9–19. [Google Scholar]
- Sagild, U.; Littauer, J.; Jespersen, C.S.; Andersen, S. Epidemiological studies in Greenland 1962–1964. I. Diabetes mellitus in Eskimos. Acta Med. Scand. 1966, 179, 29–39. [Google Scholar] [CrossRef]
- Bang, H.O.; Dyerberg, J.; Sinclair, H.M. The composition of the Eskimo food in north western Greenland. Am. J. Clin. Nutr. 1980, 33, 2657–2661. [Google Scholar] [CrossRef] [Green Version]
- Kromann, N.; Green, A. Epidemiological studies in the Upernavik district, Greenland. Incidence of some chronic diseases 1950–1974. Acta Med. Scand. 1980, 208, 401–406. [Google Scholar] [CrossRef]
- Pitsavos, C.; Panagiotakos, D.B.; Tzima, N.; Chrysohoou, C.; Economou, M.; Zampelas, A.; Stefanadis, C. Adherence to the Mediterranean diet is associated with total antioxidant capacity in healthy adults: The ATTICA study. Am. J. Clin. Nutr. 2005, 82, 694–699. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Missbach, B.; Konig, J.; Hoffmann, G. Adherence to a Mediterranean diet and risk of diabetes: A systematic review and meta-analysis. Public Health Nutr. 2015, 18, 1292–1299. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Gonzalez, M.A.; de la Fuente-Arrillaga, C.; Nunez-Cordoba, J.M.; Basterra-Gortari, F.J.; Beunza, J.J.; Vazquez, Z.; Benito, S.; Tortosa, A.; Bes-Rastrollo, M. Adherence to Mediterranean diet and risk of developing diabetes: Prospective cohort study. BMJ 2008, 336, 1348–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas-Salvado, J.; Bullo, M.; Babio, N.; Martinez-Gonzalez, M.A.; Ibarrola-Jurado, N.; Basora, J.; Estruch, R.; Covas, M.I.; Corella, D.; Aros, F.; et al. Reduction in the incidence of type 2 diabetes with the Mediterranean diet: Results of the PREDIMED-Reus nutrition intervention randomized trial. Diabetes Care 2011, 34, 14–19. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes Association. 5. Lifestyle Management: Standards of Medical Care in Diabetes-2019. Diabetes Care 2019, 42, S46–S60. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, T.A.; Ito, M.K.; Maki, K.C.; Orringer, C.E.; Bays, H.E.; Jones, P.H.; McKenney, J.M.; Grundy, S.M.; Gill, E.A.; Wild, R.A.; et al. National lipid association recommendations for patient-centered management of dyslipidemia: Part 1—Full report. J. Clin. Lipidol. 2015, 9, 129–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collaborators, G.B.D.D. Health effects of dietary risks in 195 countries, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2019, 393, 1958–1972. [Google Scholar] [CrossRef] [Green Version]
- Spector, A.A. Essentiality of fatty acids. Lipids 1999, 34, S1–S3. [Google Scholar] [CrossRef] [PubMed]
- Shetty, S.S.; Kumari, S. Fatty acids and their role in type-2 diabetes. Exp. Ther. Med. 2021, 22, 706. [Google Scholar] [CrossRef]
- Lan, Y.-L.; Lou, J.-C.; Lyu, W.; Zhang, B. Update on the synergistic effect of HSL and insulin in the treatment of metabolic disorders. Ther. Adv. Endocrinol. Metab. 2019, 10, 2042018819877300. [Google Scholar] [CrossRef]
- Galiero, R.; Caturano, A.; Vetrano, E.; Cesaro, A.; Rinaldi, L.; Salvatore, T.; Marfella, R.; Sardu, C.; Moscarella, E.; Gragnano, F.; et al. Pathophysiological mechanisms and clinical evidence of relationship between Nonalcoholic fatty liver disease (NAFLD) and cardiovascular disease. Rev. Cardiovasc. Med. 2021, 22, 755–768. [Google Scholar] [CrossRef]
- Boden, G. Effects of free fatty acids (FFA) on glucose metabolism: Significance for insulin resistance and type 2 diabetes. Exp. Clin. Endocrinol. Diabetes 2003, 111, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Lepretti, M.; Martucciello, S.; Burgos Aceves, M.A.; Putti, R.; Lionetti, L. Omega-3 Fatty Acids and Insulin Resistance: Focus on the Regulation of Mitochondria and Endoplasmic Reticulum Stress. Nutrients 2018, 10, 350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, U.N. A defect in the activity of Δ6 and Δ5 desaturases may be a factor predisposing to the development of insulin resistance syndrome. Prostaglandins Leukot. Essent. Fat. Acids 2005, 72, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Prattichizzo, F.; De Nigris, V.; La Sala, L.; Procopio, A.D.; Olivieri, F.; Ceriello, A. “Inflammaging” as a druggable target: A senescence-associated secretory phenotype—centered view of type 2 diabetes. Oxidative Med. Cell. Longev. 2016, 1810327. [Google Scholar] [CrossRef]
- Behl, T.; Grover, M.; Shah, K.; Makkar, R.; Kaur, L.; Sharma, S.; Gupta, J. Role of omega-3-fatty acids in the management of diabetes and associated complications. In Bioactive Food as Dietary Interventions for Diabetes; Elsevier: Amsterdam, The Netherlands, 2019; pp. 185–192. [Google Scholar]
- Perez-Matute, P.; Perez-Echarri, N.; Martinez, J.A.; Marti, A.; Moreno-Aliaga, M.J. Eicosapentaenoic acid actions on adiposity and insulin resistance in control and high-fat-fed rats: Role of apoptosis, adiponectin and tumour necrosis factor-alpha. Br. J. Nutr. 2007, 97, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar]
- González-Périz, A.; Horrillo, R.; Ferre, N.; Gronert, K.; Dong, B.; Morán-Salvador, E.; Titos, E.; Martínez-Clemente, M.; López-Parra, M.; Arroyo, V.; et al. Obesity-induced insulin resistance and hepatic steatosis are alleviated by ω-3 fatty acids: A role for resolvins and protectins. FASEB J. 2009, 23, 1946–1957. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N.; Chiang, N.; Van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, N.; Zhang, X.; Sugiyama, E.; Kono, H.; Horiuchi, A.; Nakajima, T.; Kanbe, H.; Tanaka, E.; Gonzalez, F.J.; Aoyama, T. Eicosapentaenoic acid improves hepatic steatosis independent of PPARalpha activation through inhibition of SREBP-1 maturation in mice. Biochem. Pharmacol. 2010, 80, 1601–1612. [Google Scholar] [CrossRef]
- Neschen, S.; Morino, K.; Dong, J.; Wang-Fischer, Y.; Cline, G.W.; Romanelli, A.J.; Rossbacher, J.C.; Moore, I.K.; Regittnig, W.; Munoz, D.S.; et al. n-3 Fatty acids preserve insulin sensitivity in vivo in a peroxisome proliferator-activated receptor-alpha-dependent manner. Diabetes 2007, 56, 1034–1041. [Google Scholar] [CrossRef] [Green Version]
- Sato, A.; Kawano, H.; Notsu, T.; Ohta, M.; Nakakuki, M.; Mizuguchi, K.; Itoh, M.; Suganami, T.; Ogawa, Y. Antiobesity effect of eicosapentaenoic acid in high-fat/high-sucrose diet-induced obesity: Importance of hepatic lipogenesis. Diabetes 2010, 59, 2495–2504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jump, D.B. Fatty acid regulation of hepatic lipid metabolism. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 115–120. [Google Scholar] [CrossRef] [Green Version]
- De Caterina, R.; Madonna, R.; Bertolotto, A.; Schmidt, E.B. n-3 fatty acids in the treatment of diabetic patients: Biological rationale and clinical data. Diabetes Care 2007, 30, 1012–1026. [Google Scholar] [CrossRef] [Green Version]
- Watson, H.; Mitra, S.; Croden, F.C.; Taylor, M.; Wood, H.M.; Perry, S.L.; Spencer, J.A.; Quirke, P.; Toogood, G.J.; Lawton, C.L.; et al. A randomised trial of the effect of omega-3 polyunsaturated fatty acid supplements on the human intestinal microbiota. Gut 2018, 67, 1974–1983. [Google Scholar] [CrossRef] [PubMed]
- Rizza, S.; Tesauro, M.; Cardillo, C.; Galli, A.; Iantorno, M.; Gigli, F.; Sbraccia, P.; Federici, M.; Quon, M.J.; Lauro, D. Fish oil supplementation improves endothelial function in normoglycemic offspring of patients with type 2 diabetes. Atherosclerosis 2009, 206, 569–574. [Google Scholar] [CrossRef]
- Wong, C.Y.; Yiu, K.H.; Li, S.W.; Lee, S.; Tam, S.; Lau, C.P.; Tse, H.F. Fish-oil supplement has neutral effects on vascular and metabolic function but improves renal function in patients with Type 2 diabetes mellitus. Diabet. Med. 2010, 27, 54–60. [Google Scholar] [CrossRef]
- Min, Y.; Djahanbakhch, O.; Hutchinson, J.; Bhullar, A.S.; Raveendran, M.; Hallot, A.; Eram, S.; Namugere, I.; Nateghian, S.; Ghebremeskel, K. Effect of docosahexaenoic acid-enriched fish oil supplementation in pregnant women with Type 2 diabetes on membrane fatty acids and fetal body composition--double-blinded randomized placebo-controlled trial. Diabet. Med. A J. Br. Diabet. Assoc. 2014, 31, 1331–1340. [Google Scholar] [CrossRef]
- Ellulu, M.S.; Khaza’ai, H.; Patimah, I.; Rahmat, A.; Abed, Y. Effect of long chain omega-3 polyunsaturated fatty acids on inflammation and metabolic markers in hypertensive and/or diabetic obese adults: A randomized controlled trial. Food Nutr. Res. 2016, 60, 29268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobo-Cejudo, M.G.; Valdes-Ramos, R.; Guadarrama-Lopez, A.L.; Pardo-Morales, R.V.; Martinez-Carrillo, B.E.; Harbige, L.S. Effect of n-3 Polyunsaturated Fatty Acid Supplementation on Metabolic and Inflammatory Biomarkers in Type 2 Diabetes Mellitus Patients. Nutrients 2017, 9, 573. [Google Scholar] [CrossRef] [Green Version]
- Durán, A.M.; Salto, L.M.; Câmara, J.; Basu, A.; Paquien, I.; Beeson, W.L.; Firek, A.; Cordero-MacIntyre, Z.; De León, M. Effects of omega-3 polyunsaturated fatty-acid supplementation on neuropathic pain symptoms and sphingosine levels in Mexican-Americans with type 2 diabetes. Diabetes Metab. Syndr. Obes. Targets Ther. 2019, 12, 109. [Google Scholar] [CrossRef] [Green Version]
- Thota, R.N.; Acharya, S.H.; Garg, M.L. Curcumin and/or omega-3 polyunsaturated fatty acids supplementation reduces insulin resistance and blood lipids in individuals with high risk of type 2 diabetes: A randomised controlled trial. Lipids Health Dis. 2019, 18, 31. [Google Scholar] [CrossRef] [Green Version]
- Usta, M.; Ersoy, A.; Ersoy, C.; Ayar, Y.; Goksel, G.; Saka Karagoz, İ. MO235 effect of omega-3 polyunsaturated fatty acid supplementation on glysemic control and renal function in type 2 diabetic patients with chronic kidney disease. Nephrol. Dial. Transplant. 2021, 36, gfab092.00113. [Google Scholar] [CrossRef]
- Sarbolouki, S.; Javanbakht, M.H.; Derakhshanian, H.; Hosseinzadeh, P.; Zareei, M.; Hashemi, S.B.; Dorosty, A.R.; Eshraghian, M.R.; Djalali, M. Eicosapentaenoic acid improves insulin sensitivity and blood sugar in overweight type 2 diabetes mellitus patients: A double-blind randomised clinical trial. Singap. Med. J. 2013, 54, 387–390. [Google Scholar] [CrossRef]
- Miller, E.R., 3rd; Juraschek, S.P.; Anderson, C.A.; Guallar, E.; Henoch-Ryugo, K.; Charleston, J.; Turban, S.; Bennett, M.R.; Appel, L.J. The effects of n-3 long-chain polyunsaturated fatty acid supplementation on biomarkers of kidney injury in adults with diabetes: Results of the GO-FISH trial. Diabetes Care 2013, 36, 1462–1469. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.M.; Chung, S.H.; Park, Y.; Park, M.K.; Son, Y.K.; Kim, S.E.; An, W.S. Effect of Omega-3 Fatty Acid on the Fatty Acid Content of the Erythrocyte Membrane and Proteinuria in Patients with Diabetic Nephropathy. Int. J. Endocrinol. 2015, 2015, 208121. [Google Scholar] [CrossRef] [Green Version]
- Toorang, F.; Djazayery, A.; Djalali, M. Effects of Omega-3 Fatty Acids Supplement on Antioxidant Enzymes Activity in Type 2 Diabetic Patients. Iran. J. Public Health 2016, 45, 340–345. [Google Scholar]
- Elajami, T.K.; Alfaddagh, A.; Lakshminarayan, D.; Soliman, M.; Chandnani, M.; Welty, F.K. Eicosapentaenoic and Docosahexaenoic Acids Attenuate Progression of Albuminuria in Patients With Type 2 Diabetes Mellitus and Coronary Artery Disease. J. Am. Heart Assoc. 2017, 6, e004740. [Google Scholar] [CrossRef]
- Rajkumar, H.; Mahmood, N.; Kumar, M.; Varikuti, S.R.; Challa, H.R.; Myakala, S.P. Effect of probiotic (VSL#3) and omega-3 on lipid profile, insulin sensitivity, inflammatory markers, and gut colonization in overweight adults: A randomized, controlled trial. Mediat. Inflamm 2014, 2014, 348959. [Google Scholar] [CrossRef] [Green Version]
- Kobyliak, N.; Abenavoli, L.; Falalyeyeva, T.; Mykhalchyshyn, G.; Boccuto, L.; Kyriienko, D.; Kononenko, L.; Komisarenko, I.; Dynnyk, O. Beneficial effects of probiotic combination with omega-3 fatty acids in NAFLD: A randomized clinical study. Minerva Med. 2018, 109, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Kobyliak, N.; Falalyeyeva, T.; Mykhalchyshyn, G.; Molochek, N.; Savchuk, O.; Kyriienko, D.; Komisarenko, I. Probiotic and omega-3 polyunsaturated fatty acids supplementation reduces insulin resistance, improves glycemia and obesity parameters in individuals with type 2 diabetes: A randomised controlled trial. Obes. Med. 2020, 19, 100248. [Google Scholar] [CrossRef]
- de Boer, I.H.; Zelnick, L.R.; Ruzinski, J.; Friedenberg, G.; Duszlak, J.; Bubes, V.Y.; Hoofnagle, A.N.; Thadhani, R.; Glynn, R.J.; Buring, J.E.; et al. Effect of Vitamin D and Omega-3 Fatty Acid Supplementation on Kidney Function in Patients With Type 2 Diabetes: A Randomized Clinical Trial. JAMA 2019, 322, 1899–1909. [Google Scholar] [CrossRef] [PubMed]
- Rajabi-Naeeni, M.; Dolatian, M.; Qorbani, M.; Vaezi, A.A. The effect of co supplementation of omega-3 and vitamin D on cardio metabolic risk factors and psychological distress in reproductive-aged women with prediabetes and hypovitaminosis D: A study protocol for a randomized controlled trial. Trials 2019, 20, 799. [Google Scholar] [CrossRef] [Green Version]
- Limonte, C.P.; Zelnick, L.R.; Ruzinski, J.; Hoofnagle, A.N.; Thadhani, R.; Melamed, M.L.; Lee, I.M.; Buring, J.E.; Sesso, H.D.; Manson, J.E.; et al. Effects of long-term vitamin D and n-3 fatty acid supplementation on inflammatory and cardiac biomarkers in patients with type 2 diabetes: Secondary analyses from a randomised controlled trial. Diabetologia 2021, 64, 437–447. [Google Scholar] [CrossRef]
- Caturano, A.; Acierno, C.; Nevola, R.; Pafundi, P.C.; Galiero, R.; Rinaldi, L.; Salvatore, T.; Adinolfi, L.E.; Sasso, F.C. Non-alcoholic fatty liver disease: From pathogenesis to clinical impact. Processes 2021, 9, 135. [Google Scholar] [CrossRef]
- Zhang, X.; Shen, D.; Fang, Z.; Jie, Z.; Qiu, X.; Zhang, C.; Chen, Y.; Ji, L. Human gut microbiota changes reveal the progression of glucose intolerance. PLoS ONE 2013, 8, e71108. [Google Scholar] [CrossRef]
- Everard, A.; Cani, P.D. Diabetes, obesity and gut microbiota. Best Pract. Research. Clin. Gastroenterol. 2013, 27, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Menni, C.; Zierer, J.; Pallister, T.; Jackson, M.A.; Long, T.; Mohney, R.P.; Steves, C.J.; Spector, T.D.; Valdes, A.M. Omega-3 fatty acids correlate with gut microbiome diversity and production of N-carbamylglutamate in middle aged and elderly women. Sci. Rep. 2017, 7, 11079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchinson, A.N.; Tingo, L.; Brummer, R.J. The Potential Effects of Probiotics and omega-3 Fatty Acids on Chronic Low-Grade Inflammation. Nutrients 2020, 12, 2402. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Sha, L.; Li, K.; Wang, Z.; Wang, T.; Li, Y.; Liu, P.; Dong, X.; Dong, Y.; Zhang, X.; et al. Dietary flaxseed oil rich in omega-3 suppresses severity of type 2 diabetes mellitus via anti-inflammation and modulating gut microbiota in rats. Lipids Health Dis. 2020, 19, 20. [Google Scholar] [CrossRef] [Green Version]
- Onishi, J.C.; Campbell, S.; Moreau, M.; Patel, F.; Brooks, A.I.; Zhou, Y.X.; Haggblom, M.M.; Storch, J. Bacterial communities in the small intestine respond differently to those in the caecum and colon in mice fed low- and high-fat diets. Microbiology 2017, 163, 1189–1197. [Google Scholar] [CrossRef]
- Costantini, L.; Molinari, R.; Farinon, B.; Merendino, N. Impact of Omega-3 Fatty Acids on the Gut Microbiota. Int. J. Mol. Sci. 2017, 18, 2645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shama, S.; Liu, W. Omega-3 fatty acids and gut microbiota: A reciprocal interaction in nonalcoholic fatty liver disease. Dig. Dis. Sci. 2020, 65, 906–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Wang, Y.; Gao, H.; Li, D.; Jiang, R.; Ge, L.; Tong, C.; Xu, K. Associations among Dietary Omega-3 Polyunsaturated Fatty Acids, the Gut Microbiota, and Intestinal Immunity. Mediat. Inflamm. 2021, 2021, 8879227. [Google Scholar] [CrossRef] [PubMed]



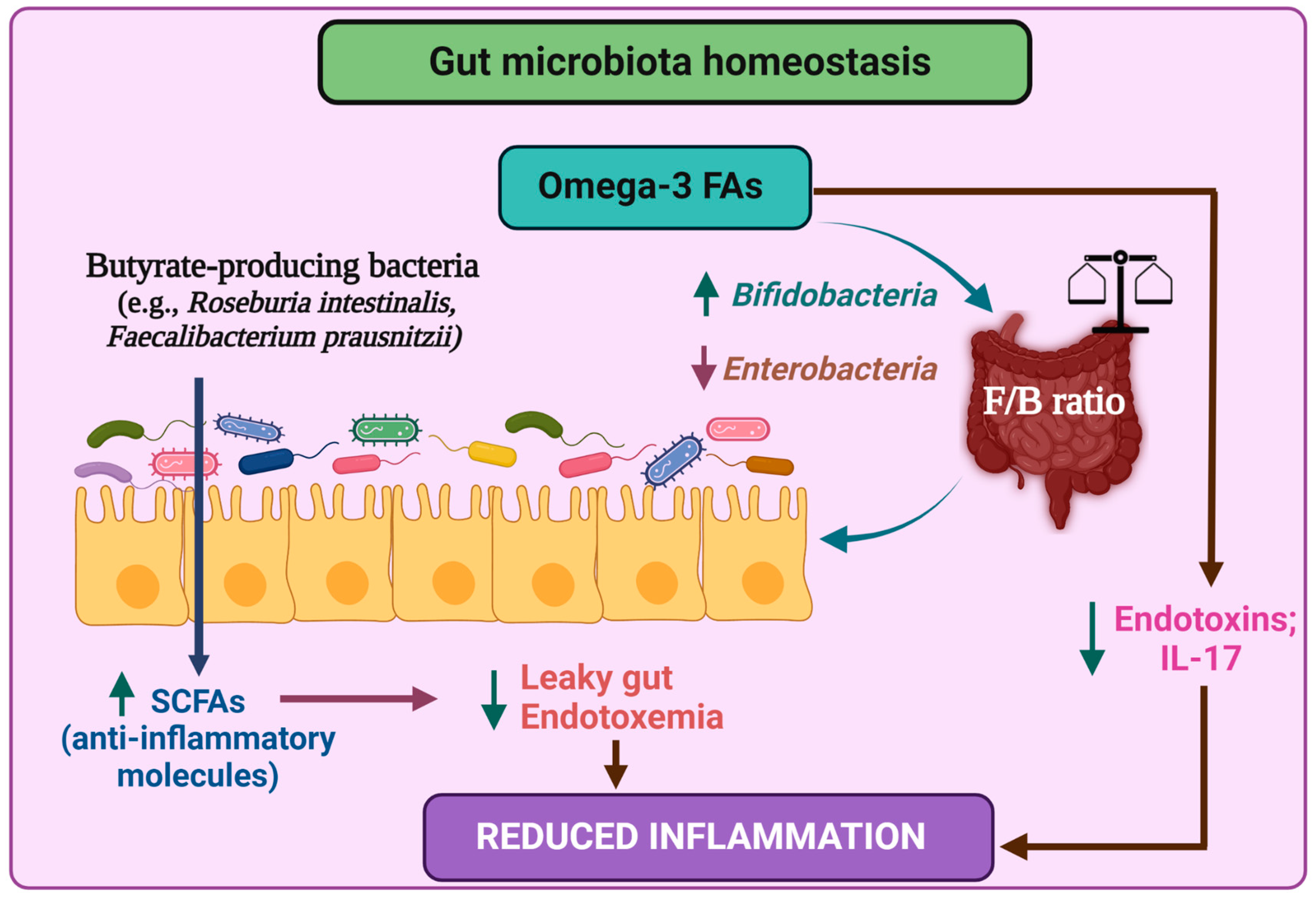{kind=link}
{kind=link}
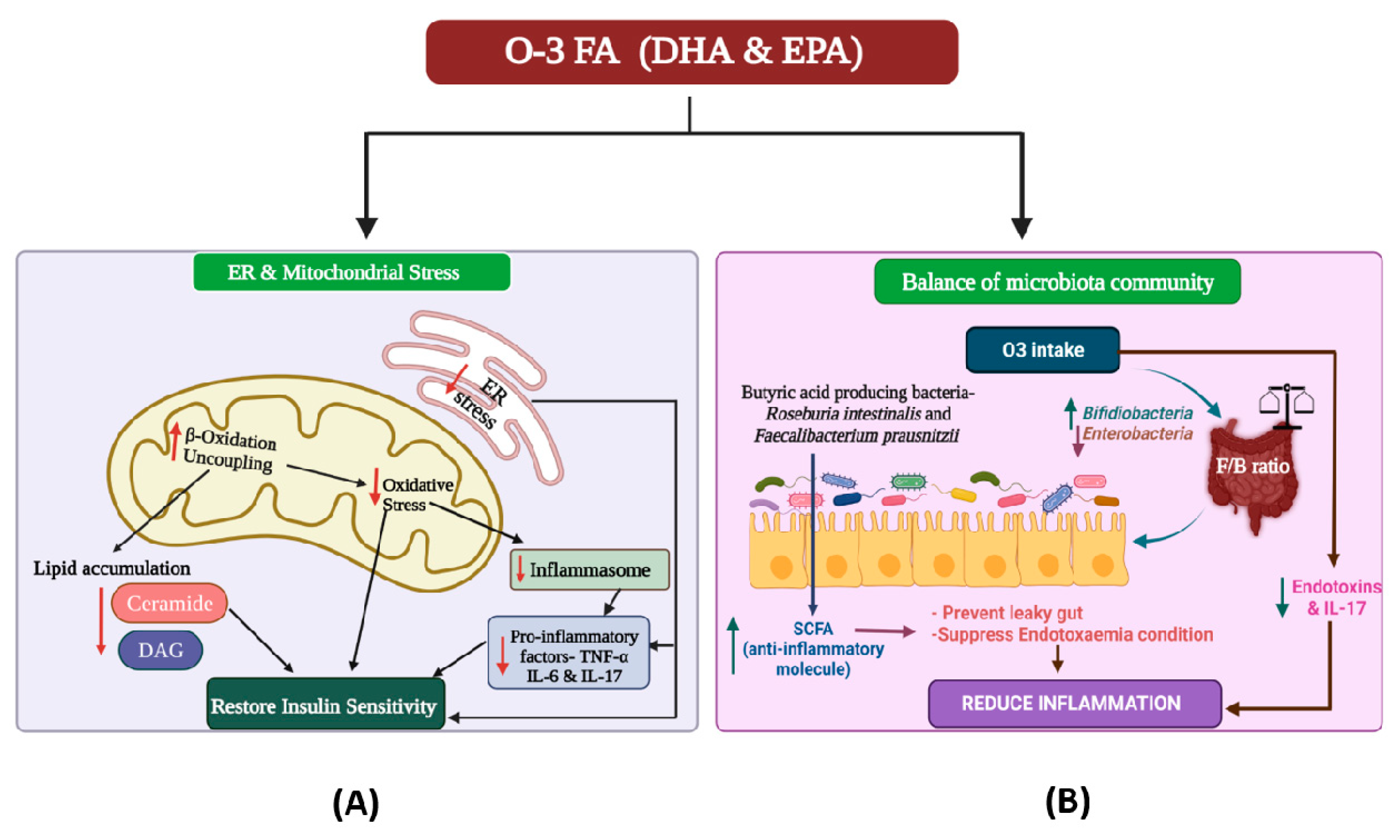{kind=link}
| Human/Animal Models | Intervention/Treatment | Outcome | References |
|---|---|---|---|
| Fish Oil Capsule (EPA & DHA) | |||
| Offspring of T2DM patients with endothelial dysfunction (n = 50) | Dose: 2 g/d Omega-3 PUFA (EPA + DHA); Fish Oil supplement; Duration: 12 weeks |
| [37] |
| T2DM patients without prior CVD (n = 97) | Dose: 4 g/d Fish Oil supplement Duration: 12 weeks |
| [38] |
| Pregnant Women with Type-2 diabetes (n = 88) and healthy women (n = 85) | Dose: 600 mg DHA; Fish Oil supplement Duration: Daily; from early pregnancy till delivery |
| [39] |
| Patients, who are hypertensive and/or Type2 diabetic obese with high levels of inflammatory markers, (n = 64) | Dose: 1.0 g fish oil supplied in soft gel capsules including 300 mg EPA and 200 mg DHA; Duration: 8 weeks |
| [40] |
| T2DM patients (BMI ≤ 29.9), aged 25–60 years, with no other chronic diseases, (n = 65) | Dose: 520 mg of DHA + EPA-enriched fish oil each per day; Duration: 24-weeks |
| [41] |
| T2DM patients (n = 40) | Dose: 100 mg/d DHA & 200 mg/d EPA supplement; Durations: 3 months |
| [42] |
| Individuals with a high risk of developing diabetes or IFG or IGT (n = 64) | Dose- fish oil capsules (1.2 g DHA + EPA) 2 capsules twice a day; Duration: 12 weeks |
| [43] |
| T2DM patients with CKD (n = 25) | Dose: 2 g/d concentrated fish oil; Duration: 3 months |
| [44] |
| Purified O-3 PUFA | |||
| Overweight patients with T2DM (n = 67) | Dose: 2 g purified EPA daily; Duration: 3 months |
| [45] |
| T2DM patients with CKD (n = 31) | Dose: Omega-3 PUFA capsules (EPA + DHA) 4 g/d; Duration: 6 weeks |
| [46] |
| T2DM nephropathic patients (n = 19) | Dose: OMACOR 3 g/d; Duration: 12 weeks |
| [47] |
| T2DM patients (n = 90) | Dose: 2714 mg/d (EPA = 1548 mg, DHA = 828 mg and 338 mg of other omega = 3 fatty acids); Duration: 2 months |
| [48] |
| T2DM with stable coronary artery disease (n = 262) | Dose: 1.86 g/d EPA and 1.5 g/d DHA |
| [49] |
| O-3 PUFA in combination with probiotics | |||
| Overweight (BMI > 25), healthy adults, aged 40–60 years (n = 60) | Dose: One capsule of VSL#3 and purified omega-3 fatty acid (180 mg EPA and 120 mg DHA per capsule) per daily; Duration: 6 weeks |
| [50] |
| Patients with NAFLD (n = 48) | Dose: Symbiter Omega—a live multi-strain probiotic mixture with flax and wheat germ oil containing O-3 FA; once daily; Duration: 8 weeks |
| [51] |
| T2DM patients (n = 54) | Dose: Symbiter Omega—a live multi-strain probiotic mixture with flax and wheat germ oil containing O-3 FA; Duration: 8 weeks |
| [52] |
| O-3 PUFA in combination with Vitamin D | |||
| T2DM patients (n = 1312) | Dose: Vit-D3 2000 IU/d and Omega-3 FA Fish oil supplementation (EPA and DHA) 1 g/d; Duration: 6 h |
| [53] |
| Pre-diabetic with hypervitaminosis D (n = 168 W) | Dose- 1000 mg omega-3 supplement (360 EPA + 240 mg DHA) twice a day + Vit D 50,000 IU every 2 weeks; Duration: 8 weeks |
| [54] |
| T2DM patients (n = 1312) | Dose: Vitamin D and Omacor (EPA + DHA) 1 g/d; Duration: 5 years |
| [55] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, M.; Pal, N.; Sharma, P.; Kumawat, M.; Sarma, D.K.; Nabi, B.; Verma, V.; Tiwari, R.R.; Shubham, S.; Arjmandi, B.; et al. Omega-3 Fatty Acids and Their Interaction with the Gut Microbiome in the Prevention and Amelioration of Type-2 Diabetes. Nutrients 2022, 14, 1723. https://doi.org/10.3390/nu14091723
Kumar M, Pal N, Sharma P, Kumawat M, Sarma DK, Nabi B, Verma V, Tiwari RR, Shubham S, Arjmandi B, et al. Omega-3 Fatty Acids and Their Interaction with the Gut Microbiome in the Prevention and Amelioration of Type-2 Diabetes. Nutrients. 2022; 14(9):1723. https://doi.org/10.3390/nu14091723
Chicago/Turabian StyleKumar, Manoj, Namrata Pal, Poonam Sharma, Manoj Kumawat, Devojit Kumar Sarma, Bilkees Nabi, Vinod Verma, Rajnarayan R. Tiwari, Swasti Shubham, Bahram Arjmandi, and et al. 2022. "Omega-3 Fatty Acids and Their Interaction with the Gut Microbiome in the Prevention and Amelioration of Type-2 Diabetes" Nutrients 14, no. 9: 1723. https://doi.org/10.3390/nu14091723
APA StyleKumar, M., Pal, N., Sharma, P., Kumawat, M., Sarma, D. K., Nabi, B., Verma, V., Tiwari, R. R., Shubham, S., Arjmandi, B., & Nagpal, R. (2022). Omega-3 Fatty Acids and Their Interaction with the Gut Microbiome in the Prevention and Amelioration of Type-2 Diabetes. Nutrients, 14(9), 1723. https://doi.org/10.3390/nu14091723