
Effects of Animal and Vegetable Proteins on Gut Microbiota in Subjects with Overweight or Obesity

Abstract
:1. Introduction
2. Search Strategy
3. The Human Gut Microbiota: Focus on Subjects with Obesity
4. Dietary Patterns Suggested for Weight Loss
5. Effect of Macronutrients on Gut Microbiota
5.1. Carbohydrates
5.2. Fats
5.3. Proteins
6. Effect of Different Nutritional Protocols According to the Amount and the Kind of Proteins on Gut Microbiota
6.1. The Western Diet
6.2. The Mediterranean Diet
6.3. The Plant-Based Diets
6.4. The Very Low-Calorie Ketogenic Diet
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vasileva, L.V.; Marchev, A.S.; Georgiev, M.I. Causes and solutions to “globesity”: The new fa(s)t alarming global epidemic. Food Chem. Toxicol. 2018, 121, 173–193. [Google Scholar] [CrossRef]
- World Health Organization. Obesity and Overweight Fact Sheet. 2018. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 19 January 2023).
- The GBD 2015 Obesity Collaborators. Health effects of overweight and obesity in 195 countries over 25 years. N. Engl. J. Med. 2017, 377, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Li, H. Obesity: Epidemiology, Pathophysiology, and Therapeutics. Front. Endocrinol. 2021, 12, 706978. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [Green Version]
- DeGruttola, A.K.; Low, D.; Mizoguchi, A.; Mizoguchi, E. Current Understanding of Dysbiosis in Disease in Human and Animal Models. Inflamm. Bowel Dis. 2016, 22, 1137–1150. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut Microbes 2018, 9, 308–325. [Google Scholar] [CrossRef] [Green Version]
- Mari, A.; Abu Baker, F.; Mahamid, M.; Sbeit, W.; Khoury, T. The Evolving Role of Gut Microbiota in the Management of Irritable Bowel Syndrome: An Overview of the Current Knowledge. J. Clin. Med. 2020, 9, 685. [Google Scholar] [CrossRef] [Green Version]
- Reddel, S.; Putignani, L.; Del Chierico, F. The Impact of Low-FODMAPs, Gluten-Free, and Ketogenic Diets on Gut Microbiota Modulation in Pathological Conditions. Nutrients 2019, 11, 373. [Google Scholar] [CrossRef] [Green Version]
- Raynor, H.A.; Champagne, C.M. Position of the Academy of Nutrition and Dietetics: Interventions for the Treatment of Overweight and Obesity in Adults. J. Acad. Nutr. Diet. 2016, 116, 129–147. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.; Koh, G. Clinical Evidence and Mechanisms of High-Protein Diet-Induced Weight Loss. J. Obes. Metab. Syndr. 2020, 29, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Astrup, A. The satiating power of protein—A key to obesity prevention? Am. J. Clin. Nutr. 2005, 82, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Tannous dit El Khoury, D.; Obeid, O.; Azar, S.T.; Hwalla, N. Variations in postprandial ghrelin status following ingestion of high-carbohydrate, high-fat, and high-protein meals in males. Ann. Nutr. Metab. 2006, 50, 260–269. [Google Scholar] [CrossRef]
- Moran, L.J.; Luscombe-Marsh, N.D.; Noakes, M.; Wittert, G.A.; Keogh, J.B.; Clifton, P.M. The Satiating Effect of Dietary Protein Is Unrelated to Postprandial Ghrelin Secretion. J. Clin. Endocrinol. Metab. 2005, 90, 5205–5211. [Google Scholar] [CrossRef] [Green Version]
- Soenen, S.; Martens, E.A.; Hochstenbach-Waelen, A.; Lemmens, S.G.; Westerterp-Plantenga, M.S. Normal protein intake is required for body weight loss and weight maintenance, and elevated protein intake for additional preservation of resting energy expenditure and fat free mass. J. Nutr. 2013, 143, 591–596. [Google Scholar] [CrossRef] [Green Version]
- Westerterp-Plantenga, M.S.; Lejeune, M.P.G.M.; Nijs, I.; Van Ooijen, M.; Kovacs, E.M.R. High protein intake sustains weight maintenance after body weight loss in humans. Int. J. Obes. 2004, 28, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Baak, M.A.; Larsen, T.M.; Jebb, S.A.; Martinez, A.; Saris, W.H.M.; Handjieva-Darlenska, T.; Kafatos, A.; Pfeiffer, A.F.H.; Kunešová, M.; Astrup, A. Dietary Intake of Protein from Different Sources and Weight Regain, Changes in Body Composition and Cardiometabolic Risk Factors after Weight Loss: The DIOGenes Study. Nutrients 2017, 9, 1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Debédat, J.; Clément, K.; Aron-Wisnewsky, J. Gut Microbiota Dysbiosis in Human Obesity: Impact of bariatric surgery. Curr. Obes. Rep. 2019, 8, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Weiss, G.A.; Hennet, T. Mechanisms and consequences of intestinal dysbiosis. Cell. Mol. Life Sci. 2017, 74, 2959–2977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E.; Backhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [Green Version]
- Armougom, F.; Henry, M.; Vialettes, B.; Raccah, D.; Raoult, D. Monitoring Bacterial Community of Human Gut Microbiota Reveals an Increase in Lactobacillus in Obese Patients and Methanogens in Anorexic Patients. PLoS ONE 2009, 4, e7125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet—Induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M.; de Vos, W.M. Akkermansia muciniphila gen. nov, sp. nov., a human intestinal mucin-degradind bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54 Pt 5, 1469–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwiertz, A.; Taras, D.; Schäfer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in lean and overweight healthy subjects. Obesity 2010, 18, 190–195. [Google Scholar] [CrossRef]
- Winer, D.A.; Luck, H.; Tsai, S.; Winer, S. The intestinal immune system in obesity and insulin resistance. Cell Metab. 2016, 23, 413–426. [Google Scholar] [CrossRef] [Green Version]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Amar, J.; Inglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [Green Version]
- Standard Italiani per la Cura Dell’obesità SIO-ADI 2016–2017; Tipografia Ceccarelli: Acquapendente, Italy, 2017.
- Verreijen, A.M.; Engberink, M.F.; Memelink, R.G.; van der Plas, S.E.; Visser, M.; Weijs, P.J. Effect of a high protein diet and/or resistance exercise on the preservation of fat free mass during weight loss in overweight and obese older adults: A randomized controlled trial. Nutr. J. 2017, 16, 10. [Google Scholar] [CrossRef] [Green Version]
- Campos-Nonato, I.; Hernandez, L.; Barquera, S. Effect of a High-Protein Diet versus Standard-Protein Diet on Weight Loss and Biomarkers of Metabolic Syndrome: A Randomized Clinical Trial. Obes. Facts 2017, 10, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Chao, A.M.; Quigley, K.M.; Wadden, T.A. Dietary interventions for obesity: Clinical and mechanistic findings. J. Clin. Investig. 2021, 131, e140065. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.T.; Luo, Y.; Zhang, Y.; Zhou, Y.; Zhao, T.T. High protein diet is of benefit for patients with type 2 diabetes: An updated meta-analysis. Medicine 2018, 97, e13149. [Google Scholar] [CrossRef] [PubMed]
- Di Rosa, C.; Lattanzi, G.; Taylor, S.F.; Manfrini, S.; Khazrai, Y.M. Very low calorie ketogenic diets in overweight and obesity treatment: Effects on anthropometric parameters, body composition, satiety, lipid profile and microbiota. Obes. Res. Clin. Pract. 2020, 14, 491–503. [Google Scholar] [CrossRef]
- Pezzana, A.; Amerio, M.L.; Fatati, G.; Caregaro Negrin, L.; Muratori, F.; Rovera, G.M.; Zanardi, M. La dieta chetogenica—Fondazione ADI: Position Paper. ADI 2014, 6, 38–43. [Google Scholar]
- Muscogiuri, G.; Barrea, L.; Laudisio, D.; Pugliese, G.; Salzano, C.; Savastano, S.; Colao, A. The management of very low-calorie ketogenic diet in obesity outpatient clinic: A practical guide. J. Transl. Med. 2019, 17, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caprio, M.; Infante, M.; Moriconi, E.; Armani, A.; Fabbri, A.; Mantovani, G.; Mariani, S.; Lubrano, C.; Poggiogalle, E.; Migliaccio, S.; et al. Very-low-calorie ketogenic diet (VLCKD) in the management of metabolic diseases: Systematic review and consensus statement from the Italian Society of Endocrinology (SIE). J. Endocrinol. Investig. 2019, 45, 1365–1386. [Google Scholar] [CrossRef] [PubMed]
- Bujnowski, D.; Xun, P.; Daviglus, M.L.; Van Horn, L.; He, K.; Stamler, J. Longitudinal Association between Animal and Vegetable Protein Intake and Obesity among Men in the United States: The Chicago Western Electric Study. J. Am. Diet. Assoc. 2011, 111, 1150–1155.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halkjær, J.; Olsen, A.; Overvad, K.; Jakobsen, M.U.; Boeing, H.; Buijsse, B.; Palli, D.; Tognon, G.; Du, H.; van der A, D.L.; et al. Intake of total, animal and plant protein and subsequent changes in weight or waist circumference in European men and women: The Diogenes project. Int. J. Obes. 2011, 35, 1104–1113. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Mouratidou, T.; Vereecken, C.; Kersting, M.; Bolca, S.; de Morales, A.C.F.; Cuenca-Garcìa, M.; Moreno, L.-A.; Gonzales-Gross, M.; Valtueña, J.; et al. Dietary animal and plant protein intakes and their associations with obesity and cardio-metabolic indicators in European adolescents: The HELENA cross-sectional study. Nutr. J. 2015, 14, 10. [Google Scholar] [CrossRef] [Green Version]
- Zhubi-Bakija, F.; Bajraktari, G.; Bytyçi, I.; Mikhailidis, D.P.; Henein, M.Y.; Latkovskis, G.; Rexhaj, Z.; Zhubi, E.; Banach, M.; Alnouri, F.; et al. The impact of type of dietary protein, animal versus vegetable, in modifying cardiometabolic risk factors: A position paper from the International Lipid Expert Panel (ILEP). Clin. Nutr. 2021, 40, 255–276. [Google Scholar] [CrossRef]
- Patterson, E.; Ryan, P.M.; Cryan, J.F.; Dinan, T.G.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Gut microbiota, obesity and diabetes. Postgrad. Med. J. 2016, 92, 286–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, K.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parvin, S.; Easmin, D.; Sheikh, A.; Biswas, M.; Sharma, S.C.D.; Jahan, G.S.; Islam, A.; Roy, N.; Shovon, M.S. Nutritional Analysis of Date Fruits (Phoenix dactylifera L.) in Perspective of Bangladesh. Am. J. Life Sci. 2015, 3, 274. [Google Scholar] [CrossRef] [Green Version]
- Eid, N.; Enani, S.; Walton, G.; Corona, G.; Costabile, A.; Gibson, G.; Rowland, I.; Spencer, J.P.E. The impact of date palm fruits and their component polyphenols, on gut microbial ecology, bacterial metabolites and colon cancer cell proliferation. J. Nutr. Sci. 2014, 3, e46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francavilla, R.; Calasso, M.; Calace, L.; Siragusa, S.; Ndagijimana, M.; Vernocchi, P.; Brunetti, L.; Mancino, G.; Tedeschi, G.; Guerzoni, E.; et al. Effect of lactose on gut microbiota and metabolome of infants with cow’s milk allergy. Pediatr. Allergy Immunol. 2012, 23, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R. Insulin signaling in the control of glucose and lipid homeostasis. Handb. Exp. Pharmacol. 2016, 233, 51–71. [Google Scholar] [PubMed]
- de Vrese, M.; Schrezenmeir, J. Probiotics, prebiotics, and synbiotics. Adv. Biochem. Eng. Biotechnol. 2008, 111, 1–66. [Google Scholar]
- Makki, K.; Deehan, E.C.; Walter, J.; Bäckhed, F. The Impact of Dietary Fiber on Gut Microbiota in Host Health and Disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Fuller, S.; Beck, E.; Salman, H.; Tapsell, L. New Horizons for the Study of Dietary Fiber and Health: A Review. Plant Foods Hum. Nutr. 2016, 71, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cotillard, A.; Kennedy, S.P.; Kong, L.C.; Prifti, E.; Pons, N.; Le Chatelier, E.; Almeida, M.; Quinquis, B.; Levenez, F.; Galleron, N.; et al. Dietary intervention impact on gut microbial gene richness. Nature 2013, 500, 585–588. [Google Scholar] [CrossRef] [PubMed]
- 2015–2020 DGfA. Nutritional Goals for Age-Sex Groups Based on Dietary Reference Intakes and Dietary Guidelines Recommendations. Available online: https://health.gov/dietaryguidelines/2015/guidelines/appendix-7 (accessed on 20 May 2023).
- Hawkins, A.T.; Wise, P.E.; Chan, T.; Lee, J.T.; Glyn, T.; Wood, V.; Eglinton, T.; Frizelle, F.; Khan, A.; Hall, J.; et al. Diverticulitis: An Update From the Age Old Paradigm. Curr. Probl. Surg. 2020, 57, 100862. [Google Scholar] [CrossRef]
- Costabile, A.; Klinder, A.; Fava, F.; Napolitano, A.; Fogliano, V.; Leonard, C.; Gibson, G.R.; Tuohy, K. Whole-grain wheat breakfast cereal has a prebiotic effect on the human gut microbiota: A double-blind, placebo-controlled, crossover study. Br. J. Nutr. 2008, 99, 110–120. [Google Scholar] [CrossRef]
- Carvalho-Wells, A.L.; Helmolz, K.; Nodet, C.; Molzer, C.; Leonard, C.; McKevith, B.; Thielecke, F.; Jackson, K.G.; Tuohy, K.M. Determination of the in vivo prebiotic potential of a maize-based whole grain breakfast cereal: A human feeding study. Br. J. Nutr. 2010, 104, 1353–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- So, D.; Whelan, K.; Rossi, M.; Morrison, M.; Holtmann, G.; Kelly, J.T.; Shanahan, E.R.; Staudacher, H.M.; Campbell, K.L. Dietary fiber intervention on gut microbiota composition in healthy adults: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2018, 107, 965–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birkeland, E.; Gharagozlian, S.; Birkeland, K.I.; Valeur, J.; Måge, I.; Rud, I.; Aas, A.-M. Prebiotic effect of inulin-type fructans on faecal microbiota and short-chain fatty acids in type 2 diabetes: A randomised controlled trial. Eur. J Nutr. 2020, 59, 3325–3338. [Google Scholar] [CrossRef] [PubMed]
- Sheflin, A.; Borresen, E.C.; Kirkwood, J.S.; Boot, C.M.; Whitney, A.; Lu, S.; Brown, R.J.; Broeckling, C.; Ryan, E.P.; Weir, T.L. Dietary supplementation with rice bran or navy bean alters gut bacterial metabolism in colorectal cancer survivors. Mol. Nutr. Food Res. 2017, 61, 1500905. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.W.; Ince, J.; Duncan, S.H.; Webster, L.M.; Holtrop, G.; Ze, X.; Brown, D.; Stares, M.D.; Scott, P.; Bergerat, A.; et al. Dominant and diet-responsive groups of bacteria within the human colonic microbiota. ISME J. 2011, 5, 220–230. [Google Scholar] [CrossRef]
- Keim, N.L.; Martin, R.J. Dietary Whole Grain–Microbiota Interactions: Insights into Mechanisms for Human Health. Adv. Nutr. Int. Rev. J. 2014, 5, 556–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leitch, E.C.M.; Walker, A.W.; Duncan, S.H.; Holtrop, G.; Flint, H.J. Selective colonization of insoluble substrates by human faecal bacteria. Environ. Microbiol. 2007, 9, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Abell, G.C.; Cooke, C.M.; Bennett, C.N.; Conlon, M.A.; McOrist, A.L. Phylotypes related to Ruminococcus bromii are abundant in the large bowel of humans and increase in response to a diet high in resistant starch. FEMS Microbiol. Ecol. 2008, 66, 505–515. [Google Scholar] [CrossRef] [Green Version]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fava, F.; Gitau, R.; Griffin, B.A.; Gibson, G.R.; Tuohy, K.M.; Lovegrove, J.A. The type and quantity of dietary fat and carbohydrate alter faecal microbiome and short-chain fatty acid excretion in a metabolic syndrome ‘at-risk’ population. Int. J. Obes. 2013, 37, 216–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, B.S.; Weisburger, J.H.; Wynder, E.L. Effects of High Risk and Low Risk Diets for Colon Carcinogenesis on Fecal Microflora and Steroids in Man. J. Nutr. 1975, 105, 878–884. [Google Scholar] [CrossRef] [PubMed]
- Teodoro, J.S.; Varela, A.T.; Rolo, A.P.; Palmeira, C.M. High-fat and obesogenic diets: Current and future strategies to fight obesity and diabetes. Genes Nutr. 2014, 9, 406. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Wang, F.; Yuan, J.; Li, J.; Jiang, D.; Zhang, J.; Li, H.; Wang, R.; Tang, J.; Huang, T.; et al. Effects of dietary fat on gut microbiota and faecal metabolites, and their relationship with cardiometabolic risk factors: A 6-month randomised controlled-feeding trial. Gut 2019, 68, 1417–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajkumar, H.; Mahmood, N.; Kumar, M.; Varikuti, S.R.; Challa, H.R.; Myakala, S.P. Effect of Probiotic (VSL#3) and Omega-3 on Lipid Profile, Insulin Sensitivity, Inflammatory Markers, and Gut Colonization in Overweight Adults: A Randomized, Controlled Trial. Mediat. Inflamm. 2014, 2014, 348959. [Google Scholar] [CrossRef] [Green Version]
- Balfegó, M.; Canivell, S.; Hanzu, F.A.; Sala-Vila, A.; Martínez-Medina, M.; Murillo, S.; Mur, T.; Ruano, E.G.; Linares, F.; Porras, N.; et al. Effects of sardine-enriched diet on metabolic control, inflammation and gut microbiota in drug-naïve patients with type 2 diabetes: A pilot randomized trial. Lipids Health Dis. 2016, 15, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jian, C.; Luukkonen, P.; Sädevirta, S.; Yki-Järvinen, H.; Salonen, A. Impact of short-term overfeeding of saturated or unsaturated fat or sugars on the gut microbiota in relation to liver fat in obese and overweight adults. Clin. Nutr. 2021, 40, 207–216. [Google Scholar] [CrossRef]
- Vijay, A.; Astbury, S.; Le Roy, C.; Spector, T.D.; Valdes, A.M. The prebiotic effects of omega-3 fatty acid supplementation: A six-week randomised intervention trial. Gut Microbes 2021, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bratlie, M.; Hagen, I.V.; Helland, A.; Erchinger, F.; Midttun, Ø.; Ueland, P.M.; Rosenlund, G.; Sveier, H.; Mellgren, G.; Hausken, T.; et al. Effects of high intake of cod or salmon on gut microbiota profile, faecal output and serum concentrations of lipids and bile acids in overweight adults: A randomised clinical trial. Eur. J. Nutr. 2021, 60, 2231–2248. [Google Scholar] [CrossRef]
- Telle-Hansen, V.H.; Gaundal, L.; Bastani, N.; Rud, I.; Byfuglien, M.G.; Gjøvaag, T.; Retterstøl, K.; Holven, K.B.; Ulven, S.M.; Myhrstad, M.C.W. Replacing saturated fatty acids with polyunsaturated fatty acids increases the abundance of Lachnospiraceae and is associated with reduced total cholesterol levels—A randomized controlled trial in healthy individuals. Lipids Health Dis. 2022, 21, 92. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, S.F.; Murphy, E.F.; O’Sullivan, O.; Lucey, A.J.; Humphreys, M.; Hogan, A.; Hayes, P.; O’Reilly, M.; Jeffery, I.B.; Wood-Martin, R.; et al. Exercise and associated dietary extremes impact on gut microbial diversity. Gut 2014, 63, 1913–1920. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.H.; Park, J.; Kim, M. Gut Microbiota-Derived Short-Chain Fatty Acids, T Cells, and Inflammation. Immune Netw. 2014, 14, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Świątecka, D.; Dominika, Ś.; Narbad, A.; Arjan, N.; Ridgway, K.P.; Karyn, R.P. The study on the impact of glycated pea proteins on human intestinal bacteria. Int. J. Food Microbiol. 2011, 145, 267–272. [Google Scholar] [PubMed]
- Beaumont, M.; Portune, K.J.; Steuer, N.; Lan, A.; Cerrudo, V.; Audebert, M.; Dumont, F.; Mancano, G.; Khodorova, N.; Andriamihaja, M.; et al. Quantity and source of dietary protein influence metabolite production by gut microbiota and rectal mucosa gene expression: A randomized, parallel, double-blind trial in overweight humans. Am. J. Clin. Nutr. 2017, 106, 1005–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hentges, D.J.; Maier, B.R.; Burton, G.C.; Flynn, M.A.; Tsutakawa, R.K. Effect of a high-beef diet on the fecal bacterial flora of humans. Cancer Res. 1977, 37, 568–571. [Google Scholar] [PubMed]
- Romond, M.-B.; Ais, A.; Guillemot, F.; Bounouader, R.; Cortot, A.; Romond, C. Cell-Free Whey from Milk Fermented with Bifidobacterium breve C50 Used to Modify the Colonic Microflora of Healthy Subjects. J. Dairy Sci. 1998, 81, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Ling, C.; Liu, L.; Zhang, J.; Wang, J.; Tong, X.; Hidayat, K.; Chen, M.; Chen, X.; Zhou, H.; et al. Effects of Whey Protein or Its Hydrolysate Supplements Combined with an Energy-Restricted Diet on Weight Loss: A Randomized Controlled Trial in Older Women. Nutrients 2022, 14, 4540. [Google Scholar] [CrossRef]
- Reimer, R.A.; Willis, H.J.; Tunnicliffe, J.M.; Park, H.; Madsen, K.L.; Soto-Vaca, A. Inulin-type fructans and whey protein both modulate appetite but only fructans alter gut microbiota in adults with overweight/obesity: A randomized controlled trial. Mol. Nutr. Food Res. 2017, 61, 1700484. [Google Scholar] [CrossRef]
- Mitchell, S.M.; McKenzie, E.J.; Mitchell, C.J.; Milan, A.M.; Zeng, N.; D’Souza, R.F.; Ramzan, F.; Sharma, P.; Rettedal, E.; Knowles, S.O.; et al. A period of 10 weeks of increased protein consumption does not alter faecal microbiota or volatile metabolites in healthy older men: A randomised controlled trial. J. Nutr. Sci. 2020, 9, e25. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.R.; Gratz, S.W.; Duncan, S.H.; Holtrop, G.; Ince, J.; Scobbie, L.; Duncan, G.; Johnstone, A.M.; Lobley, G.E.; Wallace, R.J.; et al. High-protein, reduced-carbohydrate weight-loss diets promote metabolite profiles likely to be detrimental to colonic health. Am. J. Clin. Nutr. 2011, 93, 1062–1072. [Google Scholar] [CrossRef] [Green Version]
- Machiels, K.; Joossens, M.; Sabino, J.; De Preter, V.; Arijs, I.; Eeckhaut, V.; Ballet, V.; Claes, K.; Van Immerseel, F.; Verbeke, K.; et al. A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut 2014, 63, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, C.L.J.; Önnerfält, J.; Xu, J.; Molin, G.; Ahrné, S.; Thorngren-Jerneck, K. The Microbiota of the Gut in Preschool Children With Normal and Excessive Body Weight. Obesity 2012, 20, 2257–2261. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [Green Version]
- Park, J.-E.; Seo, J.-E.; Lee, J.-Y.; Kwon, H. Distribution of Seven N-Nitrosamines in Food. Toxicol. Res. 2015, 31, 279–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Keefe, S.J.D.; Li, J.V.; Lahti, L.; Ou, J.; Carbonero, F.; Mohammed, K.; Posma, J.M.; Kinross, J.; Wahl, E.; Ruder, E.; et al. Fat, fibre and cancer risk in African Americans and rural Africans. Nat. Commun. 2015, 6, 6342. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Lacoba, R.; Pardo-Garcia, I.; Amo-Saus, E.; Escribano-Sotos, F. Mediterranean diet and health outcomes: A systematic meta-review. Eur. J. Public Health 2018, 28, 955–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Legarrea, P.; Fuller, N.R.; Zulet, M.A.; Martinez, J.A.; Caterson, I.D. The influence of Mediterranean, carbohydrate and high protein diets on gut microbiota composition in the treatment of obesity and associated inflammatory state. Asia Pac. J. Clin. Nutr. 2014, 23, 360–368. [Google Scholar] [PubMed]
- Rinott, E.; Meir, A.Y.; Tsaban, G.; Zelicha, H.; Kaplan, A.; Knights, D.; Tuohy, K.; Scholz, M.U.; Koren, O.; Stampfer, M.J.; et al. The effects of the Green-Mediterranean diet on cardiometabolic health are linked to gut microbiome modifications: A randomized controlled trial. Genome Med. 2022, 14, 29. [Google Scholar] [CrossRef]
- Pagliai, G.; Russo, E.; Niccolai, E.; Dinu, M.; Di Pilato, V.; Magrini, A.; Bartolucci, G.; Baldi, S.; Menicatti, M.; Giusti, B.; et al. Influence of a 3-month low-calorie Mediterranean diet compared to the vegetarian diet on human gut microbiota and SCFA: The CARDIVEG Study. Eur. J. Nutr. 2020, 59, 2011–2024. [Google Scholar] [CrossRef]
- Donohoe, D.R.; Collins, L.B.; Wali, A.; Bigler, R.; Sun, W.; Bultman, S.J. The Warburg effect dictates the mechanism of butyrate-mediated histone acetylation and cell proliferation. Mol. Cell. 2012, 48, 612–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez-Díaz, I.; Fernández-Navarro, T.; Sánchez, B.; Margolles, A.; González, S. Mediterranean diet and faecal microbiota: A transversal study. Food Funct. 2016, 7, 2347–2356. [Google Scholar] [CrossRef] [PubMed]
- Meslier, V.; Laiola, M.; Roager, H.M.; De Filippis, F.; Roume, H.; Quinquis, B.; Giacco, R.; Mennella, I.; Ferracane, R.; Pons, N.; et al. Mediterranean diet intervention in overweight and obese subjects lowers plasma cholesterol and causes changes in the gut microbiome and metabolome independently of energy intake. Gut 2020, 69, 1258–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahleova, H.; Rembert, E.; Alwarith, J.; Yonas, W.N.; Tura, A.; Holubkov, R.; Agnello, M.; Chutkan, R.; Barnard, N.D. Effects of a Low-Fat Vegan Diet on Gut Microbiota in Overweight Individuals and Relationships with Body Weight, Body Composition, and Insulin Sensitivity. A Randomized Clinical Trial. Nutrients 2020, 12, 2917. [Google Scholar] [CrossRef]
- Koliada, A.; Syzenko, G.; Moseiko, V.; Budovska, L.; Puchkov, K.; Perederiy, V.; Gavalko, Y.; Dorofeyev, A.; Romanenko, M.; Tkach, S.; et al. Association between body mass index and Firmicutes/Bacteroidetes ratio in an adult Ukrainian population. BMC Microbiol. 2017, 17, 120. [Google Scholar] [CrossRef] [Green Version]
- Jumpertz, R.; Le, D.S.; Turnbaugh, P.J.; Trinidad, C.; Bogardus, C.; Gordon, J.I.; Krakoff, J. Energy-balance studies reveal associations between gut microbes, caloric load, and nutrient absorption in humans123. Am. J. Clin. Nutr. 2011, 94, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez-Repiso, C.; Hernandez-Garcia, C.; Garcia-Almeida, J.M.; Bellido, D.; Martin-Nuñez, G.M.; Sanchez-Alcoholado, L.; Alcaide- Torres, J.; Sajoux, I.; Tinahones, F.J.; Moreno-Indias, I. Effect of synbiotic supplementation in a very- low-Calorie ketogenic diet on weight loss achievement and gut microbiota: A randomized controlled pilot study. Mol. Nutr. Food Res. 2019, 63, e1900167. [Google Scholar] [CrossRef]
- Basciani, S.; Camajani, E.; Contini, S.; Persichetti, A.; Risi, R.; Bertoldi, L.; Strigari, L.; Prossomariti, G.; Watanabe, M.; Mariani, S.; et al. Very-Low-Calorie Ketogenic Diets With Whey, Vegetable, or Animal Protein in Patients With Obesity: A Randomized Pilot Study. J. Clin. Endocrinol. Metab. 2020, 105, 2939–2949. [Google Scholar] [CrossRef]
- Società Italiana di Nutrizione Umana-SINU, Livelli di Assunzione di Riferimento per la Popolazione Italiana, LARN 2014. Available online: https://sinu.it/2019/07/09/proteine/ (accessed on 25 February 2023).
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Odamaki, T.; Sugahara, H.; Yonezawa, S.; Yaeshima, T.; Iwatsuki, K.; Tanabe, S.; Tominaga, T.; Togashi, H.; Benno, Y.; Xiao, J.-Z. Effect of the oral intake of yogurt containing Bifidobacterium longum BB536 on the cell numbers of enterotoxigenic Bacteroides fragilis in microbiota. Anaerobe 2012, 18, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Remely, M.; Tesar, I.; Hippe, B.; Gnauer, S.; Rust, P.; Haslberger, A. Gut microbiota composition correlates with changes in body fat content due to weight loss. Benef. Microbes 2015, 6, 431–439. [Google Scholar] [CrossRef] [PubMed]


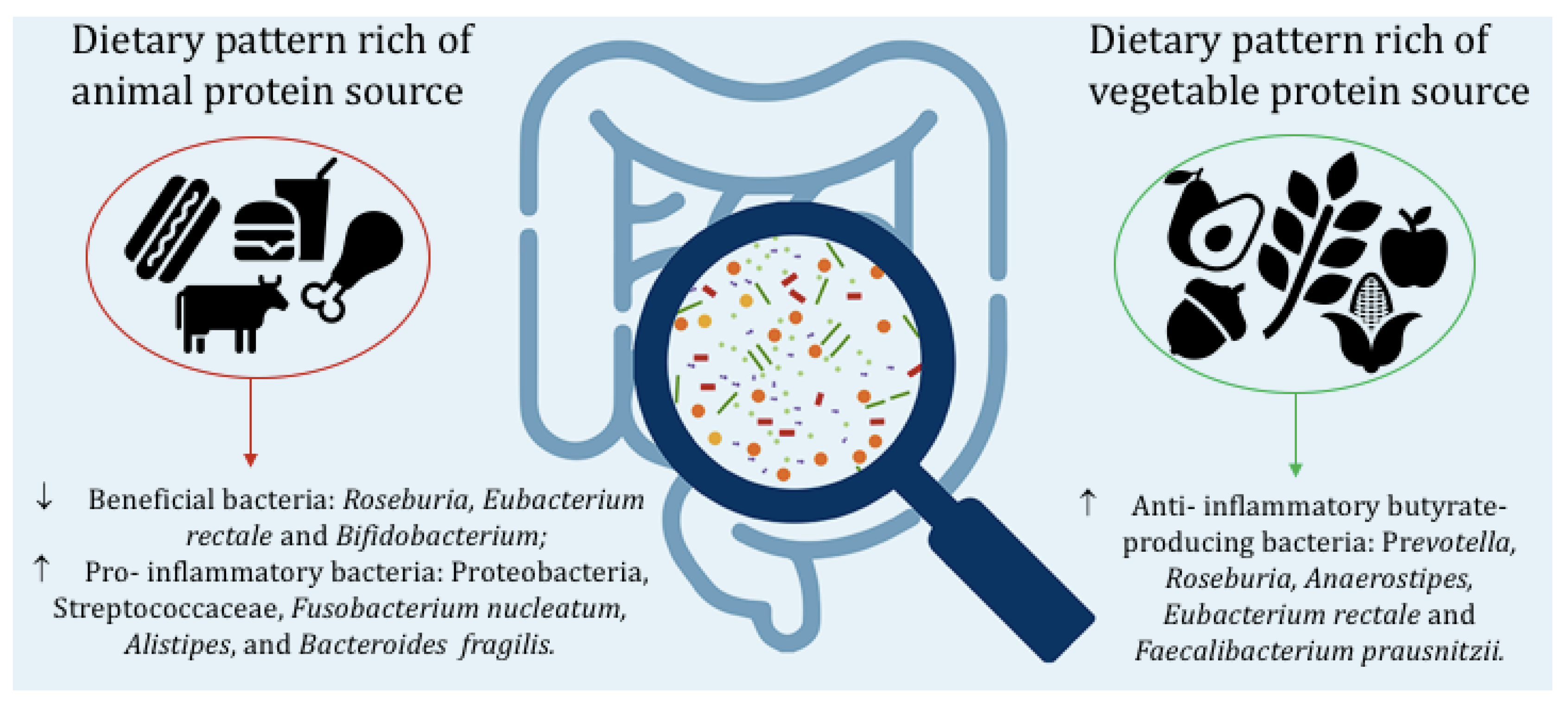{kind=link}
| Study | Population | Dietary Effects | Results |
|---|---|---|---|
| Costabile et al., 2008 [55] | 32 healthy volunteers with BMI between 20 and 30 kg/m2 | Consumption of either 48 g of whole grain (WG) or wheat bran (WB) for two 3-week periods, separated by a 2-week washout period | WG: ⇑ Bifidobacteria and lactobacilli No differences in SCFAs between the two groups |
| Cotillard et al., 2013 [52] | 38 with obesity and 11 overweight subjects | 12 weeks of energy-restricted high protein diet with 35% protein, 25% lipids, and 44% carbohydrates | Dietary intervention: ⇑ gene richness ⇓ Bifidobacterium and Eubacterium rectale. |
| Birkeland et al., 2020 [58] | 25 subjects with BMI < 40 kg/m2 affected by type II diabetes | 16 g of inulin-type fructans (a mixture of oligofructose and inulin) and 16 g placebo (maltodextrin) for 6 weeks in randomized order | The inuline-type fructans: ⇑ SCFAs ⇑ Bifidobacterium adolescentis |
| Sheflin et al., 2017 [59] | 29 overweight and with obesity volunteers | Consumption of a snack with fiber-rich, stabilized rice bran (30 g/day), or cooked navy bean powder (35 g/day) for 28 days | Supplementation with rice bran: ⇓ the Firmicutes to Bacteroidetes ratio ⇑ SCFAs |
| Walker et al., 2011 [60] | 14 overweight men | Volunteers were provided successively with a control diet, diets high in resistant starch (RS) (26 g/day of resistant starch from type III resistant starch), or non-starch polysaccharides (NSPs) (42 g/day of total non-starch polysaccharides from wheat bran) and a reduced carbohydrate weight loss (WL) diet over 10 weeks | RS diet: ⇑ Ruminococcus bromii (17%) of total bacteria compared to the 3.8% of the NSP diet ⇑ Eubacterium rectale (10.1%) RS and WL diets: ⇑ Oscillibacter WL: ⇓ Eubacterium and Collinsella aerofaciens |
| Abell et al., 2008 [63] | 46 overweight volunteers | One group received 4 weeks of a RS2—rich diet (22 g/day of RS) and the other one received 4 weeks of a RS2—low diet (1 g/day of RS). | The RS2-rich diet group showed ⇑ Ruminococcus bromii, Faecalibacterium prausnitzii and Eubacterium rectale |
| Study | Population | Dietary Effects | Results |
|---|---|---|---|
| Wan et al., 2019 [69] | 217 overweight subjects | 6 months of either a high-fat (40%) or a low-fat (20%) diet | The high-fat diet: ⇑ Alistipes, Bacteroides and the LPS ⇓ Faecalibacterium and total SCFAs |
| Fava et al., 2013 [66] | 88 overweight subjects | Low-fat (28% fat) diet and a high saturated fat (38% fat of which 18% was from saturated fats) diet for 4 weeks | Low-fat diet: ⇑ fecal abundance of Bifidobacterium High saturated fat diet: ⇑ Faecalibacterium prausnitzii |
| Rajkumar et al., 2014 [70] | 60 overweight adults | 1 capsule daily of omega-3 polyunsaturated (PUFA) supplementation (containing 180 mg of EPA and 120 mg of DHA per capsule) for 6 weeks | Did not lead to a significative change on gut microbiota composition |
| Balfegò et al., 2016 [71] | 35 overweight and with obesity patients | One group followed a standard diet and the other one a diet enriched with 100 g of sardines for 5 days/week for 6 months | The group that followed the enriched diet, compared to the baseline showed: ⇓ in the Firmicutes to Bacteroidetes ratio ⇑ in the Bacteroides to Prevotella ratio |
| Jian et al., 2021 [72] | 38 overweight and with obesity subjects | Excess of 1000 kcal/day in a diet rich in either saturated (86% of total daily energy from fat, of which 76% were saturated) or unsaturated fats (91% of total daily energy from fat, of which 79% were unsaturated) | Overfeeding of saturated fats: ⇑ Proteobacteria Overfeeding of unsaturated fats: ⇑ butyrate producers bacteria |
| Vijay et al., 2021 [73] | 69 subjects who had a BMI comprised between 20 and 39.9 kg/m2 | 6-week dietary intervention with a supplementation of either 500 mg of omega-3 or 20 g of inulin | The omega-3 supplementation resulted in: ⇑ Coprococcus spp. and Bacteroides spp. ⇓ Collinsella spp (fatty-liver associated) |
| Bratlie et al., 2021 [74] | 76 adults with overweight/obesity | Intake of salmon, cod (both in the quantity of 750 g/week) or the total exclusion of fish (control group) for 8 weeks | Salmon and Cod groups determined: ⇓ abundance of Bacteroidetes, Clostridiales; ⇑ abundance of the Selenomonadales. |
| Telle-Hansen et al., 2022 [75] | 17 humans with a BMI between 18.5 and 27 kg/m2 | Consumption of products with saturated fatty acids (SFAs) (29.9 g of SFAs/day) or polyunsaturated fatty acids (PUFAs) (26.4 g of PUFAs/day) for 3 days | The intervention with PUFAs: ⇑ Lachnospiraceae and Bifidobacterium spp. |
| Study | Population | Dietary Intervention | Results |
|---|---|---|---|
| Beaumont et al., 2017 [80] | 38 overweight subjects | 3 weeks of 15% of total daily energy supplementation with soy protein, casein, or maltodextrins as control | No significant differences in bacterial composition between the intervention groups but supplementation (of soy or casein) can increase the luminal concentrations of both beneficial and deleterious compounds |
| Hentges et al., 1977 [81] | 10 volunteers | 4 months of high beef diet with 176 g/day of proteins (double content of proteins compared to the meatless diet) vs. meatless diet with 90 g/day of protein | The high beef diet: ⇓ Bifidobacterium adolescentis ⇑ Bacteroides fragilis and vulgatus |
| Romond et al., 1998 [82] | 20 volunteers | Consumption of twice daily for 7 days 15 mL of concentrated whey from milk fermented with Bifidobacterium breve | ⇓ Bacteroides fragilis and Clostridium perfringens |
| Sun et al., 2022 [83] | 60 overweight or with obesity older women | Supplementation for 8 weeks of 15.2 g/day of whey protein or 16.8 g/day of whey protein hydrolysate to the basal quantity of 50 g/day of proteins | No significant changes in gut microbiota composition |
| Reimer et al., 2017 [84] | 125 adults with overweight/obesity | Consumption of 2 isocaloric snack bar/day with 5 g of whey protein for 12 weeks | No variation of gut microbiota |
| Mitchell et al., 2019 [85] | 31 healthy older men with BMI between 20 and 35 kg/m2 | Consumption of the recommended dietary allowance of protein (RDA: 0.8 g protein/kg bodyweight/day) or twice the RDA (2RDA) as part of a supplied diet for 10 weeks | No significant differences in proteolytic microbiota or metabolites of protein fermentation |
| Russell et al., 2011 [86] | 17 men with obesity | High protein (137 g/daily)/low carbohydrate (22 g/daily) diet for 4 weeks | ⇓ Roseburia, Eubacterium rectale and butyrate |
| Clarke et al., 2014 [77] | 40 overweight professional rugby athletes and 46 controls | The athletes usually consumed higher quantities of protein (22% of daily energy) | ⇑ gut microbiota diversity and Akkermansia compared to controls |
| Study | Population | Dietary Intervention | Results |
|---|---|---|---|
| O’keefe et al., 2015 [91] | 20 subjects African Americans and 20 rural Africans with a BMI range between 18–35 kg/m2 | African Americans were fed a high-fiber (55 g/day), low-fat (16% energy), and 14% energy from protein African-style diet, and rural Africans a high-fat (52% energy) low-fiber (12 g/day), and 27% energy from protein Western-style diet for 2 weeks | The switch to the high-fat/low-fiber diet: ⇑ Fusobacterium nucleatum The switch to the low-fat/high fiber diet: ⇓ Bilophila wadsworthia |
| Cotillard et al., 2013 [52] | 38 with obesity and 11 overweight subjects | 12 weeks of energy-restricted high protein diet with 35% protein, 25% lipids, and 44% carbohydrates | Dietary intervention: ⇑ gene richness ⇓ Bifidobacterium and Eubacterium rectale. |
| Reddy et al., 1975 [67] | 8 healthy adults | Consuming 4 weeks of high meat mixed Western diet, consisting in a consumption of 454 g/day of beef, pork, lamb, and/or chicken (23% protein, 45%fat, and 32% carbohydrate) and then switching to a nonmeat diet (20% protein, 30% fat, and 50% carbohydrate). | During the period of consumption of a high meat mixed Western diet compared to the nonmeat diet: ⇑ Bacteroides, Bifidobacterium, Peptococcus, and anaerobic Lactobacillus |
| David et al., 2014 [76] | 10 subjects with BMI range between 19 and 32 kg/m2 | 5 days of an animal-based diet composed of meat, eggs and cheeses (69.5% fat, 30.1% protein, and fiber intake nearly zero) compared to a plant-based diet composed of grains, legumes, fruits, and vegetables (22% fat and 10% protein and 25.6 g fiber/1000 kcal) | The animal-based diet: ⇑ the abundance of bile-tolerant microorganisms (Alistipes, Bilophila, and Bacteroides) ⇓ the levels of Firmicutes that metabolize dietary plant polysaccharides (Roseburia, Eubacterium rectale, and Ruminococcus bromii) |
| Study | Population | Dietary Intervention | Results |
|---|---|---|---|
| Rinott et al., 2022 [94] | 294 participants with visceral obesity | Three intervention groups: (1) healthy dietary guidelines (standard science-based nutritional counseling), (2) MED (3) Green-MED. Both isocaloric MED and Green-MED groups were rich in vegetables, with poultry and fish replacing beef and lamb intake, with a restriction of processed and red meats, a consumption of 28 g of walnuts, and with a 40% of total daily energy from mainly unsaturated fat, a daily protein intake of 123 g on average and a daily carbohydrates intake of 80 g. The Green-MED group was further provided with daily polyphenol-rich green tea and Mankai aquatic plant. | Both MED and Green-MED induced to modification of but microbiota. The Green-MED led to ⇑ Prevotella and enzymatic functions involved in branched-chain amino acid degradation, ⇓ Bifidobacterium and enzymatic functions responsible for branched-chain amino acid biosynthesis |
| Pagliai et al., 2020 [95] | 23 overweight/with obesity omnivores | Randomly assigned to follow for 3 months either a Vegetarian diet (VD) or a Mediterranean diet (MD) with 50–55% of total daily energy from carbohydrate, 25–30% from fat and 15–20% from proteins each | No significantly differences between the two diets at ranks such as phyla and families; VD significantly resulted in ⇑ Anaerostipes and Streptococcus ⇓ Clostridium, Odoribacter and Propionate production MD significantly resulted in ⇑ Lachnoclostridium and Enterohabdus ⇓ Parabacteroides |
| Gutierrez-Diaz et al., 2016 [97] | 31 overweight adults | The group with high adherence to Mediterranean diet (≥4) was characterized by 48 g/day of proteins (of which 32 g/day are from animal source and 15 g/day are from vegetable one), 103 g/day of carbohydrates, and 40 g/day of fats, compared to a 51 g/day of proteins (of which 37 g/day are from animal source and 13 g/day from vegetable one), 96 g/day of carbohydrates, and 43 g/day of fats of the lower adherence group | MDS ≥ 4 was associated with ⇑ Bacteroidetes, Prevotellacea and Prevotella, fecal propionate and butyrate. ⇓ Firmicutes and Lachnospiraceae |
| Meslier et al., 2020 [98] | 82 overweight and with obesity subjects | 43 subjects followed an isocaloric Mediterranean diet (MD) for 8 weeks with a total amount of about 63 g/day of proteins (of which 37 g/day are from vegetable source and 26 g/day from animal source) and 39 followed their habitual diet with low intake of vegetable and fruits (control group) with a total amount of about 68 g/day of proteins (of which 20 g/day are from vegetable source and 48 g/day are from animal one) | The MD intervention leads to: ⇑ levels of the fibre-degrading Faecalibacterium prausnitzii and of genes for microbial carbohydrate degradation linked to butyrate metabolism. |
| Study | Population | Dietary Intervention | Results |
|---|---|---|---|
| Pagliai et al., 2020 [95] | 23 overweight/with obesity omnivores | Randomly assigned to follow for 3 months either a Vegetarian diet (VD) or a Mediterranean diet (MD), with 50–55% of total daily energy from carbohydrate, 25–30% from fat, and 15–20% from proteins each | No significantly differences between the two diets at ranks such as phyla and families; VD significantly resulted in: ⇑ Anaerostipes and Streptococcus ⇓ Clostridium, Odoribacter and propionate production MD significantly resulted in: ⇑ Lachnoclostridium and Enterohabdus ⇓ Parabacteroides |
| Kahleova et al., 2020 [99] | 168 overweight participants | 84 subjects followed for 16 weeks a low-fat vegan diet (with 43 g/day of proteins, of which 1 g/day come from animal source and 42 g/day from vegetable one and 24 g/day of fats) and 84 subjects followed a control diet (with 69 g/day of proteins, of which 39 g/day come from animal source and 29 g/day from vegetable one and 72 g/day of fats) | After 16 weeks of low-fat vegan diet: ⇑ Increased Bacteroidetes, Faecalibacterium prausnitzii and Prevotella ⇓ Bacteroides fragilis |
| Study | Population | Dietary Intervention | Results |
|---|---|---|---|
| Gutierrez-Repiso et al., 2019 [102] | 33 patients with obesity | 2 months of VLCKD (daily protein intake between 0.8–1.5 g/kg ideal body weight) and then 2 months of a LCD. Subjects were randomly allocated to three groups: (1) supplemented with symbiotics, (2) supplemented with a placebo during the VLCKD and symbiotics during the LCD phase, and (3) placebo for the control group | VLCKD without the supplementation of symbiotics resulted in: ⇓ Proteobacteria and Enterobacteriaceae, Sinobacteraceae and Comamonadacea ⇑ Firmicutes, Ruminococcaceae and Morigibacteriaceae. |
| Basciani et al., 2020 [103] | 48 patients with obesity randomized in 3 groups of 16 subjects each | 45 days one of these 3 different VLCKD (all composed of 26 g/day of carbohydrates, 40.4% of total daily energy from fats and about 90 g/day of proteins, about 1.2–1.4 g/kg): (1) Whey protein VLCKD (WPG) (2) Vegetable protein VLCKD (VPG) (3) Animal protein VLCKD (APG) | Independently from the VLCKD type: ⇑ Bacteroidetes; ⇓ Firmicutes WPG determined the strongest effect in: ⇓ Firmicutes ⇑ Bacteroidetes compared to VPG and APG. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Rosa, C.; Di Francesco, L.; Spiezia, C.; Khazrai, Y.M. Effects of Animal and Vegetable Proteins on Gut Microbiota in Subjects with Overweight or Obesity. Nutrients 2023, 15, 2675. https://doi.org/10.3390/nu15122675
Di Rosa C, Di Francesco L, Spiezia C, Khazrai YM. Effects of Animal and Vegetable Proteins on Gut Microbiota in Subjects with Overweight or Obesity. Nutrients. 2023; 15(12):2675. https://doi.org/10.3390/nu15122675
Chicago/Turabian StyleDi Rosa, Claudia, Ludovica Di Francesco, Chiara Spiezia, and Yeganeh Manon Khazrai. 2023. "Effects of Animal and Vegetable Proteins on Gut Microbiota in Subjects with Overweight or Obesity" Nutrients 15, no. 12: 2675. https://doi.org/10.3390/nu15122675
APA StyleDi Rosa, C., Di Francesco, L., Spiezia, C., & Khazrai, Y. M. (2023). Effects of Animal and Vegetable Proteins on Gut Microbiota in Subjects with Overweight or Obesity. Nutrients, 15(12), 2675. https://doi.org/10.3390/nu15122675