
Ileal Dysbiosis Is Associated with Increased Acoustic Startle in the 22q11.2 Microdeletion Mouse Model of Schizophrenia

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
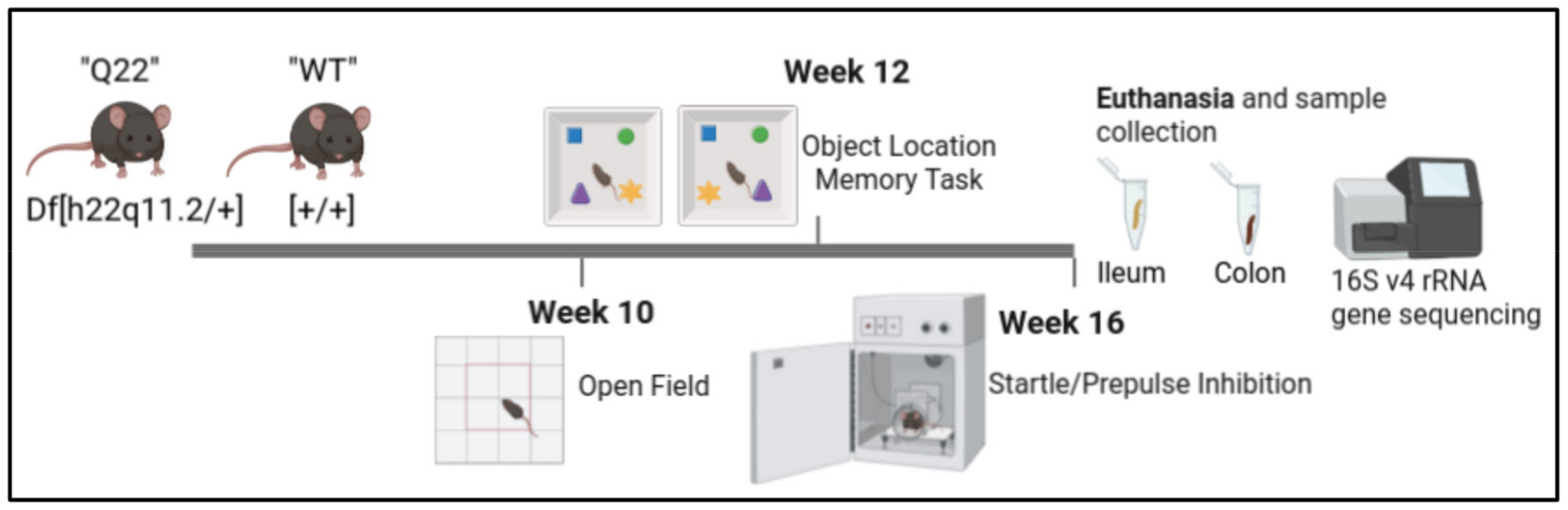
2. Materials and Methods
2.1. Animals
2.2. Handling
2.3. Acoustic Startle Response and Prepulse Inhibition of Startle
2.4. Open-Field Test
2.5. Object-in-Place Test
2.6. 16S V4 rRNA Gene Sequencing
2.7. Sequencing Data Preprocessing
2.8. Data Analysis
3. Results
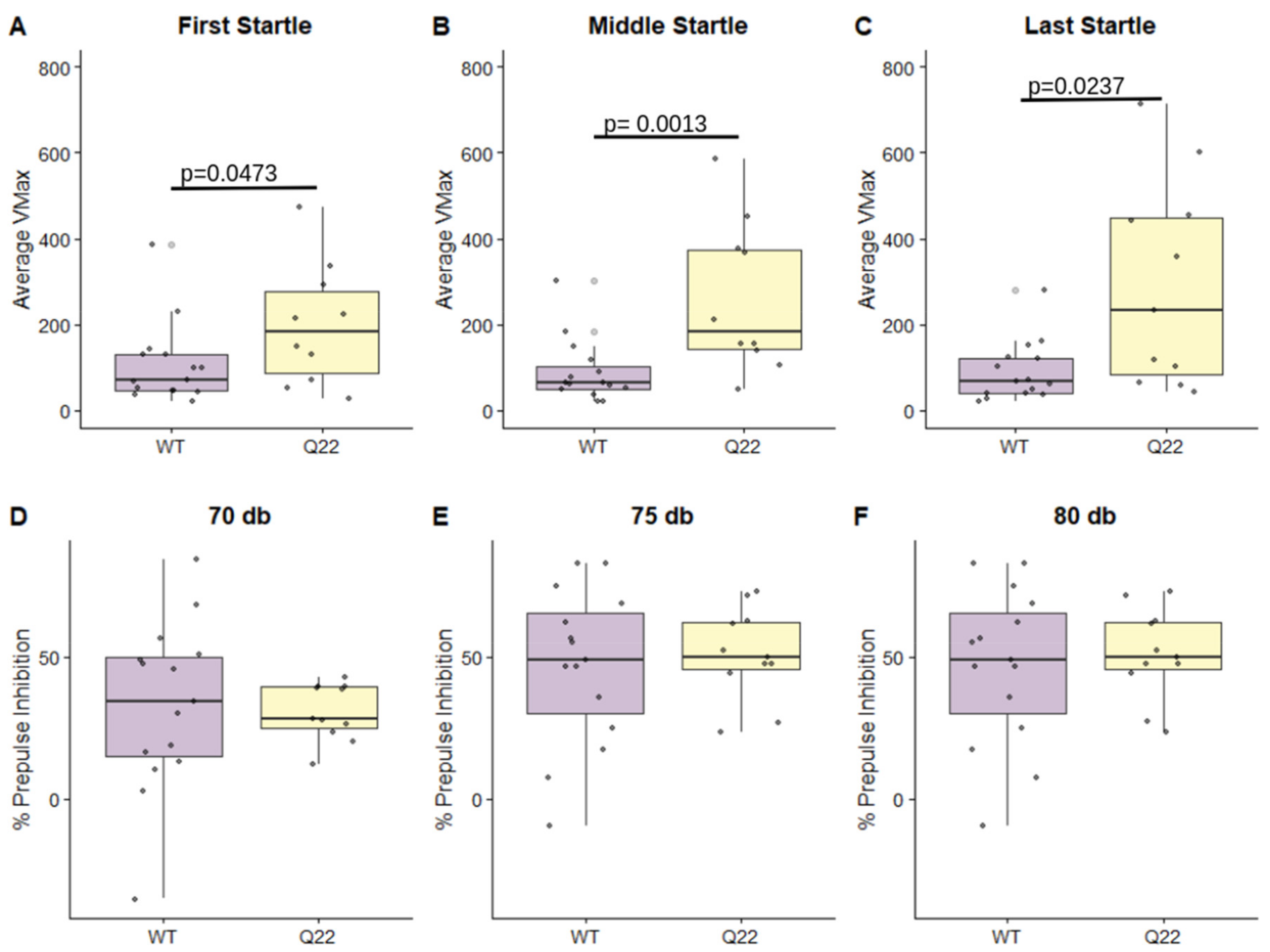
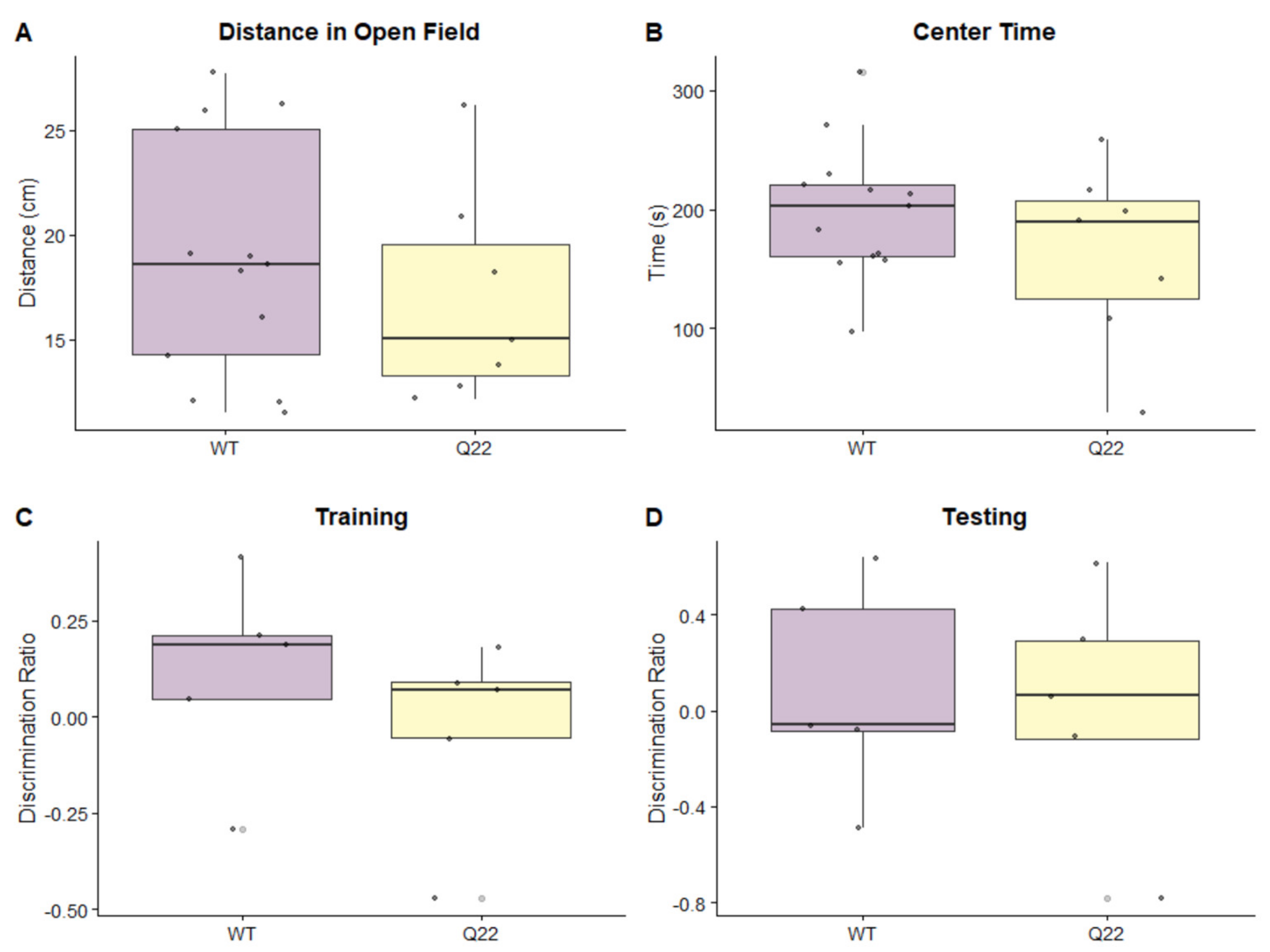
3.1. Q22 Mice Exhibit Sensorimotor Gating Deficits Compared to WT Counterparts, but Not Abnormalities in General Locomotor Activity, Anxiety-like Behavior, or Cognition
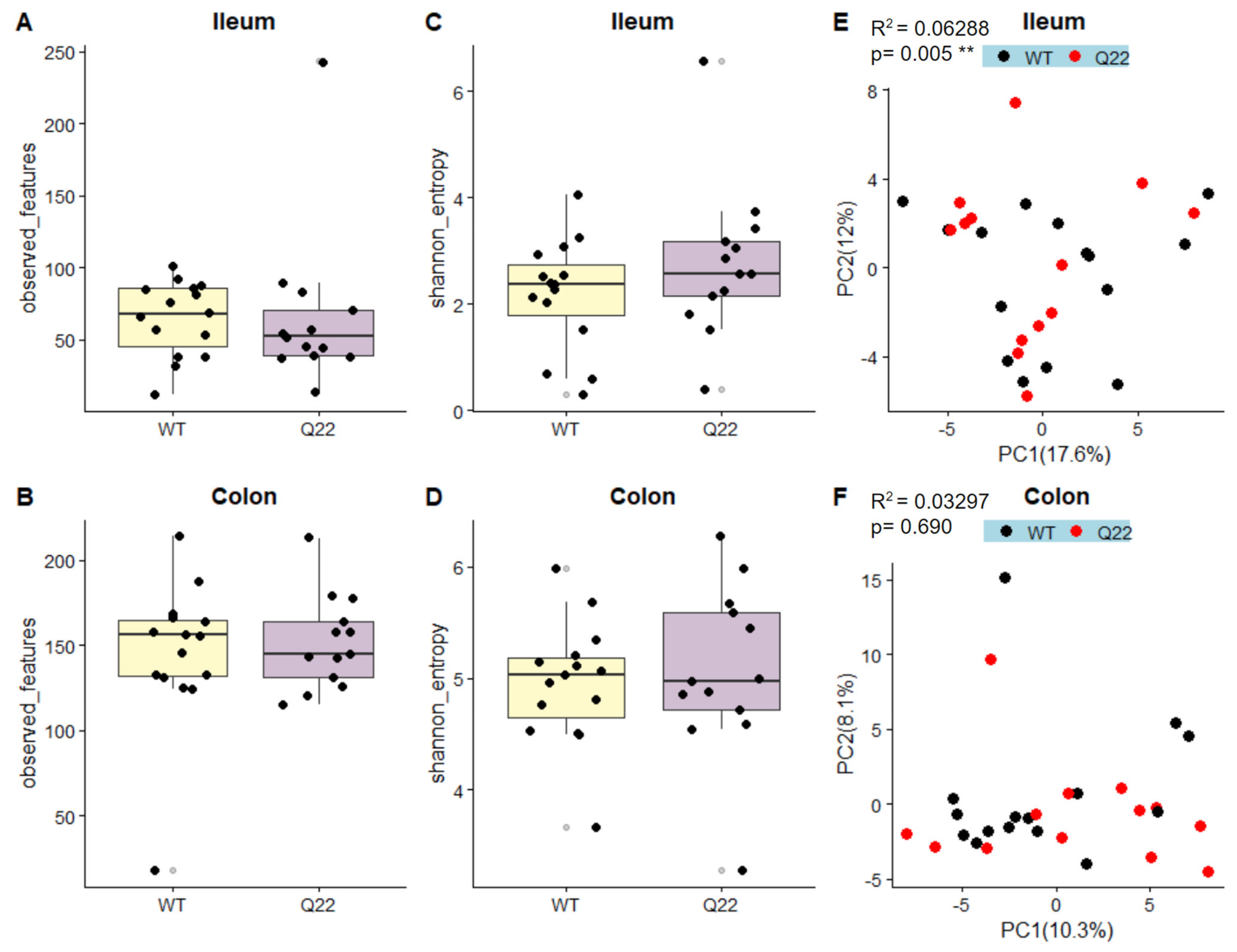
3.2. The Q22 Genotype Is Associated with Changes in the Ileal Microbiome
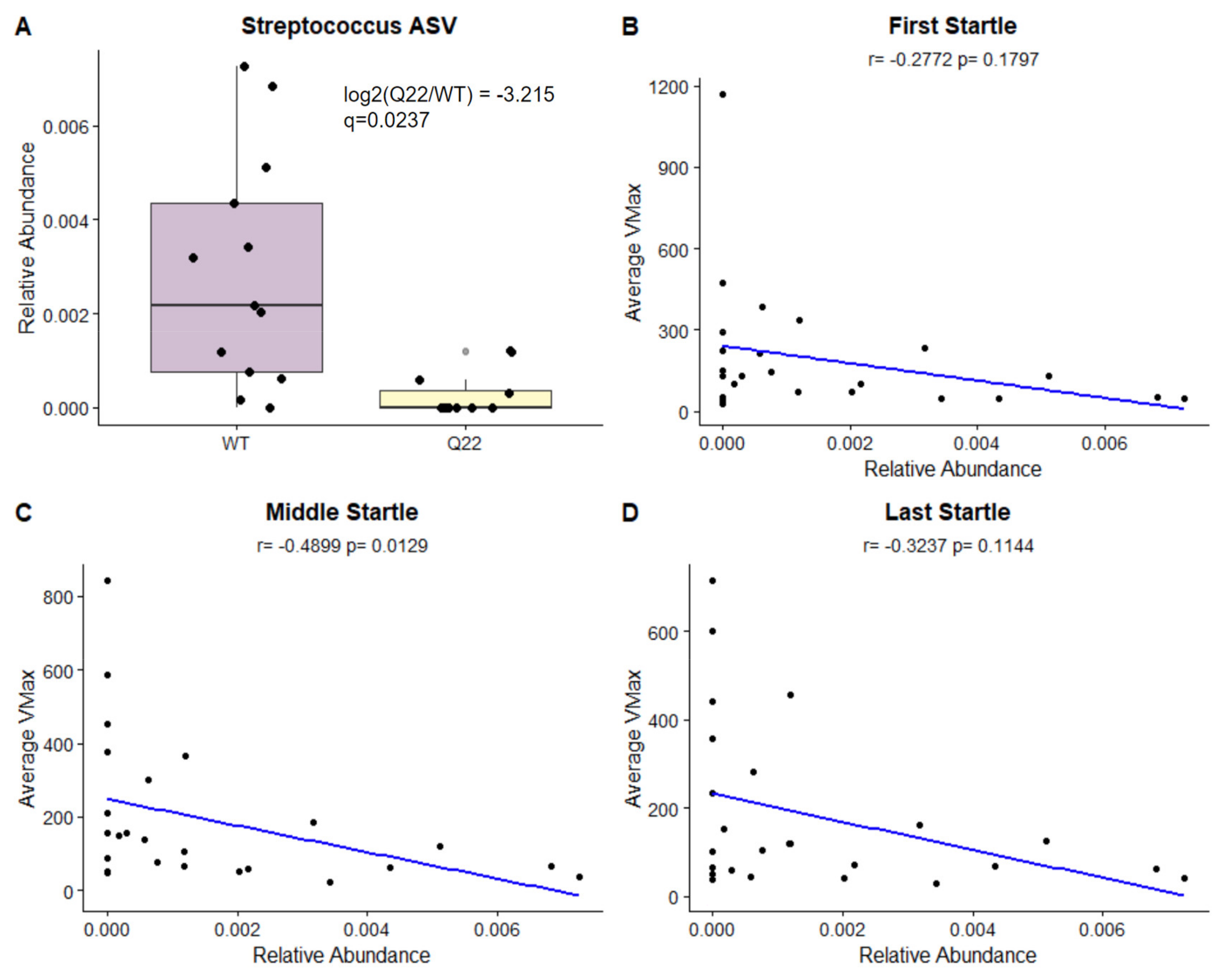
3.3. A Streptococcus ASV Distinguishes Q22 Mice from WT Mice and Is Significantly Correlated with Acoustic Startle Reflex
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kahn, R.S.; Sommer, I.E.; Murray, R.M.; Meyer-Lindenberg, A.; Weinberger, D.R.; Cannon, T.D.; O’Donovan, M.; Correll, C.U.; Kane, J.M.; van Os, J.; et al. Schizophrenia. Nat. Rev. Dis. Primer. 2015, 1, 15067. [Google Scholar] [CrossRef] [PubMed]
- Rahman, T.; Lauriello, J. Schizophrenia: An Overview. Focus. Am. Psychiatr. Publ. 2016, 14, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Brisch, R.; Saniotis, A.; Wolf, R.; Bielau, H.; Bernstein, H.-G.; Steiner, J.; Bogerts, B.; Braun, A.K.; Jankowski, Z.P.; Ekumaratilake, J.; et al. The role of dopamine in schizophrenia from a neurobiological and evolutionary perspective: Old fashioned, but still in vogue. Front. Psychiatry 2014, 5, 47. [Google Scholar] [CrossRef] [PubMed]
- Szeligowski, T.; Yun, A.L.; Lennox, B.R.; Burnet, P.W.J. The Gut Microbiome and Schizophrenia: The Current State of the Field and Clinical Applications. Front. Psychiatry 2020, 11, 156. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Kosciolek, T.; Daly, R.E.; Vázquez-Baeza, Y.; Swafford, A.; Knight, R.; Jeste, D.V. Gut microbiome in Schizophrenia: Altered functional pathways related to immune modulation and atherosclerotic risk. Brain Behav. Immun. 2021, 91, 245–256. [Google Scholar] [CrossRef]
- Li, S.; Song, J.; Ke, P.; Kong, L.; Lei, B.; Zhou, J.; Huang, Y.; Li, H.; Li, G.; Chen, J.; et al. The gut microbiome is associated with brain structure and function in schizophrenia. Sci. Rep. 2021, 11, 9743. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Zeng, B.; Liu, M.; Chen, J.; Pan, J.; Han, Y.; Liu, Y.; Cheng, K.; Zhou, C.; Wang, H.; et al. The gut microbiome from patients with schizophrenia modulates the glutamate-glutamine-GABA cycle and schizophrenia-relevant behaviors in mice. Sci. Adv. 2019, 5, eaau8317. [Google Scholar] [CrossRef]
- Zhu, F.; Guo, R.; Wang, W.; Ju, Y.; Wang, Q.; Ma, Q.; Sun, Q.; Fan, Y.; Xie, Y.; Yang, Z.; et al. Transplantation of microbiota from drug-free patients with schizophrenia causes schizophrenia-like abnormal behaviors and dysregulated kynurenine metabolism in mice. Mol. Psychiatry 2020, 25, 2905–2918. [Google Scholar] [CrossRef]
- Suganya, K.; Koo, B.S. Gut-Brain Axis: Role of Gut Microbiota on Neurological Disorders and How Probiotics/Prebiotics Beneficially Modulate Microbial and Immune Pathways to Improve Brain Functions. Int. J. Mol. Sci. 2020, 21, 7551. [Google Scholar] [CrossRef]
- Ma, Q.; Xing, C.; Long, W.; Wang, H.Y.; Liu, Q.; Wang, R.F. Impact of microbiota on central nervous system and neurological diseases: The gut-brain axis. J. Neuroinflamm. 2019, 16, 53. [Google Scholar] [CrossRef]
- Mirzaei, R.; Bouzari, B.; Hosseini-Fard, S.R.; Mazaheri, M.; Ahmadyousefi, Y.; Abdi, M.; Jalalifar, S.; Karimitabar, Z.; Teimoori, A.; Keyvani, H.; et al. Role of microbiota-derived short-chain fatty acids in nervous system disorders. Biomed. Pharmacother. 2021, 139, 111661. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.E.; Walker, A.K.; Weickert, C.S. Neuroinflammation in schizophrenia: The role of nuclear factor kappa B. Transl. Psychiatry 2021, 11, 528. [Google Scholar] [CrossRef]
- Ching, C.R.; Gutman, B.A.; Sun, D.; Reina, J.V.; Ragothaman, A.; Isaev, D.; Zavaliangos-Petropulu, A.; Lin, A.; Jonas, R.K.; Kushan, L.; et al. Psychiatric Disorders from Childhood to Adulthood in 22q11.2 Deletion Syndrome: Results From the International Consortium on Brain and Behavior in 22q11.2 Deletion Syndrome. Am. J. Psychiatry 2014, 171, 627–639. [Google Scholar] [CrossRef]
- Francisco, A.A.; Horsthuis, D.J.; Popiel, M.; Foxe, J.J.; Molholm, S. Atypical response inhibition and error processing in 22q11.2 Deletion Syndrome and schizophrenia: Towards neuromarkers of disease progression and risk. NeuroImage Clin. 2020, 27, 102351. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Spedding, M.; Schenker, E.; Didriksen, M.; Cressant, A.; Jay, T.M. Cognition- and circuit-based dysfunction in a mouse model of 22q11.2 microdeletion syndrome: Effects of stress. Transl. Psychiatry 2020, 10, 41. [Google Scholar] [CrossRef] [PubMed]
- Saito, R.; Koebis, M.; Nagai, T.; Shimizu, K.; Liao, J.; Wulaer, B.; Sugaya, Y.; Nagahama, K.; Uesaka, N.; Kushima, I.; et al. Comprehensive analysis of a novel mouse model of the 22q11.2 deletion syndrome: A model with the most common 3.0-Mb deletion at the human 22q11.2 locus. Transl. Psychiatry 2020, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Didriksen, M.; Fejgin, K.; Nilsson, S.R.; Birknow, M.R.; Grayton, H.M.; Larsen, P.H.; Lauridsen, J.B.; Nielsen, V.; Celada, P.; Santana, N.; et al. Persistent gating deficit and increased sensitivity to NMDA receptor antagonism after puberty in a new mouse model of the human 22q11.2 microdeletion syndrome: A study in male mice. J. Psychiatry Neurosci. 2017, 42, 48–58. [Google Scholar] [CrossRef]
- Tong, M.; Jacobs, J.P.; McHardy, I.H.; Braun, J. Sampling of intestinal microbiota and targeted amplification of bacterial 16S rRNA genes for microbial ecologic analysis. Curr. Protoc. Immunol. 2014, 107, 7.41.1–7.41.11. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glöckner, F.O. The SILVA and “All-species Living Tree Project (LTP)” taxonomic frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Powell, S.B.; Zhou, X.; Geyer, M.A. Prepulse inhibition and genetic mouse models of schizophrenia. Behav. Brain Res. 2009, 204, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.A.; Hills, P.J.; Dick, K.M.; Jones, S.P.; Bright, P. Cognitive mechanisms associated with auditory sensory gating. Brain Cogn. 2016, 102, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Braff, D.; Stone, C.; Callaway, E.; Geyer, M.; Glick, I.; Bali, L. Prestimulus effects on human startle reflex in normals and schizophrenics. Psychophysiology 1978, 15, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Curzon, P.; Zhang, M.; Radek, R.J.; Fox, G.B. The Behavioral Assessment of Sensorimotor Processes in the Mouse: Acoustic Startle, Sensory Gating, Locomotor Activity, Rotarod, and Beam Walking. In Methods of Behavior Analysis in Neuroscience, 2nd ed.; Buccafusco, J.J., Ed.; CRC Press: Boca Raton, FL, USA; Taylor & Francis: Abingdon, UK, 2009. [Google Scholar]
- Yang, Z.; Xiao, X.; Chen, R.; Xu, X.; Kong, W.; Zhang, T. Disc1 gene down-regulation impaired synaptic plasticity and recognition memory via disrupting neural activity in mice. Brain Res. Bull. 2021, 171, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Sintes, R.; Kvajo, M.; Gogos, J.A.; Lucas, J.J. Mice with a naturally occurring DISC1 mutation display a broad spectrum of behaviors associated to psychiatric disorders. Front. Behav. Neurosci. 2014, 8, 253. [Google Scholar] [CrossRef] [PubMed]
- Sultana, R.; Lee, C.C. Expression of Behavioral Phenotypes in Genetic and Environmental Mouse Models of Schizophrenia. Front. Behav. Neurosci. 2020, 14, 29. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Open Field Test for Measuring Locomotor Activity and Anxiety-Like Behavior. In Pre-Clinical Models: Techniques and Protocols; Guest, P.C., Ed.; Springer: New York, NY, USA, 2019; pp. 99–103. [Google Scholar] [CrossRef]
- Zhu, F.; Ju, Y.; Wang, W.; Wang, Q.; Guo, R.; Ma, Q.; Sun, Q.; Fan, Y.; Xie, Y.; Yang, Z.; et al. Metagenome-wide association of gut microbiome features for schizophrenia. Nat. Commun. 2020, 11, 1612. [Google Scholar] [CrossRef]
- Davies, E.G. Immunodeficiency in DiGeorge Syndrome and Options for Treating Cases with Complete Athymia. Front. Immunol. 2013, 4, 322. [Google Scholar] [CrossRef]
- Feifel, D.; Shilling, P.D. Promise and pitfalls of animal models of schizophrenia. Curr. Psychiatry Rep. 2010, 12, 327–334. [Google Scholar] [CrossRef]
- Gubert, C.; Kong, G.; Uzungil, V.; Zeleznikow-Johnston, A.M.; Burrows, E.L.; Renoir, T.; Hannan, A.J. Microbiome Profiling Reveals Gut Dysbiosis in the Metabotropic Glutamate Receptor 5 Knockout Mouse Model of Schizophrenia. Front. Cell Dev. Biol. 2020, 8, 582320. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Xu, J.; Li, Z.; Huang, Y.; Yuan, Y.; Wang, J.; Zhang, M.; Hu, S.; Liang, Y. Analysis of gut microbiota diversity and auxiliary diagnosis as a biomarker in patients with schizophrenia: A cross-sectional study. Schizophr. Res. 2018, 197, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Asif, H.; Dai, L.; He, Y.; Zheng, W.; Wang, D.; Ren, H.; Tang, J.; Li, C.; Jin, K.; et al. Alteration of the gut microbiome in first-episode drug-naïve and chronic medicated schizophrenia correlate with regional brain volumes. J. Psychiatr. Res. 2020, 123, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yuan, X.; Zhu, Z.; Pang, L.; Ding, S.; Li, X.; Kang, Y.; Hei, G.; Zhang, L.; Zhang, X.; et al. Multiomics Analyses Reveal Microbiome–Gut–Brain Crosstalk Centered on Aberrant Gamma-Aminobutyric Acid and Tryptophan Metabolism in Drug-Naïve Patients with First-Episode Schizophrenia. Schizophr. Bull. 2023, sbad026. [Google Scholar] [CrossRef] [PubMed]
- Forster, S.C.; Clare, S.; Beresford-Jones, B.S.; Harcourt, K.; Notley, G.; Stares, M.D.; Kumar, N.; Soderholm, A.T.; Adoum, A.; Wong, H.; et al. Identification of gut microbial species linked with disease variability in a widely used mouse model of colitis. Nat. Microbiol. 2022, 7, 590–599. [Google Scholar] [CrossRef] [PubMed]
- Ericsson, A.C.; Franklin, C.L. The gut microbiome of laboratory mice: Considerations and best practices for translational research. Mamm. Genome. 2021, 32, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Kotcher, R.E.; Chait, D.B.M.; Heckert, J.M.; Crowley, T.B.; Forde, K.A.M.; Ahuja, N.K.; Mascarenhas, M.R.M.; Emanuel, B.S.; Zackai, E.H.; McDonald-McGinn, D.M.; et al. Gastrointestinal Features of 22q11.2 Deletion Syndrome Include Chronic Motility Problems from Childhood to Adulthood. J. Pediatr. Gastroenterol. Nutr. 2022, 75, e8–e14. [Google Scholar] [CrossRef] [PubMed]
- Scarborough, J.; Mueller, F.; Weber-Stadlbauer, U.; Richetto, J.; Meyer, U. Dependency of prepulse inhibition deficits on baseline startle reactivity in a mouse model of the human 22q11.2 microdeletion syndrome. Genes. Brain Behav. 2019, 18, e12523. [Google Scholar] [CrossRef]
- Mowat, A.M.; Agace, W.W. Regional specialization within the intestinal immune system. Nat. Rev. Immunol. 2014, 14, 667–685. [Google Scholar] [CrossRef]
- Jacobson, A.; Yang, D.; Vella, M.; Chiu, I.M. The intestinal neuro-immune axis: Crosstalk between neurons, immune cells, and microbes. Mucosal Immunol. 2021, 14, 555–565. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.C.; Troutman, R.; Buri, H.; Gutta, A.; Situ, J.; Aja, E.; Jacobs, J.P. Ileal Dysbiosis Is Associated with Increased Acoustic Startle in the 22q11.2 Microdeletion Mouse Model of Schizophrenia. Nutrients 2023, 15, 3631. https://doi.org/10.3390/nu15163631
Yang JC, Troutman R, Buri H, Gutta A, Situ J, Aja E, Jacobs JP. Ileal Dysbiosis Is Associated with Increased Acoustic Startle in the 22q11.2 Microdeletion Mouse Model of Schizophrenia. Nutrients. 2023; 15(16):3631. https://doi.org/10.3390/nu15163631
Chicago/Turabian StyleYang, Julianne Ching, Ryan Troutman, Heidi Buri, Arjun Gutta, Jamilla Situ, Ezinne Aja, and Jonathan Patrick Jacobs. 2023. "Ileal Dysbiosis Is Associated with Increased Acoustic Startle in the 22q11.2 Microdeletion Mouse Model of Schizophrenia" Nutrients 15, no. 16: 3631. https://doi.org/10.3390/nu15163631
APA StyleYang, J. C., Troutman, R., Buri, H., Gutta, A., Situ, J., Aja, E., & Jacobs, J. P. (2023). Ileal Dysbiosis Is Associated with Increased Acoustic Startle in the 22q11.2 Microdeletion Mouse Model of Schizophrenia. Nutrients, 15(16), 3631. https://doi.org/10.3390/nu15163631