
Dietary Protein Regulates Female Estrous Cyclicity Partially via Fibroblast Growth Factor 21

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Approval
2.2. Animals and Diets
2.3. Serum Amino Acid and Hormone Analyses
2.4. Tissue Collection
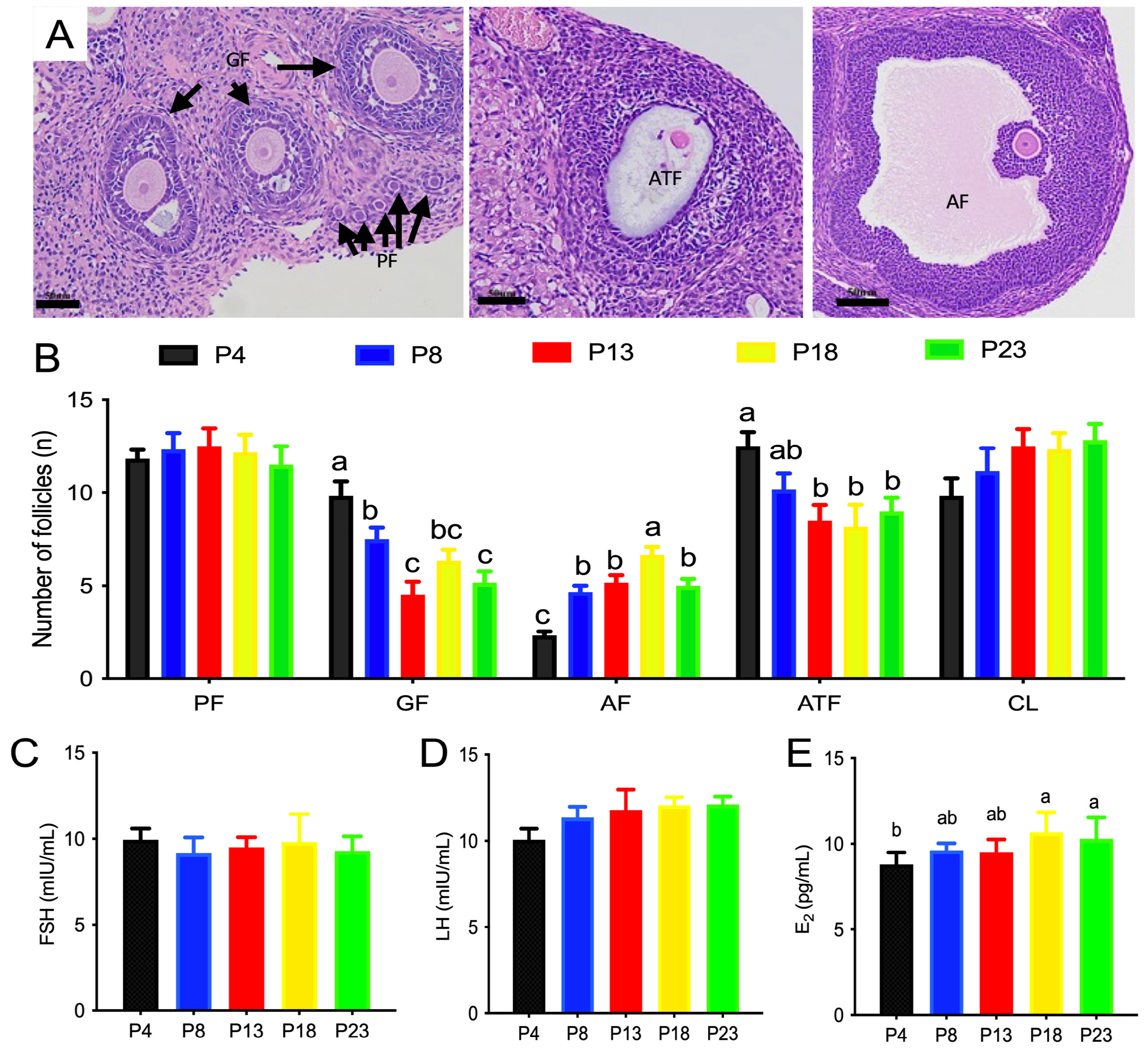
2.5. Classification of Ovarian Follicles
2.6. Re-Feeding Experiment
2.7. Real-Time Polymerase Chain Reaction Analysis
2.8. Statistical Analysis
3. Results
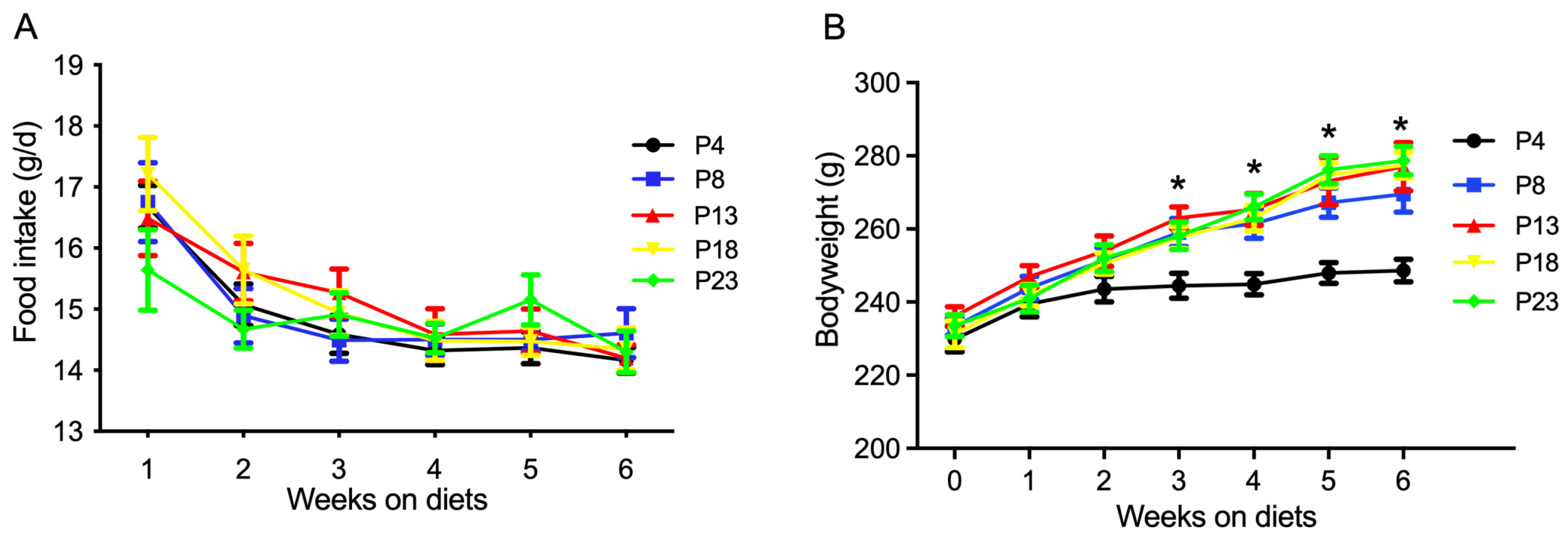
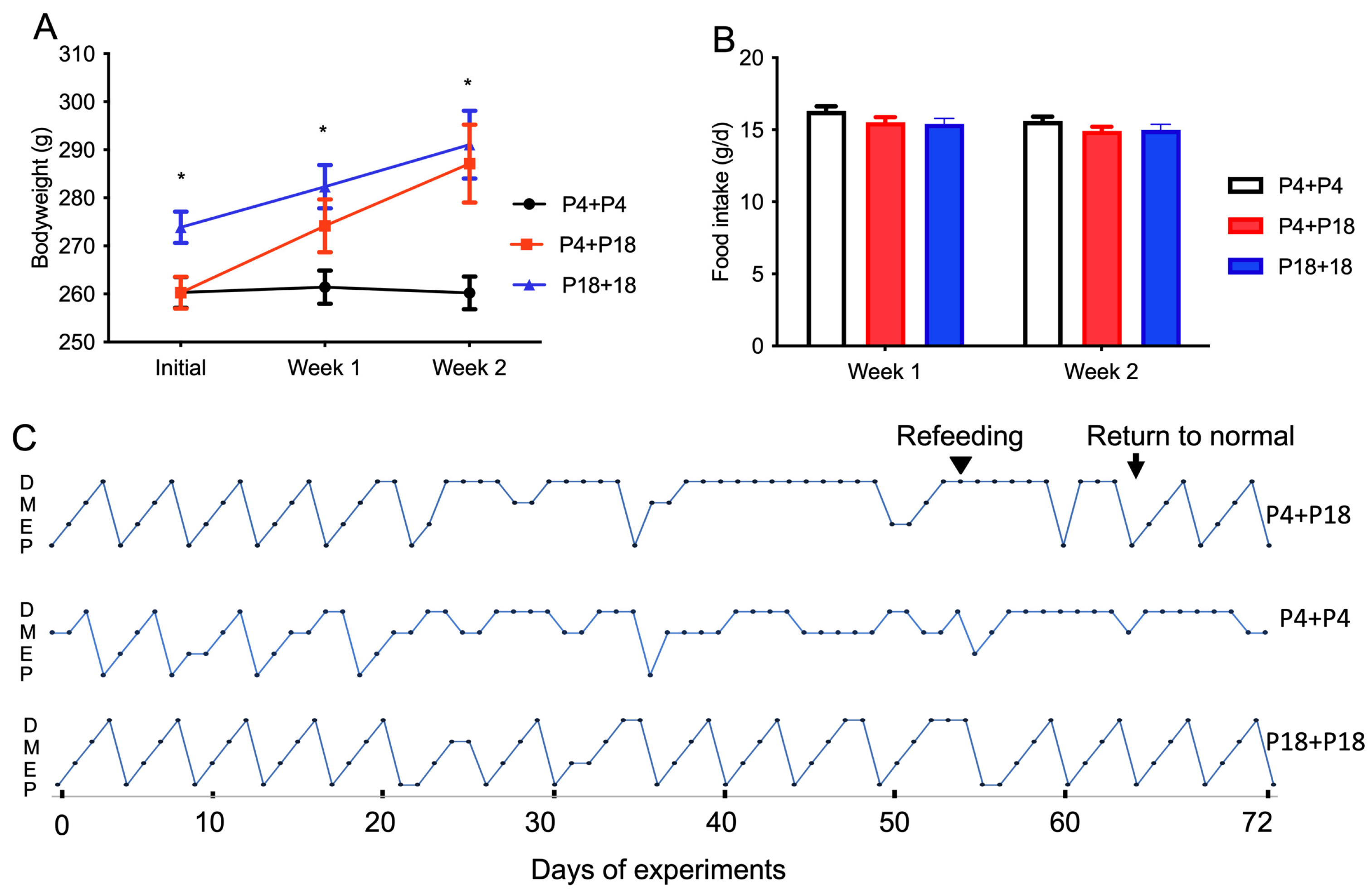
3.1. Protein Restriction or Excess Affects Bodyweight and Adipose Tissue Retention in Rats
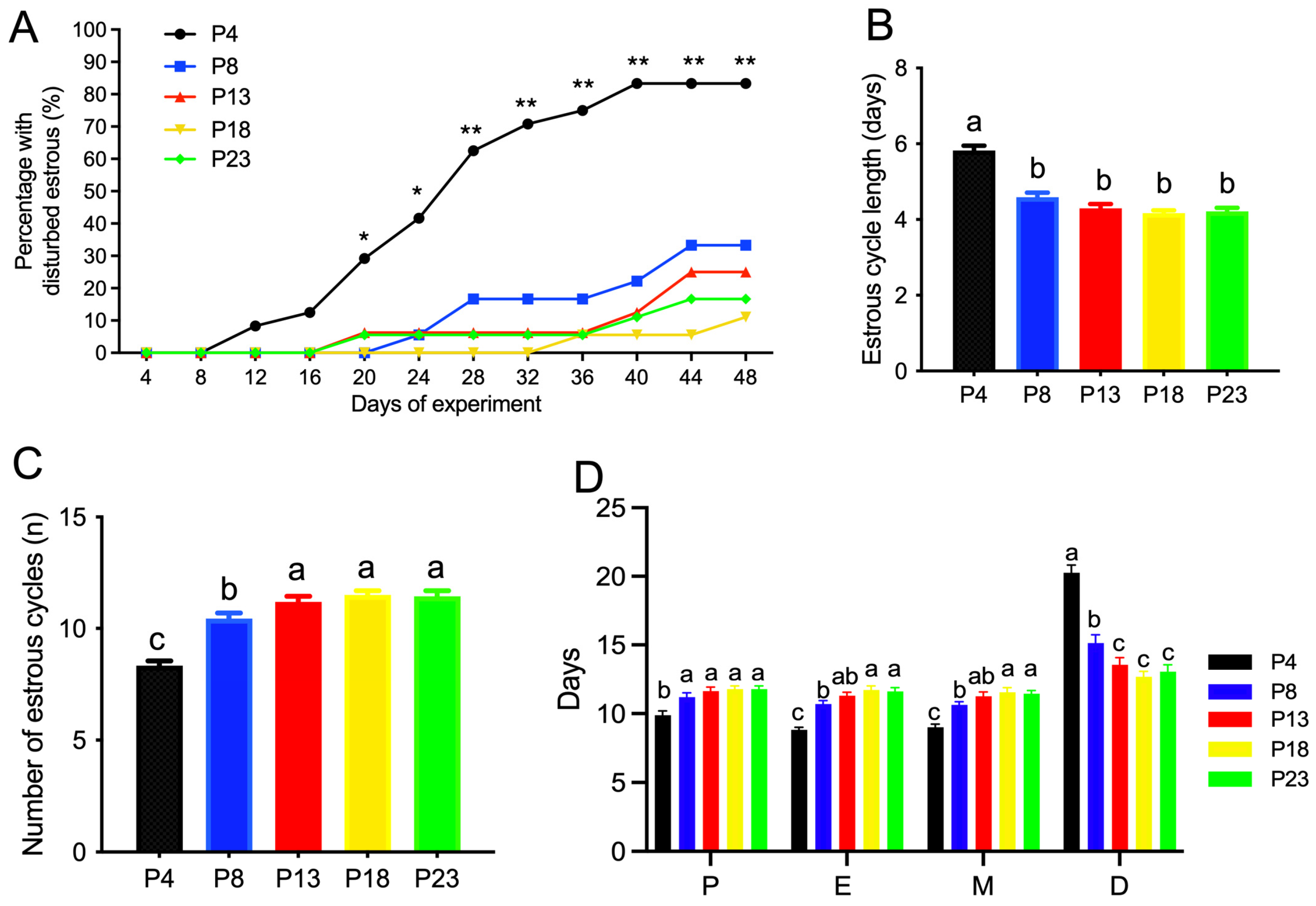
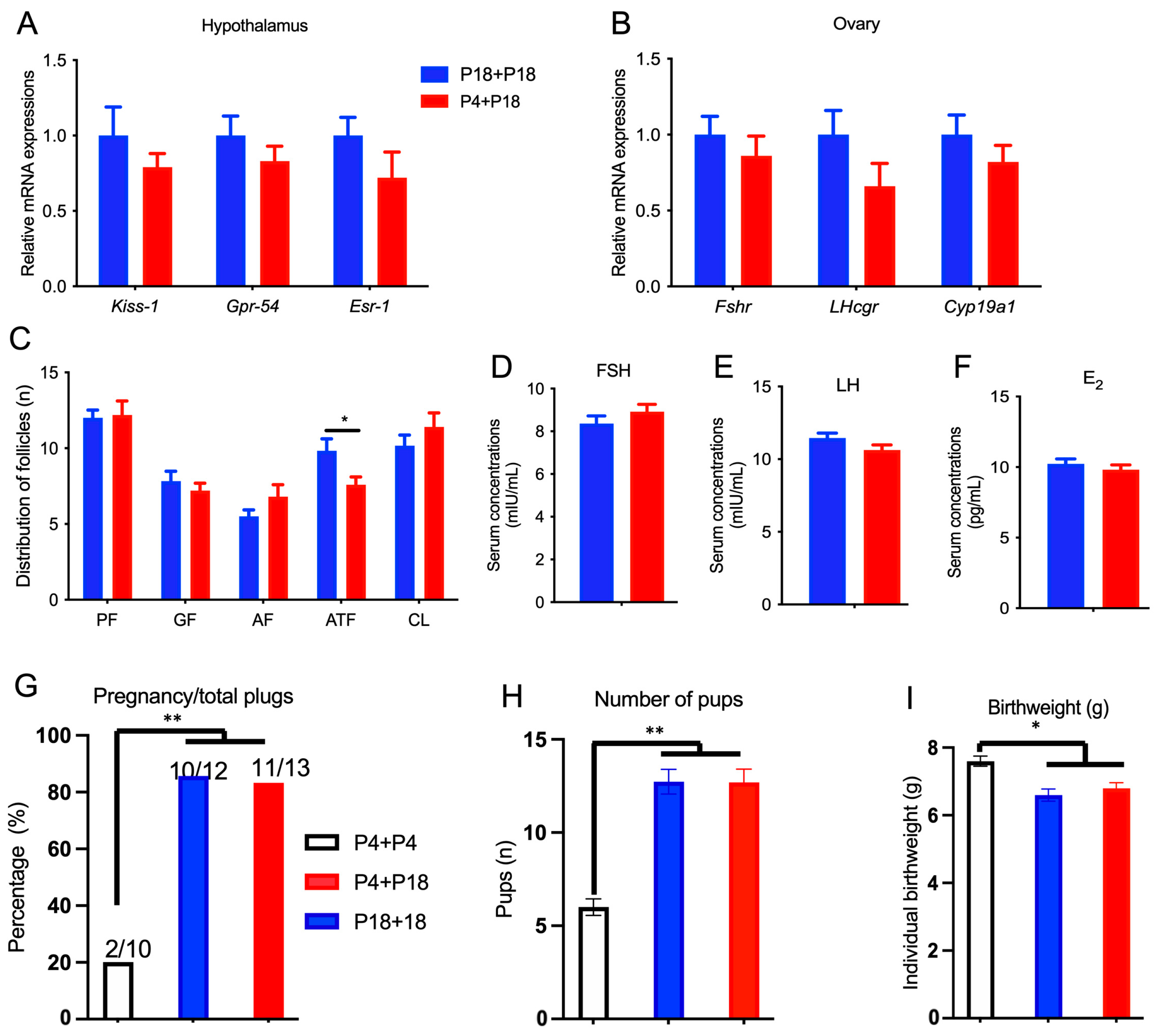
3.2. Limited Protein Intake Disrupts Estrous Cyclicity and Follicular Development
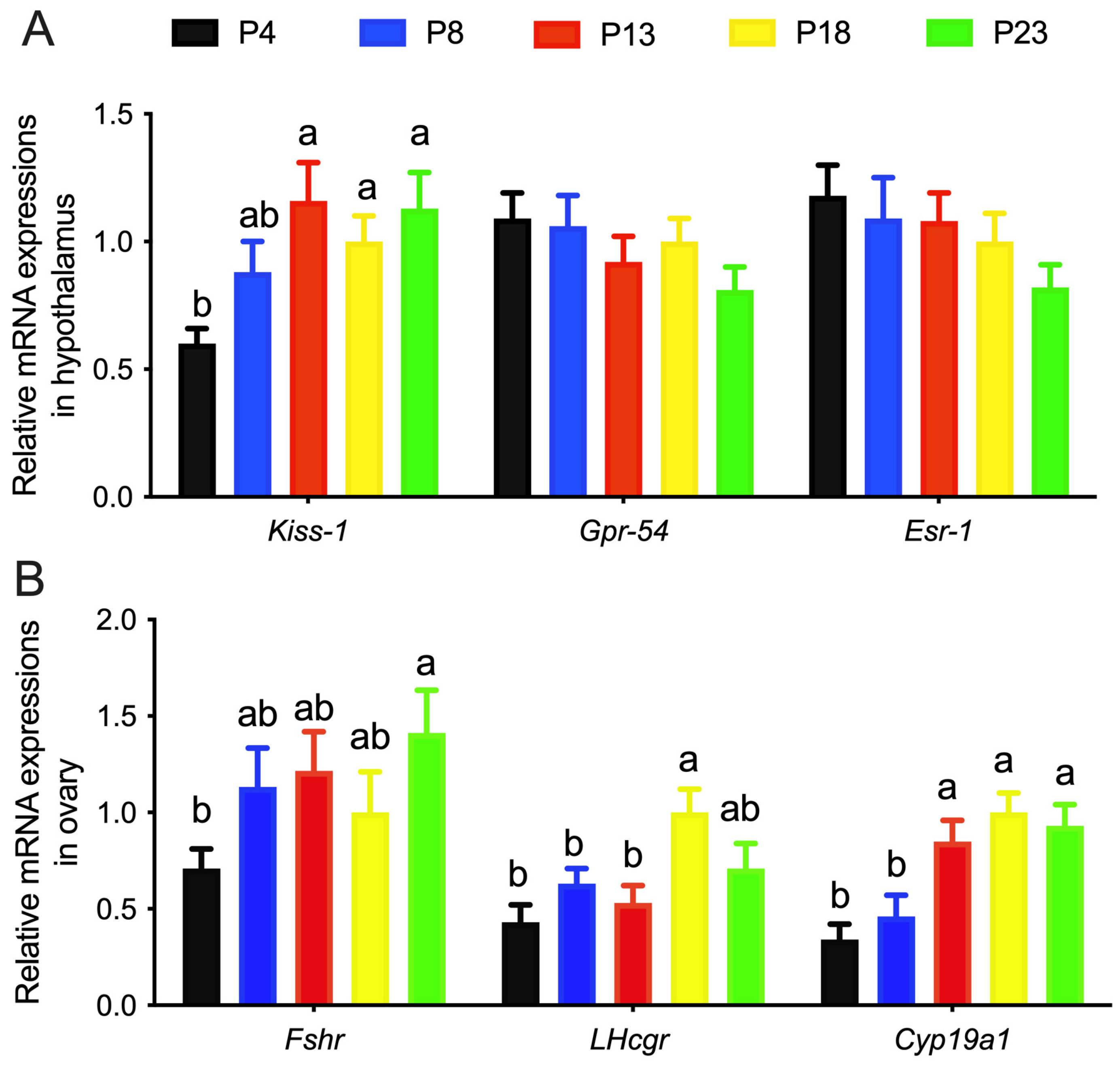
3.3. Limited Protein Intake Disrupts Gene Expression in the HPG Axis
3.4. Protein Restriction-Induced Cyclicity Disruption Is Reversed by Feeding with a Normal Diet
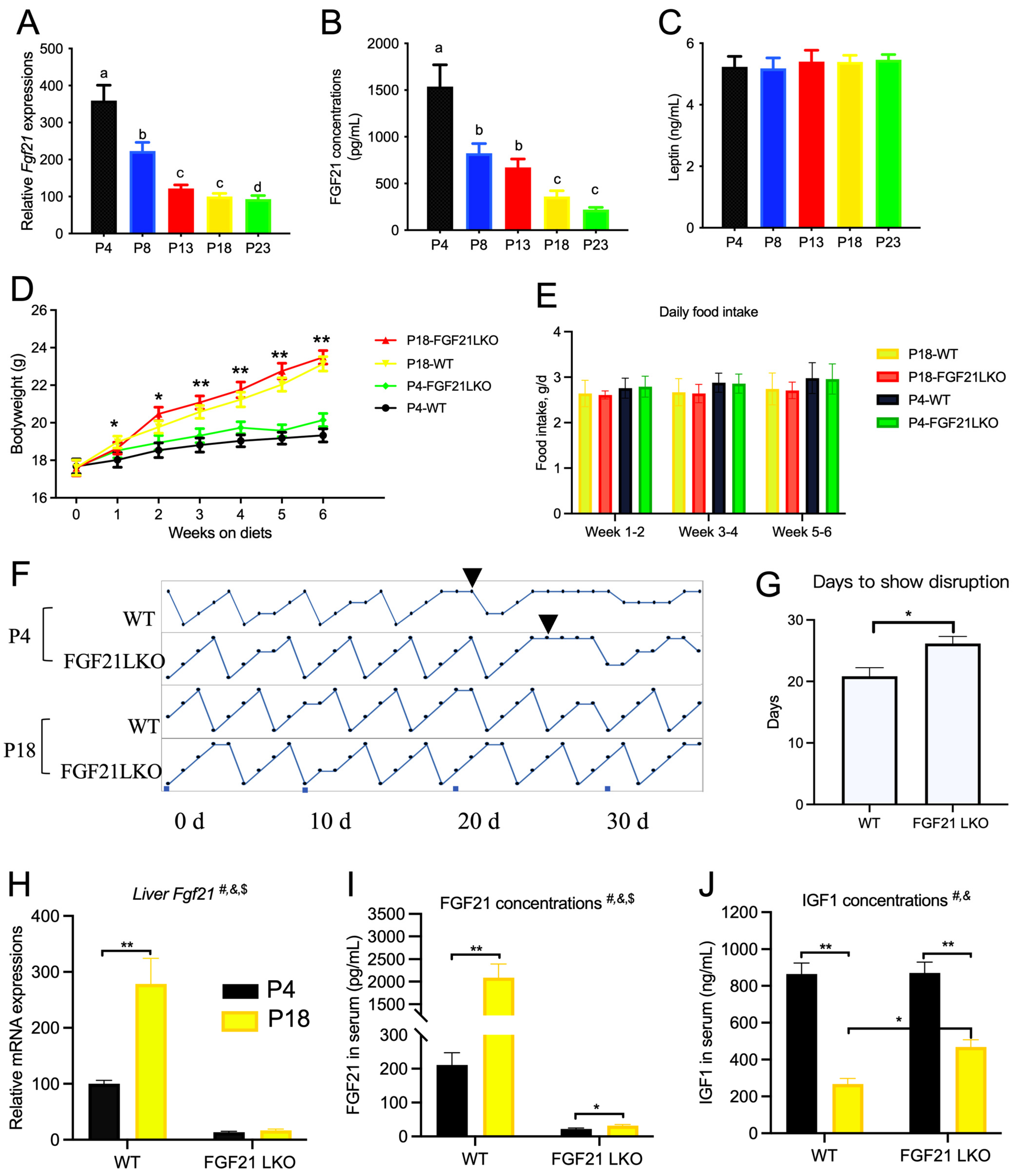
3.5. Limited Protein Intake Causes Significant Changes in Serum Amino Acid and Hormone Levels
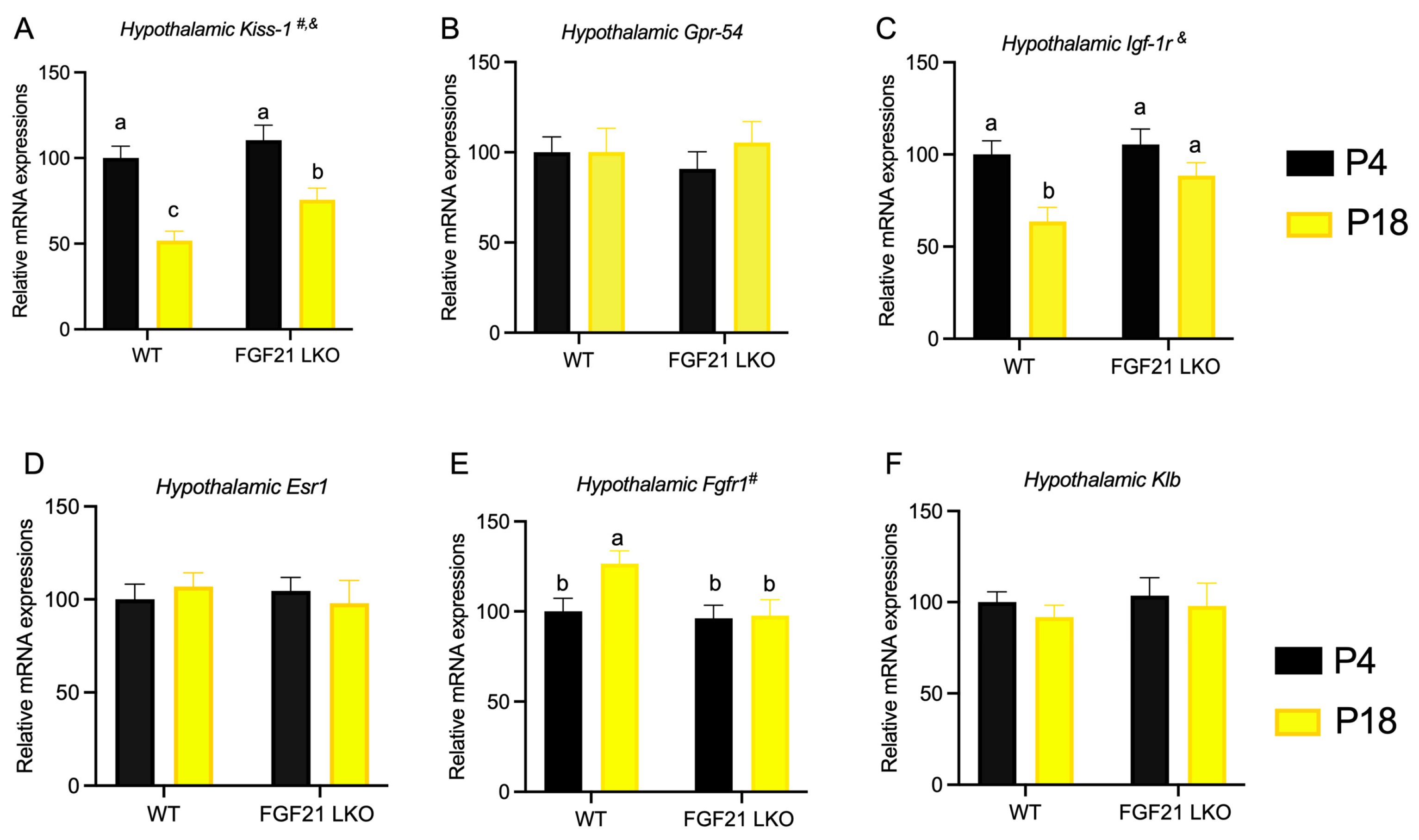
3.6. Liver FGF21 Is Partially Required for the Effect of Protein Restriction on Cyclicity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| FGF21 | factor fibroblast growth factor 21; |
| HPG | hypothalamic–pituitary–gonadal |
| GnRH | gonadotropin-releasing hormone |
| E2 | estradiol |
| GPR54 | G-protein-coupled receptor 54 |
| LH | luteinizing hormone |
| IGF-1 | insulin-like growth factor-1 |
| FGF21LKO | FGF21 liver-specific knockout |
| WT | wild-type |
| ELISA | enzyme-linked immunosorbent assay |
| EAA | essential amino acid |
| NEAA | non-essential amino acid |
References
- Ewy, M.W.; Patel, A.; Abdelmagid, M.G.; Mohamed, E.O.; Bonnes, S.L.; Salonen, B.R.; Hurt, R.T.; Mundi, M.S. Plant-Based Diet: Is It as Good as an Animal-Based Diet When It Comes to Protein? Curr. Nutr. Rep. 2022, 11, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Termannsen, A.D.; Clemmensen, K.; Thomsen, J.M.; Norgaard, O.; Diaz, L.J.; Torekov, S.S.; Quist, J.S.; Faerch, K. Effects of vegan diets on cardiometabolic health: A systematic review and meta-analysis of randomized controlled trials. Obes. Rev. 2022, 23, e13462. [Google Scholar] [CrossRef]
- Forestell, C.A.; Spaeth, A.M.; Kane, S.A. To eat or not to eat red meat. A closer look at the relationship between restrained eating and vegetarianism in college females. Appetite 2012, 58, 319–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, C.M.; Berthoud, H.R.; Munzberg, H.; Morrison, C.D. Homeostatic sensing of dietary protein restriction: A case for FGF21. Front. Neuroendocrinol. 2018, 51, 125–131. [Google Scholar] [CrossRef]
- Laeger, T.; Henagan, T.M.; Albarado, D.C.; Redman, L.M.; Bray, G.A.; Noland, R.C.; Munzberg, H.; Hutson, S.M.; Gettys, T.W.; Schwartz, M.W.; et al. FGF21 is an endocrine signal of protein restriction. J. Clin. Investig. 2014, 124, 3913–3922. [Google Scholar] [CrossRef] [Green Version]
- Laeger, T.; Albarado, D.C.; Burke, S.J.; Trosclair, L.; Hedgepeth, J.W.; Berthoud, H.R.; Gettys, T.W.; Collier, J.J.; Munzberg, H.; Morrison, C.D. Metabolic Responses to Dietary Protein Restriction Require an Increase in FGF21 that Is Delayed by the Absence of GCN2. Cell Rep. 2016, 16, 707–716. [Google Scholar] [CrossRef] [Green Version]
- Maida, A.; Zota, A.; Sjoberg, K.A.; Schumacher, J.; Sijmonsma, T.P.; Pfenninger, A.; Christensen, M.M.; Gantert, T.; Fuhrmeister, J.; Rothermel, U.; et al. A liver stress-endocrine nexus promotes metabolic integrity during dietary protein dilution. J. Clin. Investig. 2016, 126, 3263–3278. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.M.; Albarado, D.C.; Coco, L.G.; Spann, R.A.; Khan, M.S.; Qualls-Creekmore, E.; Burk, D.H.; Burke, S.J.; Collier, J.J.; Yu, S.; et al. FGF21 is required for protein restriction to extend lifespan and improve metabolic health in male mice. Nat. Commun. 2022, 13, 1897. [Google Scholar] [CrossRef]
- Levine, M.E.; Suarez, J.A.; Brandhorst, S.; Balasubramanian, P.; Cheng, C.W.; Madia, F.; Fontana, L.; Mirisola, M.G.; Guevara-Aguirre, J.; Wan, J.; et al. Low protein intake is associated with a major reduction in IGF-1, cancer, and overall mortality in the 65 and younger but not older population. Cell Metab. 2014, 19, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Patino, C.; Bossowski, J.P.; De Donatis, G.M.; Mondragon, L.; Villa, E.; Aira, L.E.; Chiche, J.; Mhaidly, R.; Lebeaupin, C.; Marchetti, S.; et al. Low-Protein Diet Induces IRE1alpha-Dependent Anticancer Immunosurveillance. Cell Metab. 2018, 27, 828–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barr, S.I. Vegetarianism and menstrual cycle disturbances: Is there an association? Am. J. Clin. Nutr. 1999, 70, 549S–554S. [Google Scholar] [CrossRef] [Green Version]
- Baines, S.; Powers, J.; Brown, W.J. How does the health and well-being of young Australian vegetarian and semi-vegetarian women compare with non-vegetarians? Public Health Nutr. 2007, 10, 436–442. [Google Scholar] [CrossRef] [Green Version]
- Dupont, J.; Reverchon, M.; Bertoldo, M.J.; Froment, P. Nutritional signals and reproduction. Mol. Cell. Endocrinol. 2014, 382, 527–537. [Google Scholar] [CrossRef]
- Lam, B.; Williamson, A.; Finer, S.; Day, F.R.; Tadross, J.A.; Goncalves, S.A.; Wade, K.; Sweeney, P.; Bedenbaugh, M.N.; Porter, D.T.; et al. MC3R links nutritional state to childhood growth and the timing of puberty. Nature 2021, 599, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Spaziani, M.; Tarantino, C.; Tahani, N.; Gianfrilli, D.; Sbardella, E.; Lenzi, A.; Radicioni, A.F. Hypothalamo-Pituitary axis and puberty. Mol. Cell. Endocrinol. 2021, 520, 111094. [Google Scholar] [CrossRef] [PubMed]
- Tena-Sempere, M. GPR54 and kisspeptin in reproduction. Hum. Reprod. Updat. 2006, 12, 631–639. [Google Scholar] [CrossRef] [Green Version]
- Sivalingam, M.; Parhar, I.S. Hypothalamic kisspeptin and kisspeptin receptors: Species variation in reproduction and reproductive behaviours. Front. Neuroendocrinol. 2022, 64, 100951. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, M.; Tsukamura, H.; Adachi, S.; Matsui, H.; Uenoyama, Y.; Iwata, K.; Yamada, S.; Inoue, K.; Ohtaki, T.; Matsumoto, H.; et al. Involvement of central metastin in the regulation of preovulatory luteinizing hormone surge and estrous cyclicity in female rats. Endocrinology 2005, 146, 4431–4436. [Google Scholar] [CrossRef] [Green Version]
- Roa, J.; Vigo, E.; Castellano, J.M.; Navarro, V.M.; Fernandez-Fernandez, R.; Casanueva, F.F.; Dieguez, C.; Aguilar, E.; Pinilla, L.; Tena-Sempere, M. Hypothalamic expression of KiSS-1 system and gonadotropin-releasing effects of kisspeptin in different reproductive states of the female Rat. Endocrinology 2006, 147, 2864–2878. [Google Scholar] [CrossRef] [Green Version]
- Childs, G.V.; Odle, A.K.; MacNicol, M.C.; MacNicol, A.M. The Importance of Leptin to Reproduction. Endocrinology 2021, 162, bqaa204. [Google Scholar] [CrossRef]
- Owen, B.M.; Bookout, A.L.; Ding, X.; Lin, V.Y.; Atkin, S.D.; Gautron, L.; Kliewer, S.A.; Mangelsdorf, D.J. FGF21 contributes to neuroendocrine control of female reproduction. Nat. Med. 2013, 19, 1153–1156. [Google Scholar] [CrossRef] [PubMed]
- Christoforidis, A.; Maniadaki, I.; Stanhope, R. Growth hormone/insulin-like growth factor-1 axis during puberty. Pediatr. Endocrinol. Rev. Per. 2005, 3, 5–10. [Google Scholar] [PubMed]
- Zhuo, Y.; Hua, L.; Feng, B.; Jiang, X.; Li, J.; Jiang, D.; Huang, X.; Zhu, Y.; Li, Z.; Yan, L.; et al. Fibroblast growth factor 21 coordinates adiponectin to mediate the beneficial effects of low-protein diet on primordial follicle reserve. EBioMedicine 2019, 41, 623–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, L.; Feng, B.; Huang, L.; Li, J.; Luo, T.; Jiang, X.; Han, X.; Che, L.; Xu, S.; Lin, Y.; et al. Time-restricted feeding improves the reproductive function of female mice via liver fibroblast growth factor 21. Clin. Transl. Med. 2020, 10, e195. [Google Scholar] [CrossRef] [PubMed]
- Hua, L.; Li, J.; Feng, B.; Jiang, D.; Jiang, X.; Luo, T.; Che, L.; Xu, S.; Lin, Y.; Fang, Z.; et al. Dietary Intake Regulates White Adipose Tissues Angiogenesis via Liver Fibroblast Growth Factor 21 in Male Mice. Endocrinology 2021, 162, bqaa244. [Google Scholar] [CrossRef]
- Hua, L.; Zhuo, Y.; Jiang, D.; Li, J.; Huang, X.; Zhu, Y.; Li, Z.; Yan, L.; Jin, C.; Jiang, X.; et al. Identification of hepatic fibroblast growth factor 21 as a mediator in 17beta-estradiol-induced white adipose tissue browning. FASEB J. 2018, 32, 5602–5611. [Google Scholar] [CrossRef] [Green Version]
- Navarro, V.M.; Castellano, J.M.; Fernandez-Fernandez, R.; Barreiro, M.L.; Roa, J.; Sanchez-Criado, J.E.; Aguilar, E.; Dieguez, C.; Pinilla, L.; Tena-Sempere, M. Developmental and hormonally regulated messenger ribonucleic acid expression of KiSS-1 and its putative receptor, GPR54, in rat hypothalamus and potent luteinizing hormone-releasing activity of KiSS-1 peptide. Endocrinology 2004, 145, 4565–4574. [Google Scholar] [CrossRef]
- Zhuo, Y.; Yang, P.; Hua, L.; Zhu, L.; Zhu, X.; Han, X.; Pang, X.; Xu, S.; Jiang, X.; Lin, Y.; et al. Effects of Chronic Exposure to Diets Containing Moldy Corn or Moldy Wheat Bran on Growth Performance, Ovarian Follicular Pool, and Oxidative Status of Gilts. Toxins 2022, 14, 413. [Google Scholar] [CrossRef]
- Liu, L.; Zhuo, Y.; Zhang, H.; Li, J.; Jiang, X.; Han, X.; Chao, J.; Feng, B.; Che, L.; Xu, S.; et al. Time-Restricted Feeding Ameliorates Uterine Epithelial Estrogen Receptor alpha Transcriptional Activity at the Time of Embryo Implantation in Mice Fed a High-Fat Diet. J. Nutr. 2023, 153, 1753–1761. [Google Scholar] [CrossRef]
- Catalano, P.M.; Shankar, K. Obesity and pregnancy: Mechanisms of short term and long term adverse consequences for mother and child. BMJ 2017, 356, j1. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; O’Kane, A.M.; Zhang, Y.; Ren, J. Maternal obesity and offspring health: Adapting metabolic changes through autophagy and mitophagy. Obes. Rev. 2023, 24, e13567. [Google Scholar] [CrossRef]
- Pedersen, A.B.; Bartholomew, M.J.; Dolence, L.A.; Aljadir, L.P.; Netteburg, K.L.; Lloyd, T. Menstrual differences due to vegetarian and nonvegetarian diets. Am. J. Clin. Nutr. 1991, 53, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Barr, S.I.; Janelle, K.C.; Prior, J.C. Vegetarian vs nonvegetarian diets, dietary restraint, and subclinical ovulatory disturbances: Prospective 6-mo study. Am. J. Clin. Nutr. 1994, 60, 887–894. [Google Scholar] [CrossRef]
- Du, F.; Higginbotham, D.A.; White, B.D. Food intake, energy balance and serum leptin concentrations in rats fed low-protein diets. J. Nutr. 2000, 130, 514–521. [Google Scholar] [CrossRef] [Green Version]
- Filho, J.C.; Hazel, S.J.; Anderstam, B.; Bergstrom, J.; Lewitt, M.; Hall, K. Effect of protein intake on plasma and erythrocyte free amino acids and serum IGF-I and IGFBP-1 levels in rats. Am. J. Physiol. 1999, 277, E693–E701. [Google Scholar] [CrossRef]
- Moro, J.; Chaumontet, C.; Even, P.C.; Blais, A.; Piedcoq, J.; Gaudichon, C.; Tomé, D.; Azzout-Marniche, D. Severe protein deficiency induces hepatic expression and systemic level of FGF21 but inhibits its hypothalamic expression in growing rats. Sci. Rep. 2021, 11, 12436. [Google Scholar] [CrossRef] [PubMed]
- Fontana, R.; Della, T.S. The Deep Correlation between Energy Metabolism and Reproduction: A View on the Effects of Nutrition for Women Fertility. Nutrients 2016, 8, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remesar, X.; Alemany, M. Dietary Energy Partition: The Central Role of Glucose. Int. J. Mol. Sci. 2020, 21, 7729. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, Y.; Zhou, D.; Che, L.; Fang, Z.; Lin, Y.; Wu, D. Feeding prepubescent gilts a high-fat diet induces molecular changes in the hypothalamus-pituitary-gonadal axis and predicts early timing of puberty. Nutrition 2014, 30, 890–896. [Google Scholar] [CrossRef]
- Parra-Vargas, M.; Ramon-Krauel, M.; Lerin, C.; Jimenez-Chillaron, J.C. Size Does Matter: Litter Size Strongly Determines Adult Metabolism in Rodents. Cell Metab. 2020, 32, 334–340. [Google Scholar] [CrossRef]
- Maida, A.; Zota, A.; Vegiopoulos, A.; Appak-Baskoy, S.; Augustin, H.G.; Heikenwalder, M.; Herzig, S.; Rose, A.J. Dietary protein dilution limits dyslipidemia in obesity through FGF21-driven fatty acid clearance. J. Nutr. Biochem. 2018, 57, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Messina, A.; Somm, E.; Miraoui, H.; Kinnunen, T.; Acierno, J.J.; Niederlander, N.J.; Bouilly, J.; Dwyer, A.A.; Sidis, Y.; et al. KLB, encoding beta-Klotho, is mutated in patients with congenital hypogonadotropic hypogonadism. EMBO Mol. Med. 2017, 9, 1379–1397. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, T.; Lin, V.Y.; Goetz, R.; Mohammadi, M.; Mangelsdorf, D.J.; Kliewer, S.A. Inhibition of growth hormone signaling by the fasting-induced hormone FGF21. Cell Metab. 2008, 8, 77–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.Y.; Ma, J.N.; Shen, Y.Y.; Xie, X.R.; Ren, W. Effects of Growth Hormone on Adult Human Gonads: Action on Reproduction and Sexual Function. Int. J. Endocrinol. 2023, 2023, 7492696. [Google Scholar] [CrossRef]
- Della, T.S.; Rando, G.; Meda, C.; Stell, A.; Chambon, P.; Krust, A.; Ibarra, C.; Magni, P.; Ciana, P.; Maggi, A. Amino acid-dependent activation of liver estrogen receptor alpha integrates metabolic and reproductive functions via IGF-1. Cell Metab. 2011, 13, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Luque, R.M.; Kineman, R.D.; Tena-Sempere, M. Regulation of hypothalamic expression of KiSS-1 and GPR54 genes by metabolic factors: Analyses using mouse models and a cell line. Endocrinology 2007, 148, 4601–4611. [Google Scholar] [CrossRef]
- Hiney, J.K.; Srivastava, V.K.; Pine, M.D.; Les, D.W. Insulin-like growth factor-I activates KiSS-1 gene expression in the brain of the prepubertal female rat. Endocrinology 2009, 150, 376–384. [Google Scholar] [CrossRef]
- Ahima, R.S. Body fat, leptin, and hypothalamic amenorrhea. N. Engl. J. Med. 2004, 351, 959–962. [Google Scholar] [CrossRef]
- Narita, K.; Nagao, K.; Bannai, M.; Ichimaru, T.; Nakano, S.; Murata, T.; Higuchi, T.; Takahashi, M. Dietary deficiency of essential amino acids rapidly induces cessation of the rat estrous cycle. PLoS ONE 2011, 6, e28136. [Google Scholar] [CrossRef] [Green Version]
- Yudkoff, M.; Daikhin, Y.; Nissim, I.; Horyn, O.; Luhovyy, B.; Luhovyy, B.; Lazarow, A.; Nissim, I. Brain amino acid requirements and toxicity: The example of leucine. J. Nutr. 2005, 135, 1531S–1538S. [Google Scholar] [CrossRef] [Green Version]
- Xie, Q.; Kang, Y.; Zhang, C.; Xie, Y.; Wang, C.; Liu, J.; Yu, C.; Zhao, H.; Huang, D. The Role of Kisspeptin in the Control of the Hypothalamic-Pituitary-Gonadal Axis and Reproduction. Front. Endocrinol. 2022, 13, 925206. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | P4 | P8 | P13 | P18 | P23 | SEM |
|---|---|---|---|---|---|---|
| BW (g) | 249.24 b | 269.98 a | 277.16 a | 279.53 a | 279.26 a | 3.19 |
| Ovary (g) | 0.035 c | 0.045 b | 0.050 b | 0.058 a | 0.061 a | 0.003 |
| Ovary (% BW) | 0.014 d | 0.017 cd | 0.018 bc | 0.021 ab | 0.022 a | 0.001 |
| Uterus (g) | 0.50 c | 0.61 b | 0.65 ab | 0.73 a | 0.64 ab | 0.04 |
| Uterus (% BW) | 0.20 b | 0.23 ab | 0.24 ab | 0.26 a | 0.23 ab | 0.01 |
| Fat pads (g) | 13.28 ab | 13.88 a | 13.39 ab | 12.83 ab | 10.99 b | 0.90 |
| Fat pads (% BW) | 5.31 a | 5.14 a | 4.81 ab | 4.56 ab | 3.92 b | 0.31 |
| Liver (g) | 6.74 b | 7.41 a | 7.95 a | 7.75 a | 8.00 a | 0.21 |
| Liver (% BW) | 2.71 | 2.74 | 2.88 | 2.77 | 2.87 | 0.08 |
| Parameters | P4 | P8 | P13 | P18 | P23 | SEM |
|---|---|---|---|---|---|---|
| Essential, μmol/L | ||||||
| Threonine | 280.31 b | 478.83 a | 481.81 a | 427.99 a | 524.74 a | 26.80 |
| Valine | 162.61 b | 219.32 a | 214.76 a | 227.42 a | 221.84 a | 10.38 |
| Methionine | 47.20 | 47.70 | 45.18 | 46.29 | 54.75 | 3.71 |
| Isoleucine | 100.74 | 125.03 | 113.89 | 136.30 | 131.39 | 12.82 |
| Leucine | 154.75 b | 230.03 a | 227.18 a | 277.60 a | 252.56 a | 21.32 |
| Phenylalanine | 88.97 c | 94.88 bc | 95.65 bc | 109.59 a | 107.88 ab | 3.41 |
| Lysine | 437.24 b | 440.75 b | 466.58 ab | 505.47 ab | 629.22 a | 53.14 |
| Total | 1271.81 c | 1636.54 b | 1645.05 ab | 1730.67 ab | 1922.38 a | 82.66 |
| Nonessential, μmol/L | ||||||
| Asparagine | 56.58 | 60.46 | 67.59 | 66.36 | 62.77 | 6.25 |
| Serine | 741.73 a | 499.04 bc | 524.92 b | 380.37 d | 414.87 cd | 22.44 |
| Glutamate | 291.07 a | 277.28 a | 218.28 b | 218.16 b | 209.49 b | 13.95 |
| Glycine | 842.13 a | 419.05 b | 310.20 c | 312.43 c | 305.64 c | 18.80 |
| Alanine | 1047.00 a | 807.59 b | 491.24 c | 600.53 c | 615.79 c | 36.01 |
| Citrulline | 127.93 a | 91.57 b | 86.20 b | 89.39 b | 76.50 b | 6.55 |
| Tyrosine | 43.96 c | 55.96 bc | 66.41 b | 70.48 b | 95.88 a | 4.75 |
| Ornithine | 139.27 | 116.99 | 127.92 | 97.78 | 97.62 | 16.26 |
| Histidine | 68.67 | 69.57 | 58.04 | 64.99 | 62.59 | 4.82 |
| Arginine | 335.45 | 241.22 | 254.91 | 265.83 | 273.32 | 31.49 |
| Proline | 439.64 a | 256.90 b | 143.13 c | 149.44 c | 118.44 c | 17.03 |
| Total | 4133.43 a | 2895.62 b | 2348.83 c | 2315.75 c | 2332.90 c | 87.76 |
| EAA/NEAA | 0.31 d | 0.56 c | 0.70 b | 0.75 ab | 0.83 a | 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Y.; Yang, M.; Song, J.; Jiang, X.; Xu, S.; Che, L.; Fang, Z.; Lin, Y.; Jin, C.; Feng, B.; et al. Dietary Protein Regulates Female Estrous Cyclicity Partially via Fibroblast Growth Factor 21. Nutrients 2023, 15, 3049. https://doi.org/10.3390/nu15133049
Cao Y, Yang M, Song J, Jiang X, Xu S, Che L, Fang Z, Lin Y, Jin C, Feng B, et al. Dietary Protein Regulates Female Estrous Cyclicity Partially via Fibroblast Growth Factor 21. Nutrients. 2023; 15(13):3049. https://doi.org/10.3390/nu15133049
Chicago/Turabian StyleCao, Yaxue, Min Yang, Jie Song, Xuemei Jiang, Shengyu Xu, Lianqiang Che, Zhengfeng Fang, Yan Lin, Chao Jin, Bin Feng, and et al. 2023. "Dietary Protein Regulates Female Estrous Cyclicity Partially via Fibroblast Growth Factor 21" Nutrients 15, no. 13: 3049. https://doi.org/10.3390/nu15133049
APA StyleCao, Y., Yang, M., Song, J., Jiang, X., Xu, S., Che, L., Fang, Z., Lin, Y., Jin, C., Feng, B., Wu, D., Hua, L., & Zhuo, Y. (2023). Dietary Protein Regulates Female Estrous Cyclicity Partially via Fibroblast Growth Factor 21. Nutrients, 15(13), 3049. https://doi.org/10.3390/nu15133049