Metabolomic Analysis of Vitamin E Supplement Use in the Prostate, Lung, Colorectal, and Ovarian Cancer Screening Trial

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
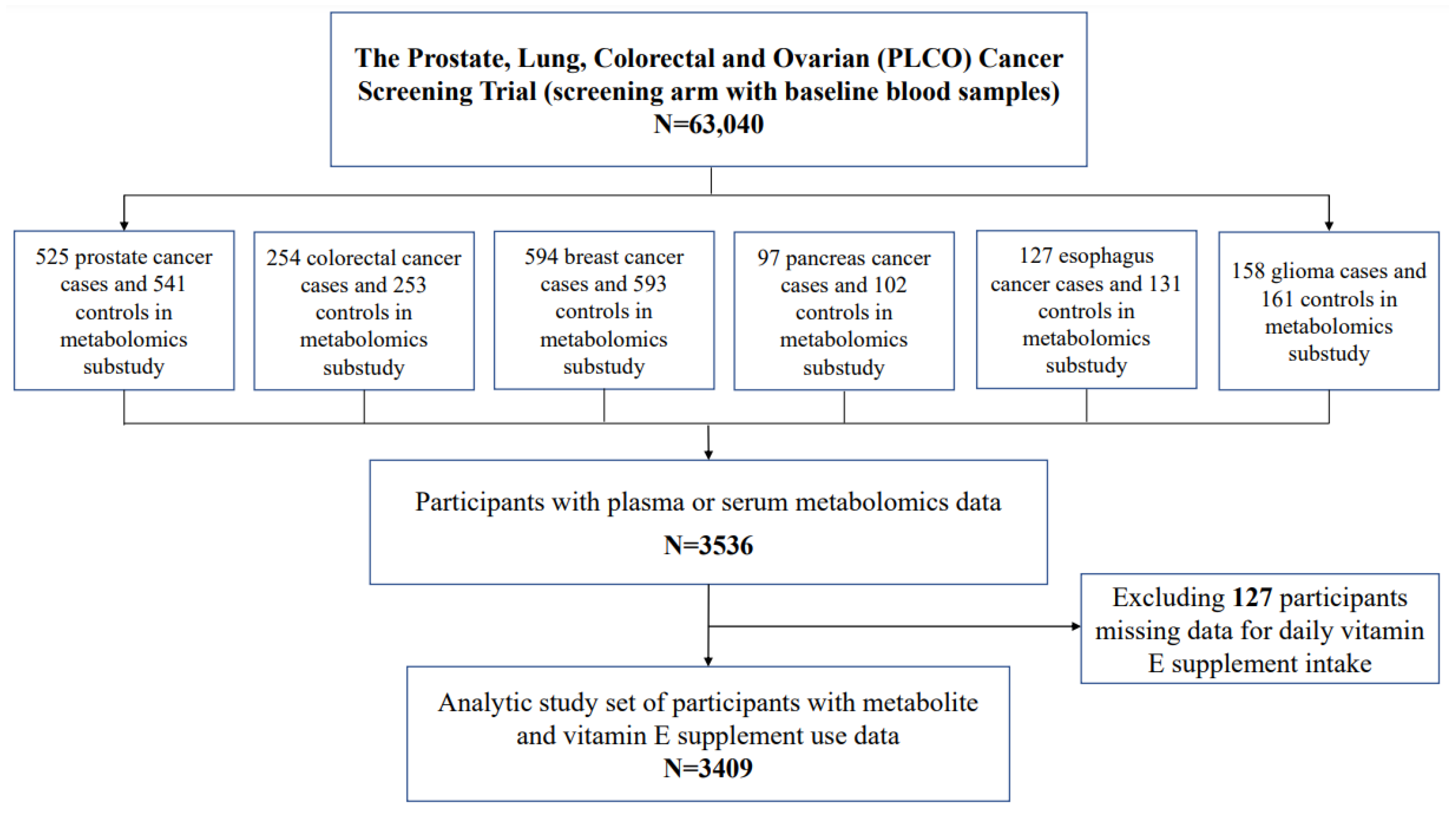
2.1. Study Population
2.2. Serum and DATA Collection
2.3. Metabolomic Analysis
2.4. Statistical Analysis
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Traber, M.G.; Sies, H. Vitamin E in humans: Demand and delivery. Annu. Rev. Nutr. 1996, 16, 321–347. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Weinstein, S.J.; Yu, K.; Mannisto, S.; Albanes, D. A Prospective Study of Serum Vitamin E and 28-Year Risk of Lung Cancer. J. Natl. Cancer Inst. 2020, 112, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, X.; Liu, A.; Wang, G.; Bosland, M.C.; Yang, C.S. delta-Tocopherol inhibits the development of prostate adenocarcinoma in prostate specific Pten-/- mice. Carcinogenesis 2018, 39, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Rao, L.; Yu, G.; Cook, T.R.; Chen, X.; Huang, F. Supramolecular cancer nanotheranostics. Chem. Soc. Rev. 2021, 50, 2839–2891. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Tiwari, A.; Verma, A.; Jain, S.K. Vitamins for Cancer Prevention and Treatment: An Insight. Curr. Mol. Med. 2017, 17, 321–340. [Google Scholar] [CrossRef] [PubMed]
- Shklar, G.; Schwartz, J.L. Vitamin E inhibits experimental carcinogenesis and tumour angiogenesis. Eur. J. Cancer B Oral Oncol. 1996, 32B, 114–119. [Google Scholar] [CrossRef]
- Sigounas, G.; Anagnostou, A.; Steiner, M. dl-alpha-tocopherol induces apoptosis in erythroleukemia, prostate, and breast cancer cells. Nutr. Cancer 1997, 28, 30–35. [Google Scholar] [CrossRef]
- Meydani, S.N.; Beharka, A.A. Recent developments in vitamin E and immune response. Nutr. Rev. 1998, 56, S49–S58. [Google Scholar] [CrossRef]
- Alpha-Tocopherol, Beta Carotene Cancer Prevention Study Group. The effect of vitamin E and beta carotene on the incidence of lung cancer and other cancers in male smokers. New Engl. J. Med. 1994, 330, 1029–1035. [Google Scholar] [CrossRef]
- Gaziano, J.M.; Glynn, R.J.; Christen, W.G.; Kurth, T.; Belanger, C.; MacFadyen, J.; Bubes, V.; Manson, J.E.; Sesso, H.D.; Buring, J.E. Vitamins E and C in the prevention of prostate and total cancer in men: The Physicians’ Health Study II randomized controlled trial. JAMA 2009, 301, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Klein, E.A.; Thompson, I.M., Jr.; Tangen, C.M.; Crowley, J.J.; Lucia, M.S.; Goodman, P.J.; Minasian, L.M.; Ford, L.G.; Parnes, H.L.; Gaziano, J.M.; et al. Vitamin E and the Risk of Prostate Cancer: Updated Results of The Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA 2011, 306, 1549–1556. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Hodis, H.N.; Weinstein, S.J.; Mack, W.J.; Sampson, J.N.; Mondul, A.M.; Albanes, D. Serum Metabolomic Response to Low- and High-Dose Vitamin E Supplementation in Two Randomized Controlled Trials. Cancer Epidemiol. Biomark. Prev. 2020, 29, 1329–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Sousa Coelho, M.; Pereira, I.C.; de Oliveira, K.G.F.; Oliveira, I.K.F.; Dos Santos Rizzo, M.; de Oliveira, V.A.; Carneiro da Silva, F.C.; Torres-Leal, F.L.; de Castro E Sousa, J.M. Chemopreventive and anti-tumor potential of vitamin E in preclinical breast cancer studies: A systematic review. Clin. Nutr. ESPEN 2023, 53, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.H.; Chen, S.J.; Liu, H.; Yan, Y.; Zheng, J.H. Vitamin E consumption and the risk of bladder cancer. Int. J. Vitam. Nutr. Res. 2019, 89, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Li, L.; Tian, Y.; Xu, F.; Qiao, T. Association between Dietary Vitamin E Intake and Esophageal Cancer Risk: An Updated Meta-Analysis. Nutrients 2018, 10, 801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prorok, P.C.; Andriole, G.L.; Bresalier, R.S.; Buys, S.S.; Chia, D.; Crawford, E.D.; Fogel, R.; Gelmann, E.P.; Gilbert, F.; Hasson, M.A.; et al. Design of the Prostate, Lung, Colorectal and Ovarian (PLCO) Cancer Screening Trial. Control Clin. Trials 2000, 21, 273S–309S. [Google Scholar] [CrossRef]
- Huang, J.; Mondul, A.M.; Weinstein, S.J.; Koutros, S.; Derkach, A.; Karoly, E.; Sampson, J.N.; Moore, S.C.; Berndt, S.I.; Albanes, D. Serum metabolomic profiling of prostate cancer risk in the prostate, lung, colorectal, and ovarian cancer screening trial. Br. J. Cancer 2016, 115, 1087–1095. [Google Scholar] [CrossRef] [Green Version]
- Cross, A.J.; Moore, S.C.; Boca, S.; Huang, W.Y.; Xiong, X.; Stolzenberg-Solomon, R.; Sinha, R.; Sampson, J.N. A prospective study of serum metabolites and colorectal cancer risk. Cancer 2014, 120, 3049–3057. [Google Scholar] [CrossRef] [Green Version]
- Playdon, M.C.; Ziegler, R.G.; Sampson, J.N.; Stolzenberg-Solomon, R.; Thompson, H.J.; Irwin, M.L.; Mayne, S.T.; Hoover, R.N.; Moore, S.C. Nutritional metabolomics and breast cancer risk in a prospective study. Am. J. Clin. Nutr. 2017, 106, 637–649. [Google Scholar] [CrossRef] [Green Version]
- Stolzenberg-Solomon, R.; Derkach, A.; Moore, S.; Weinstein, S.J.; Albanes, D.; Sampson, J. Associations between metabolites and pancreatic cancer risk in a large prospective epidemiological study. Gut 2020, 69, 2008–2015. [Google Scholar] [CrossRef]
- Ford, L.; Kennedy, A.D.; Goodman, K.D.; Pappan, K.L.; Evans, A.M.; Miller, L.A.D.; Wulff, J.E.; Wiggs, B.R.; Lennon, J.J.; Elsea, S.; et al. Precision of a Clinical Metabolomics Profiling Platform for Use in the Identification of Inborn Errors of Metabolism. J. Appl. Lab. Med. 2020, 5, 342–356. [Google Scholar] [CrossRef] [PubMed]
- Evans, A.M.; DeHaven, C.D.; Barrett, T.; Mitchell, M.; Milgram, E. Integrated, nontargeted ultrahigh performance liquid chromatography/electrospray ionization tandem mass spectrometry platform for the identification and relative quantification of the small-molecule complement of biological systems. Anal. Chem. 2009, 81, 6656–6667. [Google Scholar] [CrossRef] [PubMed]
- Dehaven, C.D.; Evans, A.M.; Dai, H.; Lawton, K.A. Organization of GC/MS and LC/MS metabolomics data into chemical libraries. J. Cheminform. 2010, 2, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondul, A.M.; Moore, S.C.; Weinstein, S.J.; Karoly, E.D.; Sampson, J.N.; Albanes, D. Metabolomic analysis of prostate cancer risk in a prospective cohort: The alpha-tocolpherol, beta-carotene cancer prevention (ATBC) study. Int. J. Cancer 2015, 137, 2124–2132. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Wang, N.; Ru, Y.; Yang, Z.; Sun, C.; Li, S.; Min, Y.; Zhao, X.; Lu, Y.; Hsing, A.W.; Zhu, S. Metabolomic Profiles of Plasma Retinol-Associated Dyslipidemia in Men and Women. Front. Nutr. 2021, 8, 740435. [Google Scholar] [CrossRef]
- Missmer, S.A.; Eliassen, A.H.; Barbieri, R.L.; Hankinson, S.E. Endogenous estrogen, androgen, and progesterone concentrations and breast cancer risk among postmenopausal women. J. Natl. Cancer Inst. 2004, 96, 1856–1865. [Google Scholar] [CrossRef] [PubMed]
- Cummings, S.R.; Lee, J.S.; Lui, L.Y.; Stone, K.; Ljung, B.M.; Cauleys, J.A. Sex hormones, risk factors, and risk of estrogen receptor–positive breast cancer in older women: A long-term prospective study. Cancer Epidemiol. Biomark. Prev. 2005, 14, 1047–1051. [Google Scholar] [CrossRef] [Green Version]
- Elebro, K.; Borgquist, S.; Simonsson, M.; Markkula, A.; Jirström, K.; Ingvar, C.; Rose, C.; Jernström, H. Combined androgen and estrogen receptor status in breast cancer: Treatment prediction and prognosis in a population-based prospective cohort. Clin. Cancer Res. 2015, 21, 3640–3650. [Google Scholar] [CrossRef] [Green Version]
- Samandari, E.; Visarius, T.; Zingg, J.M.; Azzi, A. The effect of gamma-tocopherol on proliferation, integrin expression, adhesion, and migration of human glioma cells. Biochem. Biophys. Res. Commun. 2006, 342, 1329–1333. [Google Scholar] [CrossRef]
- Heine-Bröring, R.C.; Winkels, R.M.; Renkema, J.M.; Kragt, L.; van Orten-Luiten, A.C.; Tigchelaar, E.F.; Chan, D.S.; Norat, T.; Kampman, E. Dietary supplement use and colorectal cancer risk: A systematic review and meta-analyses of prospective cohort studies. Int. J. Cancer 2015, 136, 2388–2401. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Liu, Y.; Shu, Y.; Chen, X.; Hu, J.; Zheng, R.; Ma, D.; Yang, C.; Guan, X. Link between risk of colorectal cancer and serum vitamin E levels: A meta-analysis of case-control studies. Medicine 2017, 9, e7470. [Google Scholar] [CrossRef] [PubMed]
- Hui, Y.; Jia, X.; Chen, X.; Yang, C.S.; Li, N. Time-selective chemoprevention of vitamin E and selenium on esophageal carcinogenesis in rats: The possible role of nuclear factor kappaB signaling pathway. Int. J. Cancer 2012, 131, 1517–1527. [Google Scholar]
- Korostishevsky, M.; Steves, C.J.; Malkin, I.; Spector, T.; Williams, F.M.; Livshits, G. Genomics and metabolomics of muscular mass in a community-based sample of UK females. Eur. J. Hum. Genet. 2016, 24, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.P.; Zhang, L.R.; Jia, M.; Hu, X.J. CYP3AP1*3 allele is associated with lipid-lowering efficacy of simvastatin and atorvastatin in Chinese women. J. Clin. Pharmacol. 2011, 51, 181–188. [Google Scholar] [CrossRef]
- Baris, D.; Karagas, M.R.; Koutros, S.; Colt, J.S.; Johnson, A.; Schwenn, M.; Fischer, A.H.; Figueroa, J.D.; Berndt, S.I.; Han, S.; et al. Nonsteroidal anti-inflammatory drugs and other analgesic use and bladder cancer in northern New England. Int. J. Cancer. 2013, 132, 162–173. [Google Scholar] [CrossRef] [Green Version]
- Brigelius-Flohe, R. Vitamin E and drug metabolism. Biochem. Biophys. Res. Commun. 2003, 305, 737–740. [Google Scholar] [CrossRef]
- Chen, Y.; Tang, Y.; Wang, M.T.; Zeng, S.; Nie, D. Human pregnane X receptor and resistance to chemotherapy in prostate cancer. Cancer Res. 2007, 67, 10361–10367. [Google Scholar] [CrossRef] [Green Version]
- Fredericks, W.J.; Sepulveda, J.; Lai, P.; Tomaszewski, J.E.; Lin, M.F.; McGarvey, T.; Rauscher, F.J., 3rd; Malkowicz, S.B. The tumor suppressor TERE1 (UBIAD1) prenyltransferase regulates the elevated cholesterol phenotype in castration resistant prostate cancer by controlling a program of ligand dependent SXR target genes. Oncotarget 2013, 4, 1075–1092. [Google Scholar] [CrossRef] [Green Version]
- Pondugula, S.R.; Pavek, P.; Mani, S. Pregnane X Receptor and Cancer: Context-Specificity is Key. Nucl. Recept. Res. 2016, 3, 101198. [Google Scholar] [CrossRef] [Green Version]
- Carr, D.F.; O’Meara, H.; Jorgensen, A.L.; Campbell, J.; Hobbs, M.; McCann, G.; van Staa, T.; Pirmohamed, M. SLCO1B1 genetic variant associated with statin-induced myopathy: A proof-of-concept study using the clinical practice research datalink. Clin. Pharmacol. Ther. 2013, 94, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Craig, E.L.; Stopsack, K.H.; Evergren, E.; Penn, L.Z.; Freedland, S.J.; Hamilton, R.J.; Allott, E.H. Statins and prostate cancer-hype or hope? The epidemiological perspective. Prostate Cancer Prostatic Dis. 2022, 25, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ni, J.; Messing, E.M.; Chang, E.; Yang, C.R.; Yeh, S. Vitamin E succinate inhibits the function of androgen receptor and the expression of prostate-specific antigen in prostate cancer cells. Proc. Natl. Acad. Sci. USA 2002, 99, 7408–7413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondul, A.M.; Rohrmann, S.; Menke, A.; Feinleib, M.; Nelson, W.G.; Platz, E.A.; Albanes, D. Association of serum alpha-tocopherol with sex steroid hormones and interactions with smoking: Implications for prostate cancer risk. Cancer Causes Control. 2011, 22, 827–836. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, B.T.; Hurt, E.M.; Kalathur, M.; Duhagon, M.A.; Milner, J.A.; Kim, Y.S.; Farrar, W.L. Effects of the sesquiterpene lactone parthenolide on prostate tumor-initiating cells: An integrated molecular profiling approach. Prostate 2009, 69, 827–837. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.J.; Lim, S.S.; Park, S.Y.; Shin, H.K.; Kim, J.S.; Park, J.H. Apoptosis of DU145 human prostate cancer cells induced by dehydrocostus lactone isolated from the root of Saussurea lappa. Food Chem. Toxicol. 2008, 46, 3651–3658. [Google Scholar] [CrossRef]
- Sanderson, T.; Renaud, M.; Scholten, D.; Nijmeijer, S.; van den Berg, M.; Cowell, S.; Guns, E.; Nelson, C.; Mutarapat, T.; Ruchirawat, S. Effects of lactone derivatives on aromatase (CYP19) activity in H295R human adrenocortical and (anti)androgenicity in transfected LNCaP human prostate cancer cells. Eur. J. Pharmacol. 2008, 593, 92–98. [Google Scholar] [CrossRef]


{kind=link}
| All Participants | Men | Women | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 IU | 4.3–<400 IU IU | ≥400 IU | 0 IU | 4.3–<400 IU IU | ≥400 IU | 0 IU | 4.3–<400 IU IU | ≥400 IU | |
| n | 1655 | 872 | 882 | 1002 | 446 | 389 | 653 | 426 | 493 |
| Age, years | 63.4 ± 5.0 | 63.1 ± 5.3 | 63.0 ± 5.2 | 63.6 ± 5.0 | 62.8 ± 5.3 | 63.0 ± 5.1 | 63.3 ± 5.1 | 63.3 ± 5.4 | 62.9 ± 5.2 |
| Race, % | |||||||||
| White, non-Hispanic | 79.8 | 86.6 | 85.7 | 72.8 | 81.6 | 77.6 | 90.5 | 91.8 | 92.1 |
| Black, non-Hispanic | 16.7 | 8.9 | 9.1 | 24.2 | 13.5 | 17.0 | 5.2 | 4.2 | 2.8 |
| Other races combined | 3.6 | 4.5 | 5.2 | 3.1 | 4.9 | 5.4 | 4.3 | 4.0 | 5.1 |
| Height, cm | 172.1 ± 9.9 | 171.0 ± 10.1 | 169.9 ± 9.8 | 177.9 ± 7.1 | 178.2 ± 7.0 | 178.1 ± 7.0 | 163.2 ± 6.3 | 163.5 ± 6.9 | 163.5 ± 6.4 |
| Weight, kg | 82.9 ± 16.7 | 79.9 ± 15.5 | 78.5 ± 16.5 | 88.7 ± 15.2 | 86.6 ± 14.0 | 88.0 ± 14.8 | 74.0 ± 15.0 | 72.8 ± 13.8 | 71.0 ± 13.7 |
| Body mass index, kg/m2 | 27.9 ± 4.8 | 27.3 ± 4.6 | 27.1 ± 4.7 | 28.0 ± 4.5 | 27.3 ± 4.0 | 27.7 ± 4.1 | 27.8 ± 5.3 | 27.3 ± 5.2 | 26.6 ± 5.1 |
| Tobacco smoking status, % | |||||||||
| Never smoker | 44.5 | 47.7 | 47.4 | 35.7 | 36.8 | 35.5 | 57.9 | 59.2 | 56.8 |
| Former smoker | 9.7 | 6.2 | 7.4 | 10.9 | 6.5 | 8.7 | 7.8 | 5.9 | 6.3 |
| Current smoker | 45.9 | 46.1 | 45.2 | 53.4 | 56.7 | 55.8 | 34.3 | 35.0 | 36.9 |
| Multivitamin use, % | 0 | 95.8 | 74.3 | 0 | 95.7 | 72.8 | 0 | 95.8 | 75.5 |
| 4.3–<400 IU IU | ≥400 IU | |||||||
|---|---|---|---|---|---|---|---|---|
| Metabolite | Chemical Class | Sub-Pathway | F-Value | FDR- Adjusted p-Value | Effect Size (β) | p-Value | Effect Size (β) | p-Value |
| Alpha-CEHC | Cofactors and vitamins | Tocopherol metabolism | 364.47 | <0.001 | 0.62 | <0.001 | 1.80 | <0.001 |
| Alpha-CEHC-glucuronide | Cofactors and vitamins | Tocopherol metabolism | 338.54 | <0.001 | 0.66 | <0.001 | 1.90 | <0.001 |
| Alpha-Tocopherol | Cofactors and vitamins | Tocopherol metabolism | 318.33 | <0.001 | 0.18 | <0.001 | 0.44 | <0.001 |
| Alpha-CEHC sulfate | Cofactors and vitamins | Tocopherol metabolism | 214.10 | <0.001 | 1.09 | <0.001 | 2.38 | <0.001 |
| Pantothenic acid | Cofactors and vitamins | Pantothenate and CoA metabolism | 53.15 | <0.001 | 0.17 | <0.001 | 0.29 | <0.001 |
| L-Threonic acid | Cofactors and vitamins | Ascorbate and aldarate metabolism | 43.46 | <0.001 | 0.11 | 0.002 | 0.27 | <0.001 |
| 4-Pyridoxic acid | Cofactors and vitamins | Vitamin B6 metabolism | 43.16 | <0.001 | 0.14 | 0.021 | 0.43 | <0.001 |
| Delta-Tocopherol | Cofactors and vitamins | Tocopherol metabolism | 34.05 | <0.001 | −0.20 | 0.008 | −0.49 | <0.001 |
| Glyceric acid | Carbohydrate | Glycolysis, gluconeogenesis, pyruvate metabolism | 30.86 | <0.001 | 0.08 | <0.001 | 0.13 | <0.001 |
| Oxalic acid | Carbohydrate | Glyoxylate and dicarboxylate metabolism | 28.66 | <0.001 | 0.10 | 0.013 | 0.24 | <0.001 |
| Pyridoxal | Cofactors and vitamins | Vitamin B6 metabolism | 22.79 | <0.001 | 0.03 | 0.79 | 0.45 | <0.001 |
| Hydroxypropanedioic acid | Xenobiotics | Food component/plant | 16.12 | <0.001 | 0.12 | 0.002 | 0.18 | <0.001 |
| N1-Methyl-2-pyridone-5-carboxamide | Cofactors and vitamins | Nicotinate and nicotinamide metabolism | 15.21 | <0.001 | 0.08 | 0.041 | 0.18 | <0.001 |
| Glycocholenate sulfate | Lipid | Bile acid metabolism | 15.16 | <0.001 | −0.10 | 0.001 | −0.15 | <0.001 |
| Beta-Tocopherol | Cofactors and vitamins | Tocopherol metabolism | 14.46 | <0.001 | −0.26 | 0.020 | −0.49 | <0.001 |
| Hydroxy-carboxy-4-methyl-5-propyl-2-furanpropionic acid (CMPF) | Lipid | Fatty acid, dicarboxylate | 14.36 | <0.001 | −0.12 | 0.043 | 0.13 | 0.013 |
| N1-Methyl-4-pyridone-3-carboxamide | Cofactors and vitamins | Nicotinate and nicotinamide metabolism | 11.37 | 0.001 | 0.13 | 0.004 | 0.18 | <0.001 |
| Isovaleric acid | Lipid | Fatty acid metabolism | 10.98 | 0.001 | 0.00 | 0.882 | 0.09 | <0.001 |
| 4-Androsten-3alpha,17alpha-diol monosulfate (2) or androstenediol (3alpha, 17alpha) monsulfate (2) | Lipid | Androgenic steroids | 10.34 | 0.002 | −0.15 | 0.065 | −0.18 | 0.012 |
| C22 lactone sulfate (X_12063) | Partially characterized | Partially characterized | 8.99 | 0.007 | −0.04 | 0.500 | −0.17 | <0.001 |
| Decanoylcarnitine | Lipid | Carnitine metabolism | 8.28 | 0.013 | 0.01 | 0.885 | −0.11 | 0.003 |
| Docosahexaenoic acid | Lipid | Essential fatty acid | 8.19 | 0.014 | −0.04 | 0.228 | 0.06 | 0.031 |
| Androsterone glucuronide | Lipid | Androgenic steroids | 8.08 | 0.016 | −0.19 | 0.008 | −0.26 | <0.001 |
| Phosphate | Energy | Oxidative phosphorylation | 8.02 | 0.016 | 0.06 | <0.001 | 0.06 | <0.001 |
| L-Octanoylcarnitine | Lipid | Carnitine metabolism | 7.86 | 0.017 | 0.01 | 0.806 | −0.10 | 0.004 |
| N-Stearoyltaurine or N-Stearoyl taurine | Lipid | Endocannabinoid | 7.86 | 0.018 | 0.07 | 0.244 | 0.01 | 0.863 |
| Sphingomyelin (d17:1/14:0, d16:1/15:0) | Lipid | Sphingomyelins | 7.67 | 0.020 | −0.15 | 0.014 | −0.10 | 0.063 |
| Eicosapentaenoic acid | Lipid | Essential fatty acid | 7.55 | 0.021 | 0.01 | 0.733 | 0.10 | 0.002 |
| 5alpha-Androstan-3alpha,17alpha-diol disulfate | Lipid | Sterol/steroid | 7.50 | 0.021 | 0.03 | 0.733 | −0.05 | 0.492 |
| S-Allylcysteine | Xenobiotics | Food component/plant | 7.29 | 0.026 | −0.19 | 0.111 | 0.17 | 0.100 |
| 3-Carboxy-4-methyl-5-propyl-2-furanpropionic acid (CMPF) | Lipid | Fatty acid, dicarboxylate | 7.25 | 0.026 | −0.08 | 0.210 | 0.11 | 0.054 |
| L-Aspartic acid | Amino acid | Alanine and aspartate metabolism | 7.19 | 0.027 | 0.01 | 0.663 | −0.05 | 0.010 |
| Monoglyceride (18:1(9Z)/0:0/0:0) | Lipid | Monoacylglycerol | 7.18 | 0.027 | 0.03 | 0.594 | 0.01 | 0.760 |
| L-Urobilin | Cofactors and vitamins | Hemoglobin and porphyrin metabolism | 7.05 | 0.030 | −0.02 | 0.799 | −0.24 | 0.003 |
| L-Glutamine | Amino acid | Glutamate metabolism | 7.02 | 0.030 | −0.02 | 0.032 | −0.03 | <0.001 |
| Sphingomyelin (d17:1/16:0, d18:1/15:0, d16:1/17:0) or sphingomyelin (d18:1/15:0, d16:1/17:0) | Lipid | Sphingomyelins | 6.90 | 0.034 | −0.08 | 0.001 | −0.09 | <0.001 |
| Tauro-b-muricholic acid | Lipid | Primary bile acid metabolism | 6.82 | 0.034 | 0.18 | 0.064 | 0.30 | <0.001 |
| Lactosylceramide (d18:1/22:0) | Lipid | Lactosylceramides (LCER) | 6.87 | 0.034 | −0.19 | <0.001 | −0.11 | 0.008 |
| 2-Methoxyacetaminophen glucuronide | Xenobiotics | Drug | 6.80 | 0.034 | 0.42 | 0.004 | 0.22 | 0.090 |
| 9-Hexadecenoylcarnitine | Lipid | Fatty acid metabolism (acyl carnitine, monounsaturated | 6.75 | 0.036 | 0.04 | 0.393 | 0.02 | 0.585 |
| Sphingomyelin (d17:2/16:0, d18:2/15:0) | Lipid | Sphingomyelins | 6.78 | 0.036 | −0.17 | 0.004 | −0.12 | 0.013 |
| Phosphatidylcholine (P-16:0/18:2) | Lipid | Plasmalogen | 6.67 | 0.038 | −0.11 | 0.001 | −0.09 | 0.002 |
| Homo-L-Arginine | Amino acid | Urea cycle; arginine and proline metabolism | 6.54 | 0.042 | −0.11 | 0.105 | 0.08 | 0.147 |
| Pregnanediol | Lipid | Sterol/steroid | 6.47 | 0.044 | −0.07 | 0.126 | −0.14 | 0.001 |
| 5alpha-Androstan-3alpha,17beta-diol disulfate | Lipid | Sterol/steroid | 6.46 | 0.044 | −0.11 | 0.192 | −0.18 | 0.013 |
| (R)C(S)S-Alliin | Xenobiotics | Food component/plant | 6.27 | 0.051 | −0.17 | 0.177 | 0.19 | 0.096 |
| Oxypurinol | Xenobiotics | Drug—metabolic | 6.14 | 0.057 | −0.66 | 0.002 | −0.16 | 0.371 |
| L-Serine | Amino acid | Glycine, serine and threonine metabolism | 6.13 | 0.057 | −0.04 | 0.010 | −0.04 | 0.001 |
| O-Desmethyltramadol | Xenobiotics | Drug—analgesics, anesthetics | 6.04 | 0.063 | −0.13 | 0.008 | −0.07 | 0.137 |
| Dimethyl sulfone | Xenobiotics | Chemical | 5.89 | 0.070 | 0.06 | 0.426 | 0.20 | 0.002 |
| O-Phosphoethanolamine | Lipid | Phospholipid metabolism | 5.88 | 0.070 | −0.10 | 0.239 | 0.13 | 0.090 |
| cis-4-Decenoate | Lipid | Medium-chain fatty acid | 5.87 | 0.070 | −0.03 | 0.668 | −0.11 | 0.069 |
| L-Methionine | Amino acid | Cysteine, methionine, SAM, taurine metabolism | 5.86 | 0.070 | 0.01 | 0.662 | 0.00 | 0.807 |
| 3b,17a-Dihydroxy-5a-androstane | Lipid | Androgenic steroids | 5.84 | 0.070 | −0.15 | 0.063 | −0.20 | 0.005 |
| Hydrochlorothiazide | Xenobiotics | Drug | 5.68 | 0.078 | 0.24 | 0.055 | 0.37 | 0.001 |
| Monoglyceride (22:6(4Z,7Z,10Z,13Z,16Z,19Z)/0:0/0:0) | Lipid | Monoacylglycerol | 5.63 | 0.082 | 0.00 | 0.994 | 0.09 | 0.075 |
| Sphingomyelin (d18:1/22:0) | Lipid | Sphingomyelins | 5.62 | 0.082 | −0.07 | 0.008 | −0.08 | 0.001 |
| Dehydroepiandrosterone (DHEA) sulfate | Lipid | Sterol/steroid | 5.56 | 0.083 | −0.09 | 0.034 | −0.13 | 0.001 |
| L-Cysteine | Amino acid | Cysteine, methionine, SAM, taurine metabolism | 5.56 | 0.083 | −0.02 | 0.406 | 0.03 | 0.055 |
| Etiocholanolone glucuronide | Lipid | Androgenic steroids | 5.46 | 0.089 | −0.17 | 0.026 | −0.22 | 0.001 |
| 4-Androsten-3alpha,17alpha-diol monosulfate (3) or androstenediol (3alpha, 17alpha) monsulfate (3) | Lipid | Androgenic steroids | 5.46 | 0.089 | −0.10 | 0.050 | −0.14 | 0.003 |
| Lidocaine | Xenobiotics | Drug | 5.42 | 0.091 | 0.18 | 0.001 | 0.12 | 0.017 |
| Ergothioneine | Xenobiotics | Food component/plant | 5.38 | 0.091 | −0.05 | 0.224 | 0.06 | 0.118 |
| Gamma-Glutamylalanine | Peptide | Gamma-glutamyl | 5.37 | 0.091 | 0.03 | 0.300 | 0.02 | 0.353 |
| Pioglitazone | Xenobiotics | Drug—metabolic | 5.37 | 0.091 | 0.60 | 0.001 | 0.38 | 0.018 |
| Phenol sulfate | Amino acid | Phenylalanine and tyrosine metabolism | 5.31 | 0.092 | 0.02 | 0.742 | −0.09 | 0.024 |
| 5′-Methylthioadenosine | Amino acid | Polyamine metabolism | 5.25 | 0.096 | 0.14 | 0.001 | 0.08 | 0.033 |
| Ketopioglitazone | Xenobiotics | Drug—metabolic | 5.23 | 0.098 | 0.20 | 0.001 | 0.12 | 0.020 |
| Hydroxypioglitazone (M-IV) | Xenobiotics | Drug—metabolic | 5.22 | 0.098 | 0.12 | 0.001 | 0.07 | 0.021 |
| Phosphatidylcholine (18:2(9Z,12Z)) | Lipid | Lysolipid | 5.18 | 0.099 | −0.07 | 0.001 | −0.03 | 0.055 |
| Men (n = 1837) | Women (n = 1572) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 4.3–<400 IU | ≥400 IU | 4.3–<400 IU | ≥400 IU | |||||||
| Metabolite | FDR- Adjusted p-Value | Effect Size (β) | p-Value | Effect Size (β) | p-Value | FDR- Adjusted p-Value | Effect Size (β) | p-Value | Effect Size (β) | p-Value |
| Alpha-CEHC sulfate | <0.001 | 0.94 | <0.001 | 10−18 | <0.001 | <0.001 | 1.26 | <0.001 | 2.69 | <0.001 |
| Alpha-CEHC glucuronide | <0.001 | 0.47 | <0.001 | 1.50 | <0.001 | <0.001 | 0.90 | <0.001 | 2.29 | <0.001 |
| Alpha-Tocopherol | <0.001 | 0.16 | <0.001 | 0.43 | <0.001 | <0.001 | 0.19 | <0.001 | 0.45 | <0.001 |
| Alpha-CEHC | <0.001 | 0.48 | <0.001 | 1.37 | <0.001 | <0.001 | 0.78 | <0.001 | 2.15 | <0.001 |
| Pantothenic acid | <0.001 | 0.25 | <0.001 | 0.34 | <0.001 | <0.001 | 0.10 | 0.034 | 0.24 | <0.001 |
| 4-Pyridoxic acid | <0.001 | 0.18 | 0.018 | 0.42 | <0.001 | <0.001 | 0.12 | 0.222 | 0.45 | <0.001 |
| L-Threonic acid | <0.001 | 0.15 | 0.007 | 0.29 | <0.001 | <0.001 | 0.07 | 0.118 | 0.24 | <0.001 |
| Glyceric acid | <0.001 | 0.10 | 0.001 | 0.15 | <0.001 | <0.001 | 0.06 | 0.025 | 0.11 | <0.001 |
| Delta-Tocopherol | <0.001 | −0.19 | 0.176 | −0.62 | <0.001 | <0.001 | −0.21 | 0.019 | −0.45 | <0.001 |
| Oxalic acid | <0.001 | 0.11 | 0.055 | 0.26 | <0.001 | <0.001 | 0.07 | 0.178 | 0.21 | <0.001 |
| N1-Methyl-4-pyridone-3-carboxamide | 0.003 | 0.11 | 0.027 | 0.20 | <0.001 | 0.074 | 0.04 | 0.468 | 0.15 | 0.002 |
| Pyridoxal | 0.010 | −0.01 | 0.916 | 0.29 | 0.002 | <0.001 | 0.12 | 0.472 | 0.61 | <0.001 |
| Glycocholenate sulfate | 0.052 | −0.08 | 0.055 | −0.13 | <0.001 | 0.009 | −0.13 | 0.004 | −0.17 | <0.001 |
| C22 lactone sulfate (X_12063) 3 | 0.595 | 0.02 | 0.814 | −0.08 | 0.188 | 0.046 | −0.12 | 0.186 | −0.26 | 0.001 |
| 5-Androstenediol 3 | 0.305 | −0.05 | 0.253 | −0.11 | 0.011 | 0.057 | −0.15 | 0.023 | −0.21 | <0.001 |
| Androsterone glucuronide 3 | 0.531 | −0.06 | 0.343 | −0.11 | 0.048 | 0.091 | −0.32 | 0.010 | −0.37 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, J.; Hong, H.G.; Weinstein, S.J.; Playdon, M.C.; Cross, A.J.; Stolzenberg-Solomon, R.; Freedman, N.D.; Huang, J.; Albanes, D. Metabolomic Analysis of Vitamin E Supplement Use in the Prostate, Lung, Colorectal, and Ovarian Cancer Screening Trial. Nutrients 2023, 15, 2836. https://doi.org/10.3390/nu15132836
Lim J, Hong HG, Weinstein SJ, Playdon MC, Cross AJ, Stolzenberg-Solomon R, Freedman ND, Huang J, Albanes D. Metabolomic Analysis of Vitamin E Supplement Use in the Prostate, Lung, Colorectal, and Ovarian Cancer Screening Trial. Nutrients. 2023; 15(13):2836. https://doi.org/10.3390/nu15132836
Chicago/Turabian StyleLim, Jungeun, Hyokyoung G. Hong, Stephanie J. Weinstein, Mary C. Playdon, Amanda J. Cross, Rachael Stolzenberg-Solomon, Neal D. Freedman, Jiaqi Huang, and Demetrius Albanes. 2023. "Metabolomic Analysis of Vitamin E Supplement Use in the Prostate, Lung, Colorectal, and Ovarian Cancer Screening Trial" Nutrients 15, no. 13: 2836. https://doi.org/10.3390/nu15132836
APA StyleLim, J., Hong, H. G., Weinstein, S. J., Playdon, M. C., Cross, A. J., Stolzenberg-Solomon, R., Freedman, N. D., Huang, J., & Albanes, D. (2023). Metabolomic Analysis of Vitamin E Supplement Use in the Prostate, Lung, Colorectal, and Ovarian Cancer Screening Trial. Nutrients, 15(13), 2836. https://doi.org/10.3390/nu15132836