
Potential of Fatty Acids in Treating Sarcopenia: A Systematic Review



Abstract
:1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Selection Criteria
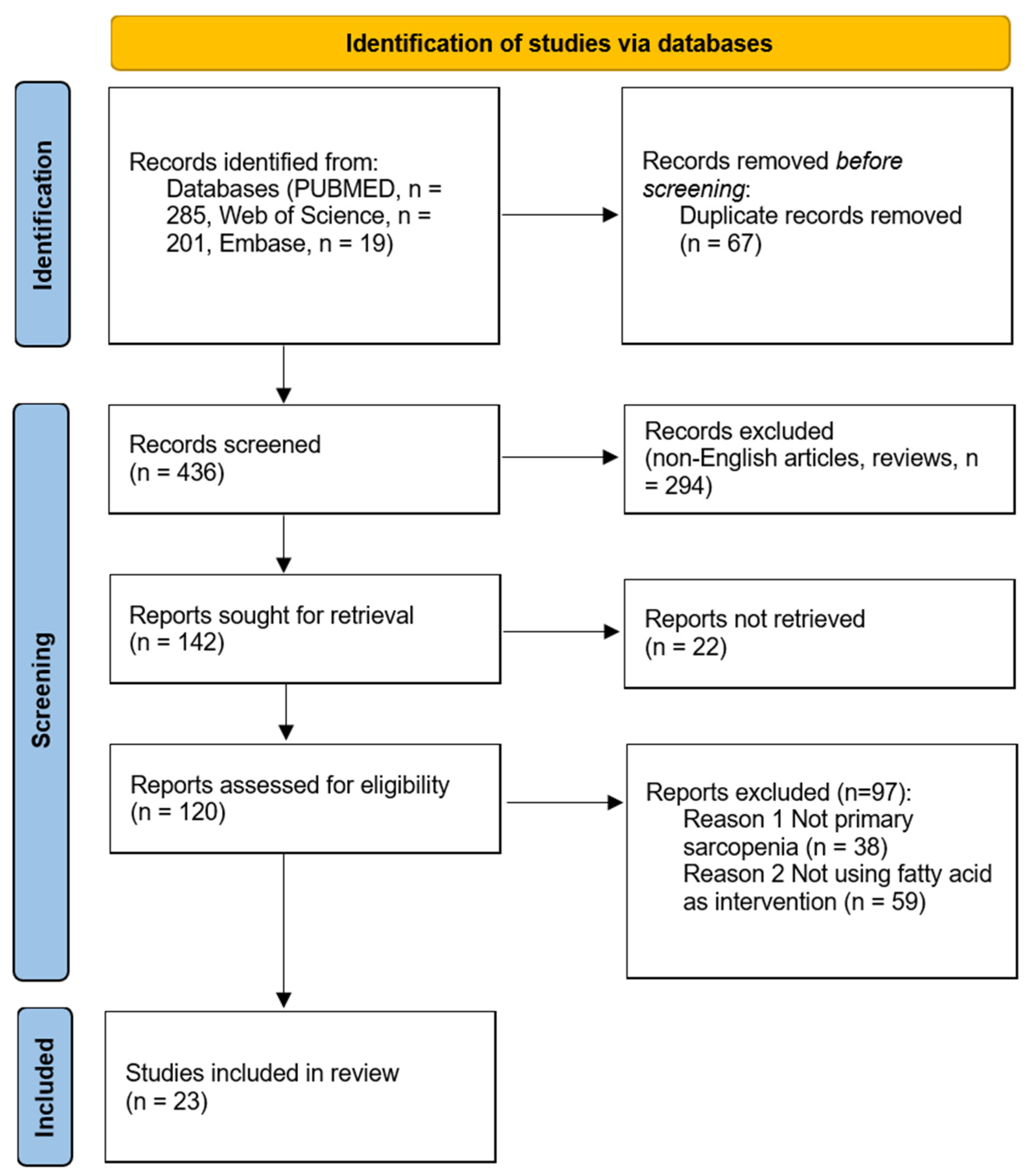
2.3. Study Selection
2.4. Risk of Bias Assessment
2.5. Data Extraction and Analysis
3. Results
3.1. Clinical Studies
3.1.1. Outcome Assessments
3.1.2. Intervention and Results
3.1.3. Key Findings
3.2. Pre-Clinical Studies
3.2.1. Cell Experiments
3.2.2. Animal Studies
3.2.3. Key Findings
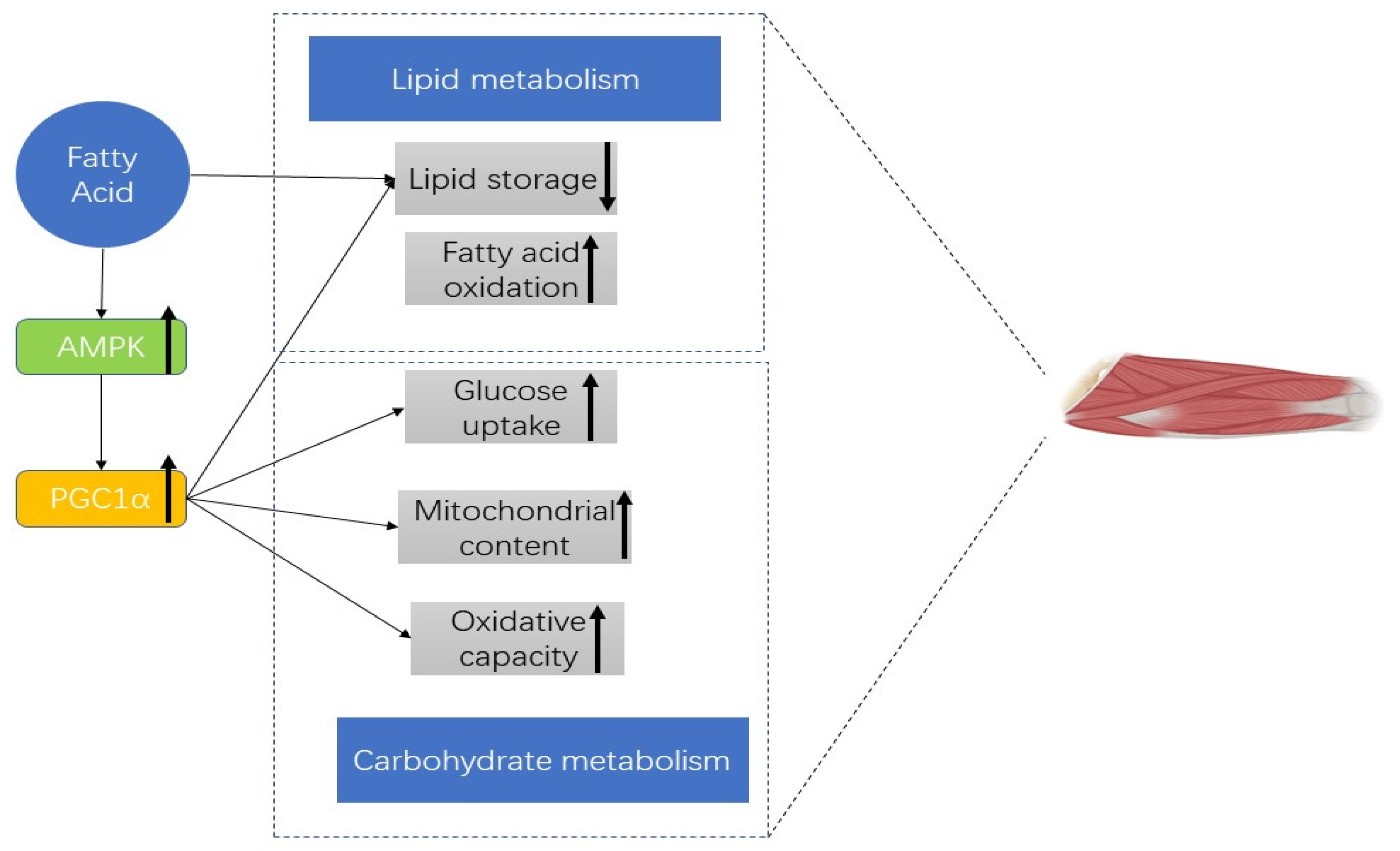
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cruz-Jentoft, A.J.; Baeyens, J.P.; Bauer, J.M.; Boirie, Y.; Cederholm, T.; Landi, F.; Martin, F.C.; Michel, J.P.; Rolland, Y.; Schneider, S.M.; et al. Sarcopenia: European consensus on definition and diagnosis: Report of the European Working Group on Sarcopenia in Older People. Age Ageing 2010, 39, 412–423. [Google Scholar] [CrossRef]
- Cruz-Jentoft, A.J.; Sayer, A.A. Sarcopenia. Lancet 2019, 393, 2636–2646. [Google Scholar] [CrossRef] [PubMed]
- Pinedo-Villanueva, R.; Westbury, L.D.; Syddall, H.E.; Sanchez-Santos, M.T.; Dennison, E.M.; Robinson, S.M.; Cooper, C. Health Care Costs Associated with Muscle Weakness: A UK Population-Based Estimate. Calcif. Tissue Int. 2019, 104, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.Y.; Chow, S.K.; Hung, V.W.; Wong, C.H.; Wong, R.M.; Tsang, C.S.; Kwok, T.; Cheung, W.H. Diagnosis of sarcopenia by evaluating skeletal muscle mass by adjusted bioimpedance analysis validated with dual-energy X-ray absorptiometry. J. Cachexia Sarcopenia Muscle 2021, 12, 2163–2173. [Google Scholar] [CrossRef]
- Traylor, D.A.; Gorissen, S.H.M.; Phillips, S.M. Perspective: Protein Requirements and Optimal Intakes in Aging: Are We Ready to Recommend More Than the Recommended Daily Allowance? Adv. Nutr. 2018, 9, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.M.Y.; Cheung, W.H.; Chow, S.K.H.; Ng, R.W.K.; Li, W.; Hsu, A.Y.; Wong, K.K.; Ho, A.W.; Choi, S.H.; Fang, C.X.; et al. Recommendations on the post-acute management of the osteoporotic fracture—Patients with “very-high” Re-fracture risk. J. Orthop. Translat. 2022, 37, 94–99. [Google Scholar] [CrossRef]
- Liu, C.; Wong, P.Y.; Tong, X.; Chow, S.K.; Hung, V.W.; Cheung, W.H.; Qin, L.; Law, S.W.; Wong, R.M.Y. Muscle plays a more superior role than fat in bone homeostasis: A cross-sectional study of old Asian people. Front. Endocrinol. 2022, 13, 990442. [Google Scholar] [CrossRef]
- Wong, R.M.Y.; Wong, H.; Zhang, N.; Chow, S.K.H.; Chau, W.W.; Wang, J.; Chim, Y.N.; Leung, K.S.; Cheung, W.H. The relationship between sarcopenia and fragility fracture-a systematic review. Osteoporos. Int. 2019, 30, 541–553. [Google Scholar] [CrossRef]
- Sluijs, I.; Beulens, J.W.; van der, A.D.; Spijkerman, A.M.; Grobbee, D.E.; van der Schouw, Y.T. Dietary intake of total, animal, and vegetable protein and risk of type 2 diabetes in the European Prospective Investigation into Cancer and Nutrition (EPIC)-NL study. Diabetes Care 2010, 33, 43–48. [Google Scholar] [CrossRef]
- Goates, S.; Du, K.; Arensberg, M.B.; Gaillard, T.; Guralnik, J.; Pereira, S.L. Economic Impact of Hospitalizations in US Adults with Sarcopenia. J. Frailty Aging 2019, 8, 93–99. [Google Scholar] [CrossRef]
- Robinson, S.M.; Reginster, J.Y.; Rizzoli, R.; Shaw, S.C.; Kanis, J.A.; Bautmans, I.; Bischoff-Ferrari, H.; Bruyere, O.; Cesari, M.; Dawson-Hughes, B.; et al. Does nutrition play a role in the prevention and management of sarcopenia? Clin. Nutr. 2018, 37, 1121–1132. [Google Scholar] [CrossRef] [PubMed]
- Frampton, J.; Murphy, K.G.; Frost, G.; Chambers, E.S. Short-chain fatty acids as potential regulators of skeletal muscle metabolism and function. Nat. Metab. 2020, 2, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, S.K. Sarcopenia: A Contemporary Health Problem among Older Adult Populations. Nutrients 2020, 12, 1293. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.X.; Du, N.; Hu, J.; Ning, F.; Mei, X.; Li, Q.; Peng, L. Intramuscular accumulation of pentadecanoic acid activates AKT1 to phosphorylate NCOR1 and triggers FOXM1-mediated apoptosis in the pathogenesis of sarcopenia. Am. J. Transl. Res. 2020, 12, 5064–5079. [Google Scholar]
- Candow, D.G.; Forbes, S.C.; Little, J.P.; Cornish, S.M.; Pinkoski, C.; Chilibeck, P.D. Effect of nutritional interventions and resistance exercise on aging muscle mass and strength. Biogerontology 2012, 13, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Bryner, R.W.; Woodworth-Hobbs, M.E.; Williamson, D.L.; Alway, S.E. Docosahexaenoic Acid protects muscle cells from palmitate-induced atrophy. ISRN Obes. 2012, 2012, 647348. [Google Scholar] [CrossRef]
- Saini, A.; Sharples, A.P.; Al-Shanti, N.; Stewart, C.E. Omega-3 fatty acid EPA improves regenerative capacity of mouse skeletal muscle cells exposed to saturated fat and inflammation. Biogerontology 2017, 18, 109–129. [Google Scholar] [CrossRef]
- Gonzalez-Hedstrom, D.; de La Fuente-Fernandez, M.; Priego, T.; Martin, A.I.; Amor, S.; Lopez-Calderon, A.; Inarejos-Garcia, A.M.; Garcia-Villalon, A.L.; Granado, M. Addition of Olive Leaf Extract to a Mixture of Algae and Extra Virgin Olive Oils Decreases Fatty Acid Oxidation and Synergically Attenuates Age-Induced Hypertension, Sarcopenia and Insulin Resistance in Rats. Antioxidants 2021, 10, 1066. [Google Scholar] [CrossRef]
- Lopez-Seoane, J.; Martinez-Ferran, M.; Romero-Morales, C.; Pareja-Galeano, H. N-3 PUFA as an ergogenic supplement modulating muscle hypertrophy and strength: A systematic review. Crit. Rev. Food Sci. Nutr. 2022, 62, 9000–9020. [Google Scholar] [CrossRef]
- Lippi, L.; Uberti, F.; Folli, A.; Turco, A.; Curci, C.; d’Abrosca, F.; de Sire, A.; Invernizzi, M. Impact of nutraceuticals and dietary supplements on mitochondria modifications in healthy aging: A systematic review of randomized controlled trials. Aging Clin. Exp. Res. 2022, 34, 2659–2674. [Google Scholar] [CrossRef]
- Li, C.W.; Yu, K.; Shyh-Chang, N.; Jiang, Z.; Liu, T.; Ma, S.; Luo, L.; Guang, L.; Liang, K.; Ma, W.; et al. Pathogenesis of sarcopenia and the relationship with fat mass: Descriptive review. J. Cachexia Sarcopenia Muscle 2022, 13, 781–794. [Google Scholar] [CrossRef] [PubMed]
- Kunz, H.E.; Michie, K.L.; Gries, K.J.; Zhang, X.; Ryan, Z.C.; Lanza, I.R. A Randomized Trial of the Effects of Dietary n3-PUFAs on Skeletal Muscle Function and Acute Exercise Response in Healthy Older Adults. Nutrients 2022, 14, 3537. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.I.; Mikhail, A.; Lan, L.; Di Carlo, A.; Hamilton, B.; Barnard, K.; Hettinga, B.P.; Hatcher, E.; Tarnopolsky, M.G.; Nederveen, J.P.; et al. A Five-Ingredient Nutritional Supplement and Home-Based Resistance Exercise Improve Lean Mass and Strength in Free-Living Elderly. Nutrients 2020, 12, 2391. [Google Scholar] [CrossRef] [PubMed]
- Strandberg, E.; Ponsot, E.; Piehl-Aulin, K.; Falk, G.; Kadi, F. Resistance Training Alone or Combined With N-3 PUFA-Rich Diet in Older Women: Effects on Muscle Fiber Hypertrophy. J. Gerontol. A Biol. Sci. Med. Sci. 2019, 74, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Rolland, Y.; Barreto, P.S.; Maltais, M.; Guyonnet, S.; Cantet, C.; Andrieu, S.; Vellas, B. Effect of Long-Term Omega 3 Polyunsaturated Fatty Acid Supplementation with or without Multidomain Lifestyle Intervention on Muscle Strength in Older Adults: Secondary Analysis of the Multidomain Alzheimer Preventive Trial (MAPT). Nutrients 2019, 11, 1931. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Sterne, J.A.C.; Savovic, J.; Page, M.J.; Elbers, R.G.; Blencowe, N.S.; Boutron, I.; Cates, C.J.; Cheng, H.Y.; Corbett, M.S.; Eldridge, S.M.; et al. RoB 2: A revised tool for assessing risk of bias in randomised trials. BMJ 2019, 366, l4898. [Google Scholar] [CrossRef]
- Jadad, A.R.; Moore, R.A.; Carroll, D.; Jenkinson, C.; Reynolds, D.J.; Gavaghan, D.J.; McQuay, H.J. Assessing the quality of reports of randomized clinical trials: Is blinding necessary? Control. Clin. Trials 1996, 17, 1–12. [Google Scholar] [CrossRef]
- Boutry-Regard, C.; Vinyes-Parés, G.; Breuillé, D.; Moritani, T. Supplementation with Whey Protein, Omega-3 Fatty Acids and Polyphenols Combined with Electrical Muscle Stimulation Increases Muscle Strength in Elderly Adults with Limited Mobility: A Randomized Controlled Trial. Nutrients 2020, 12, 1866. [Google Scholar] [CrossRef]
- Cornish, S.M.; Chilibeck, P.D. Alpha-linolenic acid supplementation and resistance training in older adults. Appl. Physiol. Nutr. Metab. 2009, 34, 49–59. [Google Scholar] [CrossRef]
- Cornish, S.M.; Myrie, S.B.; Bugera, E.M.; Chase, J.E.; Turczyn, D.; Pinder, M. Omega-3 supplementation with resistance training does not improve body composition or lower biomarkers of inflammation more so than resistance training alone in older men. Nutr. Res. 2018, 60, 87–95. [Google Scholar] [CrossRef]
- Da Boit, M.; Sibson, R.; Sivasubramaniam, S.; Meakin, J.R.; Greig, C.A.; Aspden, R.M.; Thies, F.; Jeromson, S.; Hamilton, D.L.; Speakman, J.R.; et al. Sex differences in the effect of fish-oil supplementation on the adaptive response to resistance exercise training in older people: A randomized controlled trial. Am. J. Clin. Nutr. 2017, 105, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Dalle, S.; Van Roie, E.; Hiroux, C.; Vanmunster, M.; Coudyzer, W.; Suhr, F.; Bogaerts, S.; Van Thienen, R.; Koppo, K. Omega-3 Supplementation Improves Isometric Strength but Not Muscle Anabolic and Catabolic Signaling in Response to Resistance Exercise in Healthy Older Adults. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 76, 406–414. [Google Scholar] [CrossRef]
- Krzymińska-Siemaszko, R.; Czepulis, N.; Lewandowicz, M.; Zasadzka, E.; Suwalska, A.; Witowski, J.; Wieczorowska-Tobis, K. The Effect of a 12-Week Omega-3 Supplementation on Body Composition, Muscle Strength and Physical Performance in Elderly Individuals with Decreased Muscle Mass. Int. J. Environ. Res. Public Health 2015, 12, 10558–10574. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.I.; Atherton, P.; Reeds, D.N.; Mohammed, B.S.; Rankin, D.; Rennie, M.J.; Mittendorfer, B. Dietary omega-3 fatty acid supplementation increases the rate of muscle protein synthesis in older adults: A randomized controlled trial. Am. J. Clin. Nutr. 2011, 93, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.I.; Julliand, S.; Reeds, D.N.; Sinacore, D.R.; Klein, S.; Mittendorfer, B. Fish oil-derived n-3 PUFA therapy increases muscle mass and function in healthy older adults. Am. J. Clin. Nutr. 2015, 102, 115–122. [Google Scholar] [CrossRef]
- Van Vliet, S.; Fappi, A.; Reeds, D.N.; Mittendorfer, B. No independent or combined effects of vitamin D and conjugated linoleic acids on muscle protein synthesis in older adults: A randomized, double-blind, placebo-controlled clinical trial. Am. J. Clin. Nutr. 2020, 112, 1382–1389. [Google Scholar] [CrossRef]
- Yoshino, J.; Smith, G.I.; Kelly, S.C.; Julliand, S.; Reeds, D.N.; Mittendorfer, B. Effect of dietary n-3 PUFA supplementation on the muscle transcriptome in older adults. Physiol. Rep. 2016, 4, e12785. [Google Scholar] [CrossRef]
- Cruz-Jentoft, A.J. Sarcopenia: Revised European consensus on definition and diagnosis. Age Ageing 2019, 48, 16–31. [Google Scholar] [CrossRef]
- Ghzaiel, I.; Zarrouk, A.; Nury, T.; Libergoli, M.; Florio, F.; Hammouda, S.; Ménétrier, F.; Avoscan, L.; Yammine, A.; Samadi, M.; et al. Antioxidant Properties and Cytoprotective Effect of Pistacia lentiscus L. Seed Oil against 7β-Hydroxycholesterol-Induced Toxicity in C2C12 Myoblasts: Reduction in Oxidative Stress, Mitochondrial and Peroxisomal Dysfunctions and Attenuation of Cell Death. Antioxidants 2021, 10, 1772. [Google Scholar] [CrossRef]
- Johnson, M.L.; Lalia, A.Z.; Dasari, S.; Pallauf, M.; Fitch, M.; Hellerstein, M.K.; Lanza, I.R. Eicosapentaenoic acid but not docosahexaenoic acid restores skeletal muscle mitochondrial oxidative capacity in old mice. Aging Cell 2015, 14, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Halade, G.V.; El Jamali, A.; Fernandes, G. Conjugated linoleic acid (CLA) prevents age-associated skeletal muscle loss. Biochem. Biophys. Res. Commun. 2009, 383, 513–518. [Google Scholar] [CrossRef]
- Liou, J.; Tuazon, M.A.; Burdzy, A.; Henderson, G.C. Moderate compared to low dietary intake of trans-fatty acids impairs strength of old and aerobic capacity of young SAMP8 mice in both sexes. Lipids 2013, 48, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, H.; Nishimura, M.; Uehara, M.; Kuribara-Souta, A.; Yamamoto, M.; Yoshikawa, N.; Morohashi, K.I.; Tanaka, H. Eicosapentaenoic acid changes muscle transcriptome and intervenes in aging-related fiber type transition in male mice. Am. J. Physiol. Endocrinol. Metab. 2021, 320, E346–E358. [Google Scholar] [CrossRef] [PubMed]
- Russ, D.W.; Dimova, K.; Morris, E.; Pacheco, M.; Garvey, S.M.; Scordilis, S.P. Dietary fish oil supplement induces age-specific contractile and proteomic responses in muscles of male rats. Lipids Health Dis. 2020, 19, 165. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Hedstrom, D.; Priego, T.; Lopez-Calderon, A.; Amor, S.; de la Fuente-Fernandez, M.; Inarejos-Garcia, A.M.; Garcia-Villalon, A.L.; Martin, A.I.; Granado, M. Beneficial Effects of a Mixture of Algae and Extra Virgin Olive Oils on the Age-Induced Alterations of Rodent Skeletal Muscle: Role of HDAC-4. Nutrients 2021, 13, 44. [Google Scholar] [CrossRef]
- Wong, R.M.Y.; Wong, P.Y.; Liu, C.; Wong, H.W.; Chung, Y.L.; Chow, S.K.H.; Law, S.W.; Cheung, W.H. The imminent risk of a fracture-existing worldwide data: A systematic review and meta-analysis. Osteoporos. Int. 2022, 33, 2453–2466. [Google Scholar] [CrossRef]
- Zhang, N.; Chow, S.K.H.; Leung, K.S.; Lee, H.H.; Cheung, W.H. An animal model of co-existing sarcopenia and osteoporotic fracture in senescence accelerated mouse prone 8 (SAMP8). Exp. Gerontol. 2017, 97, 1–8. [Google Scholar] [CrossRef]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef]
- Han, J.H.; Kim, I.S.; Jung, S.H.; Lee, S.G.; Son, H.Y.; Myung, C.S. The effects of propionate and valerate on insulin responsiveness for glucose uptake in 3T3-L1 adipocytes and C2C12 myotubes via G protein-coupled receptor 41. PLoS ONE 2014, 9, e95268. [Google Scholar] [CrossRef]
- Henagan, T.M.; Stefanska, B.; Fang, Z.; Navard, A.M.; Ye, J.; Lenard, N.R.; Devarshi, P.P. Sodium butyrate epigenetically modulates high-fat diet-induced skeletal muscle mitochondrial adaptation, obesity and insulin resistance through nucleosome positioning. Br. J. Pharmacol. 2015, 172, 2782–2798. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.E.; Bhattacharya, A.; Sataranatarajan, K.; Qaisar, R.; Sloane, L.; Rahman, M.M.; Kinter, M.; Van Remmen, H. The histone deacetylase inhibitor butyrate improves metabolism and reduces muscle atrophy during aging. Aging Cell 2015, 14, 957–970. [Google Scholar] [CrossRef] [PubMed]
- Maruta, H.; Yoshimura, Y.; Araki, A.; Kimoto, M.; Takahashi, Y.; Yamashita, H. Activation of AMP-Activated Protein Kinase and Stimulation of Energy Metabolism by Acetic Acid in L6 Myotube Cells. PLoS ONE 2016, 11, e0158055. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cheung, W.H.; Li, J.; Chow, S.K.; Yu, J.; Wong, S.H.; Ip, M.; Sung, J.J.Y.; Wong, R.M.Y. Understanding the gut microbiota and sarcopenia: A systematic review. J. Cachexia Sarcopenia Muscle 2021, 12, 1393–1407. [Google Scholar] [CrossRef]
- Magee, P.; Pearson, S.; Allen, J. The omega-3 fatty acid, eicosapentaenoic acid (EPA), prevents the damaging effects of tumour necrosis factor (TNF)-alpha during murine skeletal muscle cell differentiation. Lipids Health Dis. 2008, 7, 24. [Google Scholar] [CrossRef]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta 2015, 1851, 469–484. [Google Scholar] [CrossRef]
- Calder, P.C. n-3 PUFA and inflammation: From membrane to nucleus and from bench to bedside. Proc. Nutr. Soc. 2020, 79, 404–416. [Google Scholar] [CrossRef]
- Chiang, N.; Serhan, C.N. Specialized pro-resolving mediator network: An update on production and actions. Essays Biochem. 2020, 64, 443–462. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Number | Study | Clinical Focus | Experimental Design | Intervention | Duration | Variables Measured | Muscle Measurement | Physical Performance Measurement | Resistance Exercise Training | Blood Sampling/Muscle Biopsy | Results |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Roma et al. (2015) [34] | 53 subjects; 17 men and 34 women (mean age: 74.6 ± 8.0 years) | RCT | n-3 PUFA+ Vit E Vit E (Con) | 12 weeks | * MS * MM * MP | bioimpedance analysis | means of the timed up-and-go test (TUG), 4-meter walking test, and handgrip strength | NA | NA | * MS, * MM, * MP: PUFA = Con |
| 2 | Mariasole et al. (2017) [32] | 50 men and women [men: n = 27, age: 70.6 ± 4.5 y; women: n = 23, age: 70.7 ± 3.3 y, | RCT | n–3 PUFA placebo (Con), all with lower-limb resistance exercise training | 18 weeks | * MP * MS plasma triglyceride concentrations, glucose, insulin, or inflammatory markers | MRI am dmuscle anatomic cross-sectional area (ACSA) | short-performance physical battery (SPPB) knee-extensor muscles | both groups twice a week for 18 weeks | Insulin, * IL-6 and * TNF-a | Decrease in triglyceride In men: * MS, * MP: PUFA = Con In women: MS, MP: PUFA > Con |
| 3 | Emelie et al. (2019) [24] | 63 healthy recreationally active older women (65–70 years) | RCT | resistance training and healthy diet rich in n-3 PUFAs (RT-HD), resistance training only (RT) and controls (CON) | 24 weeks | inflammatory biomarkers MS | NA | NA | twice a a week for 24 weeks | * IL-1β and * mTOR | * MS: RT-HD > RT = Con RT-HD: Decrease in IL-1β and upregulate in mTOR |
| 4 | Stephen et al. (2018) [31] | elderly men (n = 23); age: 71.4 ± 6.2 | RCT | resistance training and omega-3 (Exp) and placebo (Con) | 12 weeks | * MM, * MP, blood samples: (* IL-6 and * TNF-α | dual-energy X-ray absorptiometry (DXA) | 6 min walk test, chest press, and leg press | 3 days per week for approximately 1 h in duration each day with approximately 48 h of rest between resistance training sessions for 12 weeks | * IL-6 ranged from 8.07% to 8.41% and for * TNF-α it ranged from 5.62% to 8.69%. | * MM, * MP: Exp = Con * Pre < * Post * IL-6 and * TNF-α: * Exp = * Con |
| 5 | Sebastiaan et al. (2021) [33] | elderly women (n = 23) | RCT | resistance exercise and omega-3 (Exp), and corn oil (Con) | 12 weeks | * MS * MM * MP inflammatory (p65NF-κB) and catabolic (* FOXO1 and * LC3b) markers | computed tomography scan | 5-repetition chair sit-to-stand test (5STS), 30-s chair sit-to-stand test (30STS), timed up-and-go test (TUG), maximal gait speed test (MGS), handgrip strength test (HGS), and leg press. | NA | insulin (* IL-6 blood glucose levels | * MP, * MM, * MS: Exp > Con Decrease in p65NF-κB, * FOXO1, * LC3b |
| 6 | Stephan et al. (2020) [37] | 13 men and 19 women (age: 60–85) | double-blind, RCT | (1) vitamin D-3 per day. (2) CLA per day. (3) both Vit D and CLA. (4) placebo: | 8 weeks | * MS basal myofibrillar protein synthesis | DXA | handgrip strength | NA | myofibrillar protein synthesis, plasma glucose, amino acid, and insulin concentrations | No differences between groups |
| 7 | Yves et al. (2019) [25] | Elder (n = 1680, men: 1091, women: 589) age:(75.34 ± 4.42 years) | Multi-center, RCT | (1) ω3-PUFA; (2) ω3-PUFA plus the multidomain intervention. (3) the multidomain intervention plus placebo; d) placebo. | 36 months | * MS * MP | NA | repeated chair stand test, handgrip strength, walking speed, and balance tests | NA | NA | No differences between groups |
| 8 | Jun et al. (2016) [38] | 20 healthy participants; 60–85-year-old | double-blind RCT | PUFA corn oil (Con) | 6 months | * MS * MM Gene expression | magnetic resonance imaging | handgrip strength 1-RM muscle strength, leg press, chest press, knee extension, and knee flexion | NA | Micro Array | * MS, * MM: PUFA > Con increased the expression of * UCP3 and * UQCRC1 |
| 9 | Claire. et al. (2020) [29] | 41 participants (33 females and 8 males, age: 60–90) | single-center, parallel, double-blind RCT | electrical muscle stimulation procedures + (1) placebo capsules (CHO), (2) whey protein isolate capsules (WPIs), (3) WPI + omega-4 + rutin, and curcumin (WPI + BIO) | 1.5 g/d, for 13 weeks | * MP * MM * MS | ultrasonography and MRI | isometric knee extensions, gait speed with three minutes of rest | NA | NA | * MS + *MM: Pre = Post * MP: WPI + BIO > WPI = CHO |
| 10 | Gordon. et al. (2015) [36] | 44 participants (29 females, 15 males; 60–85-year-old | double-blind RCT | Corn oil (CON) n–3 PUFA group | four 1 g pills/d, for 6 months | * MM * MP | DXA, MRI | handgrip strength, 1-repetition maximum (1-RM) muscle strength, and average isokinetic muscle power | NA | Red blood cell lipids | * MM: Pre < Post * MP: Pre < Post * Body state: Pre = Post |
| 11 | Gordon. et al. (2011) [35] | 16 older adults (10 men and 6 women, 65 y of age) | RCT | omega-3 fatty acids (Exp) and corn oil (Con) | 8 weeks | MM plasma muscle protein fractional synthesis rate (FSR) | DXA | NA | NA | muscle protein synthesis, *Akt, mTOR, and p70s6k, muscle phospholipid fatty acid, and concentrations of phenylalanine and leucine | *MM: Pre = Post FSR: Exp = Con Increased the anabolic response to amino acid Increase in p70s6k and * mTOR |
| 12 | Stephen. et al. (2009) [30] | 51 elders (28 males and 23 females; age: 65.4 ± 0.8 years) | double-blind, RCT | alpha-linolenic acid (ALA) and placebo (Con) | 12 weeks | MS MP cytokines: IL-6, TNF-α | DXA | 1-repetition maximum (1RM) chest and leg press strength, knee extensors, and flexors | 3 days per week, with at least 1 day of rest between training days, for 12 weeks. | * IL-6 and * TNF-a | * MS: Male in ALA: Pre < Post * MP: ALA = Con IL-6: Male: Pre > Post Female: Pre = Post * TNF-α: Male: Pre > Post Female: Pre < Post |
| 13 | Mats et al. (2020) [23] | sedentary men (n = 32) | double-blind, RCT | resistance exercise and omega-3 (Exp) and placebo (Con) | 12 weeks | * MM * MS * MP | DXA | 1-RM grip strength, gait speed, maximal handgrip, isometric knee extension, and SPPB | home-based resistance exercise | NA | * MM, * MS, * MP: Exp > Con; Pre < Post |
| No. | Study | Cell Line | Intervention | Muscle Samples | Method | Muscle Measurement | Physical Performance | Measurement of Cytokines | Key Findings |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Amarjit et al. (2017) [17] | C2C12 | EPA and a-3 polyunsaturated fatty acid | NA | Interactions of FFAs with TNF-a and IGF-I, qRT-PCR, flow cytometry, and creatine kinase assay | NA | NA | * IGF-II, * Id3, * IGFBP-5, RP-IIb (polr2b), and Myogenin | EPA had little impact on the cell death phenotype observed in lipotoxic conditions but did show benefits in restoring differentiation under lipo-toxic plus cytotoxic conditions. |
| 2 | Chen et al. (2020) [14] | muscle myoblast cell line HSMM-1 | different concentrations (0, 0.1, 0.3, and 0.6 mM) of PDA | 24 paired quadriceps muscle tissue | microarray, RT-qPCR, transfection, immunoprecipitation, chromatin immunoprecipitation (ChIP) assay, and luciferase assay | NA | NA | * FOXM1, PUMA, * BAX, * BAK1, * CASP3, * CASP7, * CASP9, * BCL2, FAS, * NCOR1, * AKT1, and MYC | PDA induced the expression of * FOXM1 and pro-apoptotic genes in vitro, involved in the pathogenesis of sarcopenia by activating apoptotic signaling. |
| 3 | Imen et al. (2021) [40] | C2C12 myoblasts | Pistacia lentiscus L. seed oil | NA | MTT Assay, fluorescein diacetate assay, plasma membrane permeability, oxidative stress, and mitochondrial function | NA | NA | NA | Prevention of cell death and organelle dysfunction, and attenuation of oxidative stress. |
| No. | Study | Animal | Intervention | Muscle Samples | Method | Muscle Measurement | Physical Performance | Measurement of Cytokines | Key Findings |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Matthew et al. (2015) [41] | 36 adult (6 months) and 36 old (24 months) C57BL6 mice | chow enriched with EPA or DHA (3.4% kcals) for 10 weeks | quadriceps muscle | mitochondrial energetics, protein fractional synthesis rates, RNA sequencing, and mass spectrometry-based proteomics | NA | NA | NA | EPA can improve mitochondrial function and protein quality but does not restore age-related reductions in mitochondrial protein abundance. |
| 2 | Rahman et al. (2009) [42] | 80 eleven months old female C57BL/6 (B6) mice | c9t11-CLA or t10c12-CLA for 6 months | whole hind-limb skeletal muscle | mitochondrial function and serum malondialdehyde | dual-energy X-ray absorptiometry (DXA) | NA | * TNF-α and * IL-6 | Higher muscle mass, enhanced mitochondrial ATP production, and elevated muscle antioxidant enzymes |
| 3 | Jesse et al. (2013) [43] | 51 SAMP8 (young, 25 weeks; old, 60 weeks) | trans-fatty acids started from 3 weeks of age | quadriceps muscle | collagen and intramuscular Triacylglycerol content | Echo, MRI system | grip strength and VO2peak | TNF-a | No significant diet effects or interactions. |
| 4 | Hiroki et al. (2021) [44] | 12 71-week-old C57BL/6J | EPA-enriched diet for 12 weeks | lateral gastrocnemius and plantaris muscle | measurement of triglyceride (TG) levels, RNA sequencing, and histological analysis | NA | grip strength, and treadmill exhaustion test | * MYH7, * MYH2, and * MYH4 | Increased grip strength, higher insulin sensitivity, and partially inhibited fast-to-slow fiber-type transition. |
| 5 | David et al. (2020) [45] | 14 adult (8 months) and 12 aged (22 months) male, Sprague Dawley rats | dietary fish oil for 8 weeks | not mentioned | contractile properties, proteomic analysis, and immunoblotting | NA | grip strength | 24 proteins | Increase muscle contractile force, no changes in muscle mass, and no significant associations between contractile parameters and protein abundances. |
| 6 | Daniel et al. (2021) [18] | Three months old (Young; n = 11) and twenty-four months old (Old; n = 17) male Wistar rats | olive leaf extracts and a mixture of algae oil for 21 days | gastrocnemius and soleus muscles | vascular reactivity, serum parameters, protein quantification, qRT-PCR of micro-RNAs, and immunohistochemistry | NA | NA | *Akt, p-Akt, * GSK3β, p-GSK3β | Improved the lipid profile, increased HOMA-IR, and decreased the serum levels of miRNAs 21 and 146a, preventing muscle loss. |
| 7 | Daniel et al. (2020) [46] | Young (3 months, n = 11) and old (24 months, n = 8) male Wistar rats | algae oil and extra virgin olive oil for 21 days | left gastrocnemius muscles | protein quantification and RT-qPCR | NA | NA | * IGF-I, * Akt, LC3b, * HDAC-4 | Decreased the inflammatory state, did not prevent aging- induced alterations, and decreased autophagy activity. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, T.; Liu, C.; Cui, C.; Zhang, N.; Cheung, W.H.; Wong, R.M.Y. Potential of Fatty Acids in Treating Sarcopenia: A Systematic Review. Nutrients 2023, 15, 3613. https://doi.org/10.3390/nu15163613
Huang T, Liu C, Cui C, Zhang N, Cheung WH, Wong RMY. Potential of Fatty Acids in Treating Sarcopenia: A Systematic Review. Nutrients. 2023; 15(16):3613. https://doi.org/10.3390/nu15163613
Chicago/Turabian StyleHuang, Tao, Chaoran Liu, Can Cui, Ning Zhang, Wing Hoi Cheung, and Ronald Man Yeung Wong. 2023. "Potential of Fatty Acids in Treating Sarcopenia: A Systematic Review" Nutrients 15, no. 16: 3613. https://doi.org/10.3390/nu15163613
APA StyleHuang, T., Liu, C., Cui, C., Zhang, N., Cheung, W. H., & Wong, R. M. Y. (2023). Potential of Fatty Acids in Treating Sarcopenia: A Systematic Review. Nutrients, 15(16), 3613. https://doi.org/10.3390/nu15163613