
Raspberry Leaves and Extracts-Molecular Mechanism of Action and Its Effectiveness on Human Cervical Ripening and the Induction of Labor

 , ,
, ,  , and
, and 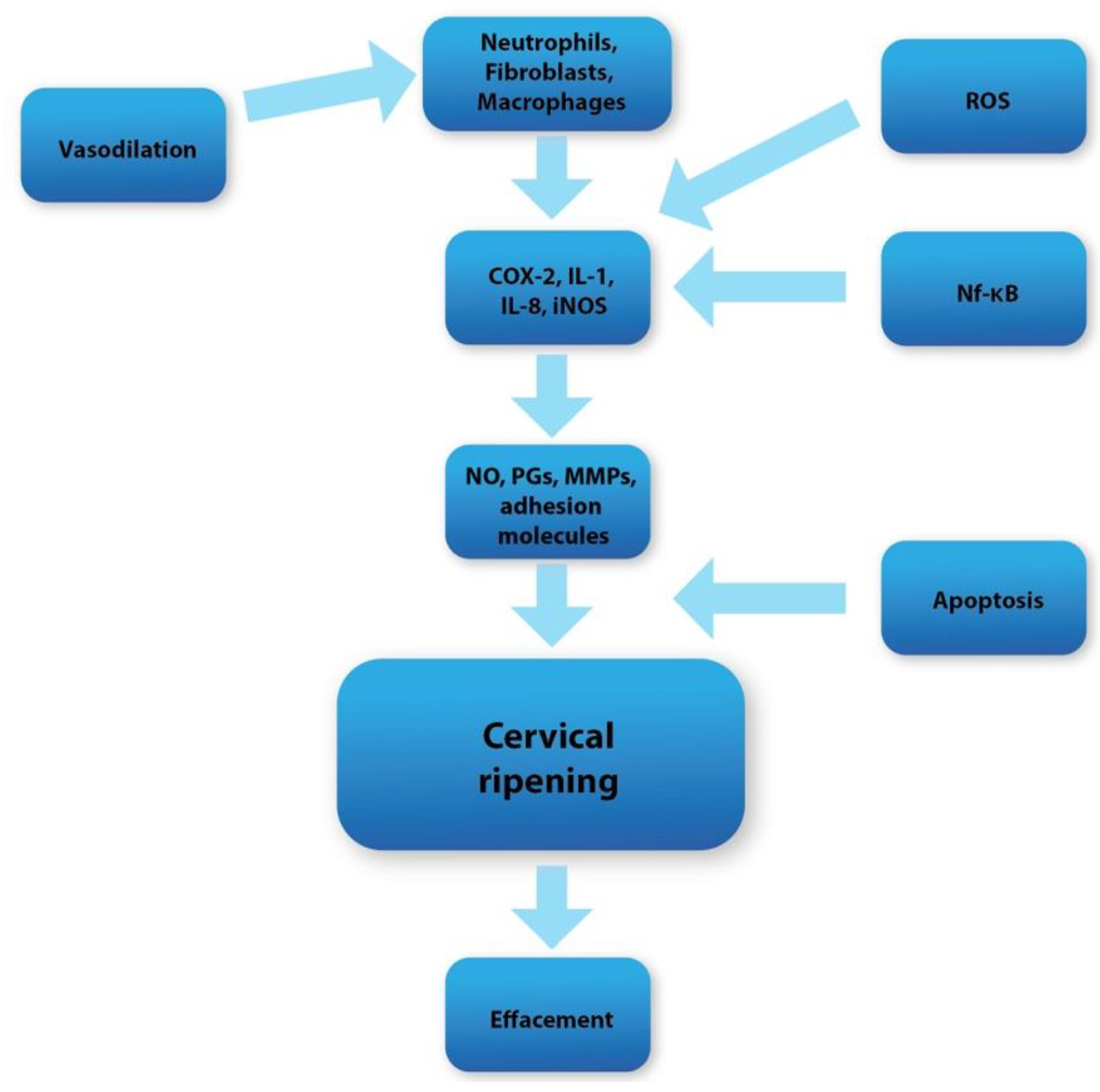{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Red Raspberry
3. Possible Impact on Labor and Cervical Ripening
4. Raspberries vs. Apoptosis and Reactive Oxygen Species (ROS)
5. Raspberries vs. Inflammation in Cervical Tissue
6. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Brown, B.; Wright, C. Safety and efficacy of supplements in pregnancy. Nutr. Rev. 2020, 78, 813–826. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, A.Z.; Moore, S.E.; Okala, S.G. Impact of Maternal Nutritional Supplementation during Pregnancy and Lactation on the Infant Gut or Breastmilk Microbiota: A Systematic Review. Nutrients 2021, 13, 1137. [Google Scholar] [CrossRef] [PubMed]
- Blondin, J.H.; LoGiudice, J.A. Pregnant women’s knowledge and awareness of nutrition. Appl. Nurs. Res. 2018, 39, 167–174. [Google Scholar] [CrossRef]
- Fallah, F.; Pourabbas, A.; Delpisheh, A.; Veisani, Y.; Shadnoush, M. Effects of Nutrition Education on Levels of Nutritional Awareness of Pregnant Women in Western Iran. Int. J. Endocrinol. Metab. 2013, 11, 175–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- A Forster, D.; Denning, A.; Wills, G.; Bolger, M.; McCarthy, E. Herbal medicine use during pregnancy in a group of Australian women. BMC Pregnancy Childbirth 2006, 6, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordeng, H.; Havnen, G.C. Use of herbal drugs in pregnancy: A survey among 400 Norwegian women. Pharmacoepidemiol. Drug Saf. 2004, 13, 371–380. [Google Scholar] [CrossRef]
- Hepner, D.L.; Harnett, M.; Segal, S.; Camann, W.; Bader, A.M.; Tsen, L.C. Herbal Medicine Use in Parturients. Obstet. Anesthesia Dig. 2002, 94, 690–693. [Google Scholar] [CrossRef]
- Westfall, R.E. Herbal healing in pregnancy: Women’s experiences. J. Herb. Pharmacother. 2003, 3, 17–39. [Google Scholar] [CrossRef]
- Mollart, L.; Adams, J.; Foureur, M. Pregnant women and health professional’s perceptions of complementary alternative medicine, and participation in a randomised controlled trial of acupressure for labour onset. Complement. Ther. Clin. Pract. 2016, 24, 167–173. [Google Scholar] [CrossRef]
- John, L.J.; Shantakumari, N. Herbal Medicines Use During Pregnancy: A Review from the Middle East. Oman Med. J. 2015, 30, 229–236. [Google Scholar] [CrossRef]
- Fan, K.W. National Center for Complementary and Alternative Medicine Website. J. Med. Libr. Assoc. 2005, 93, 410–412. [Google Scholar]
- Lans, C.; Turner, N.; Brauer, G.; Khan, T. Medicinal plants used in British Columbia, Canada for reproductive health in pets. Prev. Veter.-Med. 2009, 90, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Frawley, J.; Adams, J.; Sibbritt, D.; Steel, A.; Broom, A.; Gallois, C. Prevalence and determinants of complementary and alternative medicine use during pregnancy: Results from a nationally representative sample of Australian pregnant women. Aust. New Zealand J. Obstet. Gynaecol. 2013, 53, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Holst, L.; Haavik, S.; Nordeng, H. Raspberry leaf—Should it be recommended to pregnant women? Complement. Ther. Clin. Pr. 2009, 15, 204–208. [Google Scholar] [CrossRef]
- Bobinaitė, R.; Viskelis, P.; Bobinas, C.; Mieželienė, A.; Alenčikienė, G.; Venskutonis, P.R. Raspberry marc extracts increase antioxidative potential, ellagic acid, ellagitannin and anthocyanin concentrations in fruit purees. LWT Food Sci. Technol. 2016, 66, 460–467. [Google Scholar] [CrossRef]
- Costea, T.; Vlase, L.; Gostin, I.N.; Olah, N.K.; Predan, G.M.I. Botanical characterization, phytochemical analysis and antioxi-dant activity of indigenous red Raspberry (Rubus idaeus L.) leaves. Stud. Univ. Vasile Goldis Arad Ser. Stiint. Vietii. 2016, 26, 463–472. [Google Scholar]
- Rao, A.V.; Snyder, D.M. Raspberries and Human Health: A Review. J. Agric. Food Chem. 2010, 58, 3871–3883. [Google Scholar] [CrossRef]
- Veljkovic, B.; Djordjevic, N.; Dolicanin, Z.; Licina, B.; Topuzovic, M.; Stankovic, M.; Zlatic, N.; Dajic-Stevanovic, Z. Antioxidant and Anticancer Properties of Leaf and Fruit Extracts of the Wild Raspberry (Rubus idaeus L.). Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Belščak-Cvitanović, A.; Komes, D.; Benković, M.; Karlović, S.; Hečimović, I.; Ježek, D.; Bauman, I. Innovative formulations of chocolates enriched with plant polyphenols from Rubus idaeus L. leaves and characterization of their physical, bioactive and sensory properties. Food Res. Int. 2012, 48, 820–830. [Google Scholar] [CrossRef]
- Ferlemi, A.-V.; Lamari, F.N. Berry Leaves: An Alternative Source of Bioactive Natural Products of Nutritional and Medicinal Value. Antioxidants 2016, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Burton-Freeman, B.M.; Sandhu, A.K.; Edirisinghe, I. Red Raspberries and Their Bioactive Polyphenols: Cardiometabolic and Neuronal Health Links. Adv. Nutr. Int. Rev. J. 2016, 7, 44–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ispiryan, A.; Viškelis, J.; Viškelis, P. Red Raspberry (Rubus idaeus L.) Seed Oil: A Review. Plants 2021, 10, 944. [Google Scholar] [CrossRef] [PubMed]
- Garjonyte, R.; Budiene, J.; Labanauskas, L.; Judzentiene, A. In Vitro Antioxidant and Prooxidant Activities of Red Raspberry (Rubus idaeus L.) Stem Extracts. Molecules 2022, 27, 4073. [Google Scholar] [CrossRef] [PubMed]
- Sławińska, N.; Prochoń, K.; Olas, B. A Review on Berry Seeds—A Special Emphasis on Their Chemical Content and Health-Promoting Properties. Nutrients 2023, 15, 1422. [Google Scholar] [CrossRef]
- Gudej, J.; Tomczyk, M. Determination of Flavonoids, Tannins and Ellagic acid in leaves from Rubus L. species. Arch. Pharmacal Res. 2004, 27, 1114–1119. [Google Scholar] [CrossRef]
- Durgo, K.; Belščak-Cvitanović, A.; Stančić, A.; Franekić, J.; Komes, D. The Bioactive Potential of Red Raspberry (Rubus idaeus L.) Leaves in Exhibiting Cytotoxic and Cytoprotective Activity on Human Laryngeal Carcinoma and Colon Adenocarcinoma. J. Med. Food 2012, 15, 258–268. [Google Scholar] [CrossRef]
- Krzepiłko, A.; Prażak, R.; Święciło, A. Chemical Composition, Antioxidant and Antimicrobial Activity of Raspberry, Blackberry and Raspberry-Blackberry Hybrid Leaf Buds. Molecules 2021, 26, 327. [Google Scholar] [CrossRef]
- De Santis, D.; Carbone, K.; Garzoli, S.; Masci, V.L.; Turchetti, G. Bioactivity and Chemical Profile of Rubus idaeus L. Leaves Steam-Distillation Extract. Foods 2022, 11, 1455. [Google Scholar] [CrossRef]
- González, G.; Aguilera, F.; D’Afonseca, V. Transcriptome profiling of raspberry (Rubus idaeus Var. Amira) in response to infection by tomato ringspot virus (ToRSV). Heliyon 2020, 6, e04518. [Google Scholar] [CrossRef]
- A Manganaris, G.; Goulas, V.; Vicente, A.R.; A Terry, L. Berry antioxidants: Small fruits providing large benefits. J. Sci. Food Agric. 2014, 94, 825–833. [Google Scholar] [CrossRef] [Green Version]
- Oomah, B.; Ladet, S.; Godfrey, D.V.; Liang, J.; Girard, B. Characteristics of raspberry (Rubus idaeus L.) seed oil. Food Chem. 2000, 69, 187–193. [Google Scholar] [CrossRef]
- Nicoll, A. The Physiology of Cervical Ripening and the Induction of Labour: A Potential Role for the Nitric Oxide Donor Iso-Sorbide Mononitrate; University of Glasgow: Glasgow, UK, 2001. [Google Scholar]
- Schlembach, D.; MacKay, L.; Shi, L.; Maner, W.L.; Garfield, R.E.; Maul, H. Cervical ripening and insufficiency: From biochemical and molecular studies to in vivo clinical examination. Eur. J. Obstet. Gynecol. Reprod. Biol. 2009, 144, S70–S76. [Google Scholar] [CrossRef] [PubMed]
- Uldbjerg, N.; Ekman, G.; Malmström, A.; Olsson, K.; Ulmsten, U. Ripening of the human uterine cervix related to changes in collagen, glycosaminoglycans, and collagenolytic activity. Am. J. Obstet. Gynecol. 1983, 147, 662–666. [Google Scholar] [CrossRef]
- Iwahashi, M.; Muragaki, Y.; Ooshima, A.; Umesaki, N. Decreased Type I Collagen Expression in Human Uterine Cervix during Pregnancy. J. Clin. Endocrinol. Metab. 2003, 88, 2231–2235. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Jayyosi, C.; Lee, N.; Mahendroo, M.; Myers, K.M. Mechanics of cervical remodelling: Insights from rodent models of pregnancy. Interface Focus 2019, 9, 20190026. [Google Scholar] [CrossRef] [PubMed]
- Bollopragada, S.; Youssef, R.; Jordan, F.; Greer, I.; Norman, J.; Nelson, S. Term labor is associated with a core inflammatory response in human fetal membranes, myometrium, and cervix. Am. J. Obstet. Gynecol. 2009, 200, 104.e1–104.e11. [Google Scholar] [CrossRef]
- Tantengco, O.A.G.; Vink, J.; Medina, P.M.B.; Menon, R. Oxidative stress promotes cellular damages in the cervix: Implications for normal and pathologic cervical function in human pregnancy. Biol. Reprod. 2021, 105, 204–216. [Google Scholar] [CrossRef]
- Leppert, P.C.; Yu, S.Y. Apoptosis in the Cervix of Pregnant Rats in Association with Cervical Softening. Gynecol. Obstet. Investig. 1994, 37, 150–154. [Google Scholar] [CrossRef]
- Fulda, S.; Gorman, A.M.; Hori, O.; Samali, A. Cellular Stress Responses: Cell Survival and Cell Death. Int. J. Cell Biol. 2010, 2010, 214074. [Google Scholar] [CrossRef] [Green Version]
- Ricci, J.-E.; Muñoz-Pinedo, C.; Fitzgerald, P.; Bailly-Maitre, B.; A Perkins, G.; Yadava, N.; E Scheffler, I.; Ellisman, M.H.; Green, D.R. Disruption of Mitochondrial Function during Apoptosis Is Mediated by Caspase Cleavage of the p75 Subunit of Complex I of the Electron Transport Chain. Cell 2004, 117, 773–786. [Google Scholar] [CrossRef] [Green Version]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase Functions in Cell Death and Disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayagaki, N.; Warming, S.; Lamkanfi, M.; Walle, L.V.; Louie, S.; Dong, J.; Newton, K.; Qu, Y.; Liu, J.; Heldens, S.; et al. Non-canonical inflammasome activation targets caspase-11. Nature 2011, 479, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef] [PubMed]
- Tait, S.W.G.; Green, D.R. Mitochondria and cell death: Outer membrane permeabilization and beyond. Nat. Rev. Mol. Cell Biol. 2010, 11, 621–632. [Google Scholar] [CrossRef]
- Wang, H.-G.; Pathan, N.; Ethell, I.M.; Krajewski, S.; Yamaguchi, Y.; Shibasaki, F.; McKeon, F.; Bobo, T.; Franke, T.F.; Reed, J.C. Ca 2+ -Induced Apoptosis Through Calcineurin Dephosphorylation of BAD. Science 1999, 284, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Leibowitz, B.; Yu, J. Mitochondrial signaling in cell death via the Bcl-2 family. Cancer Biol. Ther. 2010, 9, 417–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Gubory, K.H.; Fowler, P.A.; Garrel, C. The roles of cellular reactive oxygen species, oxidative stress and antioxidants in pregnancy outcomes. Int. J. Biochem. Cell Biol. 2010, 42, 1634–1650. [Google Scholar] [CrossRef]
- Agarwal, A.; Aponte-Mellado, A.; Premkumar, B.J.; Shaman, A.; Gupta, S. The effects of oxidative stress on female reproduction: A review. Reprod. Biol. Endocrinol. 2012, 10, 49. [Google Scholar] [CrossRef] [Green Version]
- Fujii, J.; Iuchi, Y.; Okada, F. Fundamental roles of reactive oxygen species and protective mechanisms in the female reproductive system. Reprod. Biol. Endocrinol. 2005, 3, 43. [Google Scholar] [CrossRef] [Green Version]
- Landes, T.; Martinou, J.-C. Mitochondrial outer membrane permeabilization during apoptosis: The role of mitochondrial fission. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2011, 1813, 540–545. [Google Scholar] [CrossRef]
- Kaufmann, T.; Strasser, A.; Jost, P.J. Fas death receptor signalling: Roles of Bid and XIAP. Cell Death Differ. 2012, 19, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Ruder, E.H.; Hartman, T.J.; Goldman, M.B. Impact of oxidative stress on female fertility. Curr. Opin. Obstet. Gynecol. 2009, 21, 219–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-O.; Lee, C.Y. Comprehensive Study on Vitamin C Equivalent Antioxidant Capacity (VCEAC) of Various Polyphenolics in Scavenging a Free Radical and its Structural Relationship. Crit. Rev. Food Sci. Nutr. 2004, 44, 253–273. [Google Scholar] [CrossRef] [PubMed]
- Pietta, P.-G. Flavonoids as Antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, X.Q.; Weber, C.; Lee, C.Y.; Brown, J.; Liu, R.H. Antioxidant and Antiproliferative Activities of Raspberries. J. Agric. Food Chem. 2002, 50, 2926–2930. [Google Scholar] [CrossRef] [PubMed]
- Marreiro, D.D.N.; Cruz, K.J.C.; Morais, J.B.S.; Beserra, J.B.; Severo, J.S.; De Oliveira, A.R.S. Zinc and oxidative stress: Current mechanisms. Antioxidants 2017, 6, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frías-Moreno, M.N.; Parra-Quezada, R.A.; González-Aguilar, G.; Ruíz-Canizales, J.; Molina-Corral, F.J.; Sepulveda, D.R.; Salas-Salazar, N.; Olivas, G.I. Quality, Bioactive Compounds, Antioxidant Capacity, and Enzymes of Raspberries at Different Maturity Stages, Effects of Organic vs. Conventional Fertilization. Foods 2021, 10, 953. [Google Scholar] [CrossRef]
- Teng, H.; Lin, Q.; Li, K.; Yuan, B.; Song, H.; Peng, H.; Yi, L.; Wei, M.-C.; Yang, Y.-C.; Battino, M.; et al. Hepatoprotective Effects of Raspberry (Rubus coreanus Miq.) Seed Oil and Its Major Constituents. Food Chem. Toxicol. 2017, 110, 418–424. [Google Scholar] [CrossRef]
- Choi, M.-H.; Shim, S.-M.; Kim, G.-H. Protective effect of black raspberry seed containing anthocyanins against oxidative damage to DNA, protein, and lipid. J. Food Sci. Technol. 2016, 53, 1214–1221. [Google Scholar] [CrossRef] [Green Version]
- Kang, I.; Espín, J.C.; Carr, T.P.; Tomás-Barberán, F.A.; Chung, S. Raspberry Seed Flour Attenuates High-Sucrose Diet-Mediated Hepatic Stress and Adipose Tissue Inflammation. J. Nutr. Biochem. 2016, 32, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.-H.; Wang, P.-W.; Lin, T.-Y.; Yang, P.-M.; Li, W.-T.; Yeh, C.-T.; Pan, T.-L. Antioxidant properties of red raspberry extract alleviate hepatic fibrosis via inducing apoptosis and transdifferentiation of activated hepatic stellate cells. Biomed. Pharmacother. 2021, 144, 112284. [Google Scholar] [CrossRef]
- Yousuf, M.; Shamsi, A.; Khan, P.; Shahbaaz, M.; AlAjmi, M.F.; Hussain, A.; Hassan, G.M.; Islam, A.; Haque, Q.M.R.; Hassan, I. Ellagic Acid Controls Cell Proliferation and Induces Apoptosis in Breast Cancer Cells via Inhibition of Cyclin-Dependent Kinase 6. Int. J. Mol. Sci. 2020, 21, 3526. [Google Scholar] [CrossRef] [PubMed]
- Ríos, J.-L.; Giner, R.M.; Marín, M.; Recio, M.C. A Pharmacological Update of Ellagic Acid. Planta Med. 2018, 84, 1068–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Nicolay, B.N.; Chick, J.M.; Gao, X.; Geng, Y.; Ren, H.; Gao, H.; Yang, G.; Williams, J.A.; Suski, J.M.; et al. The metabolic function of cyclin D3–CDK6 kinase in cancer cell survival. Nature 2017, 546, 426–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.-W.; Hu, J.-J.; Fu, R.-Q.; Liu, X.; Zhang, Y.-H.; Li, J.; Liu, L.; Li, Y.-N.; Deng, Q.; Luo, Q.-S.; et al. Flavonoids inhibit cell proliferation and induce apoptosis and autophagy through downregulation of PI3Kγ mediated PI3K/AKT/mTOR/p70S6K/ULK signaling pathway in human breast cancer cells. Sci. Rep. 2018, 8, 11255. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Knobloch, T.J.; Seamon, L.G.; Stoner, G.D.; Cohn, D.E.; Paskett, E.D.; Fowler, J.M.; Weghorst, C.M. A black raspberry extract inhibits proliferation and regulates apoptosis in cervical cancer cells. Gynecol. Oncol. 2011, 123, 401–406. [Google Scholar] [CrossRef] [Green Version]
- Gu, I.; Brownmiller, C.; Howard, L.; Lee, S.-O. Chemical Composition of Volatile Extracts from Black Raspberries, Blueberries, and Blackberries and Their Antiproliferative Effect on A549 Non-Small-Cell Lung Cancer Cells. Life 2022, 12, 2056. [Google Scholar] [CrossRef]
- Ruscheinsky, M.; De la Motte, C.; Mahendroo, M. Hyaluronan and its binding proteins during cervical ripening and parturition: Dynamic changes in size, distribution and temporal sequence. Matrix Biol. J. Int. Soc. Matrix Biol. 2008, 27, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Lopez, N.; Guilbert, L.J.; Olson, D.M. Invasion of the leukocytes into the fetal-maternal interface during pregnancy. J. Leukoc. Biol. 2010, 88, 625–633. [Google Scholar] [CrossRef]
- Osmers, R.G.W.; Blaser, J.; Kuhn, W.; Tschesche, H. Interleukin-8 synthesis and the onset of labor. Obstet. Gynecol. 1995, 86, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.; Fischer, D.-C.C.; Ruck, P.; Marx, T.; Kaiserling, E.; Oberpichler, A.; Tschesche, H.; Rath, W.; Kaiserlîng, E.; Oberpichler, A.; et al. Parturition at term: Parallel increases in interleukin-8 and proteinase concentrations and neutrophil count in the lower uterine segment. Hum. Reprod. 1999, 14, 1096–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Maradny, E.; Kanayama, N.; Halim, A.; Maehara, K.; Sumimoto, K.; Terao, T. The effect of interleukin-1 in rabbit cervical ripening. Eur. J. Obstet. Gynecol. Reprod. Biol. 1995, 60, 75–80. [Google Scholar] [CrossRef]
- Watari, M.; Watari, H.; DiSanto, M.E.; Chacko, S.; Shi, G.P.; Strauss, J.F., III. Proinflammatory cytokines induce expression of matrix-metabolizing enzymes in human cervical smooth muscle cells. Am. J. Pathol. 1999, 154, 1755–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinaldi, S.F.; Hutchinson, J.L.; Rossi, A.G.; Norman, J.E. Anti-inflammatory mediators as physiological and pharmacological regulators of parturition. Expert Rev. Clin. Immunol. 2011, 7, 675–696. [Google Scholar] [CrossRef]
- Terzidou, V. Biochemical and endocrinological preparation for parturition. Best Pr. Res. Clin. Obstet. Gynaecol. 2007, 21, 729–756. [Google Scholar] [CrossRef]
- Sugimoto, Y.; Narumiya, S. Prostaglandin E Receptors. J. Biol. Chem. 2007, 282, 11613–11617. [Google Scholar] [CrossRef] [Green Version]
- Bakker, R.; Pierce, S.; Myers, D. The role of prostaglandins E1 and E2, dinoprostone, and misoprostol in cervical ripening and the induction of labor: A mechanistic approach. Arch. Gynecol. Obstet. 2017, 296, 167–179. [Google Scholar] [CrossRef]
- Maradny, E.E.; Kanayama, N.; Halim, A.; Maehara, K.; Sumimoto, K.; Terao, T. Biochemical changes in the cervical tissue of rabbit induced by interleukin-8, interleukin-1 dehydroepiandrosterone sulphate and prostaglandin E2: A comparative study. Hum. Reprod. 1996, 11, 1099–1104. [Google Scholar] [CrossRef] [Green Version]
- Winkler, M.; Kemp, B.; Hauptmann, S.; Rath, W. Parturition: Steroids, prostaglandin E2, and expression of adhesion molecules by endothelial cells. Obstet. Gynecol. 1997, 89, 398–402. [Google Scholar] [CrossRef]
- Astle, S.; Thornton, S.; Slater, D.M. Identification and localization of prostaglandin E2 receptors in upper and lower segment human myometrium during pregnancy. Mol. Hum. Reprod. 2005, 11, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Morris, R.J.; Bode, A.M.; Zhang, T. Prostaglandin Pathways: Opportunities for Cancer Prevention and Therapy. Cancer Res 2022, 82, 949–965. [Google Scholar] [CrossRef] [PubMed]
- Chwalisz, K.; Shao-Qing, S.; Garfield, R.E.; Beier, H.M. Cervical ripening in guinea-pigs after a local application of nitric oxide. Hum. Reprod. 1997, 12, 2093–2101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Król, M.; Kepinska, M. Human Nitric Oxide Synthase—Its Functions, Polymorphisms, and Inhibitors in the Context of Inflammation, Diabetes and Cardiovascular Diseases. Int. J. Mol. Sci. 2020, 22, 56. [Google Scholar] [CrossRef]
- Cinelli, M.A.; Do, H.T.; Miley, G.P.; Silverman, R.B. Inducible nitric oxide synthase: Regulation, structure, and inhibition. Med. Res. Rev. 2020, 40, 158–189. [Google Scholar] [CrossRef]
- Tschugguel, W.; Schneeberger, C.; Lass, H.; Stonek, F.; Zaghlula, M.B.; Czerwenka, K.; Schatten, C.; Kaider, A.; Husslein, P.; Huber, J.C. Human cervical ripening is associated with an increase in cervical inducible nitric oxide synthase expression. Biol. Reprod. 1999, 60, 1367–1372. [Google Scholar] [CrossRef] [Green Version]
- Väisänen-Tommiska, M.R.H. Nitric oxide in the human uterine cervix: Endogenous ripening factor. Ann. Med. 2008, 40, 45–55. [Google Scholar] [CrossRef]
- Ekerhovd, E.; Weijdegård, B.; Brännström, M.; Mattsby-Baltzer, I.; Norström, A. Nitric oxide induced cervical ripening in the human: Involvement of cyclic guanosine monophosphate, prostaglandin F2α, and prostaglandin E2. Am. J. Obstet. Gynecol. 2002, 186, 745–750. [Google Scholar] [CrossRef]
- Learmont, J.G.; Poston, L. Nitric oxide is involved in flow-induced dilation of isolated human small fetoplacental arteries. Am. J. Obstet. Gynecol. 1996, 174, 583–588. [Google Scholar] [CrossRef]
- Choi, B.-M.; Pae, H.-O.; Jang, S.I.; Kim, Y.-M.; Chung, H.-T. Nitric Oxide as a Pro-apoptotic as well as Anti-apoptotic Modulator. BMB Rep. 2002, 35, 116–126. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A.; Lattey, K.R.; Kelly, A.J. Nitric oxide donors for cervical ripening and induction of labour. Cochrane Database Syst. Rev. 2016, 2016, CD006901. [Google Scholar] [CrossRef] [PubMed]
- Ilchovska, D.; Barrow, M. An Overview of the NF-kB mechanism of pathophysiology in rheumatoid arthritis, investigation of the NF-kB ligand RANKL and related nutritional interventions. Autoimmun. Rev. 2021, 20, 102741. [Google Scholar] [CrossRef]
- Tilborghs, S.; Corthouts, J.; Verhoeven, Y.; Arias, D.; Rolfo, C.; Trinh, X.B.; van Dam, P.A. The role of Nuclear Factor-kappa B signaling in human cervical cancer. Crit. Rev. Oncol. 2017, 120, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Chávez, F.; Correa, D.; Navarrete-Meneses, P.; Cancino-Diaz, J.C.; Cancino-Diaz, M.E.; Rodríguez-Martínez, S. NF-κB and Its Regulators During Pregnancy. Front. Immunol. 2021, 12, 679106. [Google Scholar] [CrossRef] [PubMed]
- Kalkhoven, E.; Wissink, S.; van der Saag, P.T.; van der Burg, B. Negative Interaction between the RelA(p65) Subunit of NF-κB and the Progesterone Receptor. J. Biol. Chem. 1996, 271, 6217–6224. [Google Scholar] [CrossRef] [Green Version]
- Allport, V.; Pieber, D.; Slater, D.; Newton, R.; White, J.; Bennett, P. Human labour is associated with nuclear factor-kappaB activity which mediates cyclo-oxygenase-2 expression and is involved with the ‘functional progesterone withdrawal’. Mol. Hum. Reprod. 2001, 7, 581–586. [Google Scholar] [CrossRef] [Green Version]
- Clancy, R.; Gomez, P.; Abramson, S. Nitric oxide sustains nuclear factor kappaB activation in cytokine-stimulated chondrocytes. Osteoarthr. Cartil. 2004, 12, 552–558. [Google Scholar] [CrossRef] [Green Version]
- Gerlo, S.; Kooijman, R.; Beck, I.M.; Kolmus, K.; Spooren, A.; Haegeman, G. Cyclic AMP: A selective modulator of NF-κB action. Cell. Mol. Life Sci. 2011, 68, 3823–3841. [Google Scholar] [CrossRef]
- Orsi, N.M.; Tribe, R.M. Cytokine Networks and the Regulation of Uterine Function in Pregnancy and Parturition. J. Neuroendocr. 2008, 20, 462–469. [Google Scholar] [CrossRef]
- Cindrova-Davies, T.; Spasic-Boskovic, O.; Jauniaux, E.; Charnock-Jones, D.S.; Burton, G.J. Nuclear Factor-κB, p38, and Stress-Activated Protein Kinase Mitogen-Activated Protein Kinase Signaling Pathways Regulate Proinflammatory Cytokines and Apoptosis in Human Placental Explants in Response to Oxidative Stress: Effects of Antioxidant Vitamins. Am. J. Pathol. 2007, 170, 1511–1520. [Google Scholar] [CrossRef]
- Gloire, G.; Legrand-Poels, S.; Piette, J. NF-κB activation by reactive oxygen species: Fifteen years later. Biochem. Pharmacol. 2006, 72, 1493–1505. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Engelhardt, J.F. Interleukin-1β Induction of NFκB Is Partially Regulated by H2O2-mediated Activation of NFκB-inducing Kinase. J. Biol. Chem. 2006, 281, 1495–1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Kim, S.C.; Yu, T.; Yi, Y.-S.; Rhee, M.H.; Sung, G.-H.; Yoo, B.C.; Cho, J.Y. Functional Roles of p38 Mitogen-Activated Protein Kinase in Macrophage-Mediated Inflammatory Responses. Mediat. Inflamm. 2014, 2014, 352371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Boehm, J.; Lee, J.C. p38 MAP kinases: Key signalling molecules as therapeutic targets for inflammatory diseases. Nat. Rev. Drug Discov. 2003, 2, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.-T.; Wei, W.; Ma, B.; Xu, Y.; Liu, W.-J.; Wang, Y.; Lv, K.-Y.; Tang, H.-T.; Wei, D.; Xia, Z.-F. Activation of p38 MAPK by Reactive Oxygen Species Is Essential in a Rat Model of Stress-Induced Gastric Mucosal Injury. J. Immunol. 2007, 179, 7808–7819. [Google Scholar] [CrossRef] [Green Version]
- Svineng, G.; Ravuri, C.; Rikardsen, O.; Huseby, N.-E.; Winberg, J.-O. The Role of Reactive Oxygen Species in Integrin and Matrix Metalloproteinase Expression and Function. Connect. Tissue Res. 2008, 49, 197–202. [Google Scholar] [CrossRef]
- Jin, J.; Richardson, L.; Sheller-Miller, S.; Zhong, N.; Menon, R. Oxidative stress induces p38MAPK-dependent senescence in the feto-maternal interface cells. Placenta 2018, 67, 15–23. [Google Scholar] [CrossRef]
- Golmohamadi, A.; Möller, G.; Powers, J.; Nindo, C. Effect of ultrasound frequency on antioxidant activity, total phenolic and anthocyanin content of red raspberry puree. Ultrason. Sonochem. 2013, 20, 1316–1323. [Google Scholar] [CrossRef]
- Nistor, M.; Pop, R.; Daescu, A.; Pintea, A.; Socaciu, C.; Rugina, D. Anthocyanins as Key Phytochemicals Acting for the Prevention of Metabolic Diseases: An Overview. Molecules 2022, 27, 4254. [Google Scholar] [CrossRef]
- Itoh, K.; Wakabayashi, N.; Katoh, Y.; Ishii, T.; Igarashi, K.; Engel, J.D.; Yamamoto, M. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes Dev. 1999, 13, 76–86. [Google Scholar] [CrossRef] [Green Version]
- Dinkova-Kostova, A.T.; Holtzclaw, W.D.; Cole, R.N.; Itoh, K.; Wakabayashi, N.; Katoh, Y.; Yamamoto, M.; Talalay, P. Direct evidence that sulfhydryl groups of Keap1 are the sensors regulating induction of phase 2 enzymes that protect against carcinogens and oxidants. Proc. Natl. Acad. Sci. USA 2002, 99, 11908–11913. [Google Scholar] [CrossRef] [PubMed]
- Baird, L.; Dinkova-Kostova, A.T. The cytoprotective role of the Keap1–Nrf2 pathway. Arch. Toxicol. 2011, 85, 241–272. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Wang, X.J. Induction of the Keap1/Nrf2/ARE pathway by oxidizable diphenols. Chem. Interact. 2011, 192, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Kropat, C.; Mueller, D.; Boettler, U.; Zimmermann, K.; Heiss, E.H.; Dirsch, V.M.; Rogoll, D.; Melcher, R.; Richling, E.; Marko, D. Modulation of Nrf2-dependent gene transcription by bilberry anthocyanins in vivo. Mol. Nutr. Food Res. 2013, 57, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Groh, I.A.M.; Bakuradze, T.; Pahlke, G.; Richling, E.; Marko, D. Consumption of anthocyanin-rich beverages affects Nrf2 and Nrf2-dependent gene transcription in peripheral lymphocytes and DNA integrity of healthy volunteers. BMC Chem. 2020, 14, 39. [Google Scholar] [CrossRef]
- Kähkönen, M.; Kylli, P.; Ollilainen, V.; Salminen, J.-P.; Heinonen, M. Antioxidant Activity of Isolated Ellagitannins from Red Raspberries and Cloudberries. J. Agric. Food Chem. 2012, 60, 1167–1174. [Google Scholar] [CrossRef]
- Gođevac, D.; Tešević, V.; Vajs, V.; Milosavljević, S.; Stanković, M. Antioxidant properties of raspberry seed extracts on micronucleus distribution in peripheral blood lymphocytes. Food Chem. Toxicol. 2009, 47, 2853–2859. [Google Scholar] [CrossRef]
- Sangiovanni, E.; Vrhovsek, U.; Rossoni, G.; Colombo, E.; Brunelli, C.; Brembati, L.; Trivulzio, S.; Gasperotti, M.; Mattivi, F.; Bosisio, E.; et al. Ellagitannins from Rubus Berries for the Control of Gastric Inflammation: In Vitro and In Vivo Studies. PLoS ONE 2013, 8, e71762. [Google Scholar] [CrossRef] [Green Version]
- Hendawy, O.; Gomaa, H.A.M.; Hussein, S.; Alzarea, S.I.; Qasim, S.; Abdel Rahman, F.E.Z.S.; Ali, A.T.; Ahmed, S.R. Cold-Pressed Raspberry Seeds Oil Ameliorates High-Fat Diet Triggered Non-Alcoholic Fatty Liver Disease. Saudi Pharm. J. 2021, 29, 1303–1313. [Google Scholar] [CrossRef]
- Morgan, M.J.; Liu, Z.-G. Crosstalk of reactive oxygen species and NF-kappaκB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wang, L.; Wu, Z.; Yao, L.; Wu, Y.; Huang, L.; Liu, K.; Zhou, X.; Gou, D. Anthocyanin-rich fractions from red raspberries attenuate inflammation in both RAW264.7 macrophages and a mouse model of colitis. Sci. Rep. 2014, 4, 6234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeram, N.P.; Momin, R.A.; Nair, M.G.; Bourquin, L.D. Cyclooxygenase inhibitory and antioxidant cyanidin glycosides in cherries and berries. Phytomedicine 2001, 8, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Vuorela, S.; Kreander, K.; Karonen, M.; Nieminen, R.; Hämäläinen, M.; Galkin, A.; Laitinen, L.; Salminen, J.-P.; Moilanen, E.; Pihlaja, K.; et al. Preclinical Evaluation of Rapeseed, Raspberry, and Pine Bark Phenolics for Health Related Effects. J. Agric. Food Chem. 2005, 53, 5922–5931. [Google Scholar] [CrossRef]
- Favarin, D.C.; Teixeira, M.M.; de Andrade, E.L.; Alves, C.d.F.; Chica, J.E.L.; Sorgi, C.A.; Faccioli, L.H.; Rogerio, A.P. Anti-Inflammatory Effects of Ellagic Acid on Acute Lung Injury Induced by Acid in Mice. Mediat. Inflamm. 2013, 2013, 164202. [Google Scholar] [CrossRef] [Green Version]
- Karimi, M.Y.; Fatemi, I.; Kalantari, H.; Mombeini, M.A.; Mehrzadi, S.; Goudarzi, M. Ellagic Acid Prevents Oxidative Stress, Inflammation, and Histopathological Alterations in Acrylamide-Induced Hepatotoxicity in Wistar Rats. J. Diet. Suppl. 2020, 17, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Hwang, H.; Rose, M.E.; Nines, R.G.; Stoner, G.D. Chemopreventive Properties of Black Raspberries in N-Nitrosomethylbenzylamine-Induced Rat Esophageal Tumorigenesis: Down-regulation of Cyclooxygenase-2, Inducible Nitric Oxide Synthase, and c-Jun. Cancer Res 2006, 66, 2853–2859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majewski, M.; Kucharczyk, E.; Kaliszan, R.; Markuszewski, M.; Fotschki, B.; Juśkiewicz, J.; Borkowska-Sztachańska, M.; Ognik, K. The Characterization of Ground Raspberry Seeds and the Physiological Response to Supplementation in Hypertensive and Normotensive Rats. Nutrients 2020, 12, 1630. [Google Scholar] [CrossRef] [PubMed]
- El Hajj, M.; Holst, L. Herbal Medicine Use During Pregnancy: A Review of the Literature With a Special Focus on Sub-Saharan Africa. Front. Pharmacol. 2020, 11, 866. [Google Scholar] [CrossRef]
- Illamola, S.M.; Amaeze, O.; Krepkova, L.; Birnbaum, A.K.; Karanam, A.; Job, K.M.; Bortnikova, V.V.; Sherwin, C.; Enioutina, E.Y. Use of Herbal Medicine by Pregnant Women: What Physicians Need to Know. Front. Pharmacol. 2020, 10, 1483. [Google Scholar] [CrossRef]
- Zheng, J.; Pistilli, M.J.; Holloway, A.C.; Crankshaw, D.J. The Effects of Commercial Preparations of Red Raspberry Leaf on the Contractility of the Rat’s Uterus In Vitro. Reprod. Sci. 2010, 17, 494–501. [Google Scholar] [CrossRef]
- Rojas-Vera, J.; Patel, A.V.; Dacke, C.G. Relaxant activity of raspberry? (Rubus idaeus) leaf extract in guinea-pig ileum in vitro. Phytotherapy Res. 2002, 16, 665–668. [Google Scholar] [CrossRef] [PubMed]
- Bowman, R.; Taylor, J.; Muggleton, S.; Davis, D. Biophysical effects, safety and efficacy of raspberry leaf use in pregnancy: A systematic integrative review. BMC Complement Med. Ther. 2021, 21, 56. [Google Scholar] [CrossRef]
- Chwalisz, K.; Garfield, R.E. Role of nitric oxide in the uterus and cervix: Implications for the management of labor. J. Perinat. Med. 1998, 26, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Parsons, M.; Simpson, M.; Ponton, T. Raspberry leaf and its effect on labour: Safety and efficacy. Aust. Coll. Midwives Inc. J. 1999, 12, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Cheang, K.I.; Nguyen, T.T.; Karjane, N.W.; Salley, K.E.S. Raspberry Leaf and Hypoglycemia in Gestational Diabetes Mellitus. Obstet. Gynecol. 2016, 128, 1421–1424. [Google Scholar] [CrossRef]
- Langhammer, A.J.; Nilsen, O.G. In vitro inhibition of human CYP1A2, CYP2D6, and CYP3A4 by six herbs commonly used in pregnancy. Phytother. Res. 2014, 28, 603–610. [Google Scholar] [CrossRef]
- Na, L.; Jun-tian, L.; Qiang-zong, Z. Ellagic acid-induced hypercoagulable state in animals: A potentially useful animal hyper-coagulable model for evaluation of anticoagulants. Chin. Med. Sci. J. 2010, 25, 237–242. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Socha, M.W.; Flis, W.; Wartęga, M.; Szambelan, M.; Pietrus, M.; Kazdepka-Ziemińska, A. Raspberry Leaves and Extracts-Molecular Mechanism of Action and Its Effectiveness on Human Cervical Ripening and the Induction of Labor. Nutrients 2023, 15, 3206. https://doi.org/10.3390/nu15143206
Socha MW, Flis W, Wartęga M, Szambelan M, Pietrus M, Kazdepka-Ziemińska A. Raspberry Leaves and Extracts-Molecular Mechanism of Action and Its Effectiveness on Human Cervical Ripening and the Induction of Labor. Nutrients. 2023; 15(14):3206. https://doi.org/10.3390/nu15143206
Chicago/Turabian StyleSocha, Maciej W., Wojciech Flis, Mateusz Wartęga, Monika Szambelan, Miłosz Pietrus, and Anita Kazdepka-Ziemińska. 2023. "Raspberry Leaves and Extracts-Molecular Mechanism of Action and Its Effectiveness on Human Cervical Ripening and the Induction of Labor" Nutrients 15, no. 14: 3206. https://doi.org/10.3390/nu15143206
APA StyleSocha, M. W., Flis, W., Wartęga, M., Szambelan, M., Pietrus, M., & Kazdepka-Ziemińska, A. (2023). Raspberry Leaves and Extracts-Molecular Mechanism of Action and Its Effectiveness on Human Cervical Ripening and the Induction of Labor. Nutrients, 15(14), 3206. https://doi.org/10.3390/nu15143206