
Auraptene Has Antiviral Activity against Human Coronavirus OC43 in MRC-5 Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Compound
2.2. Cells and Virus
2.3. Cytopathic Effect Reduction Assay
2.4. HCoV-OC43 Viral RNA Copy Number Analysis
2.5. Western Blot Assay
2.6. Immunofluorescence Staining Assay
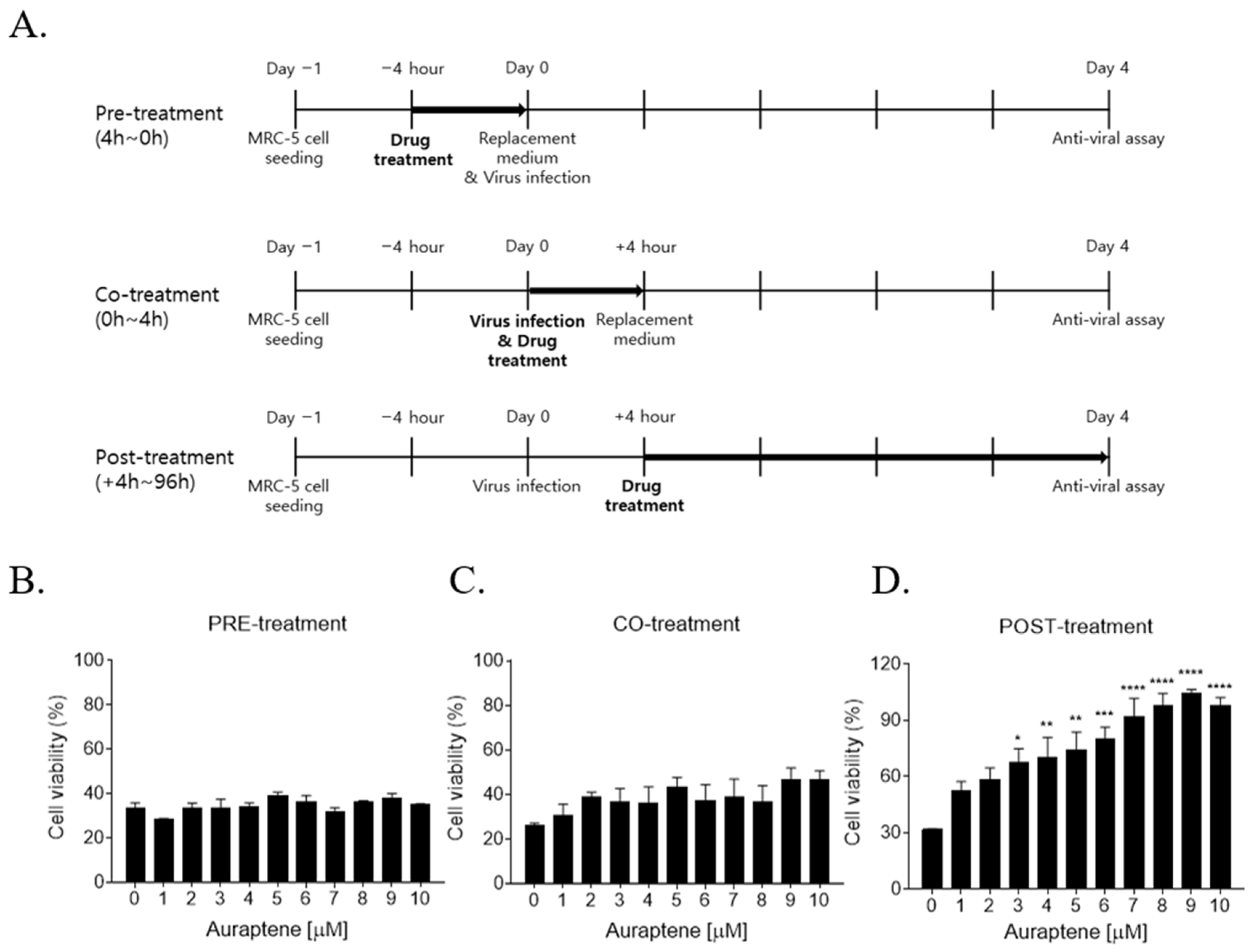
2.7. Time-of-Addition Assay
2.8. Quantitative Real-Time PCR (qRT-PCR)
2.9. Statistical Analysis
3. Results
3.1. Auraptene Inhibits the HCoV-OC43-Induced Cytopathic Effects
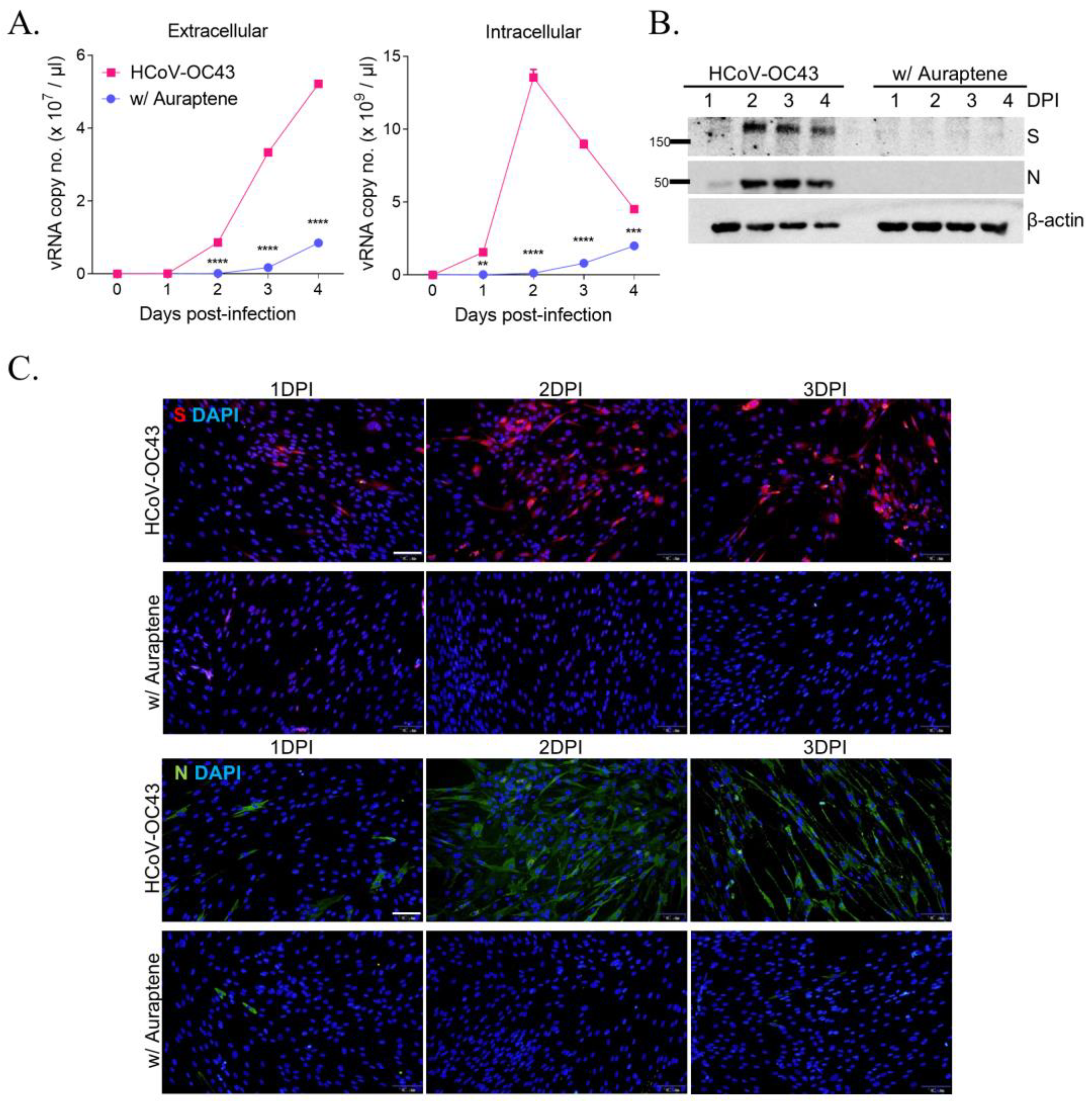
3.2. Auraptene Inhibits HCoV-OC43 Viral RNA Replication and Viral RNA Expression
3.3. Auraptene Inhibited HCoV-OC43 Infection at the Post-Entry Infection Stage
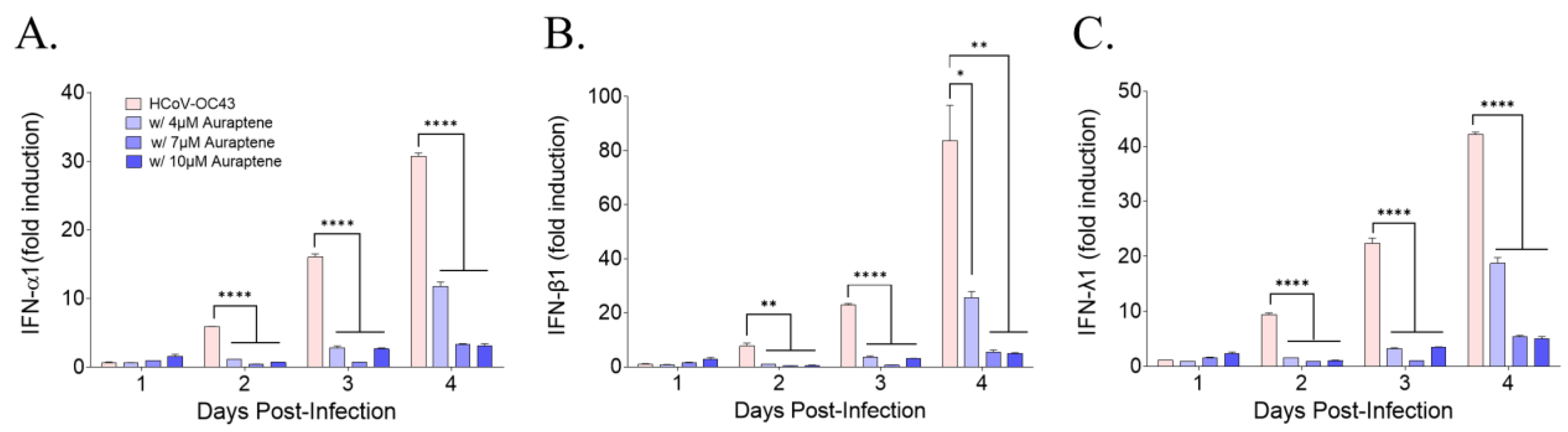
3.4. Auraptene Did Not Induce the Antiviral IFN Gene Expression
3.5. Auraptene Decreased MMP-9 mRNA Levels, which Increased with HCoV-OC43 Infection in MRC-5 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- V’Kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus biology and replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.; Lau, S.K.; Lam, C.S.; Lau, C.C.; Tsang, A.K.; Lau, J.H.; Bai, R.; Teng, J.L.; Tsang, C.C.; Wang, M.; et al. Discovery of seven novel Mammalian and avian coronaviruses in the genus deltacoronavirus supports bat coronaviruses as the gene source of alphacoronavirus and betacoronavirus and avian coronaviruses as the gene source of gammacoronavirus and deltacoronavirus. J. Virol. 2012, 86, 3995–4008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 25 May 2023).
- Monto, A.S. Medical reviews. Coronaviruses. Yale J. Biol. Med. 1974, 47, 234–251. [Google Scholar] [PubMed]
- Choi, W.I.; Kim, I.B.; Park, S.J.; Ha, E.H.; Lee, C.W. Comparison of the clinical characteristics and mortality of adults infected with human coronaviruses 229E and OC43. Sci. Rep. 2021, 11, 4499. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.H.; Min, J.S.; Kwon, S. Cardamonin as a p38 MAPK Signaling Pathway Activator Inhibits Human Coronavirus OC43 Infection in Human Lung Cells. Nutrients 2023, 15, 1335. [Google Scholar] [CrossRef]
- Hickerson, B.T.; Sheikh, F.; Donnelly, R.P.; Ilyushina, N.A. Comparison of the Antiviral Activity of Remdesivir, Chloroquine, and Interferon-beta as Single or Dual Agents Against the Human Beta-Coronavirus OC43. J. Interferon. Cytokine Res. 2023, 43, 35–42. [Google Scholar] [CrossRef]
- Komatsu, S.; Tanaka, S.; Ozawa, S.; Kubo, R.; Ono, Y.; Matsuda, Z. Biochemical studies on grape fruits, Citrus aurantium. Nippon. Kagaku Kaishi 1930, 51, 478–498. [Google Scholar] [CrossRef]
- Curini, M.; Cravotto, G.; Epifano, F.; Giannone, G. Chemistry and biological activity of natural and synthetic prenyloxycoumarins. Curr. Med. Chem. 2006, 13, 199–222. [Google Scholar] [CrossRef]
- Bibak, B.; Shakeri, F.; Barreto, G.E.; Keshavarzi, Z.; Sathyapalan, T.; Sahebkar, A. A review of the pharmacological and therapeutic effects of auraptene. Biofactors 2019, 45, 867–879. [Google Scholar] [CrossRef]
- Genovese, S.; Epifano, F. Auraptene: A natural biologically active compound with multiple targets. Curr. Drug. Targets 2011, 12, 381–386. [Google Scholar] [CrossRef]
- Tayarani-Najaran, Z.; Tayarani-Najaran, N.; Eghbali, S. A Review of Auraptene as an Anticancer Agent. Front. Pharmacol. 2021, 12, 698352. [Google Scholar] [CrossRef]
- Khomenko, T.M.; Zarubaev, V.V.; Orshanskaya, I.R.; Kadyrova, R.A.; Sannikova, V.A.; Korchagina, D.V.; Volcho, K.P.; Salakhutdinov, N.F. Anti-influenza activity of monoterpene-containing substituted coumarins. Bioorg. Med. Chem. Lett. 2017, 27, 2920–2925. [Google Scholar] [CrossRef]
- Li, G.; Gao, Q.; Yuan, S.; Wang, L.; Altmeyer, R.; Lan, K.; Yin, F.; Zou, G. Characterization of three small molecule inhibitors of enterovirus 71 identified from screening of a library of natural products. Antiviral. Res. 2017, 143, 85–96. [Google Scholar] [CrossRef]
- Kim, D.E.; Min, J.S.; Jang, M.S.; Lee, J.Y.; Shin, Y.S.; Song, J.H.; Kim, H.R.; Kim, S.; Jin, Y.H.; Kwon, S. Natural Bis-Benzylisoquinoline Alkaloids-Tetrandrine, Fangchinoline, and Cepharanthine, Inhibit Human Coronavirus OC43 Infection of MRC-5 Human Lung Cells. Biomolecules 2019, 9, 696. [Google Scholar] [CrossRef] [Green Version]
- Lei, X.; Dong, X.; Ma, R.; Wang, W.; Xiao, X.; Tian, Z.; Wang, C.; Wang, Y.; Li, L.; Ren, L.; et al. Activation and evasion of type I interferon responses by SARS-CoV-2. Nat. Commun. 2020, 11, 3810. [Google Scholar] [CrossRef]
- Kawabata, K.; Murakami, A.; Ohigashi, H. Auraptene decreases the activity of matrix metalloproteinases in dextran sulfate sodium-induced ulcerative colitis in ICR mice. Biosci. Biotechnol. Biochem. 2006, 70, 3062–3065. [Google Scholar] [CrossRef] [Green Version]
- Jamialahmadi, K.; Salari, S.; Alamolhodaei, N.S.; Avan, A.; Gholami, L.; Karimi, G. Auraptene Inhibits Migration and Invasion of Cervical and Ovarian Cancer Cells by Repression of Matrix Metalloproteinasas 2 and 9 Activity. J. Pharmacopunct. 2018, 21, 177–184. [Google Scholar] [CrossRef]
- Marquis, A.; Genovese, S.; Epifano, F.; Grenier, D. The plant coumarins auraptene and lacinartin as potential multifunctional therapeutic agents for treating periodontal disease. BMC Complement. Altern. Med. 2012, 12, 80. [Google Scholar] [CrossRef]
- Wang, P.; Dai, J.; Bai, F.; Kong, K.F.; Wong, S.J.; Montgomery, R.R.; Madri, J.A.; Fikrig, E. Matrix metalloproteinase 9 facilitates West Nile virus entry into the brain. J. Virol. 2008, 82, 8978–8985. [Google Scholar] [CrossRef] [Green Version]
- Ju, S.M.; Song, H.Y.; Lee, J.A.; Lee, S.J.; Choi, S.Y.; Park, J. Extracellular HIV-1 Tat up-regulates expression of matrix metalloproteinase-9 via a MAPK-NF-kappaB dependent pathway in human astrocytes. Exp. Mol. Med. 2009, 41, 86–93. [Google Scholar] [CrossRef]
- Mishra, M.K.; Dutta, K.; Saheb, S.K.; Basu, A. Understanding the molecular mechanism of blood-brain barrier damage in an experimental model of Japanese encephalitis: Correlation with minocycline administration as a therapeutic agent. Neurochem. Int. 2009, 55, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Calderon-Pelaez, M.A.; Velandia-Romero, M.L.; Bastidas-Legarda, L.Y.; Beltran, E.O.; Camacho-Ortega, S.J.; Castellanos, J.E. Dengue Virus Infection of Blood-Brain Barrier Cells: Consequences of Severe Disease. Front. Microbiol. 2019, 10, 1435. [Google Scholar] [CrossRef] [Green Version]
- Hui, L.; Nie, Y.; Li, S.; Guo, M.; Yang, W.; Huang, R.; Chen, J.; Liu, Y.; Lu, X.; Chen, Z.; et al. Matrix metalloproteinase 9 facilitates Zika virus invasion of the testis by modulating the integrity of the blood-testis barrier. PLoS Pathog. 2020, 16, e1008509. [Google Scholar] [CrossRef] [PubMed]
- Yoshizaki, T.; Sato, H.; Furukawa, M.; Pagano, J.S. The expression of matrix metalloproteinase 9 is enhanced by Epstein-Barr virus latent membrane protein 1. Proc. Natl. Acad. Sci. USA 1998, 95, 3621–3626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, M.Y.; Whitley, R.J.; Peng, N.; Oster, R.; Schoeb, T.R.; Sullender, W.; Ambalavanan, N.; Clancy, J.P.; Gaggar, A.; Blalock, J.E. Matrix Metalloproteinase-9 Mediates RSV Infection in Vitro and in Vivo. Viruses 2015, 7, 4230–4253. [Google Scholar] [CrossRef] [Green Version]
- Dabo, A.J.; Cummins, N.; Eden, E.; Geraghty, P. Matrix Metalloproteinase 9 Exerts Antiviral Activity against Respiratory Syncytial Virus. PLoS ONE 2015, 10, e0135970. [Google Scholar] [CrossRef]
- Rojas-Quintero, J.; Wang, X.; Tipper, J.; Burkett, P.R.; Zuniga, J.; Ashtekar, A.R.; Polverino, F.; Rout, A.; Yambayev, I.; Hernandez, C.; et al. Matrix metalloproteinase-9 deficiency protects mice from severe influenza A viral infection. JCI Insight 2018, 3, e99022. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Xu, W.; Chen, Y.; Xie, X.; Zhang, Y.; Ma, C.; Yang, Q.; Han, Y.; Zhu, C.; Xiong, Y.; et al. Matrix Metalloproteinase 9 Facilitates Hepatitis B Virus Replication through Binding with Type I Interferon (IFN) Receptor 1 To Repress IFN/JAK/STAT Signaling. J. Virol. 2017, 91, e01824-16. [Google Scholar] [CrossRef] [Green Version]
- Gutman, H.; Aftalion, M.; Melamed, S.; Politi, B.; Nevo, R.; Havusha-Laufer, S.; Achdout, H.; Gur, D.; Israely, T.; Dachir, S.; et al. Matrix Metalloproteinases Expression Is Associated with SARS-CoV-2-Induced Lung Pathology and Extracellular-Matrix Remodeling in K18-hACE2 Mice. Viruses 2022, 14, 1627. [Google Scholar] [CrossRef]
- Gelzo, M.; Cacciapuoti, S.; Pinchera, B.; De Rosa, A.; Cernera, G.; Scialo, F.; Comegna, M.; Mormile, M.; Fabbrocini, G.; Parrella, R.; et al. Matrix metalloproteinases (MMP) 3 and 9 as biomarkers of severity in COVID-19 patients. Sci. Rep. 2022, 12, 1212. [Google Scholar] [CrossRef]
- Avila-Mesquita, C.D.; Couto, A.E.S.; Campos, L.C.B.; Vasconcelos, T.F.; Michelon-Barbosa, J.; Corsi, C.A.C.; Mestriner, F.; Petroski-Moraes, B.C.; Garbellini-Diab, M.J.; Couto, D.M.S.; et al. MMP-2 and MMP-9 levels in plasma are altered and associated with mortality in COVID-19 patients. Biomed. Pharmacother. 2021, 142, 112067. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Min, J.S.; Jin, Y.-H.; Kwon, S. Auraptene Has Antiviral Activity against Human Coronavirus OC43 in MRC-5 Cells. Nutrients 2023, 15, 2960. https://doi.org/10.3390/nu15132960
Min JS, Jin Y-H, Kwon S. Auraptene Has Antiviral Activity against Human Coronavirus OC43 in MRC-5 Cells. Nutrients. 2023; 15(13):2960. https://doi.org/10.3390/nu15132960
Chicago/Turabian StyleMin, Jung Sun, Young-Hee Jin, and Sunoh Kwon. 2023. "Auraptene Has Antiviral Activity against Human Coronavirus OC43 in MRC-5 Cells" Nutrients 15, no. 13: 2960. https://doi.org/10.3390/nu15132960
APA StyleMin, J. S., Jin, Y.-H., & Kwon, S. (2023). Auraptene Has Antiviral Activity against Human Coronavirus OC43 in MRC-5 Cells. Nutrients, 15(13), 2960. https://doi.org/10.3390/nu15132960