
Exercise Equals the Mobilization of Visceral versus Subcutaneous Adipose Fatty Acid Molecules in Fasted Rats Associated with the Modulation of the AMPK/ATGL/HSL Axis
 ,
,  ,
, 
 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animal Handling
2.3. Lipid Analysis
2.4. GC-FID Analysis of FAMEs
2.5. Western Immunoblot Analysis
2.6. Statistical Analysis
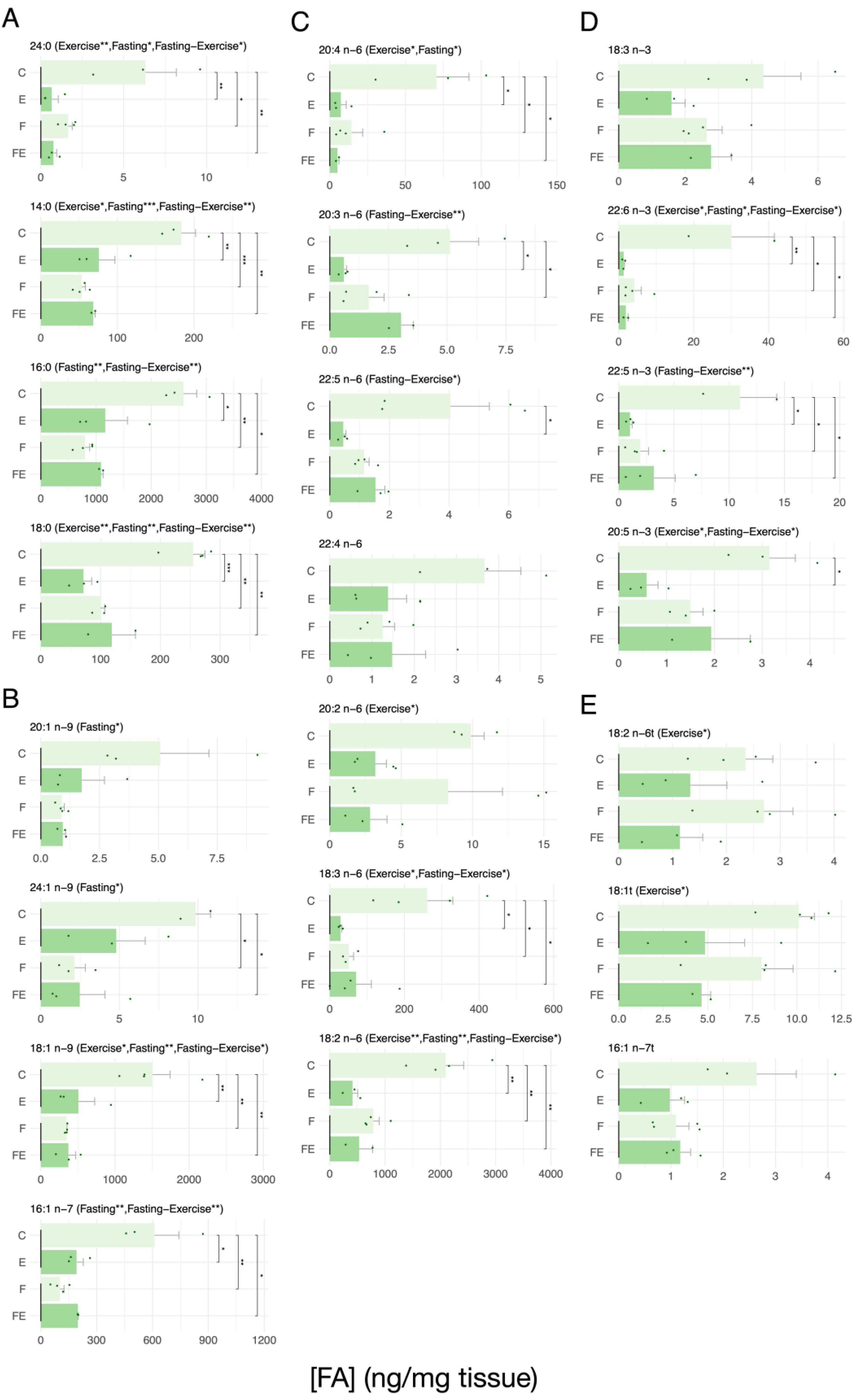
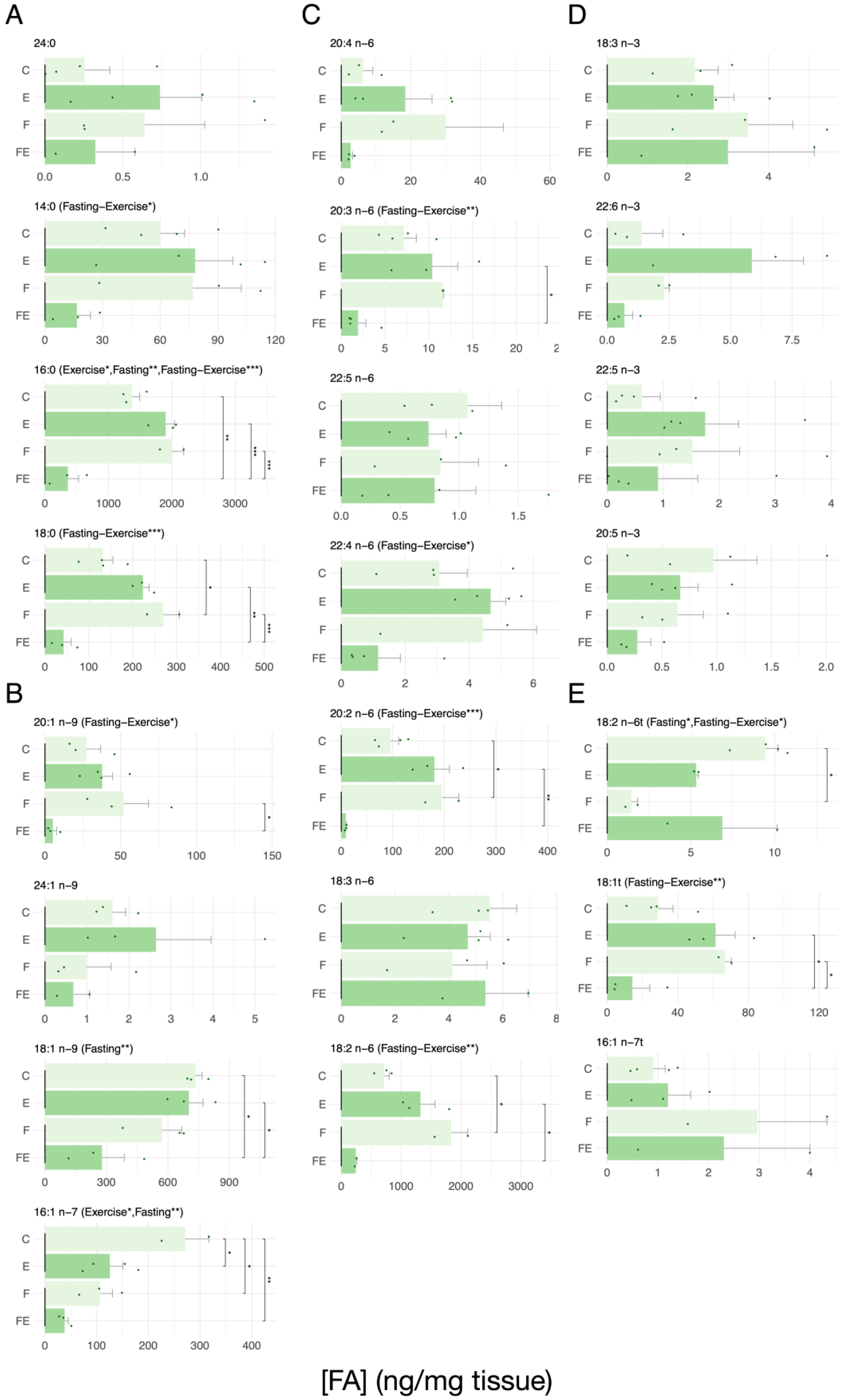
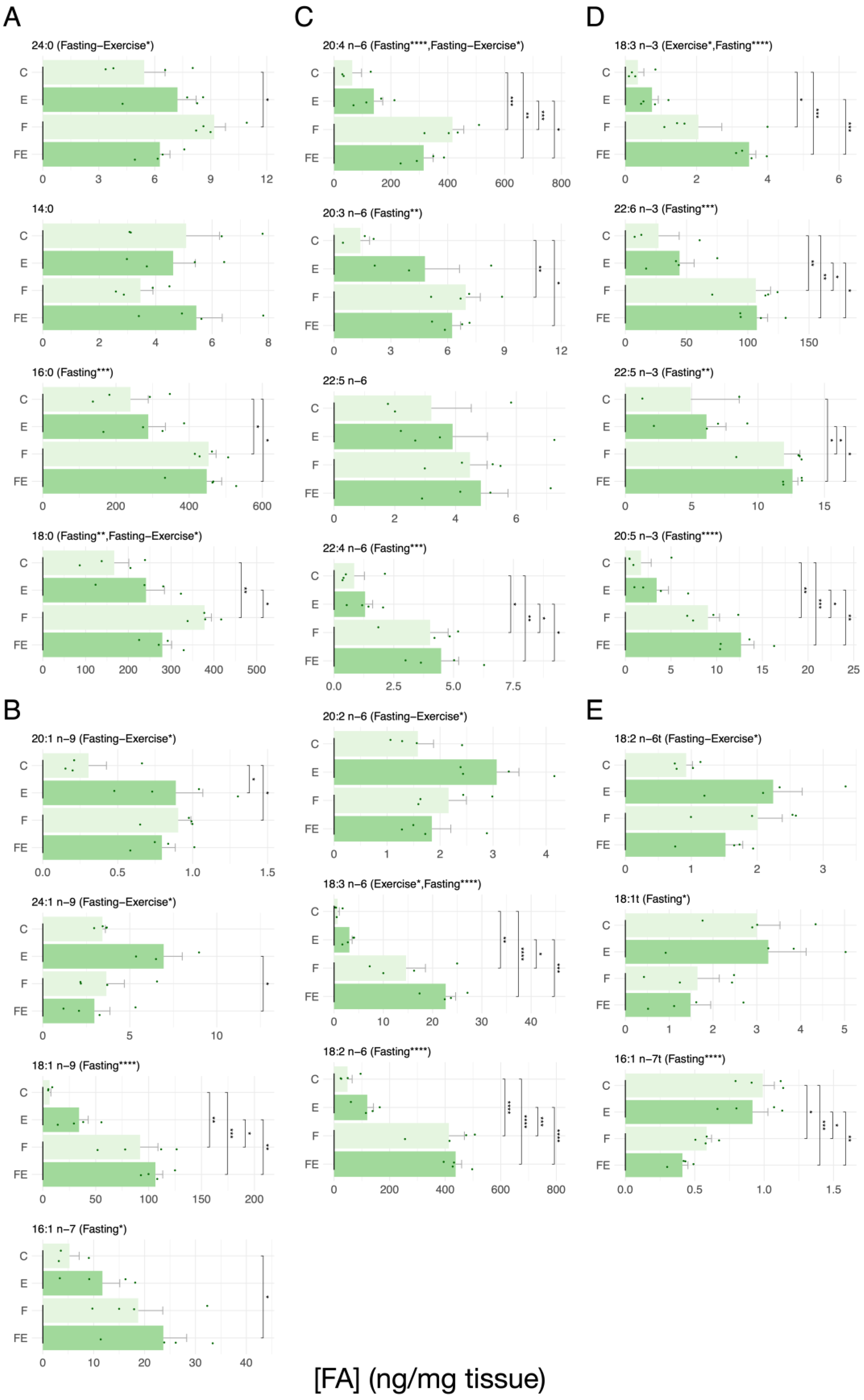
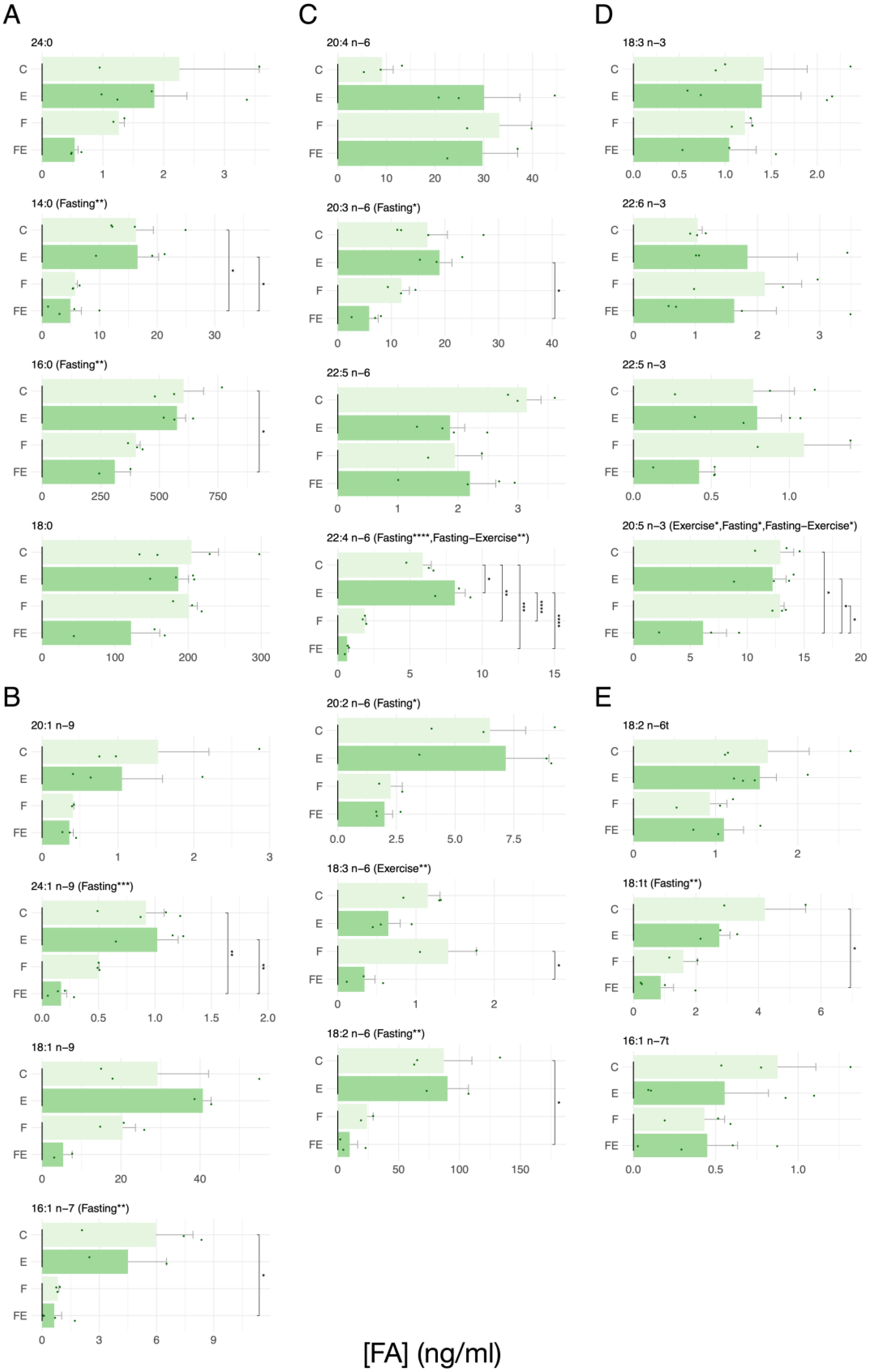
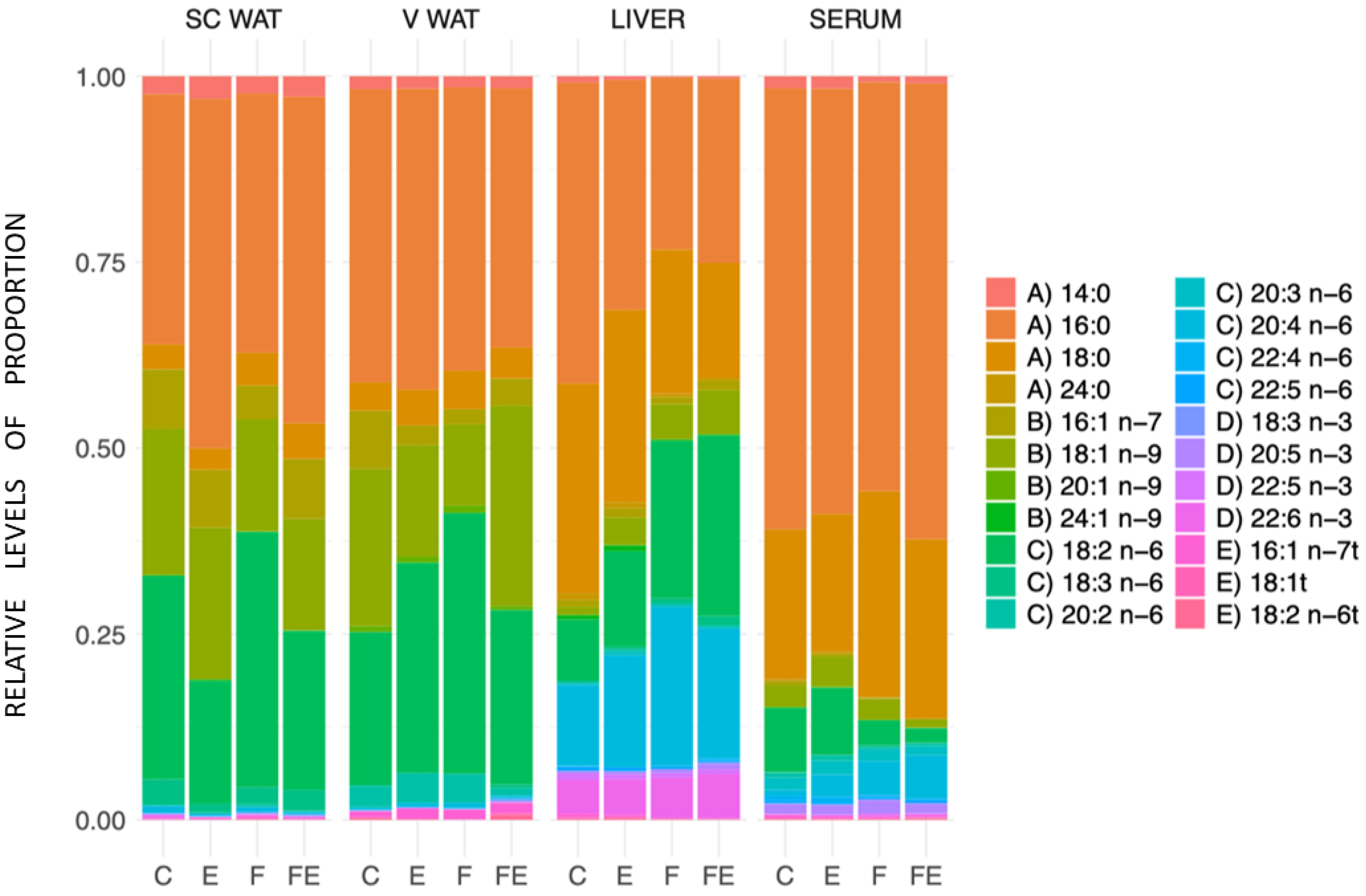
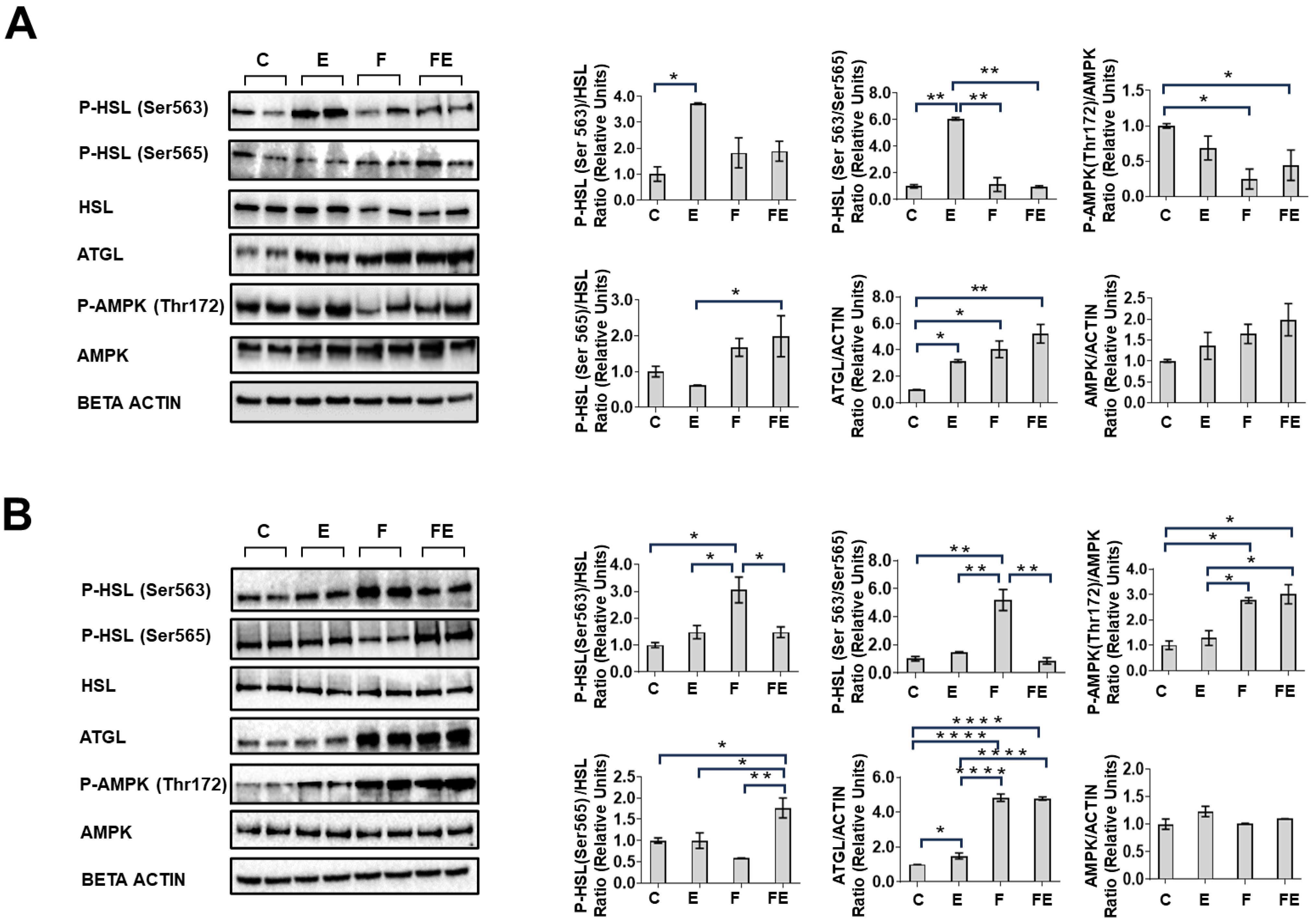
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Jaspers, R.T.; Zillikens, M.C.; Friesema, E.C.M.; delli Paoli, G.; Bloch, W.; Uitterlinden, A.G.; Goglia, F.; Lanni, A.; de Lange, P. Exercise, fasting, and mimetics: Toward beneficial combinations? FASEB J. 2017, 31, 14–28. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzo, A.; Bianchi, A.; Maroni, P.; Iannarelli, A.; Di Daniele, N.; Iacopino, L.; Di Renzo, L. Adiposity rather than BMI determines metabolic risk. Int. J. Cardiol. 2013, 166, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.R.; Braverman, E.R. Measuring adiposity in patients: The utility of BMI, percentage of body fat, and leptin. PLoS ONE 2012, 7, e33308. [Google Scholar] [CrossRef]
- Haganes, K.L.; Silva, C.P.; Eyjólfsdóttir, S.K.; Steen, S.; Grindberg, M.; Lydersen, S.; Hawley, J.A.; Moholdt, T. Time-restricted eating and exercise training improve HbA1c and body composition in women with overweight/obesity: A randomized con-trolled trial. Cell Metab. 2022, 34, 1457–1471.e4. [Google Scholar] [CrossRef]
- Merlotti, C.; Ceriani, V.; Morabito, A.; Pontiroli, A.E. Subcutaneous fat loss is greater than visceral fat loss with diet and exercise, weight-loss promoting drugs and bariatric surgery: A critical review and meta-analysis. Int. J. Obes. 2017, 41, 672–682. [Google Scholar] [CrossRef]
- Mittendorfer, B.; Magkos, F.; Fabbrini, E.; Mohammed, B.S.; Klein, S. Relationship between Body Fat Mass and Free Fatty Acid Kinetics in Men and Women. Obesity 2009, 17, 1872–1877. [Google Scholar] [CrossRef]
- Raclot, T.; Groscolas, R. Selective mobilization of adipose tissue fatty acids during energy depletion in the rat. J. Lipid Res. 1995, 36, 2164–2173. [Google Scholar] [CrossRef]
- Petridou, A.; Nikolaidis, M.G.; Matsakas, A.; Schulz, T.; Michna, H.; Mougios, V. Effect of exercise training on the fatty acid composition of lipid classes in rat liver, skeletal muscle, and adipose tissue. Eur. J. Appl. Physiol. 2005, 94, 84–92. [Google Scholar] [CrossRef]
- Deng, J.; Feng, D.; Jia, X.; Zhai, S.; Liu, Y.; Gao, N.; Zhang, X.; Li, M.; Lu, M.; Liu, C.; et al. Efficacy and mechanism of intermittent fasting in metabolic associated fatty liver disease based on ultraperformance liquid chromatography-tandem mass spectrometry. Front. Nutr. 2022, 9, 838091. [Google Scholar] [CrossRef] [PubMed]
- Kulminskaya, N.; Oberer, M. Protein-protein interactions regulate the activity of Adipose Triglyceride Lipase in intracellular lipolysis. Biochimie 2020, 169, 62–68. [Google Scholar] [CrossRef]
- Kersten, S. The impact of fasting on adipose tissue metabolism. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2023, 1868, 159262. [Google Scholar] [CrossRef] [PubMed]
- Giacco, A.; Delli Paoli, G.; Simiele, R.; Caterino, M.; Ruoppolo, M.; Bloch, W.; Kraaij, R.; Uitterlinden, A.G.; Santillo, A.; Senese, R.; et al. Exercise with fasting at thermoneutrality impacts fuel use, the microbiome, AMPK phosphorylation, muscle fibers, and thyroid hormone levels in rats. Physiol. Rep. 2020, 8(3), e14354. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Wei, C.; Tan, Y.; Dong, X.; Yang, Z.; Yan, J.; Luo, X. Both prolonged high-fat diet consumption and calorie restriction boost hepatic NAD+ metabolism in mice. J. Nutr. Biochem. 2023, 115, 109296. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gao, T.; Deng, S.; Shang, L.; Chen, X.; Chen, K.; Li, P.; Cui, X.; Zeng, J. Fasting induces hepatic lipid accumulation by stimulating peroxisomal dicarboxylic acid oxidation. J. Biol. Chem. 2021, 296, 100622. [Google Scholar] [CrossRef]
- Petersen, M.C.; Vatner, D.F.; Shulman, G.I. Regulation of hepatic glucose metabolism in health and disease. Nat. Rev. Endocrinol. 2017, 13, 572–587. [Google Scholar] [CrossRef] [PubMed]
- Petersen, K.F.; Dufour, S.; Cline, G.W.; Shulman, G.I. Regulation of hepatic mitochondrial oxidation by glucose-alanine cy-cling during starvation in humans. J. Clin. Investig. 2019, 129, 4671–4675. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.X.; Wang, J. A simplified method for analysis of polyunsaturated fatty acids. BMC Biochem. 2005, 6, 5. [Google Scholar] [CrossRef]
- Ali, S.; Intrieri, M.; Pisanti, A.; Cardinale, G.; Corbi, G.; Scapagnini, G.; Davinelli, S. Determination of n-3 index and arachidonic acid/eicosapentaenoic acid ratio in dried blood spot by gas chromatography. Biotechniques 2022, 73, 25–33. [Google Scholar] [CrossRef]
- Ali, S.; Aiello, A.; Zotti, T.; Accardi, G.; Cardinale, G.; Vito, P.; Calabrò, A.; Ligotti, M.E.; Intrieri, M.; Corbi, G.; et al. Age-associated changes in circulatory fatty acids: New insights on adults and long-lived individuals. Geroscience 2023, 45, 781–796. [Google Scholar] [CrossRef]
- R: The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 1 February 2022).
- Redman, L.M.; Heilbronn, L.K.; Martin, C.K.; Alfonso, A.; Smith, S.R.; Ravussin, E. Effect of Calorie Restriction with or without Exercise on Body Composition and Fat Distribution. J. Clin. Endocrinol. Metab. 2007, 92, 865–872. [Google Scholar] [CrossRef]
- Raclot, T.; Mioskowski, E.; Bach, A.C.; Groscolas, R. Selectivity of fatty acid mobilization: A general metabolic feature of adipose tissue. Am. J. Physiol. Content 1995, 269, R1060–R1067. [Google Scholar] [CrossRef] [PubMed]
- de Lange, P.; Farina, P.; Moreno, M.; Ragni, M.; Lombardi, A.; Silvestri, E.; Burrone, L.; Lanni, A.; Goglia, F. Sequential changes in the signal transduction responses of skeletal muscle following food deprivation. FASEB J. 2006, 20, 2579–2581. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Wang, H.; Wu, F.; Yang, C.; Zhang, H.; Xu, Z.; Guo, Y.; Guo, Z.; Xin, B.; Ma, T.; et al. Hepatic metabolite responses to 4-day complete fasting and subsequent refeeding in rats. PeerJ 2022, 10, e14009. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.W.; Cannon, B.; Nedergaard, J. The answer to the question “What is the best housing temperature to translate mouse experiments to humans?” is: Thermoneutrality. Mol. Metab. 2019, 26, 1–3. [Google Scholar] [CrossRef]
- Marks, K.A.; Marvyn, P.M.; Henao, J.J.A.; Bradley, R.M.; Stark, K.D.; Duncan, R.E. Fasting enriches liver triacylglycerol with n-3 polyunsaturated fatty acids: Implications for understanding the adipose–liver axis in serum docosahexaenoic acid regulation. Genes Nutr. 2015, 10, 39. [Google Scholar] [CrossRef]
- Giacco, A.; Cioffi, F.; Cuomo, A.; Simiele, R.; Senese, R.; Silvestri, E.; Amoresano, A.; Fontanarosa, C.; Petito, G.; Moreno, M.; et al. Mild Endurance Exercise during Fasting Increases Gastrocnemius Muscle and Prefrontal Cortex Thyroid Hormone Levels through Differential BHB and BCAA-Mediated BDNF-mTOR Signaling in Rats. Nutrients 2022, 14, 1166. [Google Scholar] [CrossRef]
- Santillo, A.; Giacco, A.; Falvo, S.; Russo, F.D.G.; Senese, R.; Di Fiore, M.M.; Baccari, G.C.; Lanni, A.; de Lange, P. Mild Exercise Rescues Steroidogenesis and Spermatogenesis in Rats Submitted to Food Withdrawal. Front. Endocrinol. 2020, 11, 302. [Google Scholar] [CrossRef]
- Delli Paoli, G.; van de Laarschot, D.; Friesema, E.C.; Verkaik, R.; Giacco, A.; Senese, R.; Arp, P.P.; Jhamai, P.M.; Pagnotta, S.M.; Broer, L.; et al. Short-Term, Combined Fasting and Exercise Improves Body Composition in Healthy Males. Int. J. Sport Nutr. Exerc. Metab. 2020, 30, 386–395. [Google Scholar] [CrossRef]
- Gaidhu, M.P.; Fediuc, S.; Anthony, N.M.; So, M.; Mirpourian, M.; Perry, R.L.; Ceddia, R.B. Prolonged AICAR-induced AMP-kinase activation promotes energy dissipation in white adipocytes: Novel mechanisms integrating HSL and ATGL. J. Lipid Res. 2009, 50, 704–715. [Google Scholar] [CrossRef]
- Bezaire, V.; Mairal, A.; Ribet, C.; Lefort, C.; Girousse, A.; Jocken, J.; Laurencikiene, J.; Anesia, R.; Rodriguez, A.-M.; Ryden, M.; et al. Contribution of Adipose Triglyceride Lipase and Hormone-sensitive Lipase to Lipolysis in hMADS Adipocytes. J. Biol. Chem. 2009, 284, 18282–18291. [Google Scholar] [CrossRef]
- Fougerat, A.; Schoiswohl, G.; Polizzi, A.; Régnier, M.; Wagner, C.; Smati, S.; Fougeray, T.; Lippi, Y.; Lasserre, F.; Raho, I.; et al. ATGL-dependent white adipose tissue lipolysis controls hepatocyte PPARα activity. Cell Rep. 2022, 39, 110910. [Google Scholar] [CrossRef] [PubMed]
- Kalderon, B.; Mayorek, N.; Berry, E.; Zevit, N.; Bar-Tana, J. Fatty acid cycling in the fasting rat. Am. J. Physiol. Metab. 2000, 279, E221–E227. [Google Scholar] [CrossRef]
- Coppack, S.W.; Persson, M.; Judd, R.L.; Miles, J.M. Glycerol and nonesterified fatty acid metabolism in human muscle and adipose tissue in vivo. Am. J. Physiol. Metab. 1999, 276, E233–E240. [Google Scholar] [CrossRef] [PubMed]
- Harney, D.J.; Cielesh, M.; Chu, R.; Cooke, K.C.; James, D.E.; Stöckli, J.; Larance, M. Proteomics analysis of adipose depots after intermittent fasting reveals visceral fat preservation mechanisms. Cell Rep. 2021, 34, 108804. [Google Scholar] [CrossRef] [PubMed]
- Bertholdt, L.; Gudiksen, A.; Stankiewicz, T.; Villesen, I.; Tybirk, J.; van Hall, G.; Bangsbo, J.; Plomgaard, P.; Pilegaard, H. Impact of training state on fasting-induced regulation of adipose tissue metabolism in humans. J. Appl. Physiol. 2018, 124, 729–740. [Google Scholar] [CrossRef]
- Sponarova, J.; Mustard, K.J.; Horakova, O.; Flachs, P.; Rossmeisl, M.; Brauner, P.; Bardova, K.; Thomason-Hughes, M.; Braunerova, R.; Janovska, P.; et al. Involvement of AMP-activated protein kinase in fat depot-specific metabolic changes during starvation. FEBS Lett. 2005, 579, 6105–6110. [Google Scholar] [CrossRef]
- Muzumdar, R.; Allison, D.B.; Huffman, D.M.; Ma, X.; Atzmon, G.; Einstein, F.H.; Fishman, S.; Poduval, A.D.; McVei, T.; Keith, S.W.; et al. Visceral adipose tissue modulates mammalian longevity. Aging Cell 2008, 7, 438–440. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zotti, T.; Giacco, A.; Cuomo, A.; Cerulo, L.; Petito, G.; Iervolino, S.; Senese, R.; Cioffi, F.; Vito, P.; Cardinale, G.; et al. Exercise Equals the Mobilization of Visceral versus Subcutaneous Adipose Fatty Acid Molecules in Fasted Rats Associated with the Modulation of the AMPK/ATGL/HSL Axis. Nutrients 2023, 15, 3095. https://doi.org/10.3390/nu15143095
Zotti T, Giacco A, Cuomo A, Cerulo L, Petito G, Iervolino S, Senese R, Cioffi F, Vito P, Cardinale G, et al. Exercise Equals the Mobilization of Visceral versus Subcutaneous Adipose Fatty Acid Molecules in Fasted Rats Associated with the Modulation of the AMPK/ATGL/HSL Axis. Nutrients. 2023; 15(14):3095. https://doi.org/10.3390/nu15143095
Chicago/Turabian StyleZotti, Tiziana, Antonia Giacco, Arianna Cuomo, Luigi Cerulo, Giuseppe Petito, Stefania Iervolino, Rosalba Senese, Federica Cioffi, Pasquale Vito, Gaetano Cardinale, and et al. 2023. "Exercise Equals the Mobilization of Visceral versus Subcutaneous Adipose Fatty Acid Molecules in Fasted Rats Associated with the Modulation of the AMPK/ATGL/HSL Axis" Nutrients 15, no. 14: 3095. https://doi.org/10.3390/nu15143095
APA StyleZotti, T., Giacco, A., Cuomo, A., Cerulo, L., Petito, G., Iervolino, S., Senese, R., Cioffi, F., Vito, P., Cardinale, G., Silvestri, E., Lombardi, A., Moreno, M., Lanni, A., & de Lange, P. (2023). Exercise Equals the Mobilization of Visceral versus Subcutaneous Adipose Fatty Acid Molecules in Fasted Rats Associated with the Modulation of the AMPK/ATGL/HSL Axis. Nutrients, 15(14), 3095. https://doi.org/10.3390/nu15143095